
The Insulin-like Growth Factor System and Colorectal Cancer
by
 , , ,
, , ,
Nikola Gligorijević
* ,
,
Zorana Dobrijević
,
Miloš Šunderić
,
Dragana Robajac
,
Danilo Četić
,
Ana Penezić
,
Goran Miljuš
and
Olgica Nedić
Institute for the Application of Nuclear Energy, Department for Metabolism, University of Belgrade, Banatska 31b, 11080 Belgrade, Serbia
*
Author to whom correspondence should be addressed.
Life 2022, 12(8), 1274; https://doi.org/10.3390/life12081274
Submission received: 20 July 2022
/
Revised: 10 August 2022
/
Accepted: 12 August 2022
/
Published: 20 August 2022
(This article belongs to the Special Issue Insulin-Like Growth Factor from Physiology to Cancer)
Abstract
:Insulin-like growth factors (IGFs) are peptides which exert mitogenic, endocrine and cytokine activities. Together with their receptors, binding proteins and associated molecules, they participate in numerous pathophysiological processes, including cancer development. Colorectal cancer (CRC) is a disease with high incidence and mortality rates worldwide, whose etiology usually represents a combination of the environmental and genetic factors. IGFs are most often increased in CRC, enabling excessive autocrine/paracrine stimulation of the cell growth. Overexpression or increased activation/accessibility of IGF receptors is a coinciding step which transmits IGF-related signals. A number of molecules and biochemical mechanisms exert modulatory effects shaping the final outcome of the IGF-stimulated processes, frequently leading to neoplastic transformation in the case of irreparable disbalance. The IGF system and related molecules and pathways which participate in the development of CRC are the focus of this review.
1. Introduction
Insulin-like growth factors (IGFs), mitogenic and metabolic peptides, are involved in the etiology and progression of the colorectal cancer (CRC). The association was confirmed at the level of cell lines, animal models and patients with CRC. Initial studies on the role of IGFs, their receptors (IGFRs), high-affinity binding proteins (IGFBPs), IGFBP proteases, highly related insulin and its receptor (IR) expanded to investigate connections with an array of physiological molecules and systems which together build a very complex network not yet fully defined and characterized. Current research includes IGFBP-related proteins (IGFBP-RP, also known as low-affinity IGFBPs), messenger RNA (mRNA), responsible genes, circular-, micro- and long non-coding RNAs (circRNA, miRNA and lncRNA), RNA-binding proteins, other protein and nucleic acid binding partners inside specific compartments of cells and membrane-bound, present in the extracellular matrix and in the circulation. There are many pathways of interference, with the effects primarily based on the activation of the IGF system.
IGFs are known to exert endocrine, paracrine and autocrine effects. Many cell types synthesize IGFs, IGFRs and IGFBPs. The response of cells to IGFs depends on various factors, many of them being specific for the particular microenvironment at a specific pathophysiological moment. Modulatory effects of different agents have been detected, positive and negative, both at the expression level and on the activity of individual components of the IGF system. Tight control mechanisms are necessary to maintain the equilibrium in the normal physiological state since overexpression or excessive activation/accessibility of some components, together with a reduced activity of suppressor molecules, can lead to disbalance and neoplastic transformation. In this review, we focus on the IGF system and related molecules/mechanisms that participate in the initiation and the development of CRC.
2. Epidemiology of Colorectal Cancer
The world’s cancer burden represents one of the biggest hurdles for human life improvement [1]. According to estimates made by the World Health Organization (WHO) in 2019 [2], cancer is the primary or secondary cause of death of people below the age of 70 in 112 out of 183 countries, and the third or fourth cause in 23 additional countries [3].
Colorectal cancer is highly ranked for its incidence and mortality—it is the third most frequently occurring malignancy, responsible for 10.0% of all cancer cases for both sexes, and it was the second most fatal cancer in 2018, with 9.4% of reported deaths [3]. More than 1.9 million new CRC cases occurred in 2020, resulting in 935,000 new deaths globally [3]. Complications, mortality, treatment side effects, health care service utilization and medical costs associated with CRC present a considerable burden worldwide. There are, however, significant geographical differences in CRC incidence and mortality. The age-standardized rate (ASR) of CRC incidence was found to be six times higher in high human development index (HDI) countries compared to low HDI countries, with a similar ratio found for ASR of mortality (Figure 1) [4,5,6].
Categorization of CRC cases can be performed by age—patients less than 50 years old belong to an early-onset CRC category, whereas older patients belong to a late-onset CRC category. In the late-onset CRC, HDI and EAPC (estimated annual percentage change) are negatively correlated for both sexes, implying that present lower incidence in low HDI countries might worsen in the next 30 to 50 years, posing a more serious health issue [5].
Men are more prone to developing CRC than women. Even though there is no discernable difference in the early onset of colon cancer between sexes (IRR = 0.98; 95% CI 0.94, 1.07), the incidence of early-onset rectal cancer (IRR = 1.08; 95% CI 1.04, 1.12), late-onset colon cancer (IRR = 1.50; 95% CI 1.37, 1.62) and late-onset rectal cancer (IRR = 2.03; 95% CI 1.84, 2.21) is considerably higher in males [5].
Smaller scale (i.e., continent- or region-specific) studies confirmed the correlation between an increased CRC incidence and high HDI. Chung et al. [7] reported such findings for Asia, while Sierra et al. [8] corroborated the impact of HDI on CRC in Central and South America.
Worldwide CRC incidence and mortality have generally increased in the 21st century and will continue to rise in the future, as estimated by numerous global [4,5,9,10], continental [7,8,11,12], regional [13,14,15,16,17] and country-specific [18,19,20,21] studies. Such trends warrant new and more effective screening and prevention strategies to be developed from evidence-based research.
3. Pathophysiology of the Colorectal Cancer
The etiology of CRC is multifactorial and usually represents a combination of the environmental and genetic factors. The process of neoplastic transformation consists of a series of (epi)genetic alterations that lead to changes in normal mucosa of the colon, resulting in cancer, which has the potential to culminate in metastasis in distant tissues [22]. Colon modifications can be divided into discrete stages which tissue passes, as proposed by Fearon and Vogelstein (Figure 2). Each stage is characterized by specific shifts in the genetic make-up of the cell [23].
There are several mechanisms that lead to neoplastic changes in colonocytes. The most common is the chromosomal instability (CIN) pathway, which is characterized by the loss of heterozygosity and various chromosomal abnormalities [24]. One of the first changes that occurs in this mechanism is dysregulation in the WNT/APC/β-cat pathway, responsible for the expression of APC gene, a tumor suppressor, leading to an increased presence of β-catenin, which induces proliferation, differentiation, migration and adhesion of colorectal cells [25,26]. Other changes follow, accompanied by new mutations and progression from benign to malignant state. The process includes mutations in KRAS and p53 genes and their binding partners [27,28,29,30,31]. Another cause of CRC, which roughly accounts for 15% of cases, is a derangement in the DNA mismatch repair system (MMR), which is in charge of the production of proteins that recognize and repair single nucleotide mismatches arising in the replication process [32,33]. The third mechanism leading to CRC is based on hypermethylation of CpG islands in promoter regions of genes involved in cell cycle regulation, apoptosis, adhesion and invasion [34,35,36].
Chronic inflammation plays a pivotal role in disease etiology. Various inflammatory markers, such as tumor necrosis factor-α (TNF-α), signal transducer and activator of transcription 3 (STAT3), interleukin-6 (IL-6) and C-reactive protein (CRP), are associated with pathogenesis of CRC. TNF-α promotes tumor growth, proliferation and metastasis. IL-6 stimulates the expression of STAT3, which is a transcription factor that induces the expression of various genes that play active roles in cell proliferation, differentiation and apoptosis, such as Bcl-2, CyclinD1, ICAM-1 and MMP2-9 [37,38,39]. Cyclooxygenase-2 (COX-2) is an inducible cyclooxygenase that is up-regulated by cytokines, growth factors and tumor promoters. It is overexpressed in 40% of human colorectal adenomas, compared to normal epithelial tissue [40]. COX-2 regulates prostaglandin (PG) synthesis, apoptosis, angiogenesis and tumor invasiveness, being a mediator between the inflammation and neoplastic transformation in CRC.
4. Insulin-like Growth Factors and Colorectal Cancer
4.1. Insulin-like Growth Factors
The IGF system is composed of two IGF peptides (IGF-I and IGF-II), two receptor types (IGF1R and IGF2R) and six high-affinity IGF-binding proteins (IGFBP-1 to -6) [41]. The first information on the so-called somatomedins, now known as IGFs, appeared in the 1950s [42]. Many cell types synthesize IGFs, but IGFs in circulation are mostly derived from the liver [41]. IGF-I synthesis in the liver is under the control of the growth hormone (GH), whereas the production of IGF-II is less GH-dependent. IGF-II is the major growth factor during fetal development. During adult life, its role seems to be of less significance compared to IGF-I. The importance of IGF-II increases in tumor development, as some tumors, including those of the colon and rectum, produce high concentrations enabling autocrine/paracrine stimulation of the cell growth [41,43]. Not only the fully processed IGF-II, but also the so-called high molecular weight IGF-II pro-peptides can exert growth-promoting effects in tumors, as well as general hypoglycemia. Non-islet cell tumor hypoglycemia in CRC patient is a rare event [44].
IGF-I regulates growth and development during post-natal life, and its circulating concentration declines with age after puberty and depends on lifestyle and nutritional habits [41]. Circulating and local concentrations of IGF-I and IGF-II often do not correlate, as was shown for the serum and colorectal tissues [45]. The expression level of IGF-I mRNA, and particularly IGF-II mRNA, is several times greater in colon cancer specimens than the actual concentration of proteins [46]. In some cases, a positive correlation was found between tumor grade and IGF-II expression. Paracrine and autocrine mechanisms of IGF-II action in colon cancer were documented, as cancer-associated fibroblasts in tumor stroma were identified as a source of up-regulated IGF-II which, in turn, activates pro-survival IGF1R/IR signaling [47].
4.2. Insulin-like Growth Factor Receptors
Receptor IGF1R is a tyrosine kinase which becomes activated after binding of IGF-I or IGF-II, inducing its auto-phosphorylation and a signaling pathway which will end up primarily in the growth-promoting events [41,48]. Phosphorylated receptor substrates are a common signaling meeting point of several receptor systems including those for other growth factors, integrins, cytokines [49]. IGF1R is widely expressed in the gastrointestinal tract, the most in the colonic crypt proliferating cells [50].
Insulin and insulin receptor (IR) are tightly connected to the IGF system, as IGFs exert some of their functions (primarily metabolic) after binding to IR. Structural homology between IGF1R and IR enables formation of the hybrid receptor (HyR) consisting of IGF1R and IR hemi-receptors, capable of binding both insulin and IGFs [51]. IR exists in two isoforms, IR-A and IR-B, with the IR-A form mainly present in fetal and cancer tissues [52,53]. HyR containing IR-A is approximately two times more expressed in CRC tissue than in normal epithelial cells [54].
There is also functional homology between IGF1R, IR and HyR, as they all initiate cell signaling cascade, relying on the activation of insulin receptor substrates (IRS-1 to -6) [55]. The relative importance of the intracellular pathways that will serve as effectors of the signal transduction is dependent on the specific cell context and related to other intrinsic and external factors. Besides by synthesis, the availability of receptors is regulated by endocytosis and proteolysis [56].
Receptor IGF2R (also known as cation-independent mannose-6-phosphate receptor, CI-MPR) binds only IGF-II with high affinity [57]. This interaction most often leads to the removal of IGF-II and its degradation, but also some signaling events are mediated through this pathway. The extracellular domain of IGF2R can be found in a soluble form in physiological fluids exerting a role of the circulating IGF-II scavenger [58].
As previously mentioned, COX-2 and PGs play important roles in the development of CRC and the expression of COX-2 mRNA, and PGE2 is higher in intestinal cells which overexpress IGF-II, the effect being mediated via IGF1R [59]. Both IGF1R and IGF2R are present on the epithelial cells of the intestine. Overexpression of IGF1R is detected in CRC samples compared to adjacent normal mucosa [60,61]. A degree of IGF1R expression is correlated with an increased risk of metastasis in CRC patients with Dukes’ score C [62], but not with patient survival [63]. The glycosylation pattern of both IGF1R and IGF2R changes in colon tissue due to CRC [64], and receptors are also modified by oxidation [65]. These post-translational modifications possibly affect ligand binding, activation and/or other interactions of receptors.
When measuring the presence of specific components of the IGF system, one should be aware of the difference in their expression levels at different locations in healthy tissue. The expression of IGF-I, IGF-II and IGF1R mRNAs is significantly higher in rectum compared to ascending colon, while there is no difference in the expression of IGF2R mRNA [45]. Furthermore, the analysis of two samples from the same location obtained during the same biopsy procedure may differ due to different expression of individual members of the IGF system in different cell types, i.e., epithelial and stromal cells.
4.3. Cell Signaling via Insulin-like Growth Factor Receptors
Binding of IGFs to IGF1R initiates auto-phosphorylation of its intracellular tyrosine (Tyr) residues, docking of IRS and activation of intracellular kinases, such as mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase (PI3K), inducing a phosphorylation cascade leading to growth-stimulating events [41,48]. IGF1R and IR signaling rely predominantly on the activation of IRS-1 and IRS-2 (Figure 3). The type of the substrate activated influences downstream signaling. IRS-1 expression seems to be inversely correlated with cell differentiation in CRC [66], whereas IRS-2 expression positively correlates with the transformation of the intestinal epithelium to adenocarcinoma [67]. Certain polymorphisms regarding these two proteins are associated with a risk of CRC [68]. An increase in phosphorylated IR was found to accompany transformation of normal colorectal epithelium to CRC, suggesting the involvement of IR in carcinogenesis [69].
Activated PI3K further stimulates the activation of Akt and the expression of B-cell lymphoma extra-large (Bcl-xL) protein, which regulates apoptosis, and the mammalian target of rapamycin (mTOR) signaling pathway, which regulates cell growth, proliferation, motility, survival and the general metabolism. Overexpression of IGF1R, Bcl-xL and mTOR inhibits apoptosis and contributes to colon cancer cell survival and invasion [70]. At the same time, activated Akt inhibits glycogen synthase kinase 3-β (GSK3β), causing translocation of β-catenin to the nucleus and transcription of genes involved in cell proliferation [71]. IGF1R-dependent pathways also play a significant role in the resistance to therapeutic agents, which we examine in the last section of this review.
IGF-I signaling was identified as a contributing factor in the Warburg effect, increased glucose uptake and a metabolic switch from butyrate oxidation to aerobic glycolysis, which is one of the hallmarks of CRC [72]. Insulin/IGF signaling was shown to induce the expression of hypoxia-inducible factor 1α (HIF-1α), which is the main regulator of Warburg effect [73]. Activation of both IGF1R-related pathways, PI3K/Akt/mTOR and Raf/MAPK, was found to be associated with glycolytic metabolism in CRC [72]. Furthermore, the relation between these signaling pathways and glucose metabolic reprograming in CRC could be partially attributed to the activities of lncRNA CRNDE [74,75].
4.4. Insulin-like Growth Factor Binding Proteins
IGF-binding proteins are synthetized in different cell types. They form complexes with IGFs which may reside within blood vessels (mostly tertiary complexes, maintaining the IGF reservoir) or they may cross the endothelial barrier to transport IGFs to peripheral tissues (mostly binary complexes) [41]. The most prevalent IGFBPs in the circulation, namely, IGFBP-3, IGFBP-2 and IGFBP-1, are derived from the liver. Some IGFBPs are synthetized locally where they bind IGFs and regulate their presentation to receptors. IGFBPs can exert either inhibitory or potentiating effect on IGFs, and their affinity for IGFs can be altered by post-translational modifications and interactions with other proteins, cells or extracellular matrix [76]. IGFs need to be in a free form in order to interact with receptors and IGFBP proteases enable their liberation [76]. IGFBPs cleaved by proteolysis have reduced affinity for ligands facilitating IGFR capture of IGFs.
IGFBP-3 is the major IGF-binding protein. Approximately 85% of the circulating IGFs are bound to it, together with an acid-labile subunit [77]. Investigations on cancer risk take into account not only the concentration of IGFs and IGFBPs, but particularly the ratio of IGFs and IGFBP-3. An unfavorable ratio was found in patients with CRC [78]. Although an increased ratio IGF-I/IGFBP-3 was related to the initiation of CRC, the relation could not be clearly established with the progression or the outcome of CRC. Most studies report on the protective role of IGFBP-3 in CRC [79,80], but there are also contrasting results [81,82]. Higher expression level of IGFBP-3 mRNA was detected in normal colon than in cancer. In contrast to normal colon, where this mRNA was distributed both in epithelial tissue and stroma, it is mainly present in a stromal component of the cancer tissue, suggesting its paracrine role [82]. A correlation between the expression of mRNA IGFBP-3, disease staging or tumor location in the colon was reported [83]. High expression of IGFBP-3 mRNA correlates with lymph node metastasis and poor outcome. An increased sialylation of IGFBP-3 originating from patients with CRC was also detected [84].
Colon cancer cell lines most often secrete IGFBP-2 and it is frequently overexpressed in CRC tissues, especially glandular [60]. According to some studies, an increased circulating concentration of IGFBP-2 is associated with CRC and in correlation with the advancement of a disease [60,85,86]. According to the others, IGFBP-1 and IGFBP-2 concentrations in blood are not associated with colon or rectal cancer [77,87,88].
IGFBP-4, which exerts only inhibitory action on IGFs, is often overexpressed in cancer and in correlation with a state of differentiation. IGFBP-4 was shown to inhibit growth of some colon cancer cells [89]. Stimulation of Caco-2 cells with IL-1β and IL-6 was shown to modulate secretion of IGFBP-2 and IGFBP-4 [90], confirming a connection between inflammation and carcinogenesis. IGFBP-6 is supposed to act as tumor suppressor and its concentration is lower in metastatic compared to non-metastatic CRC cells [91]. Furthermore, IGFBP-6 expression in CRC tissue is inversely correlated with the survival of patients [92] and connected to the tumor suppressor activity of SEMA3B [93].
IGFBPs exert IGF-dependent and IGF-independent roles. Some IGFBPs bind to specific receptors on the cell surface and/or to nuclear receptors [89,94]. IGF-independent actions of IGFBP-3 in CRC are primarily connected to its pro-apoptotic role through NF-κB inhibition [95], interaction with retinoid X receptor (RXR)/Nur77 [96] or p53-dependent signaling cascade [97]. No clear IGF-independent action of IGFBP-1 was observed in CRC [98], although increased levels of IGFBP-1 and decreased incidence of CRC seem to be linked in women [99]. Elevated serum level of IGFBP-2 was nominated as a potential biomarker of CRC [85]. However, an increased expression of IGFBP-2 mRNA in colon cancer cells and tissue is accompanied by an increased proteolytic degradation [100], calling for further investigations to define its potential local role in CRC. IGF-independent actions of IGFBP-6 are mainly tumor protective and are considered as possible paths for cancer therapy [101]. The same authors also reported a promotion of LIM 1215 colon cancer cell migration stimulated by IGFBP-6 [102], which leaves the final conclusion on the role of IGFBP-6 in colon cancer still to be assessed.
Some data concerning IGFBP concentrations and involvement in CRC (and in other malignancies as well) seem controversial due to the complexity of the IGF system, its relations with other physiological molecules and a variety of signaling mechanisms. Age, BMI, gender, physical activity, nutritional habits and possibly ethnicity contribute to disagreements reported in different studies [103]. Obesity-related insulin/IGF signaling pathways influence CRC development by evading apoptosis, whereas reported effects of exercise are inconsistent and without clear conclusions [104,105]. Maintenance of glucose control via nutritional habits has a major influence on the insulin/IGF axis in relation to CRC [106].
IGFBP-7, the most studied member of the IGFBP-RP family, shares certain signaling pathways with IGFBP-3 [107]. According to some studies, it is up-regulated in colon cancer, but according to others, it is down-regulated. Contradictory results were reported both in patients and colon cancer cell lines. Some results suggest IGFBP-3/IGFBP-7 interaction, but this crosstalk needs additional confirmation. The association seems to participate in cancer progression and metastasis [108].
The putative roles of IGFBPs are summarized in Table 1.
Due to the complexity of CRC etiology as well as the complex role of the IGF system in CRC pathology, it is hard to distinguish mechanisms of IGFBP regulation which can be directly linked to the incidence of CRC from those primarily involved in further development of cancer. The choice of the study group has a significant influence on the results. A study conducted on older men from Australia suggested that higher IGFBP-3 concentrations are associated with an increased incidence of CRC [109]. In a study on the general population in China (both men and women), the initiation of CRC was associated with an increased ratio IGF-I/IGFBP-3 as well as reduced IGFBP-3 concentration [78]. As for IGFBP-2 concentration, the data are controversial, since some suggest association with CRC incidence, while others fail to confirm the link. In IGFBP-2 transgenic mice which overexpress IGFBP-2, CRC incidence was similar to that of the control group of animals, but the volume of adenomas was more than two times smaller [110]. Although it is accepted that IGFBP-4 inhibits the action of IGF-I, and thus, CRC progression, very little is known about its role in CRC incidence. IGFBP-4 gene therapy could not prevent CRC in mice inoculated with HT-29 colon cancer cells, but an increased apoptosis was detected [111]. In a case-controlled study conducted on women from New York, lower concentrations of IGFBP-1 were linked to a decreased incidence of CRC [99]. The same study also demonstrated that women in a quintile with the highest IGFBP-3 concentration were at higher risk for CRC development, compared to those in a quintile with the lowest IGFBP-3 concentration.
4.5. Insulin-like Growth Factor Binding Protein Proteinases
Matrix metalloproteinases (MMPs) regulate remodeling of the extracellular matrix (ECM), which is an important event for metastatic progression of tumor, and some of them also act as IGFBP proteases, ensuring a simultaneous release of IGFs to stimulate IGF1R signaling. MMP-7, often increased in malignant diseases, is capable of degrading all six IGFBPs [112]. CRC cells which overexpress MMP-7 resist chemotherapy due to intensive IGFBP-3 degradation and release of IGFs [113]. MMP-19 is another proteinase which degrades IGFBP-3 in CRC [114]. IGF1R activation was identified as a promoting factor in the synthesis of MMPs.
Other MMPs, such as ADAM17 and DAM28, specifically cleave IGFBP-3 and increase IGF-I availability in CRC [115]. Pregnancy-associated protein A is IGFBP-4 specific proteinase and is expressed in the colon tissue [89]. Less IGFBP-4 is found in the cancer tissue than in the normal. Cathepsins, plasmin, kallikreins, pepsin, calpain and caspase also act as IGFBP proteinases [81,91,116,117] and are increased in CRC. An interplay between IGFs, IGF1R, IGFBPs and IGFBP proteinases plus other modifying factors define a response of the cell, including its (un)responsiveness to potential tumor-suppressive agents [48,118].
4.6. Genetic, Epigenetic and Post-Transcriptional Regulators of the Insulin-like Growth Factor System
4.6.1. Genetic Variants
Profiling of genetic alterations in various components of the insulin/IGF system in CRC tissue and cell lines demonstrated a whole spectrum of mutations which can contribute to carcinogenesis. Common mutations in genes encoding downstream members of the insulin/IGF pathway, such as KRAS and BRAF, are known predictors of the biological potential of CRC and also dictate a response to treatment relying on IR/IGF1R inhibitors [119]. Another type of canonically driven mutations in CRC, affecting TP53, alters insulin/IGF signaling due to an interplay between p53/MDM2 and IGF1R-activated pathways [120]. Mutations in PIK3CA are observed in 20–25% of patients with CRC, suggesting potential application of IGF1R-targeting immunotherapeutic approaches [121]. Additionally, a loss of expression of PTEN, an important negative regulator of the PI3K/AKT signaling pathway, characterizes a large portion of CRC cases and is partially caused by mutational events [122].
Apart from somatic mutations, inherited germline variants in the insulin/IGF pathway genes were investigated in the context of CRC carcinogenesis. However, associations of these candidate variants with CRC susceptibility were found in a relatively small number of studies and were rarely replicated. Reports include both potentially functional and tagging variants within IGF-I (rs12423791, rs1019731, rs5742632, rs5742678, rs5742694, rs2033178, rs2373722, rs1520220, rs10735380, rs6220, rs6214, rs35767, 19-CA repeat polymorphism), IR (rs10426094, rs1052371), IGFBP-3 (rs2854744, rs2854746, rs3110697, rs2132572, rs35440925), IGF1R (rs2229765, rs7166348), IRS-1 (rs1801278) and IRS-2 (rs1805097, rs2289046, rs754204, rs4773082) [68,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139].
4.6.2. DNA Methylation and Imprinting
Epigenetic mechanisms were determined as a cause of dysregulated expression of several IGF-related genes abundantly occurring in CRC patients. For instance, IGFBP-3 promoter region is commonly hypermethylated in CRC, representing potential diagnostic and predictive biomarker [140,141]. A reduced expression of PTEN is partially caused by hypermethylation, which is frequent in CRC with microsatellite instability [122,142]. On the other hand, a genome-wide DNA methylation analysis demonstrated hypomethylation of more than 30 genes involved in PI3K/AKT signaling [143].
IGF-II was the first gene found to be paternally expressed due to genomic imprinting, since differentially methylated region (DMR) is methylated on the maternal allele [144,145]. An aberrant imprinting IGF-II locus was detected in CRC, leading to an overexpression of IGF-II and, subsequently, to higher proliferation rate and invasiveness [146,147,148,149]. The same imprinted region also contains a gene encoding H19, a long non-coding RNA (lncRNA) with a known regulatory role in the molecular basis of CRC carcinogenesis [150,151].
4.6.3. Regulatory RNAs
MicroRNAs (miR) are important regulators of genes involved in the insulin/IGF signaling with their oncogenic or tumor-suppressive properties in CRC and in malignancy in general. For instance, the expression of IRS-1 is regulated by miR-145, a commonly deregulated microRNA in CRC [152,153]. The same gene was also identified as a target of tumor-suppressive miR-497 and miR-126 [154,155]. MiR-143, originating from the same precursor as miR-145, as well as members of the let-7 family, miR-497, miR-184 and miR-98, were identified as regulators of IGF1R in CRC [156,157,158,159,160]. Common tumor-suppressive microRNA, miR-375, directly targets PI3K (PIK3CA) in CRC, while miR-92 exhibits oncogenic activity through the inhibition of a set of tumor-suppressors, including PTEN [161,162]. Similarly, miR-21 and miR-26a target PTEN, but lead to enhanced PI3K/AKT signaling [163,164]. Additional IGF-related targets of miR-143 include components of the RAS/RAF/MAPK pathway, highlighting the importance of tumor-suppressive activities of this microRNA in the molecular basis of CRC [153,165,166]. Various genetic variants within the miR-143/145 locus were found to be associated with CRC susceptibility, risk of developing other malignancies and cancer aggressiveness [167,168,169,170,171,172,173,174,175,176].
Overexpression of an oncogenic lncRNA, colorectal neoplasia differentially expressed (CRNDE), is triggered in early stages of carcinogenesis in colorectal tissue [74]. Multiple splice variants are generated and the resulting isoforms have different intracellular localizations, which underlies their functional differences. Nuclear isoforms contribute to the Warburg effect in CRC by modulating transcription and acting as regulators of glucose and lipid metabolism. The expression of splice variants with retained introns is strongly regulated by insulin and IGF-I, as these variants are downstream targets of both PI3K/Akt/mTOR and RAF/MAPK signaling cascades. Furthermore, CRNDE nuclear transcripts regulate insulin/IGF signaling by a feedback mechanism [74]. Another mechanism connecting oncogenic properties of CRNDE with insulin/IGF signaling refers to sponging of microRNAs known to regulate the expression of key components of this signaling cascade, such as IRS-1 [177]. An increased expression of CRNDE was found both in tumor tissue and in plasma from CRC patients, qualifying this lncRNA as a potential circulatory biomarker of CRC [178].
Another lncRNA potentially involved in CRC pathogenesis is PTENP1, a homologue of tumor-suppressive gene PTEN [179]. By acting as a microRNA sponge, PTENP1 regulates the expression of PTEN. A similar mechanism is proposed for KRAS1P transcript, which regulates KRAS/RAF/MAPK signaling cascade [179]. PVT1, another lncRNA up-regulated in CRC, functions as an indirect regulator of IRS-1 expression through interaction with miR-214-3p [180].
4.6.4. RNA-Binding Proteins
RNA-binding proteins (RNBs), together with mRNAs, ribosome subunits and other proteins, form ribonucleoproteins which function as RNA regulators. RNBs, such as IGF-II mRNA binding proteins (IGF2BPs), can be differentially expressed in cancer. IGF2BP family consists of three members which exert negative effects in the late phase of embryo development; their expression is lost in adults and may be reactivated in tumors [181,182]. IGF2BP1 was evaluated as an independent prognostic marker in CRC associated with the responsiveness to chemotherapy [183]. Although IGF2BP expression is generally related to worse prognosis, opposing overall effects of different IGF2BP members in different cells were detected [48].
4.7. Insulin-like Growth Factor Signaling and the Associated Signaling Pathways
A complex and interrelated network of events governs growth and spreading of cancer cells, with the IGF system being both modulated by other factors and exerting modulation of signals driven by a variety of other agents. This section briefly overviews other signaling pathways which interfere with IGF signaling.
Tumor growth depends on the oxygen supply and CRC is an example of a solid cancer where hypoxia emerges due to the inability of the vascular system to adequately supply oxygen to the growing tumor tissue [184]. IGF-I activity can induce the expression of HIF-1α, which may further trigger neovascularization in order to secure oxygen and nutrients for further tumor progression. Thus, hypoxia and activity of IGF-I are two parallel paths for the induction of HIF-1 in cancer [73], and the expression of HIF-1α and vascular endothelial growth factor (VEGF) in CRC tissues was proposed to serve as a biomarker of cancer progression [185]. Even more, hypoxia induces up-regulation of IGF synthesis through the STAT5b pathway [186], promoting tumor growth [187].
A crosstalk between IGF1R and several other membrane-associated molecules modifies IGF signaling. Cadherins are calcium-dependent transmembrane proteins which mediate cell–cell adhesion, whereas their intracellular domain participates in cell signaling. Cadherins mediate epithelial–mesenchymal transition (EMT), which is a crucial process in embryonic and tumor development. E-cadherin is down-regulated and N-cadherin is up-regulated during EMT [188]. IGF1R without IGF and E-cadherin in close proximity interact. This partnership is uncoupled upon IGF binding, as was shown in colon cancer cell lines [189]. Alpha V integrin is another binding partner creating a dynamic complex controlled by α-catenin, which may contribute to the cancer cell motility [189]. Involvement of α-catenin in IGF-I-induced cellular migration, but not invasion, in colon cancer cells was documented [190].
Discoidin domain receptor 1 (DDR1) is a tyrosine kinase which binds extracellular collagen, undergoes phosphorylation of the intracellular domain and participates in a signaling cascade leading to tumor progression [191]. Contrary to E-cadherin, IGF1R and DDR1 associate upon IGF binding, further stimulating the expression of IGF1R and downstream signaling [192]. DDR1 is expressed in colon mucosa and to a greater extent in cancer than in normal tissue. The association of its expression and tumor prognosis, however, awaits further evaluation [193].
Decorin is proteoglycan able to suppress the activity of some tyrosine kinases, including IGF1R [194,195]. A dichotomy of its activity on the IGF system was noted, depending on the cell type [196]. Decorin deficiency in Dcn-/- mice was shown to stimulate EMT and colon cancer metastasis [197].
IGFBP-3 and transferrin (Tf), an iron transporter and growth-promoting factor [198], form complexes which interact with Tf receptor (TfR). A reduced serum concentration of IGFBP-3/Tf complexes and an increased expression of TfR1 on colon cell membranes was found in patients with CRC [199]. IGFBP-1 and IGFBP-2 form complexes with alpha-2-macroglobulin (α2M) in the circulation [200,201]. The amount of IGFBP-2/α2M complexes is decreased in patients with CRC, although the total concentration of IGFBP-2 is increased [86]. Oxidative conditions modify the structure of IGFBP-binding partners, influencing the formation of complexes [202]. The exact role of these complexes is still unknown, but they contribute to the (re)distribution of IGFBPs between several molecular forms.
5. Therapeutic Potential of the Insulin-like Growth Factor Signaling Pathways in Treating Colorectal Cancer
As already said, neoplastic cell growth and proliferation is driven via PI3K, Akt, mTOR and MAPK pathways after IGF-I binding to IGF1R [203]. The primary strategy in treating colon cancer includes the arrest of IGF1R overexpression by small inhibitors, antibodies or the inhibition of its ligands. A discovery of OSI-906, later termed linsitinib, a drug with a promising inhibitory effect on two receptor kinases, IR and IGF1R, was reported in 2009 [204]. This dual inhibitor intervenes in the process of autophosphorylation, demonstrating an antiproliferative effect on different tumor cell lines, including colorectal. One possibility to treat metastatic CRC is to use regorafenib (multi-kinase inhibitor) together with linsitinib and aspirin (both being IGF1R inhibitors) [205]. Linsitinib and aspirin reduce the resistance of CRC cells to regorafenib, enabling body weight gain and an increase in the survival rate of model animals (i.e., male mice). Leiphrakpam et al. reported on the down-regulation of X-linked inhibitor of apoptosis (XIAP) in CRC xenografted male mice driven by MK-0646 (mAb that blocks IGF1R and IGF2R, also known as dalotuzumab) and linsitinib [206].
An interplay of various factors keeps cancer cells alive. Hyperactivation of IGF1R, for example, results in the resistance to epidermal growth factor receptor (EGFR) inhibition in RAS wild-type metastatic CRC via up-regulation of PI3K/AKT pathway, implying that targeting both receptors might be a promising therapy for metastatic CRC. Unfortunately, a combination of cetuximab (mAb targeting EGFR) and MK-0646 or IMC-A12 (mAb that blocks IGF1R, also known as cixutumumab) did not lead to the expected results, as suggested by two studies both involving male and female participants [207,208]. A phase I trial employing a combination of cixutumumab (anti-IGF1R antibody) and selumetinib (MEK ½ inhibitor) obtained promising results offering the evidence of the health benefit and target inhibition in a cohort of 30 patients, including those with CRC [209]. A combination of ganitumab (IGF1R mAb) and conatumumab (a pro-apoptotic death receptor 5 agonist) also exerted promising effects in the Colo-205 xenograft model, but caused no response in approximately 80 tested individuals (both male and female), some of whom were patients with CRC [210].
MEDI-573 (mAb to IGF-I and IGF-II) application inhibited tumor growth in a CRC female mouse model that over-expresses IGF-II, and this effect was enhanced if MEDI-573 was combined with other known therapeutics such as trastuzumab, AZD2014, AZD5363, selumetinib or cetuximab [211]. AvFc lectibody, a combination of the human immunoglobulin G1 Fc and Avaren (high mannose glycan-binding lectin), selectively recognizes a range of cancer cell lines including colon cells [212]. Although the observed cytotoxic effects were thoroughly investigated only in the lung cancer cell lines, it was confirmed that targets of this lectibody are EGFR and IGF1R. Since aberrant glycosylation is a hallmark of cancer, and altered glycosylation of IGFRs in the colon tissue of CRC patients was reported [64], the lectibody treatment may be considered as a novel approach in cancer therapy.
IGF1R depletion/inhibition sensitizes CRC cells to radiotherapy (converting them to radiosensitive), as shown in HT-29 and SW480 cell lines where IGF1R was inhibited by NVP-ADW742 [213] and in HT-29, SW480 and DLD-1 cells pretreated with BMS-754807 [214]. The inhibitory effect of BMS-754807 on colon cancer cell growth was stronger compared to the effect of linsitinib, and the anti-neoplastic effect was mostly independent of IGF1R [215]. A prolonged treatment of colon cancer cells with BMS-754807 and GSK1838705A (inhibitors of IGF1R and IR) leads to cell survival, due to the activation of ribosomal protein S6 kinase 1 [216]. In GEO tumors, BI885578 administration (inhibitor of both IGF1R and IR) induces apoptosis and inhibition of cell proliferation in female mice [217]. Zhu et al. reported on an anti-proliferative, anti-migration, anti-invasion and inhibitory effect of isovitexin on EMT in human colon cancer cell lines and xenograft tumor model on female mice [218]. The levels of signaling molecules involved in the IGF1R signaling pathway were decreased after this treatment, pointing to the mechanism of the isovitexin action. Simvastatin down-regulates IGF1R expression and pro-apoptotic ERK activation in human HT-29 cells [219]. Phloroglucinol treatment inhibits or decreases the expression of various IGF1R downstream signaling molecules, such as RAS, MAPK, ERK, PI3K, Akt and mTOR, in HT-29 cells [220].
In addition to synthetic molecules and antibodies, numerous plant metabolites were investigated as potential therapeutics in the treatment of cancer. Curcumol, isolated from Rhizoma Curcumae, inhibited proliferation and induced apoptosis in LoVo cells, and inhibited CRC in xenograft models of nude mice, via inhibition of IGF1R and activation of p38 MAPKs [221]. Application of manuka honey (alone or in combination with 5-fluorouracil) on HCT-116 cell line decreased physical parameters of colonospheres and the survival ability of cancer cells, but also induced apoptosis via down-regulation of apoptosis inhibitors, including IGFs and IGF1R [222]. The expression levels of IGF-I and IGF1R were reduced, whereas the level of IGFBP-3 was normalized after Bifidiobacterium longum BAA-999 (with or without lycopene) was administrated to CD-A male mice in an azoxymethane-dextrane sulfate sodium-induced CRC model [223]. Polypeptides from Arca subcrenata Lischke inhibited growth of HT-29 cells and suppressed tumor growth in male mouse xenograft by reducing IGF1R phosphorylation and inhibiting IGF-I/IGF1R signaling activation [224]. Carnosic acid treatment also suppressed the growth of HT-29 cells; decreased the number of tumors and circulating concentrations of leptin, adiponectin, insulin and IGF-I; and reduced the expression of IR in a male mice model [225]. Fucoidan, sulfated polysaccharide from brown algae, inhibited IGF1R signaling through the IRS-1/PI3K/Akt pathway in HT-29 cells [226]. Laminarin, another polysaccharide from brown algae, decreased phosphorylation of ERK and MAPK and, consequently, IGF1R-dependent proliferation in the same cells [227]. Curcumin decreased the expression of IR and IGF1R in 5-fluorouracil-treated SW480 cells, and this down-regulation correlated with decreased proliferation and migration of cells [228]. Luteolin, a fruit and vegetable flavone, decreased IGF-II production in HT-29 cells and down-regulated the activation of PI3K/Akt and ERK1/2 pathways [229]. Cinnamaldehyde, isolated from stem bark of Cinnamomum cassia, inhibited PI3K/Akt signaling in SW480, HCT116 and LoVo cells [230]. Considering the incidence and mortality rates of CRC, together with limitations of the existing therapeutic procedures, the search for both natural and synthetic anti-CRC agents is expected to intensify.
It is sometimes necessary to apply multiple approaches, i.e., a combination of different therapeutics and therapies. A serious limitation in examining therapeutic potentials of various substances can be found in translation from cell line models to xenografts and further to humans as the results are unsatisfactory or the cancer cells are nonresponsive to just one therapeutic. It must also be noted that the gender equality was neglected in most of the mentioned studies, as some of the experiments were performed only on male or female animals (rarely both). Furthermore, the number of studies on humans is still low, which is understandable due to the limited number of potentially effective therapeutics, their toxicity and effective dose, as well as ethical principles related to studies on humans.
6. Conclusions
The insulin-like growth factor system exerts multiple effects and at multiple levels in colon cancer transformation. The results obtained with tumor cells, animals and humans, however, show differences, some related to a tumor/cell type, but other factors (besides methodological) seem to be very influential. Caution is suggested with the generalization of effects, as crosstalk between the IGF system and other biochemical pathways regularly occurs and contributes to the final cell signaling event. Similarly, the complexity of interrelated (patho)physiological paths and outcomes contributes to the resistance to therapeutic approaches in CRC treatment.
Author Contributions
Conceptualization, N.G. and O.N.; writing—original draft preparation, N.G., O.N., D.R., G.M., A.P., M.Š., Z.D. and D.Č.; writing—review and editing, N.G. and O.N.; visualization, N.G., D.Č., M.Š. and O.N.; supervision, O.N.; project administration, O.N.; funding acquisition, O.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Education, Science and Technological Development of the Republic of Serbia (grant number 451-03-68/2022-14/200019).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The Ever-increasing Importance of Cancer as a Leading Cause of Premature Death Worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Global Health Estimates 2020: Deaths by Cause, Age, Sex, by Country and by Region. 2000–2019. Available online: https://www.who.int/data/gho/data/themes/mortality-and-global-health-estimates/ghe-leading-causes-of-death (accessed on 15 April 2022).
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Patterns and Trends in Colorectal Cancer Incidence and Mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.; Li, Y.; Wang, W.; Feng, W.; Shi, O.; Wang, Q. International Incidence Trends in Early- and Late-Onset Colorectal Cancer: A Population-Based Study. Int. J. Colorectal Dis. 2020, 35, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- United Nations Development Programme. Human Development Reports. 2020. Available online: https://hdr.undp.org/content/human-development-report-2020 (accessed on 20 April 2022).
- Chung, R.Y.-N.; Tsoi, K.K.F.; Kyaw, M.H.; Lui, A.R.; Lai, F.T.T.; Sung, J.J.-Y. A Population-Based Age-Period-Cohort Study of Colorectal Cancer Incidence Comparing Asia against the West. Cancer Epidemiol. 2019, 59, 29–36. [Google Scholar] [CrossRef]
- Sierra, M.S.; Forman, D. Burden of Colorectal Cancer in Central and South America. Cancer Epidemiol. 2016, 44, S74–S81. [Google Scholar] [CrossRef] [Green Version]
- Araghi, M.; Soerjomataram, I.; Jenkins, M.; Brierley, J.; Morris, E.; Bray, F.; Arnold, M. Global Trends in Colorectal Cancer Mortality: Projections to the Year 2035. Int. J. Cancer 2019, 144, 2992–3000. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.C.S.; Huang, J.; Lok, V.; Wang, J.; Fung, F.; Ding, H.; Zheng, Z.-J. Differences in Incidence and Mortality Trends of Colorectal Cancer Worldwide Based on Sex, Age, and Anatomic Location. Clin. Gastroenterol. Hepatol. 2021, 19, 955–966.e61. [Google Scholar] [CrossRef]
- Arhin, N.; Ssentongo, P.; Taylor, M.; Olecki, E.J.; Pameijer, C.; Shen, C.; Oh, J.; Eng, C. Age-Standardised Incidence Rate and Epidemiology of Colorectal Cancer in Africa: A Systematic Review and Meta-Analysis. BMJ Open 2022, 12, e052376. [Google Scholar] [CrossRef]
- Cardoso, R.; Guo, F.; Heisser, T.; Hackl, M.; Ihle, P.; de Schutter, H.; van Damme, N.; Valerianova, Z.; Atanasov, T.; Májek, O.; et al. Colorectal Cancer Incidence, Mortality, and Stage Distribution in European Countries in the Colorectal Cancer Screening Era: An International Population-Based Study. Lancet Oncol. 2021, 22, 1002–1013. [Google Scholar] [CrossRef]
- Makhlouf, N.A.; Abdel-Gawad, M.; Mahros, A.M.; Lashen, S.A.; Zaghloul, M.; Eliwa, A.; Elshemy, E.E.; Ali-Eldin, Z.; Abdeltawab, D.; El-Raey, F.; et al. Colorectal Cancer in Arab World: A Systematic Review. World J. Gastrointest. Oncol. 2021, 13, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Vekic, B.; Dragojevic-Simic, V.; Jakovljevic, M.; Kalezic, M.; Zagorac, Z.; Dragovic, S.; Zivic, R.; Pilipovic, F.; Simic, R.; Jovanovic, D.; et al. A Correlation Study of the Colorectal Cancer Statistics and Economic Indicators in Selected Balkan Countries. Front. Public Health 2020, 8, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokki, I.; Papana, A.; Campbell, H.; Theodoratou, E. Estimating the Incidence of Colorectal Cancer in South East Asia. Croat. Med. J. 2013, 54, 532–540. [Google Scholar] [CrossRef] [Green Version]
- Graham, A.; Adeloye, D.; Grant, L.; Theodoratou, E.; Campbell, H. Estimating the Incidence of Colorectal Cancer in Sub–Saharan Africa: A Systematic Analysis. J. Glob. Health 2012, 2, 020404. [Google Scholar] [CrossRef] [Green Version]
- Ali Hussein Alhurry, A.M.; Rezaianzadeh, A.; Rahimikazerooni, S.; Abdzaid Akool, M.; Bahrami, F.; Shahidinia, S.S.; Pourahmad, M. A Review of the Incidence of Colorectal Cancer in the Middle East. Ann. Colorectal Res. 2017, 5, e46292. [Google Scholar] [CrossRef] [Green Version]
- Soriano, L.C.; Soriano-Gabarró, M.; García Rodríguez, L.A. Trends in the Contemporary Incidence of Colorectal Cancer and Patient Characteristics in the United Kingdom: A Population-Based Cohort Study Using the Health Improvement Network. BMC Cancer 2018, 18, 402. [Google Scholar] [CrossRef] [Green Version]
- Cayuela, L.; Rodriguez, S.; Giráldez Gallego, Á.; Cayuela, A. Regional Differences in Colorectal Cancer Mortality Trends, Spain (1980–2018). Rev. Española Enferm. Dig. 2020, 113, 570–575. [Google Scholar] [CrossRef]
- Austin, H.; Jane Henley, S.; King, J.; Richardson, L.C.; Eheman, C. Changes in Colorectal Cancer Incidence Rates in Young and Older Adults in the United States: What Does It Tell Us about Screening. Cancer Causes Control 2014, 25, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Zhang, L.; Liu, W.; Ding, Y.; Yin, J.; Ren, R.; Li, Q.; Chen, Y.; Shen, J.; Tan, X.; et al. Trends in Cancer Mortality in China from 2004 to 2018: A Nationwide Longitudinal Study. Cancer Commun. 2021, 41, 1024–1036. [Google Scholar] [CrossRef]
- Colussi, D.; Brandi, G.; Bazzoli, F.; Ricciardiello, L. Molecular Pathways Involved in Colorectal Cancer: Implications for Disease Behavior and Prevention. Int. J. Mol. Sci. 2013, 14, 16365–16385. [Google Scholar] [CrossRef] [Green Version]
- Fearon, E.R.; Vogelstein, B. A Genetic Model for Colorectal Tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Bardi, G.; Sukhikh, T.; Pandis, N.; Fenger, C.; Kronborg, O.; Heim, S. Karyotypic Characterization of Colorectal Adenocarcinomas. Genes Chromosomes Cancer 1995, 12, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.M.; Zilz, N.; Beazer-Barclay, Y.; Bryan, T.M.; Hamilton, S.R.; Thibodeau, S.N.; Vogelstein, B.; Kinzler, K.W. APC Mutations Occur Early during Colorectal Tumorigenesis. Nature 1992, 359, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, S.; Bodmer, W.F.; Bicknell, D.; Kaklamanis, L. Molecular Analysis of APC Mutations in Familial Adenomatous Polyposis and Sporadic Colon Carcinomas. Lancet 1992, 340, 626–630. [Google Scholar] [CrossRef]
- Malumbres, M.; Barbacid, M. RAS Oncogenes: The First 30 Years. Nat. Rev. Cancer 2003, 3, 459–465. [Google Scholar] [CrossRef]
- Guerrero, S.; Casanova, I.; Farré, L.; Mazo, A.; Capellà, G.; Mangues, R. K-Ras Codon 12 Mutation Induces Higher Level of Resistance to Apoptosis and Predisposition to Anchorage-Independent Growth than Codon 13 Mutation or Proto-Oncogene Overexpression. Cancer Res. 2000, 60, 6750–6756. [Google Scholar]
- Imamura, Y.; Morikawa, T.; Liao, X.; Lochhead, P.; Kuchiba, A.; Yamauchi, M.; Qian, Z.R.; Nishihara, R.; Meyerhardt, J.A.; Haigis, K.M.; et al. Specific Mutations in KRAS Codons 12 and 13, and Patient Prognosis in 1075 BRAF Wild-Type Colorectal Cancers. Clin. Cancer Res. 2012, 18, 4753–4763. [Google Scholar] [CrossRef] [Green Version]
- Baker, S.J.; Preisinger, A.C.; Jessup, J.M.; Paraskeva, C.; Markowitz, S.; Willson, J.K.; Hamilton, S.; Vogelstein, B. P53 Gene Mutations Occur in Combination with 17p Allelic Deletions as Late Events in Colorectal Tumorigenesis. Cancer Res. 1990, 50, 7717–7722. [Google Scholar]
- Lanza, G.; Matteuzzi, M.; Gafá, R.; Orvieto, E.; Maestri, I.; Santini, A.; del Senno, L. Chromosome 18q Allelic Loss and Prognosis in Stage II and III Colon Cancer. Int. J. Cancer 1998, 79, 390–395. [Google Scholar] [CrossRef]
- Fishel, R. Mismatch Repair, Molecular Switches, and Signal Transduction. Genes Dev. 1998, 12, 2096–2101. [Google Scholar] [CrossRef] [Green Version]
- Sinicrope, F.A.; Sargent, D.J. Molecular Pathways: Microsatellite Instability in Colorectal Cancer: Prognostic, Predictive, and Therapeutic Implications. Clin. Cancer Res. 2012, 18, 1506–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samowitz, W.S.; Albertsen, H.; Herrick, J.; Levin, T.R.; Sweeney, C.; Murtaugh, M.A.; Wolff, R.K.; Slattery, M.L. Evaluation of a Large, Population-Based Sample Supports a CpG Island Methylator Phenotype in Colon Cancer. Gastroenterology 2005, 129, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Teodoridis, J.M.; Hardie, C.; Brown, R. CpG Island Methylator Phenotype (CIMP) in Cancer: Causes and Implications. Cancer Lett. 2008, 268, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Bettington, M.; Walker, N.; Clouston, A.; Brown, I.; Leggett, B.; Whitehall, V. The Serrated Pathway to Colorectal Carcinoma: Current Concepts and Challenges. Histopathology 2013, 62, 367–386. [Google Scholar] [CrossRef]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and Colon Cancer. Gastroenterology 2010, 138, 2101–2114.e5. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.-T.; Wang, S.; Ye, Y.-J.; Du, R.-Y.; Cui, Z.-R.; Somsouk, M. Constitutive Activation of Stat3 Signaling Pathway in Human Colorectal Carcinoma. World J. Gastroenterol. 2004, 10, 1569. [Google Scholar] [CrossRef]
- Corvinus, F.M.; Orth, C.; Moriggl, R.; Tsareva, S.A.; Wagner, S.; Pfitzner, E.B.; Baus, D.; Kaufman, R.; Huber, L.A.; Zatloukal, K.; et al. Persistent STAT3 Activation in Colon Cancer Is Associated with Enhanced Cell Proliferation and Tumor Growth. Neoplasia 2005, 7, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Sinicrope, F.A.; Gill, S. Role of Cyclooxygenase-2 in Colorectal Cancer. Cancer Metastasis Rev. 2004, 23, 63–75. [Google Scholar] [CrossRef]
- LeRoith, D.; Holly, J.M.P.; Forbes, B.E. Insulin-like Growth Factors: Ligands, Binding Proteins, and Receptors. Mol. Metab. 2021, 52, 101245. [Google Scholar] [CrossRef]
- Salmon, W.D.; Daughaday, W.H. A Hormonally Controlled Serum Factor Which Stimulates Sulfate Incorporation by Cartilage in Vitro. J. Lab. Clin. Med. 1957, 49, 825–836. [Google Scholar]
- Kawamoto, K.; Onodera, H.; Kondo, S.; Kan, S.; Ikeuchi, D.; Maetani, S.; Imamura, M. Expression of Insulin-Like Growth Factor-2 Can Predict the Prognosis of Human Colorectal Cancer Patients: Correlation with Tumor Progression, Proliferative Activity and Survival. Oncology 1998, 55, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Teramae, S.; Miyamoto, H.; Muguruma, N.; Okada, Y.; Goji, T.; Kitamura, S.; Kimura, T.; Kimura, M.; Bando, Y.; Takayama, T. Insulin-like Growth Factor II-Producing Metastatic Colon Cancer with Recurrent Hypoglycemia. Clin. J. Gastroenterol. 2015, 8, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Vrieling, A.; Voskuil, D.W.; Bosma, A.; Majoor, D.M.; van Doorn, J.; Cats, A.; Depla, A.C.T.M.; Timmer, R.; Witteman, B.J.M.; Wesseling, J.; et al. Expression of Insulin-like Growth Factor System Components in Colorectal Tissue and Its Relation with Serum IGF Levels. Growth Horm. IGF Res. 2009, 19, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, A.; Adamek, A. Insulin-Like Growth Factor 2 (IGF2) Signaling in Colorectal Cancer—From Basic Research to Potential Clinical Applications. Int. J. Mol. Sci. 2019, 20, 4915. [Google Scholar] [CrossRef] [Green Version]
- Unger, C.; Kramer, N.; Unterleuthner, D.; Scherzer, M.; Burian, A.; Rudisch, A.; Stadler, M.; Schlederer, M.; Lenhardt, D.; Riedl, A.; et al. Stromal-Derived IGF2 Promotes Colon Cancer Progression via Paracrine and Autocrine Mechanisms. Oncogene 2017, 36, 5341–5355. [Google Scholar] [CrossRef]
- Mancarella, C.; Morrione, A.; Scotlandi, K. Novel Regulators of the IGF System in Cancer. Biomolecules 2021, 11, 273. [Google Scholar] [CrossRef]
- Samani, A.A.; Yakar, S.; LeRoith, D.; Brodt, P. The Role of the IGF System in Cancer Growth and Metastasis: Overview and Recent Insights. Endocr. Rev. 2007, 28, 20–47. [Google Scholar] [CrossRef]
- Davies, M.; Gupta, S.; Goldspink, G.; Winslet, M. The Insulin-like Growth Factor System and Colorectal Cancer: Clinical and Experimental Evidence. Int. J. Colorectal Dis. 2006, 21, 201–208. [Google Scholar] [CrossRef]
- Pandini, G.; Frasca, F.; Mineo, R.; Sciacca, L.; Vigneri, R.; Belfiore, A. Insulin/Insulin-like Growth Factor I Hybrid Receptors Have Different Biological Characteristics Depending on the Insulin Receptor Isoform Involved. J. Biol. Chem. 2002, 277, 39684–39695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belfiore, A.; Malaguarnera, R. Insulin Receptor and Cancer. Endocr. Relat. Cancer 2011, 18, R125–R147. [Google Scholar] [CrossRef] [Green Version]
- Scalia, P.; Giordano, A.; Williams, S.J. The IGF-II–Insulin Receptor Isoform-A Autocrine Signal in Cancer: Actionable Perspectives. Cancers 2020, 12, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frasca, F.; Pandini, G.; Scalia, P.; Sciacca, L.; Mineo, R.; Costantino, A.; Goldfine, I.D.; Belfiore, A.; Vigneri, R. Insulin Receptor Isoform A, a Newly Recognized, High-Affinity Insulin-Like Growth Factor II Receptor in Fetal and Cancer Cells. Mol. Cell. Biol. 1999, 19, 3278–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama, T.; Kyohara, M.; Terauchi, Y.; Shirakawa, J. The Roles of the IGF Axis in the Regulation of the Metabolism: Interaction and Difference between Insulin Receptor Signaling and IGF-I Receptor Signaling. Int. J. Mol. Sci. 2021, 22, 6817. [Google Scholar] [CrossRef]
- Rieger, L.; O’Connor, R. Controlled Signaling—Insulin-Like Growth Factor Receptor Endocytosis and Presence at Intracellular Compartments. Front. Endocrinol. 2021, 11, 620013. [Google Scholar] [CrossRef]
- El-Shewy, H.M.; Luttrell, L.M. Insulin-Like Growth Factor-2/Mannose-6 Phosphate Receptors. Vitam. Horm. 2009, 80, 667–697. [Google Scholar]
- Duplomb, L.; Chaigne-Delalande, B.; Vusio, P.; Raher, S.; Jacques, Y.; Godard, A.; Blanchard, F. Soluble Mannose 6-Phosphate/Insulin-Like Growth Factor II (IGF-II) Receptor Inhibits Interleukin-6-Type Cytokine-Dependent Proliferation by Neutralization of IGF-II. Endocrinology 2003, 144, 5381–5389. [Google Scholar] [CrossRef] [Green Version]
- di Popolo, A.; Memoli, A.; Apicella, A.; Tuccillo, C.; di Palma, A.; Ricchi, P.; Acquaviva, A.M.; Zarrilli, R. IGF-II/IGF-I Receptor Pathway up-Regulates COX-2 MRNA Expression and PGE2 Synthesis in Caco-2 Human Colon Carcinoma Cells. Oncogene 2000, 19, 5517–5524. [Google Scholar] [CrossRef] [Green Version]
- Mishra, L.; Bass, B.; Ooi, B.S.; Sidawy, A.; Korman, L.; Mönig, H. Role of Insulin-like Growth Factor-I (IGF-I) Receptor, IGF-I, and IGF Binding Protein-2 in Human Colorectal Cancers. Growth Horm. IGF Res. 1998, 8, 473–479. [Google Scholar] [CrossRef]
- Weber, M.M.; Fottner, C.; Liu, S.B.; Jung, M.C.; Engelhardt, D.; Baretton, G.B. Overexpression of the Insulin-like Growth Factor I Receptor in Human Colon Carcinomas. Cancer 2002, 95, 2086–2095. [Google Scholar] [CrossRef]
- Nakamura, M.; Miyamoto, S.; Maeda, H.; Zhang, S.; Sangai, T.; Ishii, G.; Hasebe, T.; Endoh, Y.; Saito, N.; Asaka, M.; et al. Low Levels of Insulin-Like Growth Factor Type 1 Receptor Expression at Cancer Cell Membrane Predict Liver Metastasis in Dukes’ C Human Colorectal Cancers. Clin. Cancer Res. 2004, 10, 8434–8441. [Google Scholar] [CrossRef] [Green Version]
- Heckl, S.M.; Pellinghaus, M.; Behrens, H.-M.; Krüger, S.; Schreiber, S.; Röcken, C. Questioning the IGF1 Receptor’s Assigned Role in CRC-a Case for Rehabilitation? BMC Cancer 2020, 20, 704. [Google Scholar] [CrossRef] [PubMed]
- Robajac, D.; Križáková, M.; Masnikosa, R.; Miljuš, G.; Šunderić, M.; Nedić, O.; Katrlík, J. Sensitive Glycoprofiling of Insulin-like Growth Factor Receptors Isolated from Colon Tissue of Patients with Colorectal Carcinoma Using Lectin-Based Protein Microarray. Int. J. Biol. Macromol. 2020, 144, 932–937. [Google Scholar] [CrossRef] [PubMed]
- Nedić, O.; Robajac, D.; Šunderić, M.; Miljuš, G.; Đukanović, B.; Malenković, V. Detection and Identification of Oxidized Insulin-like Growth Factor-Binding Proteins and Receptors in Patients with Colorectal Carcinoma. Free Radic. Biol. Med. 2013, 65, 1195–1200. [Google Scholar] [CrossRef]
- Esposito, D.L.; Aru, F.; Lattanzio, R.; Morgano, A.; Abbondanza, M.; Malekzadeh, R.; Bishehsari, F.; Valanzano, R.; Russo, A.; Piantelli, M.; et al. The Insulin Receptor Substrate 1 (Irs1) in Intestinal Epithelial Differentiation and in Colorectal Cancer. PLoS ONE 2012, 7, e36190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, E.; Poulogiannis, G.; McCaughan, F.; Mulholland, S.; Arends, M.J.; Ibrahim, A.E.K.; Dear, P.H. IRS2 is a Candidate Driver Oncogene on 13q34 in Colorectal Cancer. Int. J. Exp. Pathol. 2013, 94, 203–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slattery, M.L.; Samowitz, W.; Curtin, K.; Ma, K.N.; Hoffman, M.; Caan, B.; Neuhausen, S. Associations among IRS1, IRS2, IGF1, and IGFBP3 Genetic Polymorphisms and Colorectal Cancer. Cancer Epidemiol. Biomark. Prev. 2004, 13, 1206–1214. [Google Scholar] [CrossRef]
- Abbruzzese, C.; Diodoro, M.G.; Sperduti, I.; Mileo, A.M.; Pattaro, G.; de Salvo, L.; Cosimelli, M.; Perrotti, N.; Paggi, M.G. Detection of Phosphorylated Insulin Receptor in Colorectal Adenoma and Adenocarcinoma: Implications for Prognosis and Clinical Outcome. J. Cell. Physiol. 2015, 230, 562–567. [Google Scholar] [CrossRef]
- Sekharam, M.; Zhao, H.; Sun, M.; Fang, Q.; Zhang, Q.; Yuan, Z.; Dan, H.C.; Boulware, D.; Cheng, J.Q.; Coppola, D. Insulin-like Growth Factor 1 Receptor Enhances Invasion and Induces Resistance to Apoptosis of Colon Cancer Cells through the Akt/Bcl-x(L) Pathway. Cancer Res. 2003, 63, 7708–7716. [Google Scholar]
- Zhang, Q.; Wang, L.; Song, Z.; Qu, X. Knockdown of Type I Insulin-like Growth Factor Receptor Inhibits Human Colorectal Cancer Cell Growth and Downstream PI3K/Akt, WNT/β-Catenin Signal Pathways. Biomed. Pharmacother. 2015, 73, 12–18. [Google Scholar] [CrossRef]
- Kasprzak, A. Insulin-Like Growth Factor 1 (IGF-1) Signaling in Glucose Metabolism in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 6434. [Google Scholar] [CrossRef]
- Fukuda, R.; Hirota, K.; Fan, F.; Jung, Y.D.; Ellis, L.M.; Semenza, G.L. Insulin-like Growth Factor 1 Induces Hypoxia-Inducible Factor 1-Mediated Vascular Endothelial Growth Factor Expression, Which Is Dependent on MAP Kinase and Phosphatidylinositol 3-Kinase Signaling in Colon Cancer Cells. J. Biol. Chem. 2002, 277, 38205–38211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, B.C.; Graham, L.D.; Molloy, P.L. CRNDE, a Long Non-Coding RNA Responsive to Insulin/IGF Signaling, Regulates Genes Involved in Central Metabolism. Biochim. Biophys. Acta Mol. Cell Res. 2014, 1843, 372–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Wang, Y.; Tao, C.; Li, Y.; Cao, S.; Yang, X. CRNDE silencing promotes apoptosis and enhances cisplatin sensitivity of colorectal carcinoma cells by inhibiting the Akt/mTORC1-mediated Warburg effect. Oncol. Lett. 2022, 23, 70. [Google Scholar] [CrossRef] [PubMed]
- Bach, L.A. 40 YEARS OF IGF1: IGF-Binding Proteins. J. Mol. Endocrinol. 2018, 61, T11–T28. [Google Scholar] [CrossRef] [Green Version]
- Palmqvist, R.; Stattin, P.; Rinaldi, S.; Biessy, C.; Stenling, R.; Riboli, E.; Hallmans, G.; Kaaks, R. Plasma Insulin, IGF-Binding Proteins-1 and -2 and Risk of Colorectal Cancer: A Prospective Study in Northern Sweden. Int. J. Cancer 2003, 107, 89–93. [Google Scholar] [CrossRef]
- Jiang, B.; Zhang, X.; Du, L.-L.; Wang, Y.; Liu, D.-B.; Han, C.-Z.; Jing, J.-X.; Zhao, X.-W.; Xu, X.-Q. Possible Roles of Insulin, IGF-1 and IGFBPs in Initiation and Progression of Colorectal Cancer. World J. Gastroenterol. 2014, 20, 1608–1613. [Google Scholar] [CrossRef]
- Ollberding, N.J.; Cheng, I.; Wilkens, L.R.; Henderson, B.E.; Pollak, M.N.; Kolonel, L.N.; le Marchand, L. Genetic Variants, Prediagnostic Circulating Levels of Insulin-like Growth Factors, Insulin, and Glucose and the Risk of Colorectal Cancer: The Multiethnic Cohort Study. Cancer Epidemiol. Biomark. Prev. 2012, 21, 810–820. [Google Scholar] [CrossRef] [Green Version]
- Alami, N.; Page, V.; Yu, Q.; Jerome, L.; Paterson, J.; Shiry, L.; Leyland-Jones, B. Recombinant Human Insulin-like Growth Factor-Binding Protein 3 Inhibits Tumor Growth and Targets the Akt Pathway in Lung and Colon Cancer Models. Growth Horm. IGF Res. 2008, 18, 487–496. [Google Scholar] [CrossRef]
- Ali, O.; Cohen, P.; Lee, K.-W. Epidemiology and Biology of Insulin-like Growth Factor Binding Protein-3 (IGFBP-3) as an Anti-Cancer Molecule. Horm. Metab. Res. 2003, 35, 726–733. [Google Scholar] [CrossRef]
- Jenkins, P.J.; Khalaf, S.; Ogunkolade, W.; McCarthy, K.; David, T.; Hands, R.E.; Davies, D.; Bustin, S.A. Differential Expression of IGF-Binding Protein-3 in Normal and Malignant Colon and Its Influence on Apoptosis. Endocr. Relat. Cancer 2005, 12, 891–901. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, N.; Oshima, T.; Yoshihara, K.; Aoyama, T.; Hayashi, T.; Yamada, T.; Sato, T.; Shiozawa, M.; Yoshikawa, T.; Morinaga, S.; et al. Clinicopathological Significance and Impact on Outcomes of the Gene Expression Levels of IGF-1, IGF-2 and IGF-1R, IGFBP-3 in Patients with Colorectal Cancer: Overexpression of the IGFBP-3 Gene Is an Effective Predictor of Outcomes in Patients with Colorectal Cancer. Oncol. Lett. 2017, 13, 3958–3966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zámorová, M.; Holazová, A.; Miljuš, G.; Robajac, D.; Šunderić, M.; Malenković, V.; Đukanović, B.; Gemeiner, P.; Katrlík, J.; Nedić, O. Analysis of Changes in the Glycan Composition of Serum, Cytosol and Membrane Glycoprotein Biomarkers of Colorectal Cancer Using a Lectin-Based Protein Microarray. Anal. Methods 2017, 9, 2660–2666. [Google Scholar] [CrossRef]
- Vocka, M.; Langer, D.; Fryba, V.; Petrtyl, J.; Hanus, T.; Kalousova, M.; Zima, T.; Petruzelka, L. Novel Serum Markers HSP60, CHI3L1, and IGFBP-2 in Metastatic Colorectal Cancer. Oncol. Lett. 2019, 18, 6284–6292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šunderić, M.; Đukanović, B.; Malenković, V.; Nedić, O. Molecular Forms of the Insulin-like Growth Factor-Binding Protein-2 in Patients with Colorectal Cancer. Exp. Mol. Pathol. 2014, 96, 48–53. [Google Scholar] [CrossRef]
- Jenab, M.; Riboli, E.; Cleveland, R.J.; Norat, T.; Rinaldi, S.; Nieters, A.; Biessy, C.; Tjønneland, A.; Olsen, A.; Overvad, K.; et al. Serum C-Peptide, IGFBP-1 and IGFBP-2 and Risk of Colon and Rectal Cancers in the European Prospective Investigation into Cancer and Nutrition. Int. J. Cancer 2007, 121, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Otani, T.; Iwasaki, M.; Sasazuki, S.; Inoue, M.; Tsugane, S. Plasma C-Peptide, Insulin-like Growth Factor-I, Insulin-like Growth Factor Binding Proteins and Risk of Colorectal Cancer in a Nested Case-Control Study: The Japan Public Health Center-Based Prospective Study. Int. J. Cancer 2007, 120, 2007–2012. [Google Scholar] [CrossRef]
- Durai, R.; Davies, M.; Yang, W.; Yang, S.; Seifalian, A.; Goldspink, G.; Winslet, M. Biology of Insulin-like Growth Factor Binding Protein-4 and Its Role in Cancer (Review). Int. J. Oncol. 2006, 28, 1317–1325. [Google Scholar] [CrossRef]
- Street, M.; Miraki-Moud, F.; Sanderson, I.; Savage, M.; Giovannelli, G.; Bernasconi, S.; Camacho-Hubner, C. Interleukin-1beta (IL-1beta) and IL-6 Modulate Insulin-like Growth Factor-Binding Protein (IGFBP) Secretion in Colon Cancer Epithelial (Caco-2) Cells. J. Endocrinol. 2003, 179, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Bach, L.A. Recent Insights into the Actions of IGFBP-6. J. Cell Commun. Signal. 2015, 9, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zhu, X.; Wang, G.; Wang, W.; Ju, S.; Wang, X. Decreased Expression of IGFBP6 Correlates with Poor Survival in Colorectal Cancer Patients. Pathol. Res. Pract. 2020, 216, 152909. [Google Scholar] [CrossRef]
- Koyama, N.; Zhang, J.; Huqun; Miyazawa, H.; Tanaka, T.; Su, X.; Hagiwara, K. Identification of IGFBP-6 as an Effector of the Tumor Suppressor Activity of SEMA3B. Oncogene 2008, 27, 6581–6589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Dozmorov, M.; Oh, Y. IGFBP-3/IGFBP-3 Receptor System as an Anti-Tumor and Anti-Metastatic Signaling in Cancer. Cells 2020, 9, 1261. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.C.; Smartt, H.; Zadeh, H.A.M.; MacFarlane, M.; Paraskeva, C.; Collard, T.J. Insulin-like Growth Factor Binding Protein 3 (IGFBP-3) Potentiates TRAIL-Induced Apoptosis of Human Colorectal Carcinoma Cells through Inhibition of NF-ΚB. Cell Death Differ. 2007, 14, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-W.; Ma, L.; Yan, X.; Liu, B.; Zhang, X.; Cohen, P. Rapid Apoptosis Induction by IGFBP-3 Involves an Insulin-like Growth Factor-Independent Nucleomitochondrial Translocation of RXRα/Nur77. J. Biol. Chem. 2005, 280, 16942–16948. [Google Scholar] [CrossRef] [Green Version]
- Williams, A.C.; Collard, T.J.; Perks, C.M.; Newcomb, P.; Moorghen, M.; Holly, J.M.; Paraskeva, C. Increased P53-Dependent Apoptosis by the Insulin-like Growth Factor Binding Protein IGFBP-3 in Human Colonic Adenoma-Derived Cells. Cancer Res. 2000, 60, 22–27. [Google Scholar]
- Lin, Y.-W.; Weng, X.-F.; Huang, B.-L.; Guo, H.-P.; Xu, Y.-W.; Peng, Y.-H. IGFBP-1 in Cancer: Expression, Molecular Mechanisms, and Potential Clinical Implications. Am. J. Transl. Res. 2021, 13, 813–832. [Google Scholar]
- Kaaks, R.; Toniolo, P.; Akhmedkhanov, A.; Lukanova, A.; Biessy, C.; Dechaud, H.; Rinaldi, S.; Zeleniuch-Jacquotte, A.; Shore, R.E.; Riboli, E. Serum C-Peptide, Insulin-Like Growth Factor (IGF)-I, IGF-Binding Proteins, and Colorectal Cancer Risk in Women. J. Natl. Cancer Inst. 2000, 92, 1592–1600. [Google Scholar] [CrossRef] [PubMed]
- Pickard, A.; McCance, D.J. IGF-Binding Protein 2—Oncogene or Tumor Suppressor? Front. Endocrinol. 2015, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach, L.A.; Fu, P.; Yang, Z. Insulin-like Growth Factor-Binding Protein-6 and Cancer. Clin. Sci. 2013, 124, 215–229. [Google Scholar] [CrossRef] [Green Version]
- Fu, P.; Thompson, J.A.; Bach, L.A. Promotion of Cancer Cell Migration. J. Biol. Chem. 2007, 282, 22298–22306. [Google Scholar] [CrossRef] [Green Version]
- Devin, J.L.; Bolam, K.A.; Jenkins, D.G.; Skinner, T.L. The Influence of Exercise on the Insulin-like Growth Factor Axis in Oncology: Physiological Basis, Current, and Future Perspectives. Cancer Epidemiol. Biomark. Prev. 2016, 25, 239–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durai, R.; Yang, W.; Gupta, S.; Seifalian, A.M.; Winslet, M.C. The Role of the Insulin-like Growth Factor System in Colorectal Cancer: Review of Current Knowledge. Int. J. Colorectal Dis. 2005, 20, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Giovannucci, E. Insulin, Insulin-Like Growth Factors and Colon Cancer: A Review of the Evidence. J. Nutr. 2001, 131, 3109S–3120S. [Google Scholar] [CrossRef] [Green Version]
- Berk, Ş.; Janssen, J.A.M.J.L.; van Koetsveld, P.M.; Dogan, F.; Değerli, N.; Özcan, S.; Kelestimur, F.; Hofland, L.J. Modifying Effects of Glucose and Insulin/Insulin-Like Growth Factors on Colon Cancer Cells. Front. Oncol. 2021, 11, 645732. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Shen, F.; Weinfeld, M.; Sergi, C. Insulin Growth Factor Binding Protein 7 (IGFBP7)-Related Cancer and IGFBP3 and IGFBP7 Crosstalk. Front. Oncol. 2020, 10, 727. [Google Scholar] [CrossRef]
- Georges, R.B.; Adwan, H.; Hamdi, H.; Hielscher, T.; Linnemann, U.; Berger, M.R. The Insulin-like Growth Factor Binding Proteins 3 and 7 Are Associated with Colorectal Cancer and Liver Metastasis. Cancer Biol. Ther. 2011, 12, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.X.; Alfonso, H.; Chubb, S.A.P.; Ho, K.K.Y.; Fegan, P.G.; Hankey, G.J.; Golledge, J.; Flicker, L.; Yeap, B.B. Higher IGFBP3 is associated with increased incidence of colorectal cancer in older men independently of IGF1. Clin. Endocrinol. 2018, 88, 333–340. [Google Scholar] [CrossRef]
- Diehl, D.; Hessel, E.; Oesterle, D.; Renner-Müller, I.; Elmlinger, M.; Langhammer, M.; Göttlicher, M.; Wolf, E.; Lahm, H.; Hoeflich, A. IGFBP-2 overexpression reduces the appearance of dysplastic aberrant crypt foci and inhibits growth of adenomas in chemically induced colorectal carcinogenesis. Int. J. Cancer 2009, 124, 2220–2225. [Google Scholar] [CrossRef]
- Durai, R.; Yang, S.Y.; Seifalian, A.M.; Goldspink, G.; Winslet, M.C. Role of insulin-like growth factor binding protein-4 in prevention of colon cancer. World J. Surg. Oncol. 2007, 5, 128. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Miyamoto, S.; Maeda, H.; Ishii, G.; Hasebe, T.; Chiba, T.; Asaka, M.; Ochiai, A. Matrix Metalloproteinase-7 Degrades All Insulin-like Growth Factor Binding Proteins and Facilitates Insulin-like Growth Factor Bioavailability. Biochem. Biophys. Res. Commun. 2005, 333, 1011–1016. [Google Scholar] [CrossRef]
- Ohashi, S.; Natsuizaka, M.; Nakagawa, H. MMP7 and Activation of IGF-1R: A New Insight into Anti-EGFR Therapeutic Resistance in Metastatic Colorectal Cancer. Cancer Biol. Ther. 2011, 11, 184–187. [Google Scholar] [CrossRef] [Green Version]
- Sadowski, T.; Dietrich, S.; Koschinsky, F.; Sedlacek, R. Matrix Metalloproteinase 19 Regulates Insulin-like Growth Factor-Mediated Proliferation, Migration, and Adhesion in Human Keratinocytes through Proteolysis of Insulin-like Growth Factor Binding Protein-3. Mol. Biol. Cell 2003, 14, 4569–4580. [Google Scholar] [CrossRef] [PubMed]
- Walkiewicz, K.; Nowakowska-Zajdel, E.; Kozieł, P.; Muc-Wierzgoń, M. The Role of Some ADAM-Proteins and Activation of the Insulin Growth Factor-Related Pathway in Colorectal Cancer. Cent. Eur. J. Immunol. 2018, 43, 109–113. [Google Scholar] [CrossRef]
- Firth, S.M.; Baxter, R.C. Cellular Actions of the Insulin-Like Growth Factor Binding Proteins. Endocr. Rev. 2002, 23, 824–854. [Google Scholar] [CrossRef] [PubMed]
- Forbes, B.E.; McCarthy, P.; Norton, R.S. Insulin-Like Growth Factor Binding Proteins: A Structural Perspective. Front. Endocrinol. 2012, 3, 38. [Google Scholar] [CrossRef] [Green Version]
- Philippou, A.; Christopoulos, P.F.; Koutsilieris, M. Clinical Studies in Humans Targeting the Various Components of the IGF System Show Lack of Efficacy in the Treatment of Cancer. Mutat. Res. Rev. Mutat. Res. 2017, 772, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Chang, H.; Greer, A.; Hillerman, S.; Reeves, K.A.; Hurlburt, W.; Cogswell, J.; Patel, D.; Qi, Z.; Fairchild, C.; et al. IRS2 Copy Number Gain, KRAS and BRAF Mutation Status as Predictive Biomarkers for Response to the IGF-1R/IR Inhibitor BMS-754807 in Colorectal Cancer Cell Lines. Mol. Cancer Ther. 2015, 14, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wei, F.; Lv, G.; Li, C.; Liu, T.; Hadjipanayis, C.G.; Zhang, G.; Hao, C.; Bellail, A.C. The Association of TP53 Mutations with the Resistance of Colorectal Carcinoma to the Insulin-like Growth Factor-1 Receptor Inhibitor Picropodophyllin. BMC Cancer 2013, 13, 521. [Google Scholar] [CrossRef] [Green Version]
- Voutsadakis, I.A. The Landscape of PIK3CA Mutations in Colorectal Cancer. Clin. Colorectal Cancer 2021, 20, 201–215. [Google Scholar] [CrossRef]
- Lin, P.-C.; Lin, J.-K.; Lin, H.-H.; Lan, Y.-T.; Lin, C.-C.; Yang, S.-H.; Chen, W.-S.; Liang, W.-Y.; Jiang, J.-K.; Chang, S.-C. A Comprehensive Analysis of Phosphatase and Tensin Homolog Deleted on Chromosome 10 (PTEN) Loss in Colorectal Cancer. World J. Surg. Oncol. 2015, 13, 186. [Google Scholar] [CrossRef] [Green Version]
- Wong, H.-L.; Delellis, K.; Probst-Hensch, N.; Koh, W.-P.; van den Berg, D.; Lee, H.-P.; Yu, M.C.; Ingles, S.A. A New Single Nucleotide Polymorphism in the Insulin-like Growth Factor I Regulatory Region Associates with Colorectal Cancer Risk in Singapore Chinese. Cancer Epidemiol. Biomark. Prev. 2005, 14, 144–151. [Google Scholar] [CrossRef]
- Samowitz, W.S.; Wolff, R.K.; Ma, K.N.; Andersen, K.; Caan, B.; Slattery, M.L. Polymorphisms in Insulin-Related Genes Predispose to Specific KRAS2 and TP53 Mutations in Colon Cancer. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2006, 595, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Zecevic, M.; Amos, C.I.; Gu, X.; Campos, I.M.; Jones, J.S.; Lynch, P.M.; Rodriguez-Bigas, M.A.; Frazier, M.L. IGF1 Gene Polymorphism and Risk for Hereditary Nonpolyposis Colorectal Cancer. JNCI J. Natl. Cancer Inst. 2006, 98, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pechlivanis, S.; Wagner, K.; Chang-Claude, J.; Hoffmeister, M.; Brenner, H.; Försti, A. Polymorphisms in the Insulin like Growth Factor 1 and IGF Binding Protein 3 Genes and Risk of Colorectal Cancer. Cancer Detect. Prev. 2007, 31, 408–416. [Google Scholar] [CrossRef]
- Xiang, H.; Liu, L.; Chu, G.; Wei, S.; Liu, J.; Xu, Y.; Chen, J.; Wang, C.; Wei, Q.-Y.; Nie, S. Association Between Two Functional Polymorphisms of Insulin-Like Growth Factor Binding Protein 3 and Colorectal Cancer Risk in a Chinese Population. J. Toxicol. Environ. Health Part A 2009, 72, 706–711. [Google Scholar] [CrossRef]
- Feik, E.; Baierl, A.; Hieger, B.; Führlinger, G.; Pentz, A.; Stättner, S.; Weiss, W.; Pulgram, T.; Leeb, G.; Mach, K.; et al. Association of IGF1 and IGFBP3 Polymorphisms with Colorectal Polyps and Colorectal Cancer Risk. Cancer Causes Control 2010, 21, 91–97. [Google Scholar] [CrossRef]
- LeRoy, E.C.; Moore, J.H.; Hu, C.; Martínez, M.E.; Lance, P.; Duggan, D.; Thompson, P.A. Genes in the Insulin and Insulin-like Growth Factor Pathway and Odds of Metachronous Colorectal Neoplasia. Hum. Genet. 2011, 129, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Keku, T.O.; Vidal, A.; Oliver, S.; Hoyo, C.; Hall, I.J.; Omofoye, O.; McDoom, M.; Worley, K.; Galanko, J.; Sandler, R.S.; et al. Genetic Variants in IGF-I, IGF-II, IGFBP-3, and Adiponectin Genes and Colon Cancer Risk in African Americans and Whites. Cancer Causes Control 2012, 23, 1127–1138. [Google Scholar] [CrossRef] [Green Version]
- Karimi, K.; Mahmoudi, T.; Karimi, N.; Dolatmoradi, H.; Arkani, M.; Farahani, H.; Vahedi, M.; Parsimehr, E.; Dabiri, R.; Nobakht, H.; et al. Is There an Association between Variants in Candidate Insulin Pathway Genes IGF-I, IGFBP-3, INSR, and IRS2 and Risk of Colorectal Cancer in the Iranian Population? Asian Pac. J. Cancer Prev. 2013, 14, 5011–5016. [Google Scholar] [CrossRef] [Green Version]
- Stanilov, N.S.; Karakolev, I.A.; Deliysky, T.S.; Jovchev, J.P.; Stanilova, S.A. Association of Insulin-like Growth Factor-I Receptor Polymorphism with Colorectal Cancer Development. Mol. Biol. Rep. 2014, 41, 8099–8106. [Google Scholar] [CrossRef]
- Simons, C.C.J.M.; Schouten, L.J.; Godschalk, R.W.L.; van Engeland, M.; van den Brandt, P.A.; van Schooten, F.J.; Weijenberg, M.P. Genetic Variants in the Insulin-like Growth Factor Pathway and Colorectal Cancer Risk in the Netherlands Cohort Study. Sci. Rep. 2015, 5, 14126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoudi, T.; Majidzadeh-A, K.; Karimi, K.; Farahani, H.; Dabiri, R.; Nobakht, H.; Asadi, A.; Karimi, N.; Arkani, M.; Zali, M.R. Gly972Arg Variant of Insulin Receptor Substrate 1 Gene and Colorectal Cancer Risk in Overweight/Obese Subjects. Int. J. Biol. Markers 2016, 31, 68–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhou, X.; Liu, R.; Cao, Y.; Wang, L.; Chao, X.; Zhang, S. Relationship of SNP Rs35767 in IGF-1 Promoter Region with Susceptibility to Colorectal Cancer. Int. J. Clin. Exp. Pathol. 2018, 11, 5110–5116. [Google Scholar] [PubMed]
- Chao, X.L.; Wang, L.L.; Liu, R.; Li, Y.; Zhou, X.J. Association between CA Repeat Polymorphism in IGF1 Gene Promoter and Colorectal Cancer Risk in a Native Chinese Population. Neoplasma 2019, 66, 1002–1008. [Google Scholar] [CrossRef]
- Dhifallah, H.; Aissi, S.; Njima, M.; Zakhama, A.; Kenani, A. IGF1 Polymorphisms and Colon Cancer Risk in Tunisian Population. La Tunis. Med. 2020, 98, 60–69. [Google Scholar]
- Kattan, S.W.; Allah, A.M.K.A.; Mohamed, K.I.; Alruwetei, A.M.; Hegazy, A.H.; el Gayed, E.M.A. Linking Insulin like Growth Factor-1 (IGF-1) Rs6214 Gene Polymorphism and Its Serum Level with Risk of Colorectal Cancer. Beni-Suef Univ. J. Basic Appl. Sci. 2022, 11, 74. [Google Scholar] [CrossRef]
- Li, X.; Yang, J.; Wang, X.; Gao, X. The Association of IGF1 Rs35767 Polymorphism with Colorectal Cancer Risk in the Chinese Han Population. Nucleosides Nucleotides Nucleic Acids 2022, 1–9. [Google Scholar] [CrossRef]
- Kawasaki, T.; Nosho, K.; Ohnishi, M.; Suemoto, Y.; Kirkner, G.J.; Fuchs, C.S.; Ogino, S. IGFBP3 Promoter Methylation in Colorectal Cancer: Relationship with Microsatellite Instability, CpG Island Methylator Phenotype, P53. Neoplasia 2007, 9, 1091–1098. [Google Scholar] [CrossRef] [Green Version]
- Perez-Carbonell, L.; Balaguer, F.; Toiyama, Y.; Egoavil, C.; Rojas, E.; Guarinos, C.; Andreu, M.; Llor, X.; Castells, A.; Jover, R.; et al. IGFBP3 Methylation Is a Novel Diagnostic and Predictive Biomarker in Colorectal Cancer. PLoS ONE 2014, 9, e104285. [Google Scholar] [CrossRef]
- Goel, A.; Arnold, C.N.; Niedzwiecki, D.; Carethers, J.M.; Dowell, J.M.; Wasserman, L.; Compton, C.; Mayer, R.J.; Bertagnolli, M.M.; Boland, C.R. Frequent Inactivation of PTEN by Promoter Hypermethylation in Microsatellite Instability-High Sporadic Colorectal Cancers. Cancer Res. 2004, 64, 3014–3021. [Google Scholar] [CrossRef] [Green Version]
- Baharudin, R.; Ishak, M.; Muhamad Yusof, A.; Saidin, S.; Syafruddin, S.E.; Wan Mohamad Nazarie, W.F.; Lee, L.-H.; Ab Mutalib, N.-S. Epigenome-Wide DNA Methylation Profiling in Colorectal Cancer and Normal Adjacent Colon Using Infinium Human Methylation 450K. Diagnostics 2022, 12, 198. [Google Scholar] [CrossRef] [PubMed]
- Barlow, D.P.; Stöger, R.; Herrmann, B.G.; Saito, K.; Schweifer, N. The Mouse Insulin-like Growth Factor Type-2 Receptor Is Imprinted and Closely Linked to the Tme Locus. Nature 1991, 349, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Giannoukakis, N.; Deal, C.; Paquette, J.; Goodyer, C.G.; Polychronakos, C. Parental Genomic Imprinting of the Human IGF2 Gene. Nat. Genet. 1993, 4, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Takano, Y.; Shiota, G.; Kawasaki, H. Analysis of Genomic Imprinting of Insulin-Like Growth Factor 2 in Colorectal Cancer. Oncology 2000, 59, 210–216. [Google Scholar] [CrossRef]
- Cui, H.; Cruz-Correa, M.; Giardiello, F.M.; Hutcheon, D.F.; Kafonek, D.R.; Brandenburg, S.; Wu, Y.; He, X.; Powe, N.R.; Feinberg, A.P. Loss of IGF2 Imprinting: A Potential Marker of Colorectal Cancer Risk. Science 2003, 299, 1753–1755. [Google Scholar] [CrossRef]
- Tian, F.; Tang, Z.; Song, G.; Pan, Y.; He, B.; Bao, Q.; Wang, S. Loss of Imprinting of IGF2 Correlates with Hypomethylation of the H19 Differentially Methylated Region in the Tumor Tissue of Colorectal Cancer Patients. Mol. Med. Rep. 2012, 5, 1536–1540. [Google Scholar] [CrossRef]
- Belharazem, D.; Magdeburg, J.; Berton, A.-K.; Beissbarth, L.; Sauer, C.; Sticht, C.; Marx, A.; Hofheinz, R.; Post, S.; Kienle, P.; et al. Carcinoma of the Colon and Rectum with Deregulation of Insulin-like Growth Factor 2 Signaling: Clinical and Molecular Implications. J. Gastroenterol. 2016, 51, 971–984. [Google Scholar] [CrossRef]
- Schwarzenbach, H. Biological and Clinical Relevance of H19 in Colorectal Cancer Patients. EBioMedicine 2016, 13, 9–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Qi, M.; Fei, X.; Wang, X.; Wang, K. LncRNA H19: A Novel Oncogene in Multiple Cancers. Int. J. Biol. Sci. 2021, 17, 3188–3208. [Google Scholar] [CrossRef]
- Yin, Y.; Yan, Z.-P.; Lu, N.-N.; Xu, Q.; He, J.; Qian, X.; Yu, J.; Guan, X.; Jiang, B.-H.; Liu, L.-Z. Downregulation of MiR-145 Associated with Cancer Progression and VEGF Transcriptional Activation by Targeting N-RAS and IRS1. Biochim. Biophys. Acta Gene Regul. Mech. 2013, 1829, 239–247. [Google Scholar] [CrossRef]
- Pekow, J.; Meckel, K.; Dougherty, U.; Butun, F.; Mustafi, R.; Lim, J.; Crofton, C.; Chen, X.; Joseph, L.; Bissonnette, M. Tumor Suppressors MiR-143 and MiR-145 and Predicted Target Proteins API5, ERK5, K-RAS, and IRS-1 Are Differentially Expressed in Proximal and Distal Colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G179–G187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Chen, J.; Gao, C.; Zhu, D.; Xu, X.; Wu, C.; Jiang, J. MicroRNA 497 Inhibits Tumor Growth through Targeting Insulin Receptor Substrate 1 in Colorectal Cancer. Oncol. Lett. 2017, 14, 6379–6386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, S.; Yu, C.; Zhang, G.; Shi, F.; Chen, Y.; Yang, J.; Wu, W.; Zhou, Y. Downregulation of MicroRNA 126 Is Inversely Correlated with Insulin Receptor Substrate 1 Protein Expression in Colorectal Cancer and Is Associated with Advanced Stages of Disease. Oncol. Lett. 2020, 20, 2411–2419. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.T.; Jiang, C.C.; Wang, G.P.; Li, Y.P.; Wang, C.Y.; Guo, X.Y.; Yang, R.H.; Feng, Y.; Wang, F.H.; Tseng, H.-Y.; et al. MicroRNA-497 Targets Insulin-like Growth Factor 1 Receptor and Has a Tumour Suppressive Role in Human Colorectal Cancer. Oncogene 2013, 32, 1910–1920. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Liang, H.; Yao, W.; Wang, N.; Zhang, S.; Yan, X.; Feng, H.; Pang, W.; Wang, Y.; Wang, X.; et al. MiR-143 and MiR-145 Regulate IGF1R to Suppress Cell Proliferation in Colorectal Cancer. PLoS ONE 2014, 9, e114420. [Google Scholar] [CrossRef]
- Wu, G.; Liu, J.; Wu, Z.; Wu, X.; Yao, X. MicroRNA-184 Inhibits Cell Proliferation and Metastasis in Human Colorectal Cancer by Directly Targeting IGF-1R. Oncol. Lett. 2017, 14, 3215–3222. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Pan, W.; Shen, Y.; Chen, Z.; Zhang, L.; Zhang, Y.; Luo, Q.; Ying, X. IGF1/IGF1R and MicroRNA Let-7e down-Regulate Each Other and Modulate Proliferation and Migration of Colorectal Cancer Cells. Cell Cycle 2018, 17, 1212–1219. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhou, Y.; Zhou, Y.; Wang, J.; Ji, R. Mechanism of MiR 98 Inhibiting Tumor Proliferation and Invasion by Targeting IGF1R in Diabetic Patients Combined with Colon Cancer. Oncol. Lett. 2020, 20, 1719–1726. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, Q.; Li, M.; Jiang, S.; Wang, X. MicroRNA-375 Inhibits Colorectal Cancer Growth by Targeting PIK3CA. Biochem. Biophys. Res. Commun. 2014, 444, 199–204. [Google Scholar] [CrossRef]
- Ke, T.-W.; Wei, P.-L.; Yeh, K.-T.; Chen, W.T.-L.; Cheng, Y.-W. MiR-92a Promotes Cell Metastasis of Colorectal Cancer Through PTEN-Mediated PI3K/AKT Pathway. Ann. Surg. Oncol. 2015, 22, 2649–2655. [Google Scholar] [CrossRef]
- Wu, Y.; Song, Y.; Xiong, Y.; Wang, X.; Xu, K.; Han, B.; Bai, Y.; Li, L.; Zhang, Y.; Zhou, L. MicroRNA-21 (Mir-21) Promotes Cell Growth and Invasion by Repressing Tumor Suppressor PTEN in Colorectal Cancer. Cell. Physiol. Biochem. 2017, 43, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Coronel-Hernández, J.; López-Urrutia, E.; Contreras-Romero, C.; Delgado-Waldo, I.; Figueroa-González, G.; Campos-Parra, A.D.; Salgado-García, R.; Martínez-Gutierrez, A.; Rodríguez-Morales, M.; Jacobo-Herrera, N.; et al. Cell Migration and Proliferation Are Regulated by MiR-26a in Colorectal Cancer via the PTEN–AKT Axis. Cancer Cell Int. 2019, 19, 80. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Guo, X.; Zhang, H.; Xiang, Y.; Chen, J.; Yin, Y.; Cai, X.; Wang, K.; Wang, G.; Ba, Y.; et al. Role of MiR-143 Targeting KRAS in Colorectal Tumorigenesis. Oncogene 2009, 28, 1385–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Liu, J.; Huo, J.; Li, K.; Li, K.; Guo, H.; Yang, Y. Downregulation of MiR 143 Modulates KRAS Expression in Colorectal Carcinoma Cells. Oncol. Rep. 2019, 42, 2759–2767. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Pan, X.; Li, Z.; Bai, P.; Jin, H.; Wang, T.; Song, C.; Zhang, L.; Gao, L. Association between Polymorphisms in the Promoter Region of MiR-143/145 and Risk of Colorectal Cancer. Hum. Immunol. 2013, 74, 993–997. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, R.; Li, L.; Yuan, F.; Liang, W.; Wang, L.; Nie, X.; Chen, P.; Zhang, L.; Gao, L. A Functional Polymorphism in the Promoter of MiR-143/145 Is Associated with the Risk of Cervical Squamous Cell Carcinoma in Chinese Women. Medicine 2015, 94, e1289. [Google Scholar] [CrossRef]
- Chu, H.; Zhong, D.; Tang, J.; Li, J.; Xue, Y.; Tong, N.; Qin, C.; Yin, C.; Zhang, Z.; Wang, M. A Functional Variant in MiR-143 Promoter Contributes to Prostate Cancer Risk. Arch. Toxicol. 2016, 90, 403–414. [Google Scholar] [CrossRef]
- Sun, R.; Chen, P.; Li, L.; Sun, H.; Nie, X.; Liang, Y.; Yuan, F.; Pu, Y.; Bai, P.; Zhang, L.; et al. A Polymorphism Rs4705341 in the Flanking Region of MiR-143/145 Predicts Risk and Prognosis of Colorectal Cancer. Oncotarget 2016, 7, 62084–62090. [Google Scholar] [CrossRef] [Green Version]
- Yuan, F.; Sun, R.; Li, L.; Jin, B.; Wang, Y.; Liang, Y.; Che, G.; Gao, L.; Zhang, L. A Functional Variant Rs353292 in the Flanking Region of MiR-143/145 Contributes to the Risk of Colorectal Cancer. Sci. Rep. 2016, 6, 30195. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Huang, Q.; Meng, D.; Huang, M.; Li, C.; Qin, T. A Functional Rs353293 Polymorphism in the Promoter of MiR-143/145 Is Associated with a Reduced Risk of Bladder Cancer. PLoS ONE 2016, 11, e0159115. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhang, Y.; Li, Y.; Ma, Q.; Liu, S.; Yao, Y.; Tan, F.; Shi, L.; Yao, Y. The Association of Polymorphisms in MiRNAs with Nonsmall Cell Lung Cancer in a Han Chinese Population. Cancer Manag. Res. 2018, 10, 697–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradshaw, G.; Haupt, L.M.; Aquino, E.M.; Lea, R.A.; Sutherland, H.G.; Griffiths, L.R. Single Nucleotide Polymorphisms in MIR143 Contribute to Protection Against Non-Hodgkin Lymphoma (NHL) in Caucasian Populations. Genes 2019, 10, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotarac, N.; Dobrijevic, Z.; Matijasevic, S.; Savic-Pavicevic, D.; Brajuskovic, G. Analysis of Association of Potentially Functional Genetic Variants within Genes Encoding MiR-34b/c, MiR-378 and MiR-143/145 with Prostate Cancer in Serbian Population. EXCLI J. 2019, 18, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, X.; Quan, X.; Li, H.; Hao, X.; Jiang, M.; Zhou, B. Association between Two Polymorphisms in the Promoter Region of MiR-143/MiR-145 and the Susceptibility of Lung Cancer in Northeast Chinese Nonsmoking Females. DNA Cell Biol. 2019, 38, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Pan, J.; Zhang, L.; Wei, Y.; Wang, C. Long Non-Coding RNA CRNDE Sponges MiR-384 to Promote Proliferation and Metastasis of Pancreatic Cancer Cells through Upregulating IRS1. Cell Prolif. 2017, 50, e12389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Sha, H.; Sun, X.; Zhang, Y.; Wu, Y.; Zhang, J.; Zhang, H.; Wu, J.; Feng, J. CRNDE: An Oncogenic Long Non-Coding RNA in Cancers. Cancer Cell Int. 2020, 20, 162. [Google Scholar] [CrossRef]
- Poliseno, L.; Salmena, L.; Zhang, J.; Carver, B.; Haveman, W.J.; Pandolfi, P.P. A Coding-Independent Function of Gene and Pseudogene MRNAs Regulates Tumour Biology. Nature 2010, 465, 1033–1038. [Google Scholar] [CrossRef] [Green Version]
- Shang, A.-Q.; Wang, W.-W.; Yang, Y.-B.; Gu, C.-Z.; Ji, P.; Chen, C.; Zeng, B.-J.; Wu, J.-L.; Lu, W.-Y.; Sun, Z.-J.; et al. Knockdown of Long Noncoding RNA PVT1 Suppresses Cell Proliferation and Invasion of Colorectal Cancer via Upregulation of MicroRNA-214-3p. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G222–G232. [Google Scholar] [CrossRef]
- Mancarella, C.; Scotlandi, K. IGF2BP3 from Physiology to Cancer: Novel Discoveries, Unsolved Issues, and Future Perspectives. Front. Cell Dev. Biol. 2020, 7, 363. [Google Scholar] [CrossRef]
- Cao, J.; Mu, Q.; Huang, H. The Roles of Insulin-Like Growth Factor 2 MRNA-Binding Protein 2 in Cancer and Cancer Stem Cells. Stem Cells Int. 2018, 2018, 4217259. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-M.; Lin, C.-C.; Chen, W.-S.; Jiang, J.-K.; Yang, S.-H.; Chang, S.-C.; Ho, C.-L.; Yang, C.-C.; Huang, S.-C.; Chao, Y.; et al. Insulin-Like Growth Factor 2 MRNA-Binding Protein 1 (IGF2BP1) Is a Prognostic Biomarker and Associated with Chemotherapy Responsiveness in Colorectal Cancer. Int. J. Mol. Sci. 2021, 22, 6940. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, A.M.; Serra, M.E.S.; Gonçalves, A.C.; Rio, J.; Oliveiros, B.; Laranjo, M.; Rocha-Gonsalves, A.M.; Sarmento-Ribeiro, A.B.; Botelho, M.F. Hypoxia-Induced Redox Alterations and Their Correlation with 99mTc-MIBI and 99mTc-HL-91 Uptake in Colon Cancer Cells. Nucl. Med. Biol. 2010, 37, 125–132. [Google Scholar] [CrossRef]
- Cao, D.; Hou, M.; Guan, Y.; Jiang, M.; Yang, Y.; Gou, H. Expression of HIF-1alpha and VEGF in Colorectal Cancer: Association with Clinical Outcomes and Prognostic Implications. BMC Cancer 2009, 9, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Xu, Z.; Mao, S.; Chen, W.; Zeng, R.; Zhou, S.; Liu, J. Effect of Hypoxia on Hypoxia Inducible Factor-1α, Insulin-like Growth Factor I and Vascular Endothelial Growth Factor Expression in Hepatocellular Carcinoma HepG2 Cells. Oncol. Lett. 2015, 9, 1142–1148. [Google Scholar] [CrossRef] [Green Version]
- Joung, Y.-H.; Lee, M.-Y.; Lim, E.-J.; Kim, M.-S.; Hwang, T.S.; Kim, S.-Y.; Ye, S.-K.; Lee, J.-D.; Park, T.; Woo, Y.-S.; et al. Hypoxia Activates the IGF-1 Expression through STAT5b in Human HepG2 Cells. Biochem. Biophys. Res. Commun. 2007, 358, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Loh, C.-Y.; Chai, J.; Tang, T.; Wong, W.; Sethi, G.; Shanmugam, M.; Chong, P.; Looi, C. The E-Cadherin and N-Cadherin Switch in Epithelial-to-Mesenchymal Transition: Signaling, Therapeutic Implications, and Challenges. Cells 2019, 8, 1118. [Google Scholar] [CrossRef] [Green Version]
- Canonici, A.; Steelant, W.; Rigot, V.; Khomitch-Baud, A.; Boutaghou-Cherid, H.; Bruyneel, E.; van Roy, F.; Garrouste, F.; Pommier, G.; André, F. Insulin-like Growth Factor-I Receptor, E-Cadherin and Av Integrin Form a Dynamic Complex under the Control of α-Catenin. Int. J. Cancer 2008, 122, 572–582. [Google Scholar] [CrossRef]
- André, F.; Janssens, B.; Bruyneel, E.; van Roy, F.; Gespach, C.; Mareel, M.; Bracke, M. Alpha-Catenin Is Required for IGF-I-Induced Cellular Migration but Not Invasion in Human Colonic Cancer Cells. Oncogene 2004, 23, 1177–1186. [Google Scholar] [CrossRef] [Green Version]
- Vella, V.; Malaguarnera, R.; Nicolosi, M.L.; Morrione, A.; Belfiore, A. Insulin/IGF Signaling and Discoidin Domain Receptors: An Emerging Functional Connection. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118522. [Google Scholar] [CrossRef]
- Malaguarnera, R.; Nicolosi, M.L.; Sacco, A.; Morcavallo, A.; Vella, V.; Voci, C.; Spatuzza, M.; Xu, S.-Q.; Iozzo, R.V.; Vigneri, R.; et al. Novel Cross-Talk between IGF-IR and DDR1 Regulates IGF-IR Trafficking, Signaling and Biological Responses. Oncotarget 2015, 6, 16084–16105. [Google Scholar] [CrossRef] [Green Version]
- ben Arfi, K.; Schneider, C.; Bennasroune, A.; Bouland, N.; Wolak-Thierry, A.; Collin, G.; Le, C.C.; Toussaint, K.; Hachet, C.; Lehrter, V.; et al. Discoidin Domain Receptor 1 Expression in Colon Cancer: Roles and Prognosis Impact. Cancers 2022, 14, 928. [Google Scholar] [CrossRef] [PubMed]
- Reszegi, A.; Horváth, Z.; Karászi, K.; Regős, E.; Postniková, V.; Tátrai, P.; Kiss, A.; Schaff, Z.; Kovalszky, I.; Baghy, K. The Protective Role of Decorin in Hepatic Metastasis of Colorectal Carcinoma. Biomolecules 2020, 10, 1199. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Buraschi, S.; Genua, M.; Xu, S.-Q.; Solomides, C.C.; Peiper, S.C.; Gomella, L.G.; Owens, R.C.; Morrione, A. Decorin Antagonizes IGF Receptor I (IGF-IR) Function by Interfering with IGF-IR Activity and Attenuating Downstream Signaling. J. Biol. Chem. 2011, 286, 34712–34721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrione, A.; Neill, T.; Iozzo, R.V. Dichotomy of Decorin Activity on the Insulin-like Growth Factor-I System. FEBS J. 2013, 280, 2138–2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, L.; Yang, J.; Yue, J.; Chen, Y.; Zhou, H.; Fan, D.; Zhang, Q.; Buraschi, S.; Iozzo, R.V.; Bi, X. Decorin Deficiency Promotes Epithelial-Mesenchymal Transition and Colon Cancer Metastasis. Matrix Biol. 2021, 95, 1–14. [Google Scholar] [CrossRef] [PubMed]
- de Jong, G.; van Dijk, J.P.; van Eijk, H.G. The Biology of Transferrin. Clin. Chim. Acta 1990, 190, 1–46. [Google Scholar] [CrossRef]
- Miljuš, G.; Malenković, V.; Đukanović, B.; Kolundžić, N.; Nedić, O. IGFBP-3/Transferrin/Transferrin Receptor 1 Complexes as Principal Mediators of IGFBP-3 Delivery to Colon Cells in Non-Cancer and Cancer Tissues. Exp. Mol. Pathol. 2015, 98, 431–438. [Google Scholar] [CrossRef]
- Westwood, M.; Aplin, J.D.; Collinge, I.A.; Gill, A.; White, A.; Gibson, J.M. A2-Macroglobulin: A New Component in the Insulin-like Growth Factor/Insulin-like Growth Factor Binding Protein-1 Axis. J. Biol. Chem. 2001, 276, 41668–41674. [Google Scholar] [CrossRef] [Green Version]
- Šunderić, M.; Malenković, V.; Nedić, O. Complexes between Insulin-like Growth Factor Binding Proteins and Alpha-2-Macroglobulin in Patients with Tumor. Exp. Mol. Pathol. 2015, 98, 173–177. [Google Scholar] [CrossRef]
- Gligorijević, N.; Penezić, A.; Nedić, O. Influence of Glyco-Oxidation on Complexes between Fibrin (Ogen) and Insulin-like Growth Factor-Binding Protein-1 in Patients with Diabetes Mellitus Type 2. Free Radic. Res. 2017, 51, 64–72. [Google Scholar] [CrossRef]
- Orrù, S.; Nigro, E.; Mandola, A.; Alfieri, A.; Buono, P.; Daniele, A.; Mancini, A.; Imperlini, E. A Functional Interplay between IGF-1 and Adiponectin. Int. J. Mol. Sci. 2017, 18, 2145. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, M.J.; Cooke, A.; Rosenfeld-Franklin, M.; Buck, E.; Foreman, K.; Landfair, D.; O’Connor, M.; Pirritt, C.; Sun, Y.; Yao, Y.; et al. Discovery of OSI-906: A Selective and Orally Efficacious Dual Inhibitor of the IGF-1 Receptor and Insulin Receptor. Future Med. Chem. 2009, 1, 1153–1171. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Mehrabi Nasab, E.; Hassanpour, F.; Athari, S.S. Linsitinib and Aspirin as the IGF1-R Antagonists, Inhibit Regorafenib-Resistant Chemotherapy in Colon Cancer. Saudi J. Biol. Sci. 2022, 29, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Leiphrakpam, P.D.; Agarwal, E.; Mathiesen, M.; Haferbier, K.L.; Brattain, M.G.; Chowdhury, S. In Vivo Analysis of Insulin-like Growth Factor Type 1 Receptor Humanized Monoclonal Antibody MK-0646 and Small Molecule Kinase Inhibitor OSI-906 in Colorectal Cancer. Oncol. Rep. 2014, 31, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Reidy, D.L.; Vakiani, E.; Fakih, M.G.; Saif, M.W.; Hecht, J.R.; Goodman-Davis, N.; Hollywood, E.; Shia, J.; Schwartz, J.; Chandrawansa, K.; et al. Randomized, Phase II Study of the Insulin-Like Growth Factor-1 Receptor Inhibitor IMC-A12, With or Without Cetuximab, in Patients with Cetuximab- or Panitumumab-Refractory Metastatic Colorectal Cancer. J. Clin. Oncol. 2010, 28, 4240–4246. [Google Scholar] [CrossRef] [Green Version]
- Sclafani, F.; Kim, T.Y.; Cunningham, D.; Kim, T.W.; Tabernero, J.; Schmoll, H.J.; Roh, J.K.; Kim, S.Y.; Park, Y.S.; Guren, T.K.; et al. A Randomized Phase II/III Study of Dalotuzumab in Combination with Cetuximab and Irinotecan in Chemorefractory, KRAS Wild-Type, Metastatic Colorectal Cancer. J. Natl. Cancer Inst. 2015, 107, djv258. [Google Scholar] [CrossRef] [Green Version]
- Wilky, B.A.; Rudek, M.A.; Ahmed, S.; Laheru, D.A.; Cosgrove, D.; Donehower, R.C.; Nelkin, B.; Ball, D.; Doyle, L.A.; Chen, H.; et al. A Phase I Trial of Vertical Inhibition of IGF Signalling Using Cixutumumab, an Anti-IGF-1R Antibody, and Selumetinib, an MEK 1/2 Inhibitor, in Advanced Solid Tumours. Br. J. Cancer 2015, 112, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Tabernero, J.; Chawla, S.P.; Kindler, H.; Reckamp, K.; Chiorean, E.G.; Azad, N.S.; Lockhart, A.C.; Hsu, C.-P.; Baker, N.F.; Galimi, F.; et al. Anticancer Activity of the Type I Insulin-like Growth Factor Receptor Antagonist, Ganitumab, in Combination with the Death Receptor 5 Agonist, Conatumumab. Target. Oncol. 2015, 10, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Fazenbaker, C.; Chen, C.; Breen, S.; Huang, J.; Yao, X.; Ren, P.; Yao, Y.; Herbst, R.; Hollingsworth, R.E. Overproduction of IGF-2 Drives a Subset of Colorectal Cancer Cells, Which Specifically Respond to an Anti-IGF Therapeutic Antibody and Combination Therapies. Oncogene 2017, 36, 797–806. [Google Scholar] [CrossRef]
- Oh, Y.J.; Dent, M.W.; Freels, A.R.; Zhou, Q.; Lebrilla, C.B.; Merchant, M.L.; Matoba, N. Antitumor Activity of a Lectibody Targeting Cancer-Associated High-Mannose Glycans. Mol. Ther. 2022, 30, 1523–1535. [Google Scholar] [CrossRef]
- Zong, R.; Chen, X.; Feng, J.; Xu, S. IGF-1R Depletion Sensitizes Colon Cancer Cell Lines to Radiotherapy. Cancer Biomark. 2021, 32, 199–206. [Google Scholar] [CrossRef]
- Li, Y.; Lu, K.; Zhao, B.; Zeng, X.; Xu, S.; Ma, X.; Zhi, Y. Depletion of Insulin-like Growth Factor 1 Receptor Increases Radiosensitivity in Colorectal Cancer. J. Gastrointest. Oncol. 2020, 11, 1135–1145. [Google Scholar] [CrossRef] [PubMed]
- Fuentes-Baile, M.; Ventero, M.P.; Encinar, J.A.; García-Morales, P.; Poveda-Deltell, M.; Pérez-Valenciano, E.; Barberá, V.M.; Gallego-Plazas, J.; Rodríguez-Lescure, Á.; Martín-Nieto, J.; et al. Differential Effects of IGF-1R Small Molecule Tyrosine Kinase Inhibitors BMS-754807 and OSI-906 on Human Cancer Cell Lines. Cancers 2020, 12, 3717. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, Y.; Zhu, J.; Zheng, H.; Chen, S.; Chen, L.; Yang, H.-S. IGF-1R Inhibition Induces MEK Phosphorylation to Promote Survival in Colon Carcinomas. Signal Transduct. Target. Ther. 2020, 5, 153. [Google Scholar] [CrossRef]
- Sanderson, M.P.; Apgar, J.; Garin-Chesa, P.; Hofmann, M.H.; Kessler, D.; Quant, J.; Savchenko, A.; Schaaf, O.; Treu, M.; Tye, H.; et al. BI 885578, a Novel IGF1R/INSR Tyrosine Kinase Inhibitor with Pharmacokinetic Properties That Dissociate Antitumor Efficacy and Perturbation of Glucose Homeostasis. Mol. Cancer Ther. 2015, 14, 2762–2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Zhao, N.; Jiang, M. Isovitexin Attenuates Tumor Growth in Human Colon Cancer Cells through the Modulation of Apoptosis and Epithelial-Mesenchymal Transition via PI3K/Akt/MTOR Signaling Pathway. Biochem. Cell Biol. 2021, 99, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.J.; Hong, E.M.; Park, S.W.; Byun, H.W.; Koh, D.H.; Choi, M.H.; Kae, S.H.; Lee, J. Statin Induces Apoptosis of Human Colon Cancer Cells and Downregulation of Insulin-like Growth Factor 1 Receptor via Proapoptotic ERK Activation. Oncol. Lett. 2016, 12, 250–256. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-H.; Kim, I.-H.; Nam, T.-J. Phloroglucinol Induces Apoptosis through the Regulation of Insulin-like Growth Factor 1 Receptor Signaling Pathways in Human Colon Cancer HT-29 Cells. Int. J. Oncol. 2014, 45, 1036–1042. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, F.; Bai, Z.; Chi, B.; Wu, J.; Chen, X. Curcumol Inhibits Growth and Induces Apoptosis of Colorectal Cancer LoVo Cell Line via IGF-1R and P38 MAPK Pathway. Int. J. Mol. Sci. 2015, 16, 19851–19867. [Google Scholar] [CrossRef] [Green Version]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Regolo, L.; Alvarez-Suarez, J.M.; Quinzi, D.; Sargenti, A.; Bai, W.; Tian, L.; Giampieri, F.; Battino, M. Manuka Honey in Combination with 5-Fluorouracil Decreases Physical Parameters of Colonspheres Enriched with Cancer Stem-like Cells and Reduces Their Resistance to Apoptosis. Food Chem. 2022, 374, 131753. [Google Scholar] [CrossRef]
- Valadez-Bustos, N.; Escamilla-Silva, E.M.; García-Vázquez, F.J.; Gallegos-Corona, M.A.; Amaya-Llano, S.L.; Ramos-Gómez, M. Oral Administration of Microencapsulated B. longum BAA-999 and Lycopene Modulates IGF-1/IGF-1R/IGFBP3 Protein Expressions in a Colorectal Murine Model. Int. J. Mol. Sci. 2019, 20, 4275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Zheng, W.; Luo, Y.; Ou, X.; Song, L.; Zhang, S.; He, T.; Guo, Z.; Zhu, J.; Shi, H.; et al. Arca Subcrenata Polypeptides Inhibit Human Colorectal Cancer HT-29 Cells Growth via Suppression of IGF-1R/Akt/MTOR Signaling and ATP Production. Nutr. Cancer 2020, 72, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Kim, J.-S.; Seo, Y.R.; Park, J.H.Y.; Choi, M.-S.; Sung, M.-K. Carnosic Acid Suppresses Colon Tumor Formation in Association with Antiadipogenic Activity. Mol. Nutr. Food Res. 2014, 58, 2274–2285. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-H.; Nam, T.-J. Fucoidan Downregulates Insulin-like Growth Factor-I Receptor Levels in HT-29 Human Colon Cancer Cells. Oncol. Rep. 2018, 39, 1516–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.-K.; Kim, I.-H.; Kim, J.; Nam, T.-J. Induction of Apoptosis by Laminarin, Regulating the Insulin-like Growth Factor-IR Signaling Pathways in HT-29 Human Colon Cells. Int. J. Mol. Med. 2012, 30, 734–738. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, S.A.; Zand, H.; Cheraghpour, M. The Influence of Curcumin on the Downregulation of MYC, Insulin and IGF-1 Receptors: A Possible Mechanism Underlying the Anti-Growth and Anti-Migration in Chemoresistant Colorectal Cancer Cells. Medicina 2019, 55, 90. [Google Scholar] [CrossRef] [Green Version]
- Lim, D.Y.; Cho, H.J.; Kim, J.; Nho, C.W.; Lee, K.W.; Park, J.H.Y. Luteolin Decreases IGF-II Production and Downregulates Insulin-like Growth Factor-I Receptor Signaling in HT-29 Human Colon Cancer Cells. BMC Gastroenterol. 2012, 12, 9. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Teng, Y.; Liu, S.; Wang, Z.; Chen, Y.; Zhang, Y.; Xi, S.; Xu, S.; Wang, R.; Zou, X. Cinnamaldehyde Affects the Biological Behavior of Human Colorectal Cancer Cells and Induces Apoptosis via Inhibition of the PI3K/Akt Signaling Pathway. Oncol. Rep. 2016, 35, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Global bivariate maps representing the link between human development index (HDI) and colorectal cancer incidence (A) or mortality (B). The boundaries expressed on this map do not imply the expression of any opinion whatsoever concerning the legal status of any country, territory, city or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. Data source: WHO, UNDP [2,6].
Figure 1.
Global bivariate maps representing the link between human development index (HDI) and colorectal cancer incidence (A) or mortality (B). The boundaries expressed on this map do not imply the expression of any opinion whatsoever concerning the legal status of any country, territory, city or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. Data source: WHO, UNDP [2,6].

Figure 2.
Multiple-stage neoplastic transformation of the colon tissue.
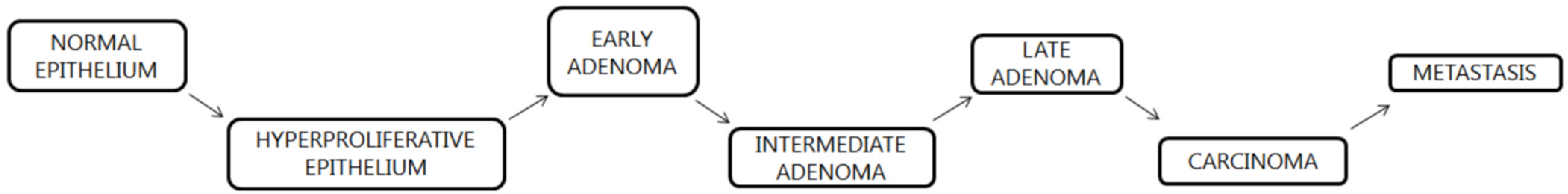
Figure 3.
Schematic presentation of IGF1R/IR-dependent cell signaling pathways and molecules altered in CRC. Upward and downward red arrows indicate a direction of change. IR—insulin receptor; IGF—insulin-like growth factor; IGF1R—insulin-like growth factor receptor; IGFBP—IGF binding protein; IRS—insulin receptor substrate; PI3K—phosphoinositide 3-kinase; Akt—protein kinase B; GSK3β—glycogen synthase kinase 3-β; Bcl-xL—B-cell lymphoma extra-large; mTOR—mammalian target of rapamycin; Shc—Src homology and collagen adaptor protein; Ras—rat sarcoma virus-related protein; Raf—serine/threonine-specific protein kinase; MEK—mitogen-activated protein kinase; ERK—extracellular signal-regulated kinase.
Figure 3.
Schematic presentation of IGF1R/IR-dependent cell signaling pathways and molecules altered in CRC. Upward and downward red arrows indicate a direction of change. IR—insulin receptor; IGF—insulin-like growth factor; IGF1R—insulin-like growth factor receptor; IGFBP—IGF binding protein; IRS—insulin receptor substrate; PI3K—phosphoinositide 3-kinase; Akt—protein kinase B; GSK3β—glycogen synthase kinase 3-β; Bcl-xL—B-cell lymphoma extra-large; mTOR—mammalian target of rapamycin; Shc—Src homology and collagen adaptor protein; Ras—rat sarcoma virus-related protein; Raf—serine/threonine-specific protein kinase; MEK—mitogen-activated protein kinase; ERK—extracellular signal-regulated kinase.
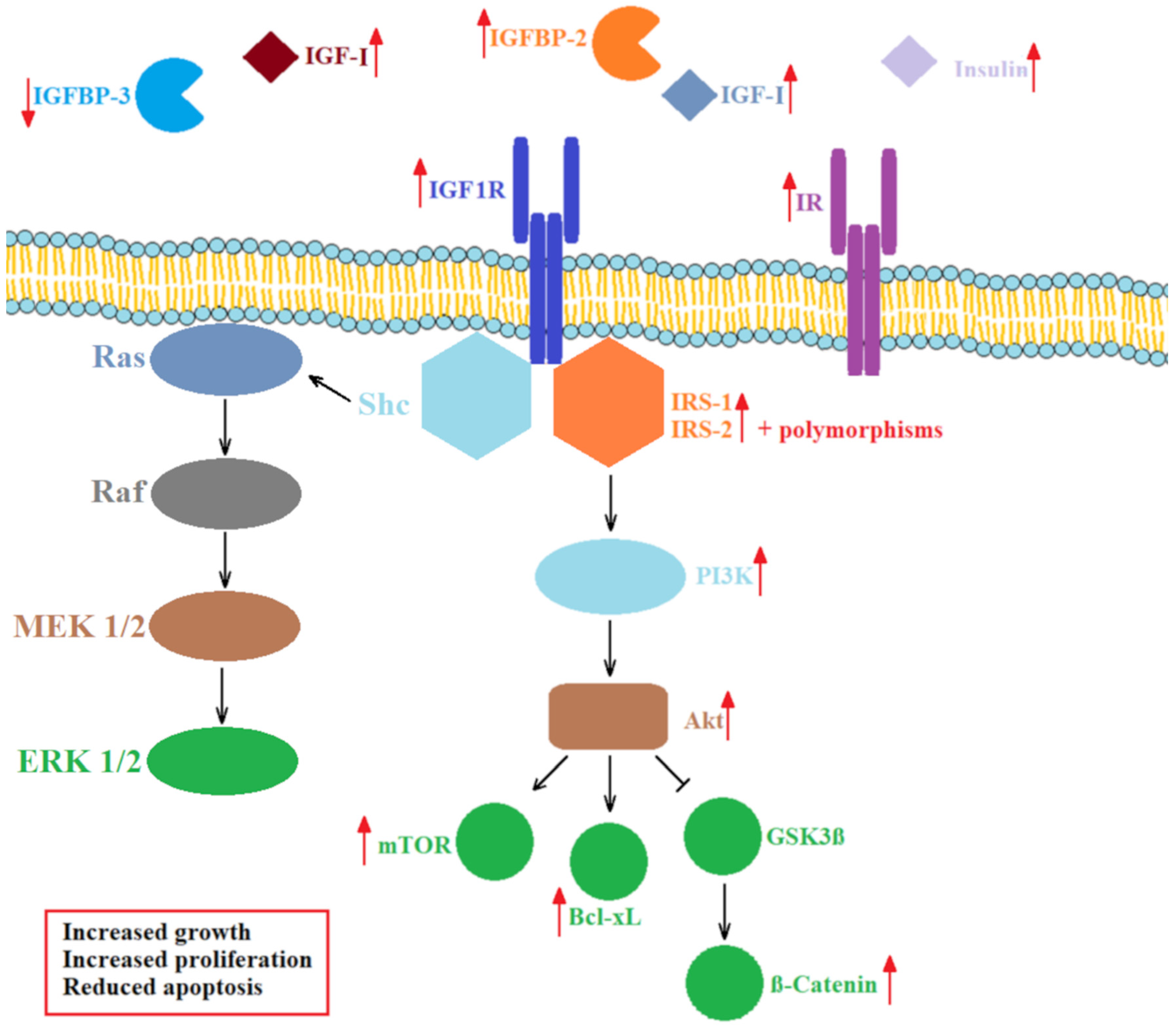

{kind=link}
{kind=link}
{kind=link}
Table 1.
Known or suspected roles of IGFBPs in CRC.
| IGFBP | Role | Possible Modes of Action | Reference |
|---|---|---|---|
| IGFBP-1 | Protective | Decreased concentration leads to an increased concentration of free IGF-I | [99] |
| IGFBP-2 | Tumor promoting | Local increase in free IGF-I due to proteolysis and binding to α5β1 integrin | [60,85,86] |
| IGFBP-3 | Protective | Binding of free IGF-I, pro-apoptotic role through NF-κB inhibition, interaction with retinoid X receptor (RXR)/Nur77 or p53-dependent signaling cascade | [79,80,95,96,97] |
| Tumor promoting | Undefined mechanisms dependent on other members of the IGF system, mainly the concentrations of IGF-I and IGF-II | [81,82] | |
| IGFBP-4 | Protective | Inhibits growth of colon cancer cells | [89,90] |
| IGFBP-5 | Not known | - | - |
| IGFBP-6 | Protective | Inhibition of IGF-II induced proliferation and migration, protection via IGF-independent pathways | [91,92,93,101] |
| Tumor promoting | Stimulation of LIM 1215 colon cancer cell migration | [102] | |
| IGFBP-7 | Possibly protective | Possibly associated with the activation of apoptosis, but further evidence is required | [108] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gligorijević, N.; Dobrijević, Z.; Šunderić, M.; Robajac, D.; Četić, D.; Penezić, A.; Miljuš, G.; Nedić, O. The Insulin-like Growth Factor System and Colorectal Cancer. Life 2022, 12, 1274. https://doi.org/10.3390/life12081274
AMA Style
Gligorijević N, Dobrijević Z, Šunderić M, Robajac D, Četić D, Penezić A, Miljuš G, Nedić O. The Insulin-like Growth Factor System and Colorectal Cancer. Life. 2022; 12(8):1274. https://doi.org/10.3390/life12081274
Chicago/Turabian StyleGligorijević, Nikola, Zorana Dobrijević, Miloš Šunderić, Dragana Robajac, Danilo Četić, Ana Penezić, Goran Miljuš, and Olgica Nedić. 2022. "The Insulin-like Growth Factor System and Colorectal Cancer" Life 12, no. 8: 1274. https://doi.org/10.3390/life12081274
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.