
Characterization of the Bacterial and Sulphate Reducing Community in the Alkaline and Constantly Cold Water of the Closed Kotalahti Mine



Abstract
:1. Introduction
2. Experimental Section
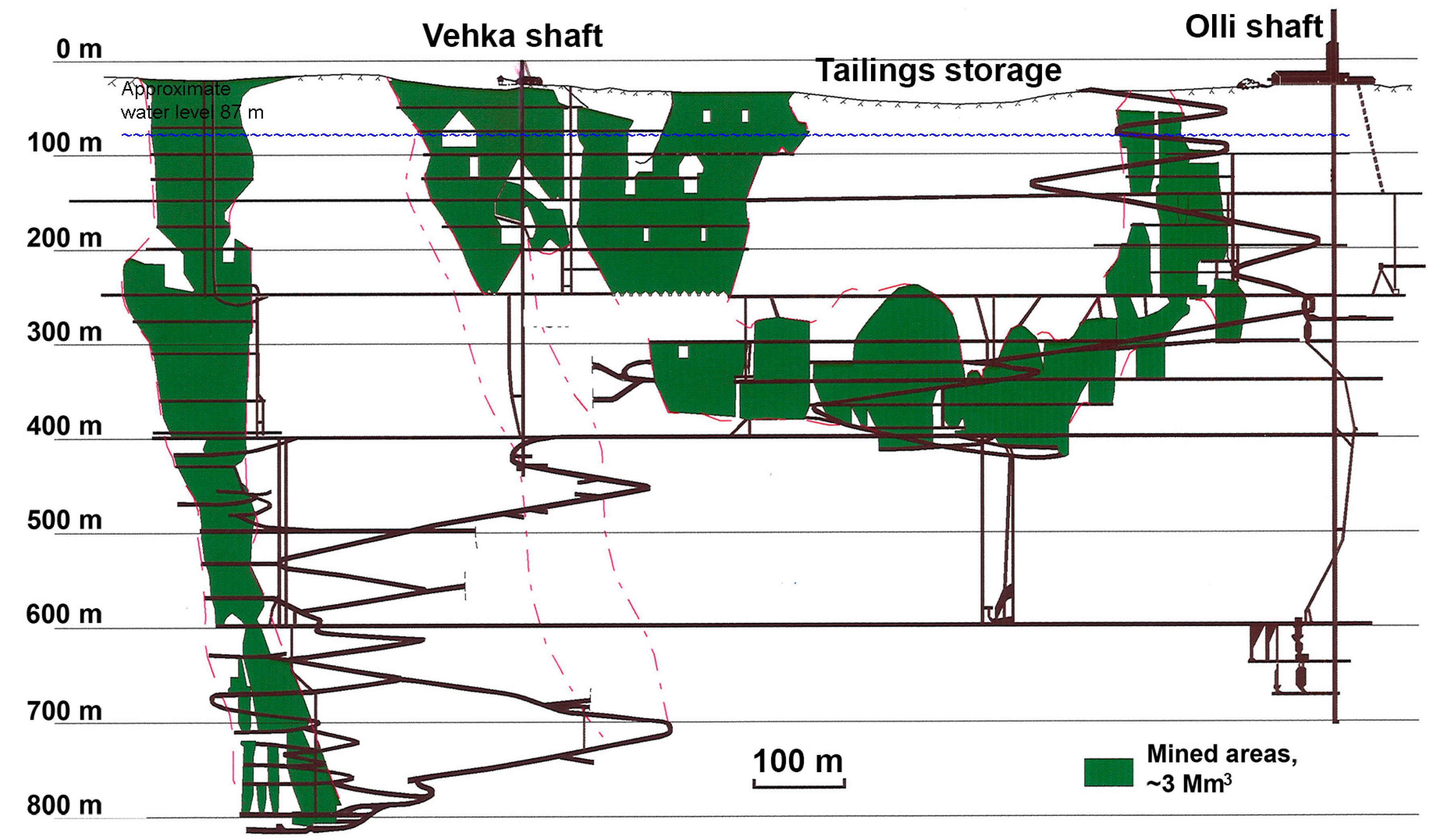
2.1. Sampling Site
2.2. Sampling
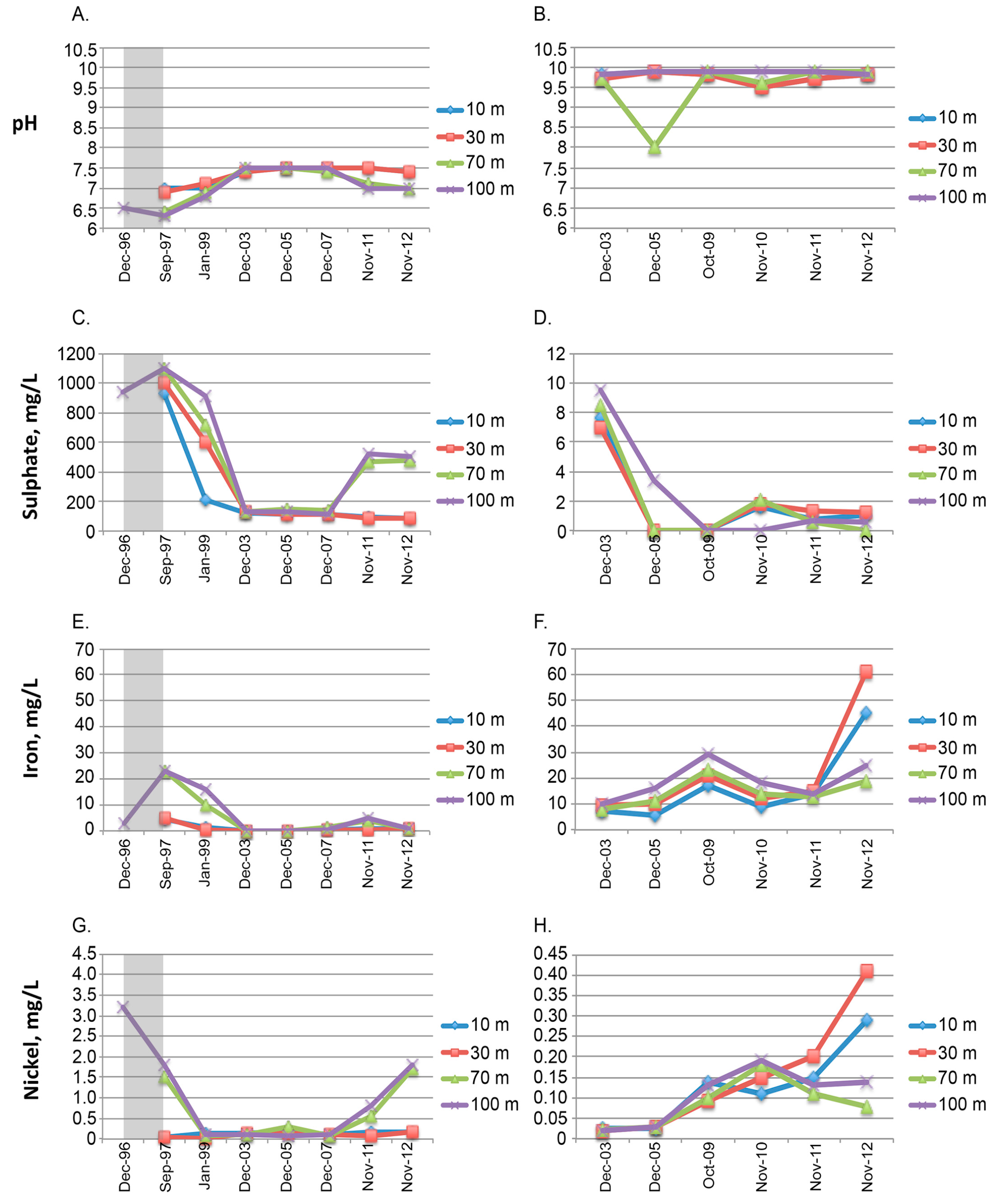
2.3. Physicochemical Measurements


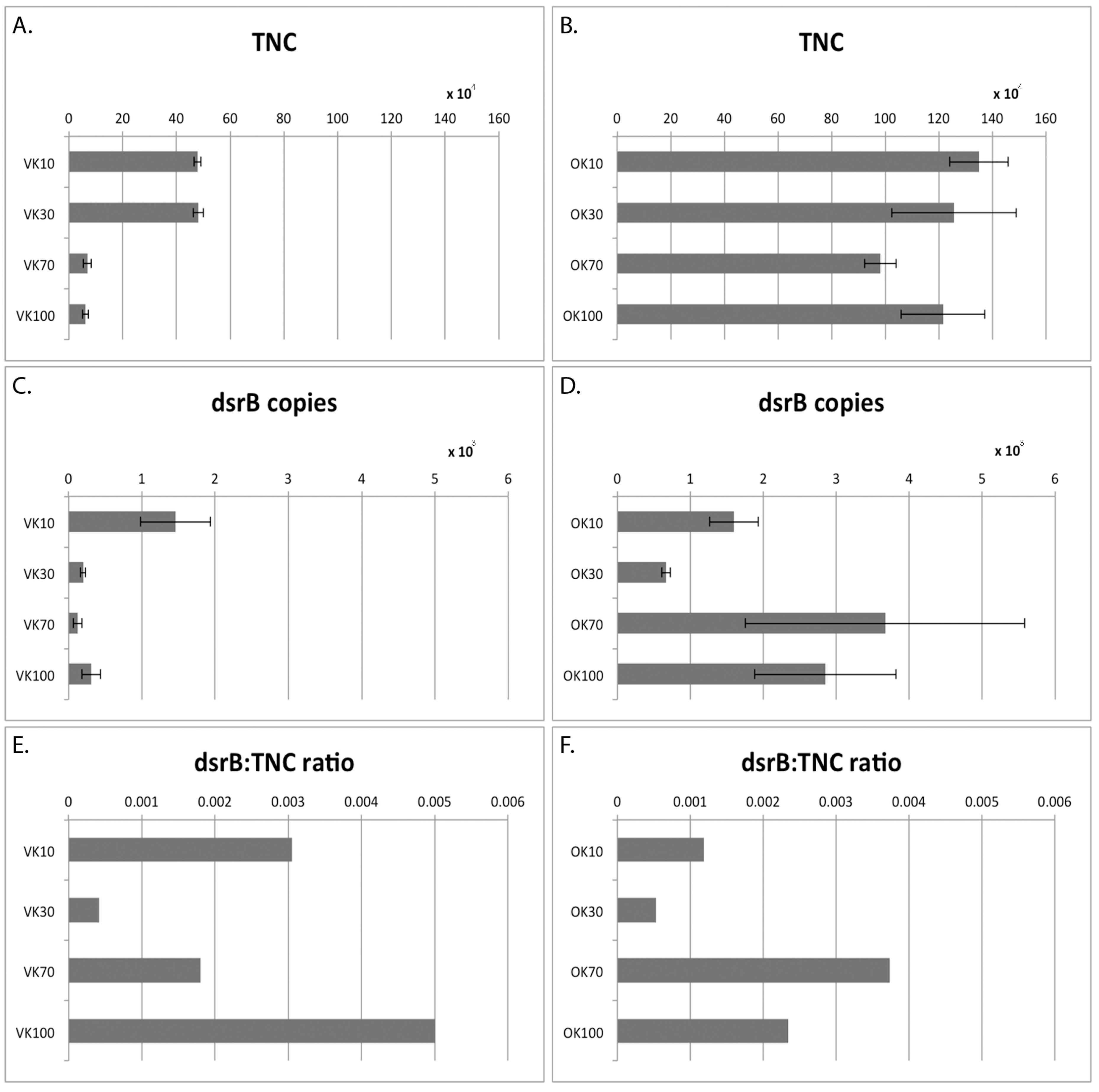
2.4. Total Number of Microbial Cells
2.5. DNA Extraction
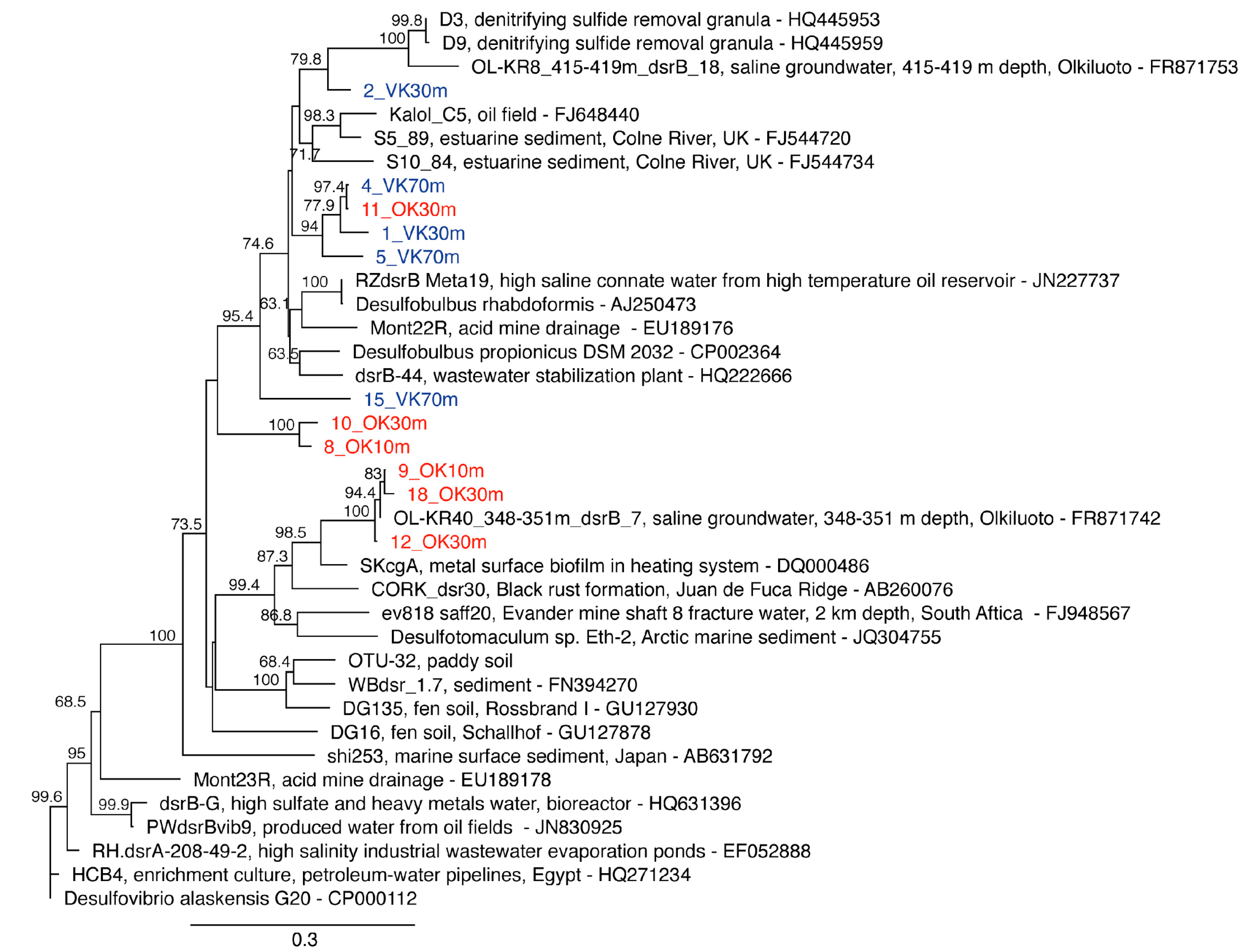
2.6. dsrB qPCR
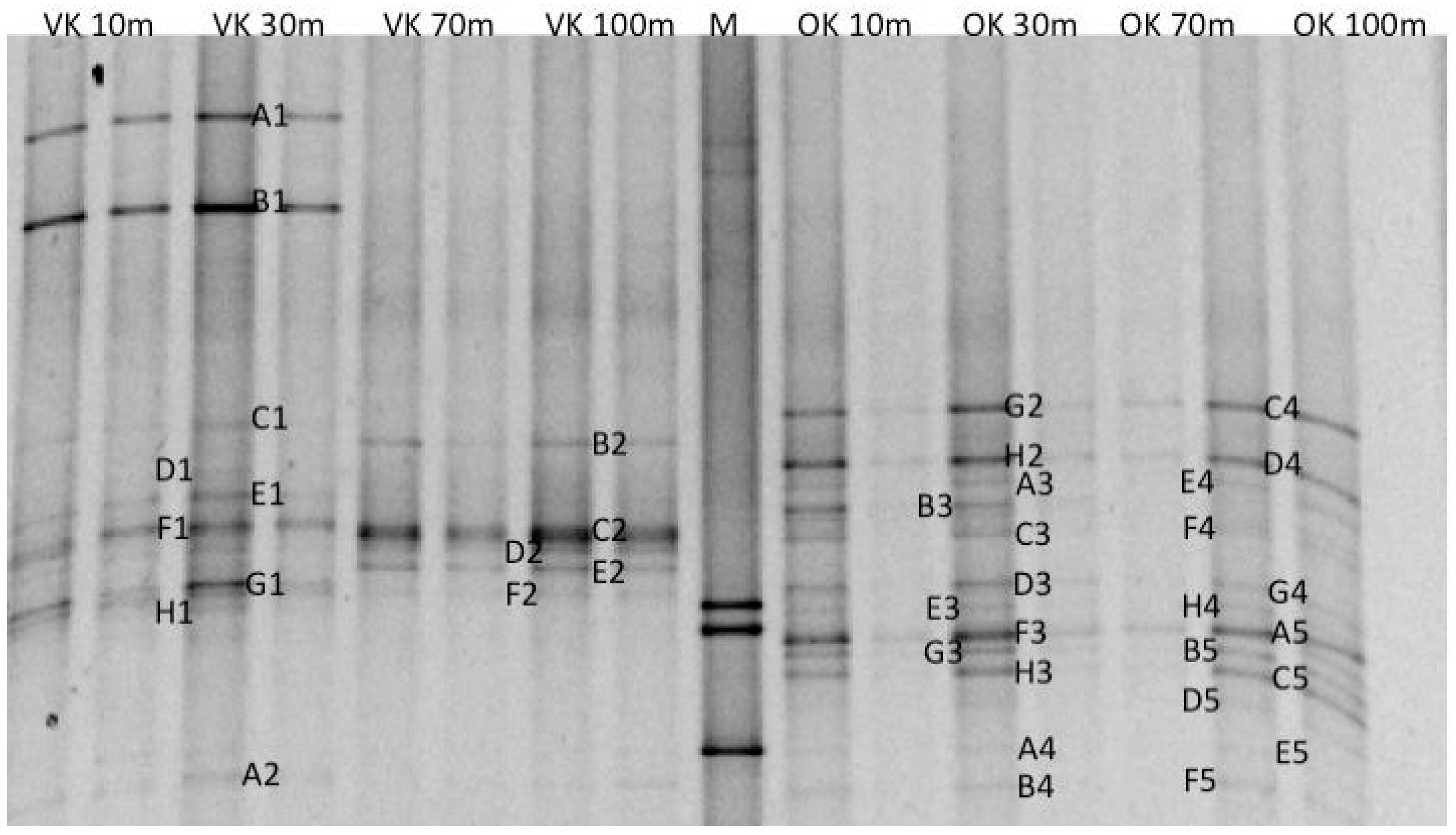
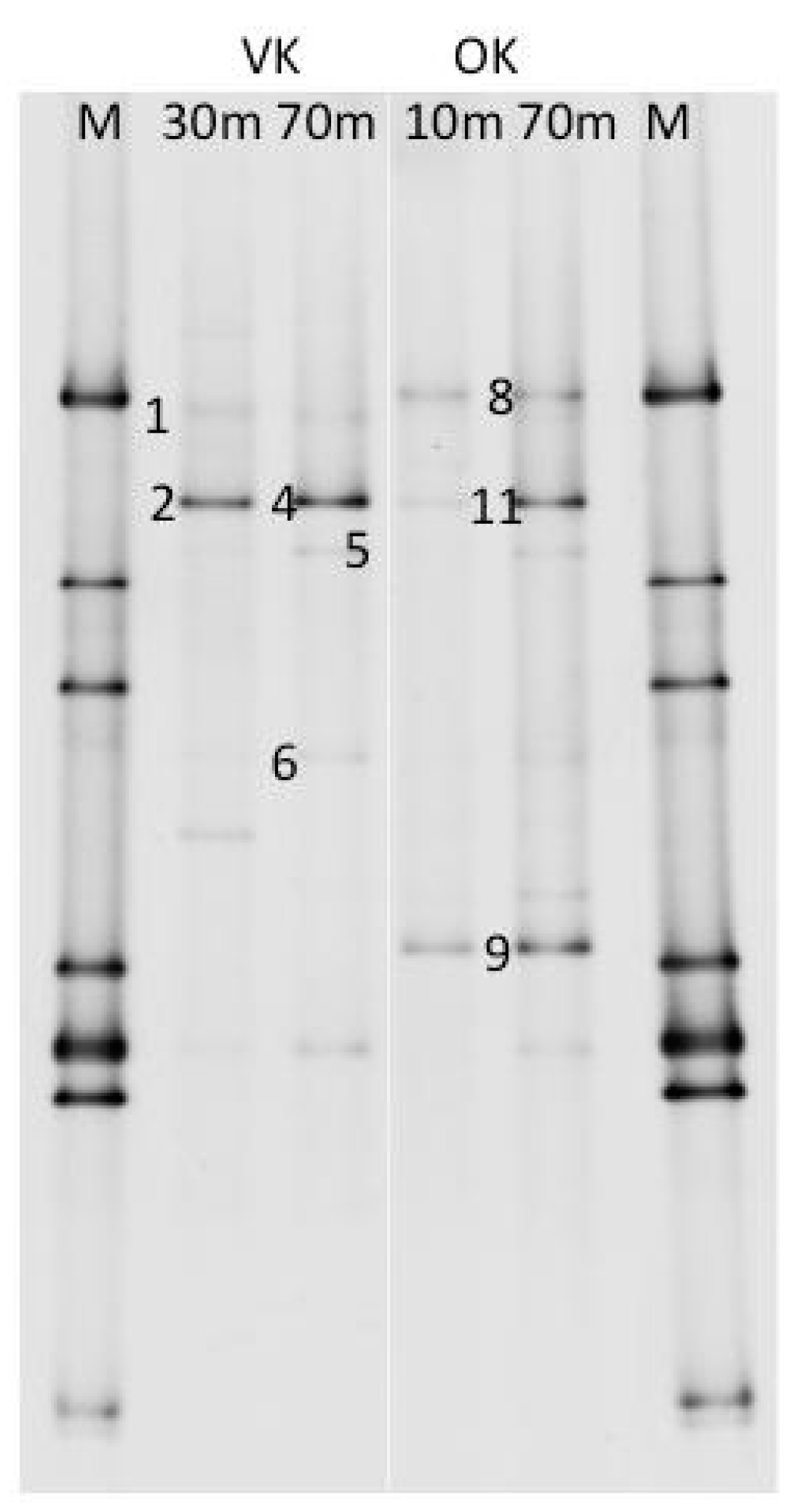
2.7. DGGE Analyses
2.8. Sequencing
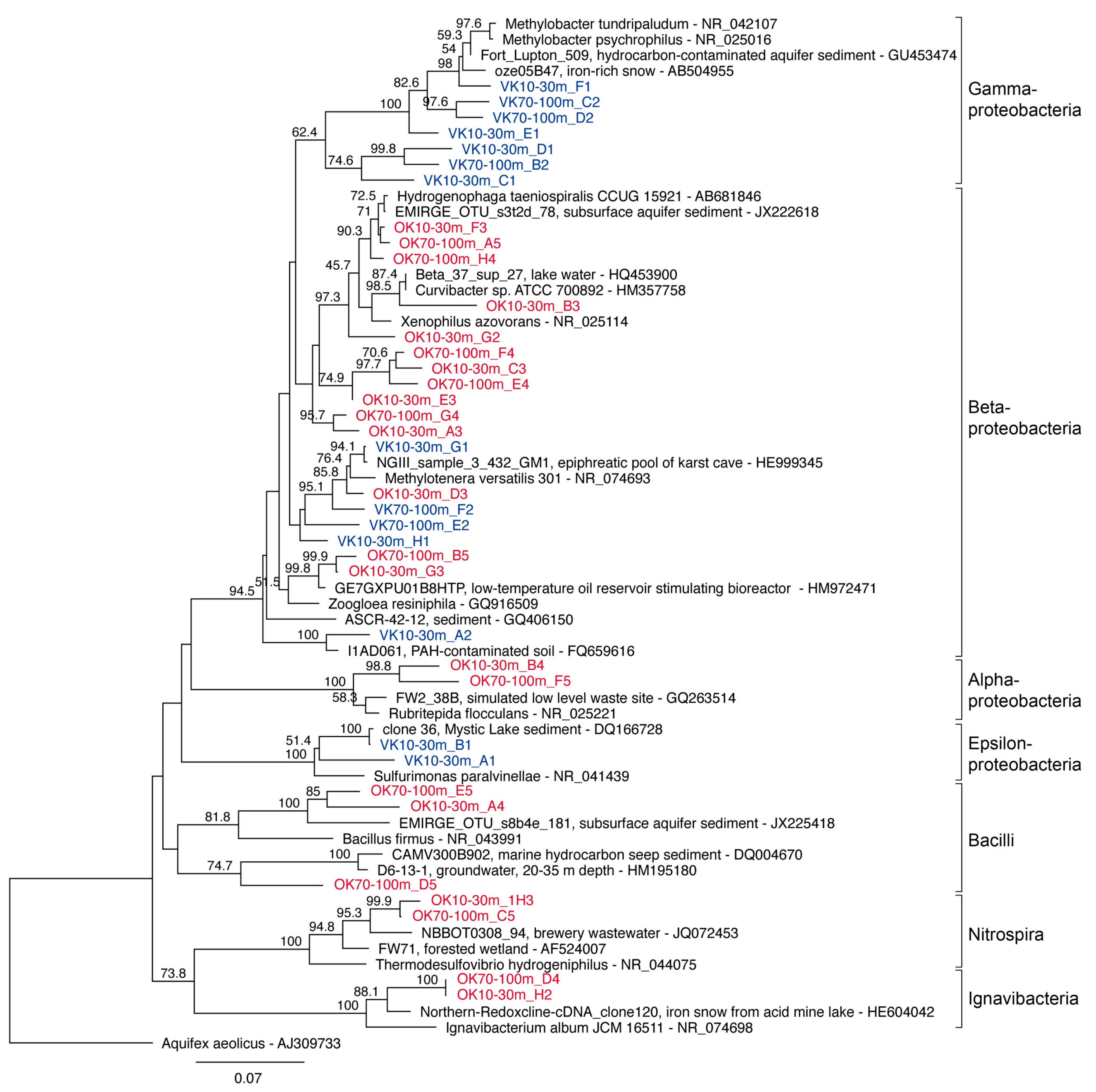
2.9. Phylogenetic Analyses
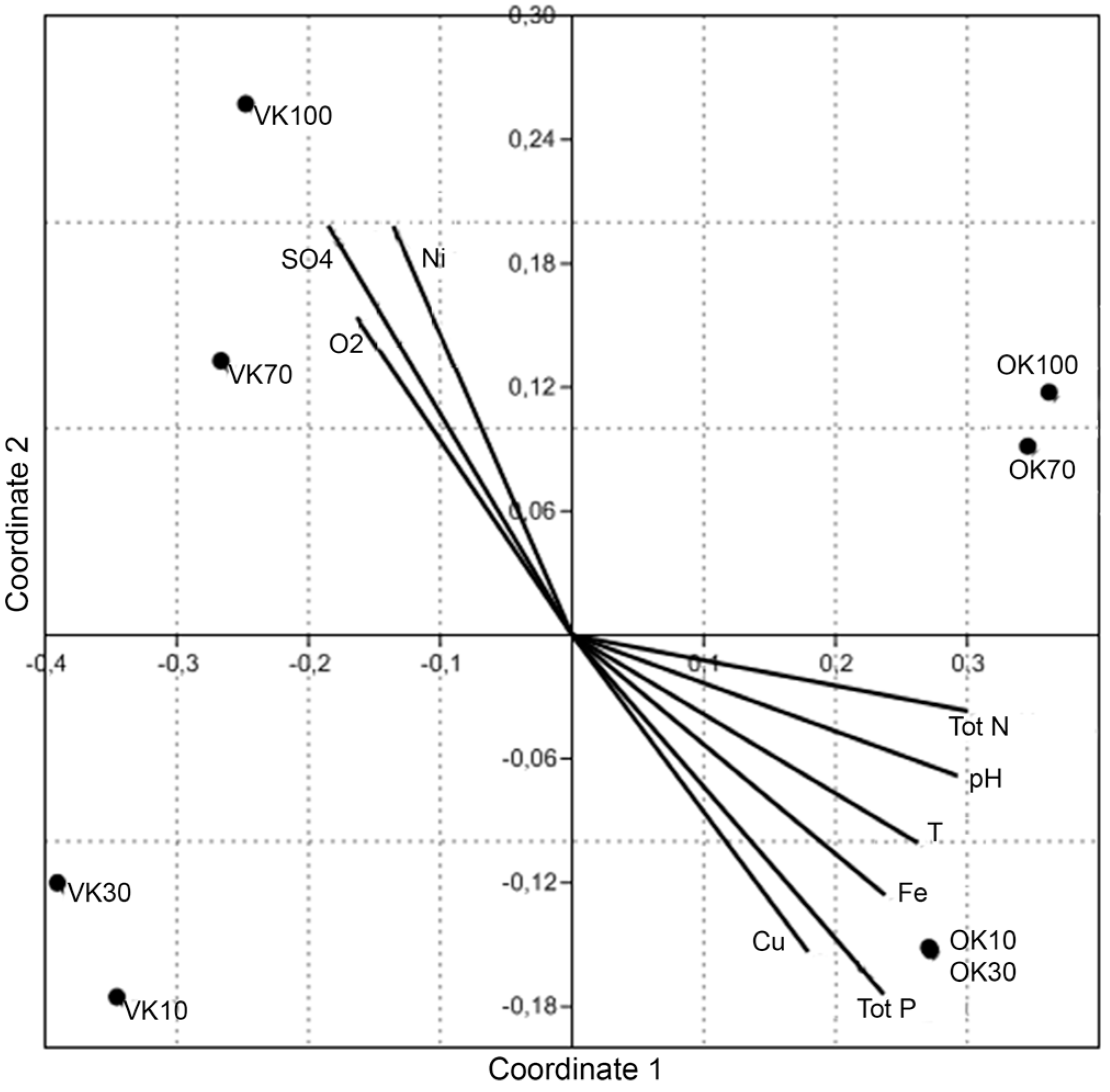
2.10. Statistical Analyses
3. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | VK 10 m | VK 30 m | VK 70 m | VK 100 m | OK 10 m | OK 30 m | OK 70 m | OK 100 m |
|---|---|---|---|---|---|---|---|---|
| T °C | 5.70 | 5.70 | 5.60 | 5.6 | 7.7 | 6.90 | 6.5 | 6.9 |
| O2 mg/L | 0 | 0.34 | 0.84 | 0.48 | 0 | 0.19 | 0.13 | 0 |
| O2% | 0 | 2.70 | 6.70 | 3.8 | 0 | 1.6 | 1.1 | 0 |
| pH | 7.4 | 7.4 | 7 | 7 | 9.80 | 9.80 | 9.0 | 9.80 |
| Tot N mg/L | 0.16 | 0.16 | 0.42 | 0.39 | 4.7 | 4.7 | 4.7 | 4.7 |
| Tot P μg/L | 31 | 32 | 9 | 9 | 76 | 86 | 46 | 58 |
| SO42− mg/L | 86 | 85 | 480 | 500 | 1.1 | 1.2 | <0.5 | 0.54 |
| Fe mg/L | 1 | 1 | 1 | 1 | 45 | 61 | 19 | 25 |
| Cu μg/L | 1.50 | 1.80 | 1.10 | 1.10 | 17 | 29 | 6.60 | 3.4 |
| Ni μg/L | 180 | 180 | 1700 | 1800 | 290 | 410 | 79 | 140 |

| Parameter | VK 10 m | VK 30 m | VK 70 m | VK 100 m | OK 10 m | OK 30 m | OK 70 m | OK 100 m |
|---|---|---|---|---|---|---|---|---|
| TNC mL−1 | 4.8 × 105 | 4.8 × 105 | 6.8 × 104 | 6.2 × 104 | 1.4 × 106 | 1.3 × 106 | 9.8 × 105 | 1.2 × 106 |
| SEM | 1.2 × 104 | 1.9 × 104 | 1.5 × 104 | 1.1 × 104 | 1.1 × 105 | 2.3 × 105 | 5.9 × 104 | 1.6 × 105 |
| dsrB mL−1 | 1.5 × 103 | 2.0 × 102 | 1.2 × 102 | 3.1 × 102 | 1.6 × 103 | 6.7 × 102 | 3.7 × 103 | 2.9 × 103 |
| SEM | 4.78 ×102 | 3.38 ×101 | 6.03 × 101 | 1.26 × 102 | 3.36 × 102 | 6.01 × 101 | 1.91 × 103 | 9.7 × 102 |
| dsrB/TNC ratio | 0.0030 | 0.0004 | 0.0018 | 0.0050 | 0.0012 | 0.0005 | 0.0027 | 0.0023 |
| Phylogenetic Group | OTU | VK 10 | VK 30 | VK 70 | VK 100 | OK 10 | OK 30 | OK 70 | OK 100 |
|---|---|---|---|---|---|---|---|---|---|
| ε-Proteobacteria/Sulfurimonas | 1 | ● | a1 | ||||||
| ε-Proteobacteria/Sulfurimonas | 2 | ● | b1 | ||||||
| β-Proteobacteria/Comamonadaceae | 3 | ● | g2 | ● | c4 | ||||
| β-Proteobacteria/unclassified | 4 | ● | c1 | ||||||
| β-Proteobacteria/unclassified | 5 | ● | b2 | ||||||
| β-Proteobacteria/unclassified | 6 | ● | d1 | ||||||
| γ-Proteobacteria/Methylobacter | 7 | ● | e1 | ||||||
| β-Proteobacteria/unclassified | 8 | ● | a3 | ||||||
| β-Proteobacteria/unclassified | 9 | ● | e4 | ||||||
| Ignavibacter | 10 | ● | h2 | ● | d4 | ||||
| β-Proteobacteria/Curvibacter | 11 | ● | b3 | ||||||
| γ-Proteobacteria/Methylobacter | 12 | ● | f1 | ● | c2 | ||||
| β-Proteobacteria/unclassified | 13 | ● | c3 | ● | f4 | ||||
| γ-Proteobacteria/Methylobacter | 14 | d2 | |||||||
| β-Proteobacteria/Methylotenera | 15 | ● | e2 | ||||||
| β-Proteobacteria/Methylotenera | 16 | ● | g1 | ● | ● | d3 | |||
| β-Proteobacteria/Methylobacter | 17 | ● | g4 | ||||||
| β-Proteobacteria/Methylotenera | 18 | ● | h1 | ● | f2 | ||||
| β-Proteobacteria/Methylobacter | 19 | ● | e3 | ||||||
| β-Proteobacteria/Hydrogenophaga | 20 | ● | h4 | ||||||
| β-Proteobacteria/Hydrogenophaga | 21 | ● | f3 | ● | a5 | ||||
| β-Proteobacteria/Rhodocyclaceae | 22 | ● | g3 | ● | b5 | ||||
| Thermodesulfovibrio | 23 | ● | h3 | ● | c5 | ||||
| Bacillus | 24 | d5 | |||||||
| Bacillus | 25 | ● | a4 | ● | e5 | ||||
| α-Proteobacteria/Rubritepida | 26 | ● | b4 | ● | f5 | ||||
| β-Proteobacteria | 27 | a2 | ● | ● |





4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Moncur, M.C.; Ptacek, C.J.; Hayashi, M.; Blowes, D.W.; Birks, S.J. Seasonal cycling and mass-loading of dissolved metals and sulfate discharging from an abandoned mine site in northern Canada. Appl. Geochem. 2014, 41, 176–188. [Google Scholar] [CrossRef]
- Luptakova, A.; Kusnierova, M. Bioremediation of acid mine drainage contaminated by SRB. Hydrometallurgy 2005, 77, 97–102. [Google Scholar] [CrossRef]
- Simate, G.S.; Ndlovu, S. Acid mine drainage: Challenges and opportunities. Int. J. Eng. Res. Appl. 2014, 2, 1785–1803. [Google Scholar] [CrossRef]
- Bosch, J.; Meckenstock, R.U. Rates and potential mechanism of anaerobic nitrate-dependent microbial pyrite oxidation. Biochem. Soc. Trans. 2012, 40, 1280–1283. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, C.J.; Jacobsen, O.S.; Elberling, B.; Aamand, J. Microbial oxidation of pyrite coupled to nitrate reduction in anoxic groundwater sediment. Environ. Sci. Technol. 2009, 43, 4851–4857. [Google Scholar] [CrossRef] [PubMed]
- Banks, D.; Younger, P.L.; Arnesen, R.-T.; Iversen, E.R.; Banks, S.B. Mine-water chemistry: The good, the bad and the ugly. Environ. Geol. 1997, 32, 157–174. [Google Scholar] [CrossRef]
- Johnson, D.B.; Hallberg, K.B. Acid mine drainage remediation options: A review. Sci. Total Environ. 2005, 338, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Wolkersdorfer, C. Water management at abandoned flooded underground mines. In Fundamentals, Tracer Tests, Modelling, Water Treatment, 1st ed.; Springer: New York, NY, USA, 2008; p. 465. [Google Scholar]
- Vestola, E.; Mroueh, U.-M. Sulfaatinpelkistyksen hyödyntäminen happamien kaivosvesien käsittelyssä. Opas louhoskäsittelyn hallintaan. In In Situ Treatment of Acid Mine Drainage by Sulphate Reducing Bacteria. Guide to the Pit Lake Treatment; VTT Tiedotteita, Research Notes 2422; VTT: Espoo, Finland, 2008; p. 58. [Google Scholar]
- Gaál, G. Geological setting and intrusion tectonics of the Kotalahti nickel-copper deposit, Finland. Bull. Geol. Soc. Finl. 1980, 52, 101–128. [Google Scholar]
- Kahma, A. The main sulphide ore belt of Finland between Lake Ladoga and the Bothnian Bay. Bull. Geol. Soc. Finl. 1978, 50, 39–43. [Google Scholar]
- Mäkinen, J.; Makkonen, H.V. Petrology and structure of the Palaeoproterozoic (1.9 Ga) Rytky nickel sulphide deposit, central Finland: A comparison with the Kotalahti nickel deposit. Miner. Depos. 2004, 39, 405–421. [Google Scholar] [CrossRef]
- Aluehallintovirasto Itä-Suomi, Dnro ISAVI/94/04.08/2013 (2014). Available online: https://www.avi.fi/documents/10191/1029174/isavi_paatos_96_2014_1–2014–12–9.pdf/d7e2df5a-b7c5–424b-9a36-e0d902bde345 (accessed on 7 July 2015).
- Aluehallintovirasto Itä-Suomi, 2011 Aluehallintovirasto Itä-Suomi, Dnro ISAVI/283/04.08/2010 (2011). Available online: http://www.avi.fi/documents/10191/56914/isavi_paatos_78_2011_1–2011–8-30.pdf (accessed on 7 July 2015).
- Determination of pH of Water; SFS 3012; Finnish Standards Association: Helsinki, Finland, 1979.
- Water Quality. Determination of Dissolved Oxygen. Iodometric Method; SFS-EN 25813; Finnish Standards Association: Helsinki, Finland, 1993.
- Water Quality. Determination of Nitrogen. Part 1: Method Using Oxidative Digestion with Peroxodisulfate; SFS-EN ISO 11905-1:en; Finnish Standards Association: Helsinki, Finland, 1998.
- Water Quality. Determination of Phosphorus. Ammonium Molybdate Spectrometric Method; SFS-EN 1189; Finnish Standards Association: Helsinki, Finland, 1997.
- Water Quality. Determination of Dissolved Anions by Liquid Chromatography of Ions. Part 1: Determination of Bromide, Chloride, Fluoride, Nitrate, Nitrite, Phosphate and Sulfate; SFS-EN ISO 10304-1; Finnish Standards Association: Helsinki, Finland, 2007.
- Water Quality. Application of Inductively Coupled Plasma Mass Spectrometry (ICP-MS). Part 1: General Guidelines (ISO 17294-1:2004); SFS-EN ISO 17294-1:en; Finnish Standards Association: Helsinki, Finland, 2004.
- Water Quality. Application of Inductively Coupled Plasma Mass Spectrometry (ICP-MS). Part 2: Determination of 62 Elements (ISO 17294-2:2003); SFS-EN ISO 17294-2:en; Finnish Standards Association: Helsinki, Finland, 2003.
- Water Quality. Digestion for the Determination of Selected Elements in Water. Part 2: Nitric Acid Digestion (ISO 15587-2:2002); SFS-EN ISO 15587-2; Finnish Standards Association: Helsinki, Finland, 2002.
- Geets, J.; Borremans, B.; Diels, L.; Springael, D.; Vangronsveld, J.; van der Lelie, D.; Vanbroekhoven, K. DsrB gene-based DGGE for community and diversity surveys of sulfate-reducing bacteria. J. Microbiol. Methods 2006, 66, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Roger, A.J.; Flax, J.L.; Brusseau, G.A.; Stahl, D.A. Phylogeny of dissimilatory sulfite reductases supports an early origin of sulfate respiration. J. Bacteriol. 1998, 180, 2975–2982. [Google Scholar] [PubMed]
- Nübel, U.; Engelen, B.; Felske, A.; Snaidr, J.; Wieshuber, A.; Amann, R.I.; Ludwig, W.; Backhaus, H. Sequence heterogeneities of genes encoding 16S rRNAs in Paenibacillus polymyxa detected by temperature gradient gel electrophoresis. J. Bacteriol. 1996, 78, 5636–5643. [Google Scholar]
- Laitila, A.; Kotaviita, E.; Peltola, P.; Home, S.; Wilhelmson, A. Indigeneous microbial community of barley greatly influences grain germination and malt quality. J. Inst. Brew. 2007, 113, 9–20. [Google Scholar] [CrossRef]
- Guidon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–123. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Castro, H.F.; Williams, N.H.; Ogram, A. Phylogeny of sulphate-reducing bacteria. FEMS Microbiol. Ecol. 2000, 31, 1–9. [Google Scholar] [PubMed]
- Nevatalo, L. Bioreactor Applications Utilizing Msophilic Sulphate-Reducing Bacteria for Treatment of Mine Wastewaters at 9–35 °C. Ph.D. Thesis, Tampere University of Technology, Tampere, Finland, 10 September 2010. [Google Scholar]
- Knoblauch, C.; Jorgensen, B.B. Effect of temperature on sulphate reduction, growth rate and growth yield in five psychrophilic sulphate-reducing bacteria from Arctic sediments. Environ. Microbiol. 1999, 1, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Knoblauch, C.; Sahm, K.; Jorgensen, B.B. Psychrophilic sulphate-reducing bacteria isolated from permanently cold Arctic marine sediments: Description of Desulfofrigus oceanense gen. nov., sp. nov., Desulfofrigus fragile sp. nov., Desulfofaba gelida gen. nov., sp. nov., Desulfotalea psychrophila gen. nov., sp. nov. and Desulfotalea arctica sp. nov. Int. J. Syst. Bacteriol. 1999, 49, 1631–1643. [Google Scholar] [PubMed]
- Isaksen, M.F.; Jorgensen, B.B. Adaptation of psychrophilic and psychrotrophic sulphate-reducing bacteria to permanently cold marine environments. Appl. Environ. Microbiol. 1996, 62, 408–414. [Google Scholar] [PubMed]
- Dar, S.A.; Bijmans, M.F.M.; Dinkla, I.J.T.; Geurkink, B.; Lens, P.N.L.; Dopson, M. Population dynamics of a single-stage sulfidogenic bioreactor treating synthetic zinc-containing waste streams. Microb. Ecol. 2009, 58, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Koschorreck, M.; Geller, W.; Neu, T.; Kleinsteuber, S.; Kunze, T.; Trosiener, A.; Wendt-Potthoff, K. Structure and function of the microbial community in an in situ reactor to treat an acidic mine pit lake. FEMS Microbiol. Ecol. 2010, 73, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Auvinen, H.; Nevatalo, L.M.; Kaksonen, A.H.; Puhakka, J.A. Low-temperature (9 °C) AMD treatment in a sulfidogenic bioreactor dominated by a mesophilic Desulfomicrobium species. Biotechnol. Bioeng. 2009, 104, 740–751. [Google Scholar] [PubMed]
- Cook, K.L.; Whitehead, T.R.; Spence, C.; Cotta, M.A. Evaluation of the sulphate-reducing bacterial population associated with stored swine slurry. Anaerobe 2008, 14, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Bomberg, M.; Arnold, M.; Kinnunen, P. Bioremediation and metal resistant bacteria in a closed, cold northern mine. Adv. Mater. Res. 2015, in press. [Google Scholar]
- Wartiainen, I.; Hestnes, A.G.; McDonald, I.R.; Svenning, M.M. Methylobacter tundripaludum sp. nov., a methane-oxidizing bacterium from Arctic wetland soil on the Svalbard islands, Norway (78° N). Int. J. Syst. Evol. Microbiol. 2006, 56, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J. The methanotrophs—The families methylococcaceae and methylocystaceae. In The Prokaryotes; Dworkin, E., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 5, pp. 266–289. [Google Scholar]
- Kalyuzhnaya, M.G.; Bowerman, S.; Lara, J.C.; Lidstrom, M.E.; Chistoserdova, L. Methylotenera mobilis gen. nov., sp. nov., an obligately methylamine-utilizing bacterium within the family Methylophilaceae. Int. J. Syst. Evol. Microbiol. 2006, 56, 2819–2823. [Google Scholar] [CrossRef] [PubMed]
- Kalyuzhnaya, M.; Lidstrom, M.; Chistoserdova, L. Real-time detection of actively metabolizing microbes by redox sensing as applied to methylotroph populations in Lake Washington. ISME J. 2008, 2, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, F.; Takai, K.; Kobayashi, H.; Nealson, K.H.; Horikoshi, K. Sulfurimonas autotrophica gen. nov., sp. nov., a novel sulfur-oxidizing ε-proteobacterium isolated from hydrothermal sediments in the Mid-Okinawa Trough. Int. J. Syst. Evol. Microbiol. 2003, 53, 1801–1805. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Suzuki, M.; Nakagawa, S.; Miyazaki, M.; Suzuki, Y.; Inagaki, F.; Horikoshi, K. Sulfurimonas paralvinellae sp. nov., a novel mesophilic, hydrogen- and sulfur-oxidizing chemolithoautotroph within the Epsilon-Proteobacteria isolated from a deep-sea hydrothermal vent polychaete nest, reclassification of Thiomicrospira denitrificans as Sulfurimonas denitrificans comb. nov. and emended description of the genus Sulfurimonas. Int. J. Syst. Evol. Microbiol. 2006, 56, 1725–1733. [Google Scholar] [PubMed]
- Heider, J.; Schüle, K. Anaerobic biodegradation of hydrocarbons including methane. In The Prokaryotes—Prokaryotic Physiology and Biochemistry; DeLonf, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 605–634. [Google Scholar]
- Widdel, F.; Boetius, A.; Rabus, R. Anaerobic biodegradation of hydrocarbons including methane. In The Prokaryotes; Dworkin, E., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 2, pp. 1028–1049. [Google Scholar]
- Alarico, S.; Rainey, F.A.; Empadinhas, N.; Schumann, P.; Nobre, M.F.; da Costa, M.S. Rubritepida flocculans gen. nov., sp. nov., a new slightly thermophilic member of the alpha-1 subclass of the Proteobacteria. Syst. Appl. Microbiol. 2002, 25, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Christova, N.; Tuleva, B.; Nikolova-Damyanova, B. Enhanced hydrocarbon biodegradation by a newly isolated Bacillus subtilis strain. Z. Naturforsch. C 2004, 59, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Iino, T.; Mori, K.; Uchino, Y.; Nakagawa, T.; Harayama, S.; Suzuki, K. Ignavibacterium album gen. nov., sp. nov., a moderately thermophilic anaerobic bacterium isolated from microbial mats at a terrestrial hot spring and proposal of Ignavibacteria classis nov., for a novel lineage at the periphery of green sulfur bacteria. Int. J. Syst. Evol. Microbiol. 2010, 60, 1376–1382. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, Y.; Muramatsu, M.; Imachi, H.; Narihiro, T.; Ohashi, A.; Harada, H.; Hanada, S.; Kamagata, Y. Thermodesulfovibrio aggregans sp. nov. and Thermodesulfovibrio thiophilus sp. nov., anaerobic, thermophilic, sulfate-reducing bacteria isolated from thermophilic methanogenic sludge, and emended description of the genus Thermodesulfovibrio. Int. J. Syst. Evol. Microbiol. 2008, 58, 2541–2548. [Google Scholar] [CrossRef] [PubMed]
- Isaksen, M.F.; Bak, F.; Jorgensen, B.B. Thermophilic sulphate-reducing bacteria in cold marine sediment. FEMS Microbiol. Ecol. 1994, 14, 1–8. [Google Scholar] [CrossRef]
- Rabus, R.; Brüchert, V.; Amann, J.; Könneke, M. Physiological response to temperature changes of the marine, sulphate-reducing bacterium Desulfobacterium autotrophicum. FEMS Microbiol. Ecol. 2002, 42, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Nevatalo, L.M.; Bijmans, M.F.M.; Lens, P.N.L.; Kaksonen, A.H.; Puhakka, J.A. The effect of sub-optimal temperature on specific sulfidogenic activity of mesophilic SRB in an H2-fed membrane bioreactor. Process Biochem. 2010, 45, 363–368. [Google Scholar] [CrossRef]
- Bless, D.; Park, B.; Nordwick, S.; Zaluski, M.; Joyce, H.; Hiebert, R.; Clavelot, C. Operational lessons learned during bioreactor demonstrations for acid rock drainage treatment. Mine Water Environ. 2008, 27, 241–250. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bomberg, M.; Arnold, M.; Kinnunen, P. Characterization of the Bacterial and Sulphate Reducing Community in the Alkaline and Constantly Cold Water of the Closed Kotalahti Mine. Minerals 2015, 5, 452-472. https://doi.org/10.3390/min5030452
Bomberg M, Arnold M, Kinnunen P. Characterization of the Bacterial and Sulphate Reducing Community in the Alkaline and Constantly Cold Water of the Closed Kotalahti Mine. Minerals. 2015; 5(3):452-472. https://doi.org/10.3390/min5030452
Chicago/Turabian StyleBomberg, Malin, Mona Arnold, and Päivi Kinnunen. 2015. "Characterization of the Bacterial and Sulphate Reducing Community in the Alkaline and Constantly Cold Water of the Closed Kotalahti Mine" Minerals 5, no. 3: 452-472. https://doi.org/10.3390/min5030452
APA StyleBomberg, M., Arnold, M., & Kinnunen, P. (2015). Characterization of the Bacterial and Sulphate Reducing Community in the Alkaline and Constantly Cold Water of the Closed Kotalahti Mine. Minerals, 5(3), 452-472. https://doi.org/10.3390/min5030452