
The Cytotoxic Properties of Extreme Fungi’s Bioactive Components—An Updated Metabolic and Omics Overview
by

Attila Kiss
1,†, 
Farhad Hariri Akbari
2,†,
Andrey Marchev
3,
Viktor Papp
4 and
Iman Mirmazloum
5,*
1
Agro-Food Science Techtransfer and Innovation Centre, Faculty for Agro, Food and Environmental Science, Debrecen University, 4032 Debrecen, Hungary
2
Department of Biology, Biotechnical Faculty, University of Ljubljana, 1000 Ljubljana, Slovenia
3
Laboratory of Metabolomics, Department of Biotechnology, The Stephan Angeloff Institute of Microbiology, Bulgarian Academy of Sciences, 4000 Plovdiv, Bulgaria
4
Department of Botany, Hungarian University of Agriculture and Life Sciences, 1118 Budapest, Hungary
5
Department of Plant Physiology and Plant Ecology, Institute of Agronomy, Hungarian University of Agriculture and Life Sciences, 1118 Budapest, Hungary
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work as co-first authors.
Life 2023, 13(8), 1623; https://doi.org/10.3390/life13081623
Submission received: 5 June 2023
/
Revised: 28 June 2023
/
Accepted: 29 June 2023
/
Published: 25 July 2023
(This article belongs to the Collection Bacterial Infections, Treatment and Antibiotic Resistance)
Abstract
:Fungi are the most diverse living organisms on planet Earth, where their ubiquitous presence in various ecosystems offers vast potential for the research and discovery of new, naturally occurring medicinal products. Concerning human health, cancer remains one of the leading causes of mortality. While extensive research is being conducted on treatments and their efficacy in various stages of cancer, finding cytotoxic drugs that target tumor cells with no/less toxicity toward normal tissue is a significant challenge. In addition, traditional cancer treatments continue to suffer from chemical resistance. Fortunately, the cytotoxic properties of several natural products derived from various microorganisms, including fungi, are now well-established. The current review aims to extract and consolidate the findings of various scientific studies that identified fungi-derived bioactive metabolites with antitumor (anticancer) properties. The antitumor secondary metabolites identified from extremophilic and extremotolerant fungi are grouped according to their biological activity and type. It became evident that the significance of these compounds, with their medicinal properties and their potential application in cancer treatment, is tremendous. Furthermore, the utilization of omics tools, analysis, and genome mining technology to identify the novel metabolites for targeted treatments is discussed. Through this review, we tried to accentuate the invaluable importance of fungi grown in extreme environments and the necessity of innovative research in discovering naturally occurring bioactive compounds for the development of novel cancer treatments.
1. Introduction
Cancer is a polygenic, multifactorial, and complex disease comprising up to 300 different types that are associated with various environmental factors and genetic disorders since the dawn of mankind [1,2,3]. Documented data indicate that cancer is the second leading cause of death after cardiovascular disease [4]. An estimated 9.6 million deaths related to cancer since 2018 were reported by a study on the global burden of disease [3,5].
Depending on the disease conditions, there are various prescribed treatments for cancer, including radiotherapy, chemotherapy, targeted surgery and immunotherapy, recent gene therapies, or a combination of the above approaches [6,7,8].
There are usually conjugated elements in cancer therapies that deliver drugs to specific cancer cells or peripheral tissues that prevent tumour cells from growing [9]. So far, several cytotoxic compounds have been identified with chemotherapeutic properties, of which most are non-tumour-cell-specific and cause toxicity to healthy tissue [10]. Moreover, the chemo-resistance phenomenon is still a weakness of these treatments [11]. Despite numerous advances in cancer treatment, the resistance of mammalian tumour cells to chemotherapy and their numerous side effects necessitates the discovery and introduction of newer alternatives or synergistic drugs [12].
In contrast, natural products derived from various microorganisms with a variety of medicinal properties such as targeted cytotoxic, antitumor, anti-inflammatory, and other activities are continuously being discovered [13]. Such secondary metabolites, also referred to as “specialized metabolites” in the recent literature, are organic compounds that, unlike primary metabolites, are not directly involved in the growth or reproduction of a living organism [14,15,16]. It is believed that these compounds are being utilized to protect the organisms from various biotic and abiotic stress factors [17,18,19].
Despite great progress in the synthesis of synthetic substances, novel metabolites produced in plants/fungi with medicinal properties often have complex biosynthetic pathways with complicated structures, which makes it difficult to synthesize them artificially on an industrial scale [19,20].
As a result, the most common accessibility options for such metabolites are natural resources and targeted extraction and purification [21].
Fungi are known to be the most diverse and abundant organisms on our planet [22]. Several fungal species are able to grow/thrive in extreme environments [23]. The presence of fungi in all kinds of ecosystems provides excellent opportunities for the research and discovery of new natural products [24,25]. Fungi-derived compounds are widely used in medical, biotechnological, and traditional bioremediation applications [26,27,28,29]. They have also been the subject of several studies to obtain new biomolecules and enzymes, serving various purposes such as biofuels, insecticides, pesticides, anthelmintic, and analgesics. Additionally, fungi’s metabolites are utilized in flavourings, cosmetics, and as supplements in single-cell proteins (SCPs) [30,31,32].
The profound value and importance of these compounds in the mentioned industries is completely consistent with the evolving desire of our society towards natural treatments. Due to this growing trend, it has become necessary to perform an extensive chemical discovery of these metabolites, with the aim of increasing our understanding of their biosynthetic origin and unlocking their enormous potential for diverse biological activities [33,34]. In the field of fungal discoveries, penicillin produced by Penicillium notatum has been considered a goldmine since 1928 [35]. Ascomycetes such as Aspergillus, Penicillium, and Fuserium, as well as imperfect fungi, are common producers of bioactive compounds, followed by the endophytic [35,36,37] and filamentous species, and Alterneria, Trichoderma, and Phoma [38]. However, there are still several metabolites of these microorganisms that are yet to be characterized [39].
The term “extremus” means extreme in Latin and Greek, and the word “philiā” conveys the meaning of love [40,41]. The extremus organisms have existed for many years and continue to grow under extreme conditions, making them excellent choices to be considered for evolutionary studies [42,43]. According to the concept derived from these terms, these organisms can grow in harsh physical and geochemical conditions that are intolerable to most organisms [44]. They can proliferate in salty, acidic, and alkaline conditions and solutions; in hot and icy recesses; in toxic wastes, heavy metals, and organic solvents; or in other extreme habitats which, as mentioned earlier, are considered unfavorable for most organisms [45,46].
These organisms are typically classified into three groups: I: extremophilic organisms, which require severe conditions to reproduce [46]; II: extremotolerant organisms, which grow optimally under “normal” conditions but can tolerate extreme amounts of one or more unfavourable physico-chemical and environmental parameters [47]; and finally, III: polyextremophilic or polyextremotolerant organisms, which adapt to or survive in areas where extremely unfavourable physico-chemical parameters are present [48,49]. In addition, they can be also classified based on the environments in which they can thrive, for instance, the organisms that grow at high temperatures are termed thermophilic and hyperthermophilic; the psychrophilics that can grow at low temperatures; or those who are adapted to acidic or basic pH environments and are referred to as acidophilic or alkaliphilic. The species that grow in high-pressure conditions, such as in the deep sea or underground, are called piezophilic or barophilic, and finally, there are organisms that require NaCl to grow or survive in environments with high salt concentrations and low water activity(aw) that are generally called halophilic [50,51].
Incidentally, the molecular perception and mechanism for survival in such extreme ecosystems has been studied [23]. Apparently, structural changes and biologically active metabolites that are catalytically active under these conditions seem to play an essential role in tolerance and adaptability [52,53]. Biodiversity in the microbial community produces a wide range of secondary metabolites that are limited by many factors and change under stress condition [17,54]. This means that under high stress condition, biodiversity tends to be lower whereas, species diversity can increase when stress level decrease [55,56]. Now, we know that increased levels of stress can induce genetic mutations in all living organisms, and sometimes, the mutations and survival are considered as an alternative method to adapt, proliferate, and grow in extreme conditions, with consequent production of important metabolites with medicinal properties [57,58,59].
Natural products with medicinal or anticancer properties are called bioactive molecules that are produced by living organisms [60]. Although the majority of natural anticancer products are derived from plants, microorganisms, especially fungi, can also be considered as excellent sources of these ingredients, with even more attractive properties [61,62]. They can be manipulated easily and screened biologically and physiologically to discover novel natural products with extraordinary diversity among living organisms [63]. The metabolites produced by extremophiles and extremotolerant fungi are very valuable to produce drugs against cancer cells [64]. In addition, fungi produce other bioactive metabolites such as mycotoxins, antimycotics [65,66,67], immunosuppressants [68], antifungals [69], antibacterials [70], and antivirals [71]. Even though intensive research has been conducted on fungi-derived natural products and their medicinal properties for more than a century, there are still several fungal metabolites with unknown biological activities that can be considered as new sources of drugs [37].
The following sections briefly review some of the major bioactive antitumor (anticancer) metabolites isolated from extremophilic, extremotolerant, and polyextremophilic (or tolerant) fungi reported in the published literature from 1984 to 2023; the reported effective doses, the types of target cells and the omics approaches used for their identification are also addressed.
2. Fungi of Extreme Temperature Conditions
Temperature is one of the important factors that cause selective pressure on microorganisms like fungi [72] and may induce the integration of cellular components, which leads to structural and functional alterations of biological molecules [73,74]. Psychrophiles are one type of extremophiles which grow and live in temperatures lower than 15 degrees Celsius [75,76], while psychrotolerants proliferate optimally and grow at 15–30 °C [77,78]. Several species of micro-fungi have been reported from the Antarctic region that can survive the subzero temperatures [79]. On the other hand, thermophilic fungi are extremophiles that can proliferate at approximately 45 °C and can tolerate the higher temperatures of up to 61 °C [74,80]. The unique special compounds of these fungi (e.g., heat-resistant enzymes) can be of high interest for modern biotechnology [81,82]. The biosynthetic pathways, structures, and applications of these bioactive compounds are yet to be fully characterized [82,83,84].
Psychrophilic or psychrotolerant fungi produce certain metabolites at low temperatures that are perhaps essential for their survival in harsh environments such as the deep sea of oceans with cold water, high pressure, and nutrient deficiencies [85,86]. These fungi are classified as polyextremophile or tolerant to multiple environmental stresses [87].
So far, only a limited number of studies have been conducted to identify biologically active compounds from this group of extreme fungi [88]. As instances of these calls of compounds with cytotoxic activity against tumour cell lines, a novel cyclic nitropeptide named Psychrophilin A–D from Penicillium algidus (a psychrophilic fungi) was isolated from soil specimens in Greenland [89]. Psychrophilin A–D showed moderate cytotoxic activity with an effective dose (ID50) value of 10.1 μg/mL against P388 murine leukemia cell lines [89]. Piperazine-type compounds, including Oidioperazines A–D and Chetracins B–D produced by the fungus Oidiodendron truncatum GW3-13, were also isolated from soil samples in the Antarctic station near the Great Wall of China [77]. The cytotoxic activity of these piperazine-type compounds against A2780, Bel-7402, HCT-8, BGC-823, and A549 cell lines was documented [42,65].
Thermophilic or thermotolerant fungi are microorganisms that live in hot springs with temperatures between 45 °C and 65 °C [74]. A review of articles on the characterization of fungi in such a harsh environment revealed the incompleteness in the structural clarification, identification, and biological activities of their metabolites. Aspergillus terreus is one thermophilic fungus that produces lovastatin, which has a selective prohibitory effect on primary AML cell growth up to 75–95%. Moreover, the detracted proliferation by lovastatin have been reported with moderate inhibitory effect and induced apoptosis on four lung cancer cell lines and in ten ovarian cancer cell lines, respectively [31,46]. In addition, it has been recently found that lovastatin with IC50 values of 0.6, 0.7, and 1.1 μg/mL inhibited the liver cancer HepG2, cervical cancer HeLa, and breast cancer MCF-7 cell lines, respectively [37].
Simvastatin is another metabolite that was extracted from A. terreus with prohibitive effect on the growth of three melanoma, two lung, and four breast cancer cell lines and with an apoptosis effect of reducing tumour growth in hepatic cancer cells with IC50 values between 0.8 and 5.4 μM, making it an anticancer drug candidate for different clinical experiments [37].
Asperlin and brefeldin A are small polyketides produced by Aspergillus terreus (JAS-2) whose anticancer activities have been demonstrated for decades after their discovery [37]. Terrein, a compound isolated also from A. terreus, is a small antifungal agent known since 1935 and widely studied for its anticancer properties for almost 80 years. Terrein with an IC50 value of 1.1 nM was reported to cause apoptosis in breast cancer (MCF-7 cell line); in addition, its activity against liver cancer cell lines of HepG2 (IC50 66.8 μM) and pancreatic PANC-1 (IC50 9.8 μM) was also reported [37]. It is noteworthy to mention that terrein’s activity against the mentioned cancer cell line was 100 times stronger than Taxol, which is extracted from Taxus baccata [37].
Fumagillin is another metabolite first isolated from the liquid culture of Aspergillus fumigatus strain H-3.77 and later found in other species of Aspergillus such as A. flavus and A. parasiticus. One of the chlorinated derivatives of fumagillin was extracted from a marine-inhabiting Penicillium strain with potent inhibitory activity against osteosarcoma cell lines [90,91,92]. TNP-470, PPI-2458, and CKD-732 are synthetically derived fumagillin analogs that have been tested in human cancer clinical experiments. These molecules interact with the enzyme methionine aminopeptidase type 2 that cleaves the N-terminal methioninyl residue of newly synthesized proteins, interrupting the tumour vessels [91].
Chromatographic separation and molecular identification of the obtained fragments of Malbranchea sulfurea metabolites revealed six light-sensitive polyketides called malbran pyrroles A–F with cytotoxic activities against PANC-1, HepG2, and MCF-7 cancer cell lines, with their IC50 values in the range from 3 to 11 μM 42 [42,51]. More metabolites of this group of fungi and their biological activities are listed in Table 1, with their half inhibitory concentrations (IC50), specifically on human cancer cell lines, unless otherwise, stated. These IC50 values serve as critical reference points in determining the potency and efficacy of these agents against various cancers.
3. Piezophilic Fungi
Piezophiles are organisms that proliferate optimally under high pressure (≤100 MPa), while piezo-tolerants can grow optimally at pressures above 0.1 MPa [42]. Piezophilic fungi adapt and grow in deep biospheres of the sea and oceans [105]. In research on natural medicinal products, the marine Acremonium fungi gained more attention for their antimicrobial cephalosporins compared to their less explored terrestrial counterparts [42,106]. Piezophiles and piezo-tolerants of extreme environments deserve more exploration due to their ability to produce novel biologically active compounds with potential therapeutic properties [107]. However, it should be emphasized that the research on these fungi and their secondary metabolites is very confined because of the sampling challenges and the limitation of adequate culture simulation in the laboratory. However, the advanced execution of deep-sea sampling and drilling methods, as well as omics technologies and culture-dependent methods, are being used as part of current approach to identify cytotoxic compounds and metabolites from the marine environment [30,68,108]. A novel hydroxyphenylacetic acid compound named Westerdijkin A, with anticancer activity against K562 and HL-60 cell lines, was isolated from Aspergillus westerdijkiae in deep-sea sediments of the South China Sea, at a depth of 4593 m [109]. Acaromyester A and Acaromycin A are other cytotoxic compounds found in Acaromyces ingoldii FS121 from the depth of 3415 m in the South China Sea. These bioactive metabolites presented cytotoxic activities against cancer cell lines such as breast cancer (cell line MCF-7) and hypotriploid lung cancer (cell line NCI-H46) [110,111,112].
Penicillium is a genus of ascomycete fungus widely distributed in deep-sea sediments and has received a lot of attention due to its important secondary metabolites. Most of the secondary metabolites of Penicillium have been reported to have anticancer properties [113,114]. Recent studies are revealing the diversity of fungi in the Arabian Sea marine environment, but despite such biodiversity, the biological potential of sea fungi from the region is rarely studied [30]. One of the fungal strains separated from marine sediment specimens was recognized as Penicillium dipodomyicola by ITS rDNA sequencing and identified as Penicillium sp. ArCSPf [30]. Due to its ability to tolerate high salt concentrations, low temperatures, and pressure changes, Penicillium sp. ArCSPf is one of the most prevalent and dominant species of the area. Studies on fungal diversity from the Arabian Sea continental slope showed that approximately 43% of fungal isolates belong to the genus Penicillium [30,115]. The results of current research show that piezophilic or barophilic fungi can be considered as rich sources of natural products with significant antibiotic and cytotoxic activity [30]. Brevione isolated from Penicillium spp., living in deepest part of the sea, has illustrated strong cytotoxic activity [93,96]. In addition, terpenoid derivatives with potential cytotoxicity are the other class of compounds, frequently found in penicillium and Aspergillus species of the deep-sea waters [105,116]. Some of the most interesting research on these fungi and their identified metabolites, with biological activities like anticancer properties, are summarized in Table 2.
4. Acidophilic and Alkaliphilic Fungi
Fungi normally grow at a slightly acidic pH of around 5.0–6.0. In contrast, acidophiles grow optimally at pH values ranging from 3–4, whereas the alkaliphiles can thrive the higher pH values of above 9 [123].
Emericellipsin A is an example compound isolated from Emericellopsis alkaline and Streptomyces SPP (alkalophilic fungi strains) and is presumed to be an effective novel antitumor substance with a significant cytotoxic effect against HepG2 and HeLa tumour cell lines [69,124].
Moreover, many peptaibols have been isolated from Emericellopsis sp such as Emericellipsin A–E, where in vitro tests displayed selective and suppressing cytotoxic activity toward tumour cells by activating calcium-mediated apoptosis against HepG2 and Hela cell lines [125]. Penicillium species of the surface waters of an acid mine produce two activators of signal transduction enzyme inhibition, including caspase-1 (Casp-1) and matrix metalloproteinase-3 (MMP-3) inhibitors [126]. Specific Casp-1 inhibitors may serve as new drug with highly potent cytotoxic activities [127,128,129,130]. The specific MMP inhibitors represented a novel therapeutic path to treat cancers by obstructing the activity of MMPs abused by tumour cells. There are reports on stabilizing the tumour progression effect of these inhibitors when applied as low-toxicity supplements for cytotoxic treatments [131,132].
Berkeley Pit Lake, located in Butte, Montana, is famous for its abandoned open-pit copper mine 540 m below the Earth’s surface, saturated with metal-sulphate-contaminated acidic water of over 140 billion L [133,134,135,136]. In recent years, several studies have focused on the metabolites from the microorganisms growing in this harsh environment [41]. A novel spiroketal compound named berkelic acid and two new hybrid polyketide–terpenoid compounds known as berkeleydione and berkeleytrione have been isolated recently from the Penicillium species of the lake. These metabolites, along with several other compounds such as berkeleyacetals A–C from an unknown species of Penicillium, were believed to have biological activities such as anticancer properties [126,133,136]. Berkelic acid was tested against 60 human cell lines displaying selective anticancer properties against ovarian cancer OVCAR-3 with a GI50 of 9.13 (91 nM concentration) [133]. The chemical structure of berkelic acid was revised by the Fürstner group using methods like synthetic NMR and crystallographic techniques in 2018 [137]. Penicillium rubrum produces Berkeleyamides A–D and berkeleyones A–C in the low micromolar range that were tested positive for the suppression of MMP-3 and Casp-1 via interleukin 1-β inhibition in THP-1 cells [99,138,139]. Penicillium solitum and Penicillium purpurogenum JS03-21 produce a new tricarboxylic acid derivative and two new drimane sesquiterpene lactones named berkedrimanes A and B, which, at low micromolar concentrations, reduce the production of cytokines interleukin (IL-1β) by inflammasomes [128,140,141]. Berkazaphilones A and B, berkedienolactone, octadienoic acid derivatives, berkedienoic acid [100,101], vermistatin, dihydrovermistatin, penisimplicissin, aldehyde, azaphilone, and methylparaconic acid were isolated from a culture broth of Penicillium rubrum [134,139,142]. The compounds were isolated either for their inhibitory effect on the signal transduction enzyme Casp-1 or because of their structural similarity to such inhibitors [139]. Selected compounds were further evaluated for their ability to inhibit interleukin-1β production by inflammasomes in induced THP-1 cells. Berkazaphilones B and C and a penisimplicissin analogue of vermistatin showed selective activity against human leukaemia cancer cell lines [134,139] (Table 3).
5. Halophiles and Other Poly Extremophiles
Halophiles are organisms found in all domains of life, including bacteria, archaea, and eukarya, and are generally referred to as “salt-loving” organisms. The halophile fungi groups have a variety of properties and valuable secondary metabolites.
Genomic and molecular studies have revealed the differential expression of many genes involved in the biosynthesis of secondary metabolites in these organisms compared to their expression under normal conditions [151,152]. The obtained compounds from these fungi can potentially be used in industrial and biotechnological fields, as well as in the production of β-carotene, ectoine, and bio-rhodopsin for optical computing, and in holograms, biosensors, photoelectric devices, cosmetics, preservatives, fermented food products, the manufacturing of bioplastics and artificial retinas, bio-surfactants and exopolysaccharides, biofuels, pigments for colouring, and even bioremediation [153,154,155,156,157].
Halophiles are confined and adapted to environments where salt is present and, in many cases, is needed for their survival. In addition to the high salt concentrations, halophilic microorganisms are exposed to several types of other abnormalities, such as nutritional deficiencies, osmotic pH, UV, and ionic stress. Unlike halophilics, the non-tolerant organisms with no adaptability to these conditions lose water through their cells and tissues faster, during which the turgor pressure loss causes cytosolic dehydration [157,158,159]. Halophiles are considered one of the most valuable resources of bioactive compounds due to their low water activity that triggers the formation of rare secondary metabolites with potential anticancer properties [160,161]. Studies have shown that the biosynthesis of anticancer compounds such as cytochalasin E, ergosterol, and rosellichalasin from Aspergillus sp. increased at higher salt concentration of the medium [37,83]. The mentioned compounds diminished the durability of BEL-7402, RKO, A549, and Hela human cancer cell lines. Among them, ergosterol had the greatest inhibitory effect on a human colon cancer cell line [160,161,162]. The temperate and subtropical regions host P. chrysogenum or P. rubens, the producers of two interesting compounds named chloctanspirone A and B [135,163]. These compounds are the first chlorinated sorbicillinoids obtained from natural origins and are distinguished by their unique ring structures [37]. Chloctanspirone A is the more active analogue with a greater prohibitory effect on human leukaemia HL-60 and lung cancer cell line A-549 compared to chloctanspirone B, with its moderate or non-existent activity against the same cell lines [164].
The pullulan polymer molecules that have been produced by A. pullulans are considered important polysaccharides with biological activities [165,166]. Studies have demonstrated pullulan’s effect on the T and B cells by means of altering the immune-stimulatory system [167]. Pullulan treatment upregulates the costimulatory molecular expression and enhances the pro-inflammatory cytokine production in bone marrow-derived dendritic cells (BMDCs) in vitro and in spleen DCs in vivo [167,168]. Pullulan induced the maturation of DCs in spleen- and tumour-draining lymph nodes (drLN) and in tumour-bearing mice and promoted the OVA-specific T cell activation and migration of the T cells into the tumour [169,170]. The combination of OVA and pullulan inhibited tumour growth and liver metastasis [171,172]. It was demonstrated that treatment with a combination of pullulan and tyrosinase-related protein 2 (TRP2) peptide suppressed the B16 melanoma growth [173]. It was also found that pullulan can not only enhance the DCs’ maturation and function, but also it acted as an adjuvant in promoting antigen (Ag)-specific immunity in mice [166]. Thus, pullulan could be a new and useful adjuvant to be considered in therapeutic cancer vaccines. Another relative compound is liamocin oil, which is structurally unique and heavier than water, produced by certain strains of fungi. One of the highest yields for liamocin production (7.0–8.6 g/L) was reported by Zhang et al. 2016 [166] from RSU 9 and RSU 21 strains of A. pullulans originated from the tropical environment of Thailand. The liamocins prohibited a human cervical cancer cell line and two human breast cancer cell lines with IC50 values of 32.2 ± 1.4–63.1 ± 2.4 μg liamocins/mL with no adverse effects on healthy cell lines [168]. This suggests that these compounds deserve more screening as potential anticancer drugs with critical and appropriate clinical trials and precise molecular identifications [174,175]. Some halophiles and other extremophiles and their anticancer biomolecules are shown in Table 4.
Fungi produce several pigments such as azaphilones, melanin, carotenoids, polyketides, etc., with positive roles in human well-being, while containing other harmful pigments. A. pullulans produces important pigments conferring strong anticancer activities with low cytotoxicity towards normal cells [166]. Alternaria sp. ZJ9-6B was screened for potential production of pigments against human breast cancer cell lines with positive outcomes. Pigments derived from Monascus purpureus like monascin presented prohibitory activity against mice skin carcinogenesis, while ankaflavin showed activity against Hep G2 and A549 human cancer cell lines [183,184]. Similarly, monaphilone A and B, isolated from M. purpureus, showed promising anti-proliferative effects on HEp-2 human laryngeal carcinoma cell lines [185,186,187]. Table 5 summarizes the other contaminated environments’ fungal metabolites with anti-cancer properties. The chemical structure of the selected novel antitumour compounds from different groups of extremophilic fungi is presented in Figure 1.
6. “Omics” Tools for Discovery of Bioactive Metabolites of Extremophilic Fungi
The biosynthetic pathways of bioactive metabolites of microorganisms in extreme environments can be revealed by the application of “Omics techniques” with better resolution and predictability to accelerate the generation of information for the natural products research communities. This approach has already been applied for studying single microorganisms or microbial communities in extreme environments [192,193,194]. In this section, an important algorithmic perspective behind several computing platforms, software, and databases is provided to provide an overview of the various computations for omics tools approaches to the elucidation of metabolic pathways of fungal metabolites in harsh environments.
Some of the important fungi-related platforms are ‘antiSMASH’ ‘fungiSMASH’, ‘NP.searcher’, ‘ClustScan’, ‘CLUSEAN’, ‘SMURF, SBSPKS, ‘MIDDAS-M’, ‘CLUster’ ‘CASSIS/SMIPS’, ‘NRPSPredictor’, ‘SBSPKS’, ‘NaPDoS’, ‘GNP/PRISM’, ‘EvoMining’, and ‘C-Hunter’ [195,196,197]. Most of these tools are freely accessible to users with limited programming skills. They usually contain algorithms that can align and search for interactive regions at the level of gene clusters and find and compare similarities with their closest relatives from other databases. This is then followed by identification prediction of metabolites that are likely to be synthesized from the gene sequences that are already available in the databases. Extraction algorithms and scalable expression platforms give very wide access to fungus-derived secondary metabolites’ biosynthetic pathways [198,199]. These are promising tools for identifying the secondary metabolites and their biosynthetic gene clusters, evaluating the genetic potential of different strains, and more effectively discovering currently unknown metabolites [195,196]. In addition, this information can be considered for the metabolic engineering of molecules by using synthetic biology approaches [192,197].
Metabolic gene clusters include a group of co-located genes that encode enzymes for the biosynthesis of a particular metabolite and are prominent features of chromosomes; therefore, the sequencing of these known gene families provides information that can be used to understand the presence of a potential secondary metabolite biosynthesis pathway in different species [198]. There are two main strategies for exploiting bioinformatics tools to identify gene clusters that are involved in biosynthetic pathways with high accuracy [199]. The first step of the gene mining process is to identify genes encoding conserved enzymes/protein domains that have associated roles in secondary metabolism [200,201], such as “adenylation”, “condensation”, glycosylation, or peptidyl carrier proteins, and domains of nonribosomal peptide synthetases. In the second step, defined algorithms are specifically used to associate the presence of such similarities with similar classes of natural products for intra- or inter-species matches [192,202].
Bioinformatics methods are mainly used to primarily locate the main so-called “backbone” enzymes, such as nonribosomal peptide synthetases, polyketide synthases, and their hybrids (peptide synthetases-polyketide synthases), in order to accurately identify gene clusters from known families and very specific classes [203].
Meanwhile, modern algorithms have been successfully designed for identifying potential gene clusters using the de novo-based method, i.e., without prior knowledge of functional domains or known patterns of key enzymes. It should be noted that gene clusters for many of the specific metabolic pathways have been discovered in common filamentous bacteria and fungi, but more are now being found in plants and higher organisms [204].
Next-generation sequencing provides more and more opportunities for studying microbial diversity and for predicting rare metabolic pathways and genes that are involved in stress tolerance and the survival of microorganisms in extreme environments [205].
One of the key shortages in the applicability of “Meta-Omics” is the absence of real interactivity when new results for the new bioactive compounds are found. In this regard, studies based on genomic data can widely explain the genetic basis of secondary metabolite biosynthesis, particularly for the genes encoding for characteristic secondary metabolites in a better organized manner [206]. Additionally, genomics has opened a new door for the engineering of new analogues of many complex structural metabolites by sequencing the genomes of fungal species and identifying biosynthetic pathways [207]. The ability of genome mining to search for the biosynthetic roots of secondary metabolites based on the distribution of polyketide synthases and non-ribosomal peptide synthetase gene clusters can reveal previously undefined pathways. Therefore, the integration of genome mining, computational techniques, and analytical biochemistry could be the future of natural product research [208,209] (Figure 2).
On the other hand, metabolomics and analytical chemistry techniques provide distinct strategies for metabolic engineering, which are provided by the hierarchical structure of the regulation of secondary metabolites’ biosynthesis, such as the manipulation of universal regulators to enhance the generation of secondary metabolites [210], or by targeting metabolic pathways for specific regulators to increase the content of a particularly interesting compound [208,211]. Culture condition screening is considered a successful and well-stablished experimental approach to detecting specialized secondary metabolites. In this method, organisms are exposed to different growth conditions or external stressors; then, the culture medium is analysed for secreted molecules using techniques such as mass spectrometry and nuclear magnetic resonance spectroscopy [212].
Developing modern computational techniques such as spectrum search algorithms, DEREPLICATOR+ is useful for identifying natural polypeptide products which contain ribosomal synthesized, or post-translationally modified, peptides and non-ribosomal peptide synthetase [213]. Machine learning can be extended for the identification of terpenes, benzenoids, flavonoids, polyketides, alkaloids, and other classes of natural products [214]. “DEREPLICATOR+” is one of the services that enable mutual validation of peptidogenomics/glycogenomics as a new genome mining strategy [214].
It should be emphasized that obtaining pure active compounds is a common problem in laboratories with limited facilities as it is accompanied by the cost and time of data reproduction and analytics. In this regard, it is important to consider the management of big data with proper analysis that can accelerate the bio-discovery efforts by providing rationale data for libraries and databases [215,216].
MeFSAT is an in silico chemical library with its primary objective being building a non-redundant resource of secondary metabolites of medicinal fungi along with information on their two-dimensional (2D) and three-dimensional (3D) chemical structures [217]. The information compiled in the MeFSAT database is openly accessible [218].
Advanced high-throughput sequencing technologies revolutionized microbiology, particularly in studies conducted in extreme environments, showing the high diversity and complexity of those microbial ecosystems. However, understanding the physiology of microorganisms in a particular environment, such as natural extreme environments or in vitro laboratory simulations, is now considered essential for completing genomic or transcriptomic studies that cannot be replaced by any other method.
As a result, a combination of traditional cultivation methods and modern culture-independent techniques can be considered the best ways to better understand the growth and multiplication of microorganisms in harsh environments [219,220,221]. In several cases, the identification of novel metabolites originates from initial screening and is not researched with further genome sequencing and dereplication [222,223].
So, to summarize the above, with the help of approaches such as “Omics” and genome mining, optical measurement, real screening, and three-dimensional models, with the support of informatics-based analysis and artificial intelligence (AI), the dosage of the effective substance in a natural product can be determined for diagnosis of cancerous conditions and its phases [224,225,226]. Today, recent advances in “Omics” techniques can be utilized as tools for detecting cellular malfunctions that are at the centre of multi-factorial diseases such as cancer [227,228]. Genomics and proteomics are thought to enable the rapid [229], complete, and parallel analysis (pipeline processes) of genes and proteins that are expressed in a particular cell or type of tissue [230]. On the other hand, turning a normal cell into a cancer stage requires molecular changes at various levels, including the genome, epigenome, transcription, proteome, and metabolome [231]. In the field of cancer diagnosis, a variety of different profiles of cancer lines and natural tissues can be used to determine genomic or proteomic characteristics [232]. Since the availability of several genomes data and “Gene and protein expression profiles”, such data have had a significant impact on cancer research and therapies and led to genomics turning into a distinguished technique that has developed continuously by using genetic sequences in cancer research [233,234]. This method extensively qualifies the rate and probability of somatic mutations in individual patients and by applying big data to identify environmental factors, which are related to cancer, mutagenesis, and germline predispositions [235,236]. “Gene and protein expression profiles” also have the potential to improve the clinical management of cancers by providing classification, either by developing class prediction or class discovery or by providing information for diagnostic classification, and to identify new molecular targets to improve patient care through the identification of profiles that predict responsiveness to therapy or prognosis [237,238,239].
Finally, the generation of such information and turning it into theories depends on genome sequencing to create information systems to assist physicians in making decisions about the diagnosis and treatment of cancer in patients [239].
A schematic view of the stages of early clinical trials leading to the discovery of cancer drugs is shown in Figure 3. The current and future of drug discovery can rely on the utilization of a combination of multi-omics and molecular and analytical approaches [240].
A collection of bioinformatics tools and databases for the identification of metabolic gene clusters related to drug discovery of extremophiles and extremotolerant fungi are summarized in Table 6. The details of the platform or tool, system requirements, input formats, function, output, and finally the accessibility options are included in this table.
A schematic view of the stages of early clinical trials leading to the discovery of cancer drugs is shown in Figure 3. In addition, we would like to address a new approach, which would be the chemical modification of the natural metabolites with conjugational alterations in laboratory and to investigate the potential functionality enhancement of such hybrid molecules [280,281].
7. Conclusions
In the fascinating world of microbiology, fungi that can survive and multiply in harsh environments have an unusual biochemistry that is highly regarded in biotechnology applications in industry, pharmaceuticals, environmental protection, medicine, and many other fields. Secondary metabolites of fungi from extreme environments are considered novel bioactive compounds that have attracted much attention in recent years. They produce essential, beneficial natural metabolites related to a variety of applications, including novel drugs and medicines to be applied in therapies as immunosuppressants, cytotoxic and anticancer agents, and antimicrobials, anti-inflammatories, and antioxidants. They also serve as resources for industrial and biotechnological applications as enzymes and nutrients, such as β-carotene, ectoine, and bio-rhodopsin for optical data processing; as bio-surfactants and exopolysaccharides in bioremediation and biofuels; and as pigments for the dye and food industries.
In the era of omics, on one hand, with the rapid development of recent tools and techniques, the alteration in culture conditions or the generation of mutants could also give rise to modified strains with the ability to produce novel important metabolites; on the other hand, these new methods can be used to identify valuable biological resources. Synthesizing such information and turning it into theories can take over the present and future of drug discovery as being referred to a combination of multi-omics and molecular and analytical approaches.
In this article, some of the most well-known and practical bioinformatics tools/platforms and databases for the study of fungal metabolism are presented, especially for those living in extreme environments.
In addition, around 260 compounds were collected from previous reports. To address the new trend of looking for medicines of natural origin, several compounds from extremophilic, extremotolerant, and poly (extremophile and tolerant) fungi were summarized in different tables of this work. These include compounds from fungi that reproduce, grow, and develop in extreme temperature conditions, compounds belonging to piezophilic or barophilic (high pressure) groups, compounds selected from fungi that can live in acidic or alkaline environments, compounds found in environments with high salt concentrations and low water activity, and, finally, compounds that were discovered in other types of extreme environment. The metabolites mentioned in this review have strong or moderate bioactive properties and may have the potential to be used in future cancer drug discovery and therapies.
Author Contributions
Conceptualization, F.H.A. and A.K.; resources, V.P. and I.M.; writing—original draft preparation, F.H.A.; writing—review and editing, I.M., A.K., A.M. and V.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
F.H.A. is thankful to Peter Pregelj, Primož Rus, Rok Tavčar, and to Sara Eshmoel for carefully checking the references.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nagai, H.; Kim, Y.H. Cancer Prevention from the Perspective of Global Cancer Burden Patterns. J. Thorac. Dis. 2017, 9, 448. [Google Scholar] [CrossRef] [PubMed]
- Fouad, Y.A.; Aanei, C. Revisiting the Hallmarks of Cancer. Am. J. Cancer Res. 2017, 7, 1016. [Google Scholar] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Cao, B.; Varghese, C.; Mikkelsen, B.; Weiderpass, E.; Soerjomataram, I. Comparing Cancer and Cardiovascular Disease Trends in 20 Middle-or High-Income Countries 2000–19: A Pointer to National Trajectories towards Achieving Sustainable Development Goal Target 3.4. Cancer Treat. Rev. 2021, 100, 102290. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.C.; Kayamba, V.; Peek, R.M., Jr.; Heimburger, D. Cancer Control in Low-and Middle-Income Countries: Is It Time to Consider Screening? J. Glob. Oncol. 2019, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.-M.; Lee, D.; Kong, H.-J.; Lee, S.; Won, Y.-J.; Jung, K.-W.; Cho, H. Causes of Death among Cancer Patients in the Era of Cancer Survivorship in Korea: Attention to the Suicide and Cardiovascular Mortality. Cancer Med. 2020, 9, 1741–1752. [Google Scholar] [CrossRef]
- Niculescu, A.-G.; Grumezescu, A.M. Novel Tumour-Targeting Nanoparticles for Cancer Treatment—A Review. IJMS 2022, 23, 5253. [Google Scholar] [CrossRef]
- DeSantis, C.E.; Lin, C.C.; Mariotto, A.B.; Siegel, R.L.; Stein, K.D.; Kramer, J.L.; Alteri, R.; Robbins, A.S.; Jemal, A. Cancer Treatment and Survivorship Statistics, 2014. CA Cancer J. Clin. 2014, 64, 252–271. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Ju, D.-T.; Chang, C.-F.; Reddy, P.M.; Velmurugan, B.K. A Review on the Effects of Current Chemotherapy Drugs and Natural Agents in Treating Non–Small Cell Lung Cancer. Biomedicine 2017, 7, 23. [Google Scholar] [CrossRef]
- Schirrmacher, V. From Chemotherapy to Biological Therapy: A Review of Novel Concepts to Reduce the Side Effects of Systemic Cancer Treatment. Inter. J. Oncol. 2019, 54, 407–419. [Google Scholar] [CrossRef]
- Falzone, L.; Salomone, S.; Libra, M. Evolution of Cancer Pharmacological Treatments at the Turn of the Third Millennium. Front. Pharmacol. 2018, 9, 1300. [Google Scholar] [CrossRef] [PubMed]
- Varghese, E.; Samuel, S.M.; Líšková, A.; Samec, M.; Kubatka, P.; Büsselberg, D. Targeting Glucose Metabolism to Overcome Resistance to Anticancer Chemotherapy in Breast Cancer. Cancers 2020, 12, 2252. [Google Scholar] [CrossRef] [PubMed]
- Narsing Rao, M.P.; Xiao, M.; Li, W.-J. Fungal and Bacterial Pigments: Secondary Metabolites with Wide Applications. Front. Microbiol. 2017, 8, 1113. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The Traditional Medicine and Modern Medicine from Natural Products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.-E.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef]
- Chavali, A.K.; Rhee, S.Y. Bioinformatics Tools for the Identification of Gene Clusters That Biosynthesize Specialized Metabolites. Brief. Bioinform. 2018, 19, 1022–1034. [Google Scholar] [CrossRef]
- Hariri Akbari, F. An overview of Stress in cellular and molecular levels and the importance of studying responses to stresses in biology. Res. J. Biotech. 2021, 16, 258–282. [Google Scholar]
- Ncube, B.; Ndhlala, A.R.; Van Staden, J. Secondary Metabolism and the Rationale for Systems Manipulation. In Transgenesis and Secondary Metabolism; Jha, S., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–22. ISBN 978-3-319-27490-4. [Google Scholar]
- Huang, M.; Lu, J.-J.; Ding, J. Natural Products in Cancer Therapy: Past, Present and Future. Nat. Prod. Bioprospect. 2021, 11, 5–13. [Google Scholar] [CrossRef]
- Alamgir, A.N.M. Biotechnology, In Vitro Production of Natural Bioactive Compounds, Herbal Preparation, and Disease Management (Treatment and Prevention). In Therapeutic Use of Medicinal Plants and their Extracts: Volume 2; Progress in Drug Research; Springer International Publishing: Cham, Switzerland, 2018; Volume 74, pp. 585–664. ISBN 978-3-319-92386-4. [Google Scholar]
- Chemat, F.; Abert Vian, M.; Ravi, H.K.; Khadhraoui, B.; Hilali, S.; Perino, S.; Fabiano Tixier, A.-S. Review of Alternative Solvents for Green Extraction of Food and Natural Products: Panorama, Principles, Applications and Prospects. Molecules 2019, 24, 3007. [Google Scholar] [CrossRef]
- Turk, M.; Gostinčar, C. Glycerol Metabolism Genes in Aureobasidium pullulans and Aureobasidium subglaciale. Fungal Biol. 2018, 122, 63–73. [Google Scholar] [CrossRef]
- Chávez, R.; Fierro, F.; García-Rico, R.O.; Vaca, I. Filamentous Fungi from Extreme Environments as a Promising Source of Novel Bioactive Secondary Metabolites. Front. Microbiol. 2015, 6, 903. [Google Scholar] [CrossRef]
- Jakubczyk, D.; Dussart, F. Selected Fungal Natural Products with Antimicrobial Properties. Molecules 2020, 25, 911. [Google Scholar] [CrossRef]
- Bozoudi, D.; Tsaltas, D. The Multiple and Versatile Roles of Aureobasidium pullulans in the Vitivinicultural Sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef]
- Vala, A.K.; Sachaniya, B.K.; Dave, B.P. Marine Fungal White Biotechnology: An Ecological and Industrial Perspective. In Recent Advancement in White Biotechnology Through Fungi; Yadav, A.N., Mishra, S., Singh, S., Gupta, A., Eds.; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2019; pp. 485–503. ISBN 978-3-030-10479-5. [Google Scholar]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The Amazing Potential of Fungi: 50 Ways We Can Exploit Fungi Industrially. Fungal Divers. 2019, 97, 1–136. [Google Scholar] [CrossRef]
- Gostinčar, C.; Ohm, R.A.; Kogej, T.; Sonjak, S.; Turk, M.; Zajc, J.; Zalar, P.; Grube, M.; Sun, H.; Han, J.; et al. Genome Sequencing of Four Aureobasidium pullulans Varieties: Biotechnological Potential, Stress Tolerance, and Description of New Species. BMC Genomics 2014, 15, 549. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-D.; Xie, L.; Wei, Y.; Zhou, X.; Jia, B.; Liu, J.; Zhang, S. Abiotic Stress Resistance, a Novel Moonlighting Function of Ribosomal Protein RPL44 in the Halophilic Fungus Aspergillus Glaucus. Appl. Environ. Microbiol. 2014, 80, 4294–4300. [Google Scholar] [CrossRef]
- Farha, A.K.; Hatha, A.M. Bioprospecting Potential and Secondary Metabolite Profile of a Novel Sediment-Derived Fungus Penicillium Sp. ArCSPf from Continental Slope of Eastern Arabian Sea. Mycology 2019, 10, 109–117. [Google Scholar] [CrossRef]
- Coker, J.A. Extremophiles and Biotechnology: Current Uses and Prospects. F1000Research 2016, 5, 396. [Google Scholar] [CrossRef]
- Demain, A.L. Valuable Secondary Metabolites from Fungi. In Biosynthesis and Molecular Genetics of Fungal Secondary Metabolites; Martín, J.-F., García-Estrada, C., Zeilinger, S., Eds.; Fungal Biology; Springer New York: New York, NY, USA, 2014; pp. 1–15. ISBN 978-1-4939-1190-5. [Google Scholar]
- Thomford, N.; Senthebane, D.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef]
- Ncube, B.; Van Staden, J. Tilting Plant Metabolism for Improved Metabolite Biosynthesis and Enhanced Human Benefit. Molecules 2015, 20, 12698–12731. [Google Scholar] [CrossRef]
- Gaynes, R. The Discovery of Penicillin—New Insights After More Than 75 Years of Clinical Use. Emerg. Infect. Dis. 2017, 23, 849–853. [Google Scholar] [CrossRef]
- Egbuta, M.; Mwanza, M.; Babalola, O. Health Risks Associated with Exposure to Filamentous Fungi. Int. J. Environ. Res. Public Health 2017, 14, 719. [Google Scholar] [CrossRef] [PubMed]
- Bladt, T.; Frisvad, J.; Knudsen, P.; Larsen, T. Anticancer and Antifungal Compounds from Aspergillus, Penicillium and Other Filamentous Fungi. Molecules 2013, 18, 11338–11376. [Google Scholar] [CrossRef]
- Bills, G.F.; Platas, G.; Fillola, A.; Jiménez, M.R.; Collado, J.; Vicente, F.; Martín, J.; González, A.; Bur-Zimmermann, J.; Tormo, J.R.; et al. Enhancement of Antibiotic and Secondary Metabolite Detection from Filamentous Fungi by Growth on Nutritional Arrays. J. Appl. Microbiol. 2008, 104, 1644–1658. [Google Scholar] [CrossRef] [PubMed]
- Ancheeva, E.; Daletos, G.; Proksch, P. Bioactive Secondary Metabolites from Endophytic Fungi. CMC 2020, 27, 1836–1854. [Google Scholar] [CrossRef] [PubMed]
- Rampelotto, P.H. Extremophiles and Extreme Environments. Life 2013, 3, 482–485. [Google Scholar] [CrossRef]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the Extremes: Extremophiles and the Limits of Life in a Planetary Context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef]
- Ibrar, M.; Ullah, M.W.; Manan, S.; Farooq, U.; Rafiq, M.; Hasan, F. Fungi from the Extremes of Life: An Untapped Treasure for Bioactive Compounds. Appl. Microbiol. Biotechnol. 2020, 104, 2777–2801. [Google Scholar] [CrossRef]
- Bang, C.; Dagan, T.; Deines, P.; Dubilier, N.; Duschl, W.J.; Fraune, S.; Hentschel, U.; Hirt, H.; Hülter, N.; Lachnit, T.; et al. Metaorganisms in Extreme Environments: Do Microbes Play a Role in Organismal Adaptation? Zoology 2018, 127, 1–19. [Google Scholar] [CrossRef]
- Al Tamie, M.S.S. Effect of Salinity on the Fungal Occurance in Al-Shega Area at Al-Qassim, Saudi Arabia. Res. J. Microbiol. 2014, 9, 287–295. [Google Scholar] [CrossRef]
- Vylkova, S. Environmental PH Modulation by Pathogenic Fungi as a Strategy to Conquer the Host. PLoS Pathog. 2017, 13, e1006149. [Google Scholar] [CrossRef]
- Lindgren, A.R.; Buckley, B.A.; Eppley, S.M.; Reysenbach, A.-L.; Stedman, K.M.; Wagner, J.T. Life on the Edge—The Biology of Organisms Inhabiting Extreme Environments: An Introduction to the Symposium. Integr. Comp. Biol. 2016, 56, 493–499. [Google Scholar] [CrossRef]
- Sibanda, T.; Selvarajan, R.; Tekere, M. Synthetic Extreme Environments: Overlooked Sources of Potential Biotechnologically Relevant Microorganisms. Microb. Biotechnol. 2017, 10, 570–585. [Google Scholar] [CrossRef]
- Segal-Kischinevzky, C.; Romero-Aguilar, L.; Alcaraz, L.D.; López-Ortiz, G.; Martínez-Castillo, B.; Torres-Ramírez, N.; Sandoval, G.; González, J. Yeasts Inhabiting Extreme Environments and Their Biotechnological Applications. Microorganisms 2022, 10, 794. [Google Scholar] [CrossRef] [PubMed]
- Plemenitaš, A.; Lenassi, M.; Konte, T.; Kejžar, A.; Zajc, J.; Gostinčar, C.; Gunde-Cimerman, N. Adaptation to High Salt Concentrations in Halotolerant/Halophilic Fungi: A Molecular Perspective. Front. Microbiol. 2014, 5, 199. [Google Scholar] [CrossRef] [PubMed]
- Reed, C.J.; Lewis, H.; Trejo, E.; Winston, V.; Evilia, C. Protein Adaptations in Archaeal Extremophiles. Archaea 2013, 2013, 373275. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Satyaprakash, K.; Das, A.K. Extremophilic Fungi as a Source of Bioactive Molecules. In Extremophilic Fungi; Sahay, S., Ed.; Springer Nature Singapore: Singapore, 2022; pp. 489–522. ISBN 9789811649066. [Google Scholar]
- Schröder, C.; Burkhardt, C.; Antranikian, G. What We Learn from Extremophiles. Chem. Texts. 2020, 6, 8. [Google Scholar] [CrossRef]
- DasSarma, S.; DasSarma, P.; Laye, V.J.; Schwieterman, E.W. Extremophilic Models for Astrobiology: Haloarchaeal Survival Strategies and Pigments for Remote Sensing. Extremophiles 2020, 24, 31–41. [Google Scholar] [CrossRef]
- Selim, M.S.M.; Abdelhamid, S.A.; Mohamed, S.S. Secondary Metabolites and Biodiversity of Actinomycetes. J. Genet. Eng. Biotechnol. 2021, 19, 72. [Google Scholar] [CrossRef]
- Hong, P.; Schmid, B.; De Laender, F.; Eisenhauer, N.; Zhang, X.; Chen, H.; Craven, D.; De Boeck, H.J.; Hautier, Y.; Petchey, O.L.; et al. Biodiversity Promotes Ecosystem Functioning despite Environmental Change. Ecol. Lett. 2022, 25, 555–569. [Google Scholar] [CrossRef]
- Hutchison, C.; Gravel, D.; Guichard, F.; Potvin, C. Effect of Diversity on Growth, Mortality, and Loss of Resilience to Extreme Climate Events in a Tropical Planted Forest Experiment. Sci. Rep. 2018, 8, 15443. [Google Scholar] [CrossRef] [PubMed]
- Brangsch, H.; Höller, M.; Krauβe, T.; Waqas, M.; Schroeckh, V.; Brakhage, A.A.; Bunk, B.; Spröer, C.; Overmann, J.; Kothe, E. Extremophile Metal Resistance: Plasmid-Encoded Functions in Streptomyces mirabilis. Appl. Environ. Microbiol. 2022, 88, e00085-22. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.S.; Nair, A.; Nivetha, K.; More, V.S.; Anantharaju, K.S.; More, S.S. Chapter 7—Molecular Adaptations in Proteins and Enzymes Produced by Extremophilic Microorganisms. In Extremozymes and Their Industrial Applications; Arora, N.K., Agnihotri, S., Mishra, J., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 205–230. ISBN 978-0-323-90274-8. [Google Scholar]
- Tafer, H.; Poyntner, C.; Lopandic, K.; Sterflinger, K.; Piñar, G. Back to the Salt Mines: Genome and Transcriptome Comparisons of the Halophilic Fungus Aspergillus salisburgensis and Its Halotolerant Relative Aspergillus sclerotialis. Genes 2019, 10, 381. [Google Scholar] [CrossRef]
- Albarano, L.; Esposito, R.; Ruocco, N.; Costantini, M. Genome Mining as New Challenge in Natural Products Discovery. Marine Drugs 2020, 18, 199. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L.; Vaishnav, P. Natural Products for Cancer Chemotherapy: Natural Products—Cancer. Microb. Biotechnol. 2011, 4, 687–699. [Google Scholar] [CrossRef]
- Wali, A.F.; Majid, S.; Rasool, S.; Shehada, S.B.; Abdulkareem, S.K.; Firdous, A.; Beigh, S.; Shakeel, S.; Mushtaq, S.; Akbar, I.; et al. Natural Products against Cancer: Review on Phytochemicals from Marine Sources in Preventing Cancer. SPJ 2019, 27, 767–777. [Google Scholar] [CrossRef]
- Ellenaa, V.; Steiger, M. The Good Fungus—About the Potential of Fungi for Our Future. In Good Microbes in Medicine, Food Production, Biotechnology, Bioremediation, and Agriculture; de Bruijn, F.J., Smidt, H., Cocolin, L.S., Sauer, M., Dowling, D., Thomashow, L., Eds.; Wiley: Hoboken, NJ, USA, 2022; pp. 287–293. ISBN 978-1-119-76254-6. [Google Scholar]
- Liu, T.; Zhang, S.; Zhu, J.; Pan, H.; Bai, J.; Li, Z.; Guan, L.; Liu, G.; Yuan, C.; Wu, X.; et al. Two New Amides from a Halotolerant Fungus, Myrothecium Sp. GS-17. J. Antibiot. 2015, 68, 267–270. [Google Scholar] [CrossRef]
- Conrado, R.; Gomes, T.C.; Roque, G.S.C.; De Souza, A.O. Overview of Bioactive Fungal Secondary Metabolites: Cytotoxic and Antimicrobial Compounds. Antibiotics 2022, 11, 1604. [Google Scholar] [CrossRef]
- Perrone, G.; Ferrara, M.; Medina, A.; Pascale, M.; Magan, N. Toxigenic Fungi and Mycotoxins in a Climate Change Scenario: Ecology, Genomics, Distribution, Prediction and Prevention of the Risk. Microorganisms 2020, 8, 1496. [Google Scholar] [CrossRef]
- Novak Babič, M.; Gunde-Cimerman, N.; Breskvar, M.; Džeroski, S.; Brandão, J. Occurrence, Diversity and Anti-Fungal Resistance of Fungi in Sand of an Urban Beach in Slovenia—Environmental Monitoring with Possible Health Risk Implications. J. Fungi 2022, 8, 860. [Google Scholar] [CrossRef]
- Sayed, A.M.; Hassan, M.H.A.; Alhadrami, H.A.; Hassan, H.M.; Goodfellow, M.; Rateb, M.E. Extreme Environments: Microbiology Leading to Specialized Metabolites. J. Appl. Microbiol. 2020, 128, 630–657. [Google Scholar] [CrossRef] [PubMed]
- Kuvarina, A.E.; Gavryushina, I.A.; Sykonnikov, M.A.; Efimenko, T.A.; Markelova, N.N.; Bilanenko, E.N.; Bondarenko, S.A.; Kokaeva, L.Y.; Timofeeva, A.V.; Serebryakova, M.V.; et al. Exploring Peptaibol’s Profile, Antifungal, and Antitumour Activity of Emericellipsin A of Emericellopsis Species from Soda and Saline Soils. Molecules 2022, 27, 1736. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, V.N.; Carvalho, C.R.; Johann, S.; Mendes, G.; Alves, T.M.A.; Zani, C.L.; Junior, P.A.S.; Murta, S.M.F.; Romanha, A.J.; Cantrell, C.L.; et al. Antibacterial, Antifungal and Antiprotozoal Activities of Fungal Communities Present in Different Substrates from Antarctica. Polar Biol. 2015, 38, 1143–1152. [Google Scholar] [CrossRef]
- Moghadamtousi, S.; Nikzad, S.; Kadir, H.; Abubakar, S.; Zandi, K. Potential Antiviral Agents from Marine Fungi: An Overview. Mar. Drugs 2015, 13, 4520–4538. [Google Scholar] [CrossRef]
- Rodríguez-Verdugo, A.; Lozano-Huntelman, N.; Cruz-Loya, M.; Savage, V.; Yeh, P. Compounding Effects of Climate Warming and Antibiotic Resistance. iScience 2020, 23, 101024. [Google Scholar] [CrossRef] [PubMed]
- Ebada, S.S.; Fischer, T.; Hamacher, A.; Du, F.-Y.; Roth, Y.O.; Kassack, M.U.; Wang, B.-G.; Roth, E.H. Psychrophilin E, a New Cyclotripeptide, from Co-Fermentation of Two Marine Alga-Derived Fungi of the Genus Aspergillus. Nat. Prod. Res. 2014, 28, 776–781. [Google Scholar] [CrossRef]
- Maheshwari, R.; Bharadwaj, G.; Bhat, M.K. Thermophilic Fungi: Their Physiology and Enzymes. Microbiol. Mol. Biol. Rev. 2000, 64, 461–488. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.K.; Chatterjee, S.; Gauri, S.S.; Das, S.S.; Mishra, A.; Patra, M.; Ghosh, A.K.; Das, A.K.; Singh, S.M.; Dey, S. Isolation and Characterization of Extracellular Polysaccharide Thelebolan Produced by a Newly Isolated Psychrophilic Antarctic Fungus Thelebolus. Carbohyd. Polym. 2014, 104, 204–212. [Google Scholar] [CrossRef]
- Hassan, N.; Rafiq, M.; Hayat, M.; Shah, A.A.; Hasan, F. Psychrophilic and Psychrotrophic Fungi: A Comprehensive Review. Rev. Environ. Sci. Biotechnol. 2016, 15, 147–172. [Google Scholar] [CrossRef]
- Li, L.; Li, D.; Luan, Y.; Gu, Q.; Zhu, T. Cytotoxic Metabolites from the Antarctic Psychrophilic Fungus Oidiodendron truncatum. J. Nat. Prod. 2012, 75, 920–927. [Google Scholar] [CrossRef]
- Amato, P.; Christner, B.C. Energy Metabolism Response to Low-Temperature and Frozen Conditions in Psychrobacter cryohalolentis. Appl. Environ. Microbiol. 2009, 75, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Gakuubi, M.M.; Munusamy, M.; Liang, Z.-X.; Ng, S.B. Fungal Endophytes: A Promising Frontier for Discovery of Novel Bioactive Compounds. J. Fungi 2021, 7, 786. [Google Scholar] [CrossRef] [PubMed]
- Ranawat, P.; Rawat, S. Radiation Resistance in Thermophiles: Mechanisms and Applications. World J. Microbiol. Biotechnol. 2017, 33, 112. [Google Scholar] [CrossRef] [PubMed]
- Conway, K.R.; Boddy, C.N. ClusterMine360: A Database of Microbial PKS/NRPS Biosynthesis. Nucleic Acids Res. 2012, 41, D402–D407. [Google Scholar] [CrossRef]
- Mehta, R.; Singhal, P.; Singh, H.; Damle, D.; Sharma, A.K. Insight into Thermophiles and Their Wide-Spectrum Applications. 3 Biotech 2016, 6, 81. [Google Scholar] [CrossRef]
- Elsakhawy, T. Marine Microbial Polysaccharides Environmental Role and Applications: An Overview. Environ. Biodivers. Soil Secur. 2017, 1, 61–70. [Google Scholar] [CrossRef]
- Tesei, D. Black Fungi Research: Out-of-This-World Implications. Encyclopedia 2022, 2, 212–229. [Google Scholar] [CrossRef]
- Oliveira, M.; Amorim, A. Microbial Forensics: New Breakthroughs and Future Prospects. Appl. Microbiol. Biotechnol. 2018, 102, 10377–10391. [Google Scholar] [CrossRef]
- Schipper, M.A.A. Mucor Strictus Hagem, a Psychrophilic Fungus, AndMucor Falcatus Sp.n. Antonie Van Leeuwenhoek 1967, 33, 189–195. [Google Scholar] [CrossRef]
- Wang, M.; Jiang, X.; Wu, W.; Hao, Y.; Su, Y.; Cai, L.; Xiang, M.; Liu, X. Psychrophilic Fungi from the World’s Roof. Persoonia 2015, 34, 100–112. [Google Scholar] [CrossRef]
- Wang, M.; Tian, J.; Xiang, M.; Liu, X. Living Strategy of Cold-Adapted Fungi with the Reference to Several Representative Species. Mycology 2017, 8, 178–188. [Google Scholar] [CrossRef]
- Dalsgaard, P.W.; Larsen, T.O.; Frydenvang, K.; Christophersen, C. Psychrophilin A and Cycloaspeptide D, Novel Cyclic Peptides from the Psychrotolerant Fungus Penicillium ribeum. J. Nat. Prod. 2004, 67, 878–881. [Google Scholar] [CrossRef] [PubMed]
- Kawata, K.; Iwai, A.; Muramatsu, D.; Aoki, S.; Uchiyama, H.; Okabe, M.; Hayakawa, S.; Takaoka, A.; Miyazaki, T. Stimulation of Macrophages with the β-Glucan Produced by Aureobasidium pullulans Promotes the Secretion of Tumour Necrosis Factor-Related Apoptosis Inducing Ligand (TRAIL). PLoS ONE 2015, 10, e0124809. [Google Scholar] [CrossRef] [PubMed]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J.C.; Terlouw, B.R.; van der Hooft, J.J.J.; van Santen, J.A.; Tracanna, V.; Suarez Duran, H.G.; Pascal Andreu, V.; et al. MIBiG 2.0: A Repository for Biosynthetic Gene Clusters of Known Function. Nucleic Acids Res. 2020, 48, D454–D458. [Google Scholar] [CrossRef] [PubMed]
- Farag, A.M.; Hassan, S.W.; Beltagy, E.A.; El-Shenawy, M.A. Optimization of Production of Anti-Tumour l-Asparaginase by Free and Immobilized Marine Aspergillus terreus. Egypt. J. Aquat. Res. 2015, 41, 295–302. [Google Scholar] [CrossRef]
- Zain ul Arifeen, M.; Ma, Y.-N.; Xue, Y.-R.; Liu, C.-H. Deep-Sea Fungi Could Be the New Arsenal for Bioactive Molecules. Mar. Drugs 2019, 18, 9. [Google Scholar] [CrossRef]
- Sajjad, W.; Din, G.; Rafiq, M.; Iqbal, A.; Khan, S.; Zada, S.; Ali, B.; Kang, S. Pigment Production by Cold-Adapted Bacteria and Fungi: Colorful Tale of Cryosphere with Wide Range Applications. Extremophiles 2020, 24, 447–473. [Google Scholar] [CrossRef]
- Peng, J.; Gao, H.; Zhang, X.; Wang, S.; Wu, C.; Gu, Q.; Guo, P.; Zhu, T.; Li, D. Psychrophilins E–H and Versicotide C, Cyclic Peptides from the Marine-Derived Fungus Aspergillus Versicolor ZLN-60. J. Nat. Prod. 2014, 77, 2218–2223. [Google Scholar] [CrossRef]
- Jiang, W.; Ye, P.; Chen, C.-T.; Wang, K.; Liu, P.; He, S.; Wu, X.; Gan, L.; Ye, Y.; Wu, B. Two Novel Hepatocellular Carcinoma Cycle Inhibitory Cyclodepsipeptides from a Hydrothermal Vent Crab-Associated Fungus Aspergillus Clavatus C2WU. Mar. Drugs 2013, 11, 4761–4772. [Google Scholar] [CrossRef]
- Giddings, L.-A.; Newman, D.J. Extremophilic Fungi from Marine Environments: Underexplored Sources of Antitumour, Anti-Infective and Other Biologically Active Agents. Mar. Drugs 2022, 20, 62. [Google Scholar] [CrossRef]
- Orfali, R.; Perveen, S. New Bioactive Metabolites from the Thermophilic Fungus Penicillium Sp. Isolated from Ghamiqa Hot Spring in Saudi Arabia. J. Chem. 2019, 2019, 7162948. [Google Scholar] [CrossRef]
- Zhang, X.; Li, S.-J.; Li, J.-J.; Liang, Z.-Z.; Zhao, C.-Q. Novel Natural Products from Extremophilic Fungi. Mar. Drugs 2018, 16, 194. [Google Scholar] [CrossRef] [PubMed]
- Dikmen, M.; Canturk, Z.; Engur, S.; Kaya Tilki, E. Inhibitory effects of secondary metabolites of halotolerant Aspergillus terreus on angiogenesis. Biomed. Res. 2017, 28, 3613–3618. [Google Scholar]
- Corral, P.; Amoozegar, M.A.; Ventosa, A. Halophiles and Their Biomolecules: Recent Advances and Future Applications in Biomedicine. Mar. Drugs 2019, 18, 33. [Google Scholar] [CrossRef]
- Park, H.B.; Kwon, H.C.; Lee, C.-H.; Yang, H.O. Glionitrin A, an Antibiotic−Antitumour Metabolite Derived from Competitive Interaction between Abandoned Mine Microbes. J. Nat. Prod. 2009, 72, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-L.; Liao, W.-Y.; Liu, W.-Y.; Liaw, C.-C.; Shen, C.-N.; Huang, Z.-Y.; Wu, S.-H. Discovery of New Natural Products by Intact-Cell Mass Spectrometry and LC-SPE-NMR: Malbranpyrroles, Novel Polyketides from Thermophilic Fungus Malbranchea sulfurea. Chem. Eur. J. 2009, 15, 11573–11580. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Nong, X.; Wang, J.; Qi, S. Brevianamides and Mycophenolic Acid Derivatives from the Deep-Sea-Derived Fungus Penicillium brevicompactum DFFSCS025. Mar. Drugs 2017, 15, 43. [Google Scholar] [CrossRef]
- Ichiye, T. Enzymes from Piezophiles. Semin. Cell Dev. Biol. 2018, 84, 138–146. [Google Scholar] [CrossRef]
- Caliò, A.; Dubois, C.; Fontanay, S.; Koza, M.M.; Hoh, F.; Roumestand, C.; Oger, P.; Peters, J. Unravelling the Adaptation Mechanisms to High Pressure in Proteins. Int. J. Mol. Sci. 2022, 23, 8469. [Google Scholar] [CrossRef]
- Luo, X.; Zhou, X.; Lin, X.; Qin, X.; Zhang, T.; Wang, J.; Tu, Z.; Yang, B.; Liao, S.; Tian, Y.; et al. Antituberculosis Compounds from a Deep-Sea-Derived Fungus Aspergillus Sp. SCSIO Ind09F01. Nat. Prod. Res. 2017, 31, 1958–1962. [Google Scholar] [CrossRef]
- Tawfike, A.F.; Viegelmann, C.; Edrada-Ebel, R. Metabolomics and Dereplication Strategies in Natural Products. Methods Mol. Biol. 2013, 1055, 227–244. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P. Fungal Secondary Metabolism: Regulation, Function and Drug Discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Bernan, V.S.; Greenstein, M.; Maiese, W.M. Marine Microorganisms as a Source of New Natural Products. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 1997; Volume 43, pp. 57–90. ISBN 978-0-12-002643-2. [Google Scholar]
- Yang, X.L.; Chen, Y.C.; Li, H.H.; Zhang, W.M. Molecular identification of 23 marine fungal strains and their activties against plant pathogenic fungi and cytotoxic activities. Biotechnol. Bull. 2014, 8, 131–137. [Google Scholar]
- Olatinwo, R.; So, C.-L.; Eberhardt, T.L. Effect of Acaromyces Ingoldii Secondary Metabolites on the Growth of Brown-Rot (Gloeophyllum Trabeum) and White-Rot (Trametes Versicolor) Fungi. Mycobiology 2019, 47, 506–511. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J. Natural Products from Marine Fungi—Still an Underrepresented Resource. Mar. Drugs 2016, 14, 19. [Google Scholar] [CrossRef]
- Ndwigah, F.I.; Boga, H.I.; Wanyoike, W.; Kachiuri, R. Characterization, Enzymatic activity, and secondary metabolites of fungal isolates from lake Sonachi in Kenya. IOSR J. Pharm. Biol. Sci. (IOSR-JPBS) 2015, 10, 65–76. [Google Scholar]
- Burgaud, G.; Le Calvez, T.; Arzur, D.; Vandenkoornhuyse, P.; Barbier, G. Diversity of Culturable Marine Filamentous Fungi from Deep-Sea Hydrothermal Vents. Environ. Microbiol. 2009, 11, 1588–1600. [Google Scholar] [CrossRef]
- Cong, M.; Pang, X.; Zhao, K.; Song, Y.; Liu, Y.; Wang, J. Deep-Sea Natural Products from Extreme Environments: Cold Seeps and Hydrothermal Vents. Mar. Drugs 2022, 20, 404. [Google Scholar] [CrossRef]
- Liu, F.; Lin, X.; Zhou, X.; Chen, M.; Huang, X.; Yang, B.; Tao, H. Xanthones and Quinolones Derivatives Produced by the Deep-Sea-Derived Fungus Penicillium Sp. SCSIO Ind16F01. Molecules 2017, 22, 1999. [Google Scholar] [CrossRef]
- Zheng, J.; Wang, Y.; Wang, J.; Liu, P.; Li, J.; Zhu, W. Antimicrobial Ergosteroids and Pyrrole Derivatives from Halotolerant Aspergillus Flocculosus PT05-1 Cultured in a Hypersaline Medium. Extremophiles 2013, 17, 963–971. [Google Scholar] [CrossRef]
- Barone, G.; Varrella, S.; Tangherlini, M.; Rastelli, E.; Dell’Anno, A.; Danovaro, R.; Corinaldesi, C. Marine Fungi: Biotechnological Perspectives from Deep-Hypersaline Anoxic Basins. Diversity 2019, 11, 113. [Google Scholar] [CrossRef]
- Enache, M.; Cojoc, R.; Kamekura, M. Halophilic Microorganisms and Their Biomolecules: Approaching into Frame of Bio(Nano) Technologies. In Halophiles; Maheshwari, D.K., Saraf, M., Eds.; Sustainable Development and Biodiversity; Springer International Publishing: Cham, Switzerland, 2015; Volume 6, pp. 161–172. ISBN 978-3-319-14594-5. [Google Scholar]
- Fredimoses, M.; Zhou, X.; Ai, W.; Tian, X.; Yang, B.; Lin, X.; Xian, J.-Y.; Liu, Y. Westerdijkin A, a New Hydroxyphenylacetic Acid Derivative from Deep Sea Fungus Aspergillus Westerdijkiae SCSIO 05233. Nat. Prod. Res. 2015, 29, 158–162. [Google Scholar] [CrossRef]
- Mamo, G.; Mattiasson, B. (Eds.) Alkaliphiles in Biotechnology; Advances in Biochemical Engineering/Biotechnology; Springer International Publishing: Cham, Switzerland, 2020; Volume 172, ISBN 978-3-030-49735-4. [Google Scholar]
- Stierle, A.A.; Stierle, D.B. Secondary Metabolites of Mine Waste Acidophilic Fungi. In Bioprospecting; Paterson, R., Lima, N., Eds.; Topics in Biodiversity and Conservation; Springer International Publishing: Cham, Switzerland, 2017; Volume 16, pp. 213–243. ISBN 978-3-319-47933-0. [Google Scholar]
- Rogozhin, E.A.; Sadykova, V.S.; Baranova, A.A.; Vasilchenko, A.S.; Lushpa, V.A.; Mineev, K.S.; Georgieva, M.L.; Kul’ko, A.B.; Krasheninnikov, M.E.; Lyundup, A.V.; et al. A Novel Lipopeptaibol Emericellipsin A with Antimicrobial and Antitumour Activity Produced by the Extremophilic Fungus Emericellopsis alkalina. Molecules 2018, 23, 2785. [Google Scholar] [CrossRef]
- Kuvarina, A.E.; Gavryushina, I.A.; Kulko, A.B.; Ivanov, I.A.; Rogozhin, E.A.; Georgieva, M.L.; Sadykova, V.S. The Emericellipsins A–E from an Alkalophilic Fungus Emericellopsis alkalina Show Potent Activity against Multidrug-Resistant Pathogenic Fungi. J. Fungi 2021, 7, 153. [Google Scholar] [CrossRef]
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Debets, A.J.M.; Bilanenko, E.N. Are Alkalitolerant Fungi of the Emericellopsis Lineage (Bionectriaceae) of Marine Origin? IMA Fungus 2013, 4, 213–228. [Google Scholar] [CrossRef]
- Kuvarina, A.E.; Rogozhin, E.A.; Sykonnikov, M.A.; Timofeeva, A.V.; Serebryakova, M.V.; Fedorova, N.V.; Kokaeva, L.Y.; Efimenko, T.A.; Georgieva, M.L.; Sadykova, V.S. Isolation and Characterization of a Novel Hydrophobin, Sa-HFB1, with Antifungal Activity from an Alkaliphilic Fungus, Sodiomyces alkalinus. J. Fungi 2022, 8, 659. [Google Scholar] [CrossRef] [PubMed]
- Stierle, D.B.; Stierle, A.A.; Girtsman, T.; McIntyre, K.; Nichols, J. Caspase-1 and -3 Inhibiting Drimane Sesquiterpenoids from the Extremophilic Fungus Penicillium solitum. J. Nat. Prod. 2012, 75, 262–266. [Google Scholar] [CrossRef]
- Stierle, A.A.; Stierle, D.B.; Kemp, K. Novel Sesquiterpenoid Matrix Metalloproteinase-3 Inhibitors from an Acid Mine Waste Extremophile. J. Nat. Prod. 2004, 67, 1392–1395. [Google Scholar] [CrossRef] [PubMed]
- Caruso, F.; Pedersen, J.Z.; Incerpi, S.; Kaur, S.; Belli, S.; Florea, R.-M.; Rossi, M. Mechanism of Caspase-1 Inhibition by Four Anti-Inflammatory Drugs Used in COVID-19 Treatment. Int. J. Mol. Sci. 2022, 23, 1849. [Google Scholar] [CrossRef]
- Winer, A.; Adams, S.; Mignatti, P. Matrix Metalloproteinase Inhibitors in Cancer Therapy: Turning Past Failures into Future Successes. Mol. Cancer Ther. 2018, 17, 1147–1155. [Google Scholar] [CrossRef]
- Hidalgo, M.; Eckhardt, S.G. Development of Matrix Metalloproteinase Inhibitors in Cancer Therapy. JNCI 2001, 93, 178–193. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.A.; Stierle, D.B.; Kelly, K. Berkelic Acid, A Novel Spiroketal with Selective Anticancer Activity from an Acid Mine Waste Fungal Extremophile. J. Org. Chem. 2006, 71, 5357–5360. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.A.; Stierle, D.B. Bioactive Secondary Metabolites from Acid Mine Waste Extremophiles. Nat. Prod. Commun. 2014, 9, 1037–1044. [Google Scholar] [CrossRef]
- Liu, S.; Su, M.; Song, S.-J.; Jung, J. Marine-Derived Penicillium Species as Producers of Cytotoxic Metabolites. Mar. Drugs 2017, 15, 329. [Google Scholar] [CrossRef]
- Stierle, D.B.; Stierle, A.A.; Hobbs, J.D.; Stokken, J.; Clardy, J. Berkeleydione and Berkeleytrione, New Bioactive Metabolites from an Acid Mine Organism. Org. Lett. 2004, 6, 1049–1052. [Google Scholar] [CrossRef]
- Ma, L.; Tai, H.; Li, C.; Zhang, Y.; Wang, Z.-H.; Ji, W.-Z. Photodynamic Inhibitory Effects of Three Perylenequinones on Human Colorectal Carcinoma Cell Line and Primate Embryonic Stem Cell Line. World J. Gastroenterol. 2003, 9, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Ngokpol, S.; Suwakulsiri, W.; Sureram, S.; Lirdprapamongkol, K.; Aree, T.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Drimane Sesquiterpene-Conjugated Amino Acids from a Marine Isolate of the Fungus Talaromyces Minioluteus (Penicillium Minioluteum). Mar. Drugs 2015, 13, 3567–3580. [Google Scholar] [CrossRef]
- Stierle, A.A.; Stierle, D.B.; Girtsman, T. Caspase-1 Inhibitors from an Extremophilic Fungus That Target Specific Leukemia Cell Lines. J. Nat. Prod. 2012, 75, 344–350. [Google Scholar] [CrossRef]
- Pettit, R.K. Small-Molecule Elicitation of Microbial Secondary Metabolites: Elicitation of Microbial Secondary Metabolites. Microb. Biotechnol. 2011, 4, 471–478. [Google Scholar] [CrossRef]
- Stierle, A.A.; Stierle, D.B. Bioprospecting in the Berkeley Pit. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2013; Volume 39, pp. 1–45. ISBN 978-0-444-62615-8. [Google Scholar]
- Dieter, A.; Hamm, A.; Fiedler, H.-P.; Goodfellow, M.; Müller, W.E.G.; Brun, R.; Beil, W.; Bringmann, G. Pyrocoll, an Antibiotic, Antiparasitic and Antitumour Compound Produced by a Novel Alkaliphilic Streptomyces Strain. J. Antibiot. 2003, 56, 639–646. [Google Scholar] [CrossRef]
- Ma, M.; Ge, H.; Yi, W.; Wu, B.; Zhang, Z. Bioactive Drimane Sesquiterpenoids and Isocoumarins from the Marine-Derived Fungus Penicillium Minioluteum ZZ1657. Tetrahedron Lett. 2020, 61, 151504. [Google Scholar] [CrossRef]
- Rydholm, C.; Szakacs, G.; Lutzoni, F. Low Genetic Variation and No Detectable Population Structure in Aspergillus fumigatus Compared to Closely Related Neosartorya Species. Eukaryot Cell 2006, 5, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Wu, G.; Gu, Q.; Zhu, T.; Li, D. New Eremophilane-Type Sesquiterpenes from an Antarctic Deepsea Derived Fungus, Penicillium Sp. PR19 N-1. Arch. Pharm. Res. 2014, 37, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Wu, Q.; Yu, Y.; Liang, Z.; Liu, Y.; Zhou, L.; Tang, L.; Zhou, X. Penicilliumin B, a Novel Sesquiterpene Methylcyclopentenedione from a Deep Sea-Derived Penicillium Strain with Renoprotective Activities. Sci. Rep. 2017, 7, 10757. [Google Scholar] [CrossRef] [PubMed]
- Stierle, D.B.; Stierle, A.A.; Patacini, B. The Berkeleyacetals, Three Meroterpenes from a Deep Water Acid Mine Waste Penicillium. J. Nat. Prod. 2007, 70, 1820–1823. [Google Scholar] [CrossRef] [PubMed]
- Kornienko, A.; Evidente, A.; Vurro, M.; Mathieu, V.; Cimmino, A.; Evidente, M.; van Otterlo, W.A.L.; Dasari, R.; Lefranc, F.; Kiss, R. Toward a Cancer Drug of Fungal Origin. Med. Res. Rev. 2015, 35, 937–967. [Google Scholar] [CrossRef]
- Tiquia-Arashiro, S.M.; Grube, M. (Eds.) Fungi in Extreme Environments: Ecological Role and Biotechnological Significance; Springer: Cham, Switzerland; Berlin/Heidelberg, Germany, 2019; ISBN 978-3-030-19029-3. [Google Scholar]
- Steward, W.P.; Brown, K. Cancer Chemoprevention: A Rapidly Evolving Field. Br. J. Cancer. 2013, 109, 1–7. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Ramos, J.; Plemenitaš, A. Halotolerant and Halophilic Fungi. Mycolo. Res. 2009, 113, 1231–1241. [Google Scholar] [CrossRef]
- Montalvo-Rodríguez, R.; Maupin-Furlow, J.A. Insights through Genetics of Halophilic Microorganisms and Their Viruses. Genes 2020, 11, 388. [Google Scholar] [CrossRef]
- Sanchez, S.; Demain, A.L. Bioactive Products from Fungi. In Food Bioactives; Puri, M., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 59–87. ISBN 978-3-319-51637-0. [Google Scholar]
- Zajc, J.; Kogej, T.; Galinski, E.A.; Ramos, J.; Gunde-Cimerman, N. Osmoadaptation Strategy of the Most Halophilic Fungus, Wallemia ichthyophaga, Growing Optimally at Salinities above 15% NaCl. Appl. Environ. Microbiol. 2014, 80, 247–256. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Zalar, P. Extremely Halotolerant and Halophilic Fungi Inhabit Brine in Solar Salterns Around the Globe, Fungi in Solar Salterns. Food Technol. Biotechnol. 2014, 52, 170–179. [Google Scholar]
- Śliżewska, W.; Struszczyk-Świta, K.; Marchut-Mikołajczyk, O. Metabolic Potential of Halophilic Filamentous Fungi—Current Perspective. Int. J. Mol. Sci. 2022, 23, 4189. [Google Scholar] [CrossRef] [PubMed]
- Venil, C.K.; Velmurugan, P.; Dufossé, L.; Renuka Devi, P.; Veera Ravi, A. Fungal Pigments: Potential Coloring Compounds for Wide Ranging Applications in Textile Dyeing. J. Fungi 2020, 6, 68. [Google Scholar] [CrossRef] [PubMed]
- Ruginescu, R.; Gomoiu, I.; Popescu, O.; Cojoc, R.; Neagu, S.; Lucaci, I.; Batrinescu-Moteau, C.; Enache, M. Bioprospecting for Novel Halophilic and Halotolerant Sources of Hydrolytic Enzymes in Brackish, Saline and Hypersaline Lakes of Romania. Microorganisms 2020, 8, 1903. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Galinski, E.A.; Grant, W.D.; Oren, A.; Ventosa, A. Halophiles 2010: Life in Saline Environments. Appl. Environ. Microbiol. 2010, 76, 6971–6981. [Google Scholar] [CrossRef]
- Sepcic, K.; Zalar, P.; Gunde-Cimerman, N. Low Water Activity Induces the Production of Bioactive Metabolites in Halophilic and Halotolerant Fungi. Mar. Drugs 2010, 9, 43–58. [Google Scholar] [CrossRef]
- Konte, T.; Terpitz, U.; Plemenitaš, A. Reconstruction of the High-Osmolarity Glycerol (HOG) Signaling Pathway from the Halophilic Fungus Wallemia ichthyophaga in Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 1821. [Google Scholar] [CrossRef]
- Méjanelle, L.; Lòpez, J.F.; Gunde-Cimerman, N.; Grimalt, J.O. Ergosterol Biosynthesis in Novel Melanized Fungi from Hypersaline Environments. J. Lipid. Res. 2001, 42, 352–358. [Google Scholar] [CrossRef]
- Peng, J.; Zhang, X.; Du, L.; Wang, W.; Zhu, T.; Gu, Q.; Li, D. Sorbicatechols A and B, Antiviral Sorbicillinoids from the Marine-Derived Fungus Penicillium chrysogenum PJX-17. J. Nat. Prod. 2014, 77, 424–428. [Google Scholar] [CrossRef]
- Ruiz-Ruiz, C.; Srivastava, G.K.; Carranza, D.; Mata, J.A.; Llamas, I.; Santamaría, M.; Quesada, E.; Molina, I.J. An Exopolysaccharide Produced by the Novel Halophilic Bacterium Halomonas Stenophila Strain B100 Selectively Induces Apoptosis in Human T Leukaemia Cells. Appl. Microbiol. Biotechnol. 2011, 89, 345–355. [Google Scholar] [CrossRef]
- Yu, X.; Yang, R.; Gu, Z.; Lai, S.; Yang, H. Anti-Tumour and Immunostimulatory Functions of Two Feruloyl Oligosaccharides Produced from Wheat Bran and Fermented by Aureobasidium pullulans. BioResources 2014, 9, 6778–6790. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, X.; Kwak, M.; Xu, L.; Zhang, L.; Yu, Q.; Jin, J.-O. Maturation of Dendritic Cells by Pullulan Promotes Anti-Cancer Effect. Oncotarget 2016, 7, 44644–44659. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Kusano, K.; Kondo, N.; Nishikawa, K.; Kuge, T.; Ohno, N. Biological Activity of High-Purity β-1,3-1,6-Glucan Derived from the Black Yeast Aureobasidium pullulans: A Literature Review. Nutrients 2021, 13, 242. [Google Scholar] [CrossRef] [PubMed]
- Manitchotpisit, P.; Watanapoksin, R.; Price, N.P.J.; Bischoff, K.M.; Tayeh, M.; Teeraworawit, S.; Kriwong, S.; Leathers, T.D. Aureobasidium pullulans as a Source of Liamocins (Heavy Oils) with Anticancer Activity. World J. Microbiol. Biotechnol. 2014, 30, 2199–2204. [Google Scholar] [CrossRef]
- Muramatsu, D.; Iwai, A.; Aoki, S.; Uchiyama, H.; Kawata, K.; Nakayama, Y.; Nikawa, Y.; Kusano, K.; Okabe, M.; Miyazaki, T. β-Glucan Derived from Aureobasidium pullulans Is Effective for the Prevention of Influenza in Mice. PLoS ONE 2012, 7, e41399. [Google Scholar] [CrossRef]
- Yujiang, F.; Huanan, L.; Yong, S.; Jie, L. Natural Polysaccharide Pullulan as Anti-Cancer Drug Carriers for Hepatic Targeting. Front. Bioeng. Biotechnol. 2016, 4, 349. [Google Scholar] [CrossRef]
- Gibbs, P.A.; Seviour, R.J. The Production of Exopolysaccharides by Aureobasidium pullulans in Fermenters with Low-Shear Configurations. Appl. Microbiol. Biotechnol. 1998, 49, 168–174. [Google Scholar] [CrossRef]
- Cheng, K.-C.; Demirci, A.; Catchmark, J.M. Pullulan: Biosynthesis, Production, and Applications. Appl. Microbiol. Biotechnol. 2011, 92, 29–44. [Google Scholar] [CrossRef]
- He, X.; Abrams, S.I.; Lovell, J.F. Peptide Delivery Systems for Cancer Vaccines. Adv. Therap. 2018, 1, 1800060. [Google Scholar] [CrossRef]
- Dardiotis, E.; Aloizou, A.-M.; Markoula, S.; Siokas, V.; Tsarouhas, K.; Tzanakakis, G.; Libra, M.; Kyritsis, A.P.; Brotis, A.G.; Aschner, M.; et al. Cancer-Associated Stroke: Pathophysiology, Detection and Management (Review). Int. J. Oncol. 2019, 54, 779–796. [Google Scholar] [CrossRef] [PubMed]
- Stierle, A.A.; Stierle, D.B.; Girtsman, T.; Mou, T.C.; Antczak, C.; Djaballah, H. Azaphilones from an Acid Mine Extremophile Strain of a Pleurostomophora Sp. J. Nat. Prod. 2015, 78, 2917–2923. [Google Scholar] [CrossRef]
- Liu, P.P.; Wang, W.L.; Gu, Q.Q.; Zhu, W.M. Alkyl Benzaldehyde derivatives and their cytotoxicities from Halotolerant Fungus Aspergillus variecolor B-17. Period. Ocean Univ. China 2008, 38, 585–589. [Google Scholar]
- Golemis, E.A.; Scheet, P.; Beck, T.N.; Scolnick, E.M.; Hunter, D.J.; Hawk, E.; Hopkins, N. Molecular Mechanisms of the Preventable Causes of Cancer in the United States. Genes Dev. 2018, 32, 868–902. [Google Scholar] [CrossRef]
- Liu, T.; Zhu, J.; Zhang, S.-Y.; Li, Z.-L.; Guan, L.-P.; Pan, H.-Q.; Wu, X.; Bai, J.; Hua, H.-M. Polyketides from the Halotolerant Fungus Myrothecium Sp. GS-17. Molecules 2013, 18, 15126–15133. [Google Scholar] [CrossRef] [PubMed]
- Rajpal, K.; Prasad, R.; Gupta, V.K.; Varma, A. Isolation and Characterization of Halophilic Soil Fungi from the Salt Desert of Little Rann of Kutch, India. PeerJ Prepr. 2016. [Google Scholar] [CrossRef]
- Horton, D.; Baker, D.C. Advances in Carbohydrate Chemistry and Biochemistry, 1st ed.; Academic Press: Oxford, UK, 2012; ISBN 978-0-12-396523-3. [Google Scholar]
- Selvasekaran, P.; Mahalakshmi; Roshini, F.; Angalene, L.A.; Chandini; Sunil, T.; Chidambaram, R. Fungal Exopolysaccharides: Production and Biotechnological Industrial Applications in Food and Allied Sectors. In Recent Trends in Mycological Research; Yadav, A.N., Ed.; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2021; pp. 311–357. ISBN 978-3-030-68259-0. [Google Scholar]
- Chung, D.; Kim, H.; Choi, H.S. Fungi in Salterns. J. Microbiol. 2019, 57, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Mohankumari, H.P.; Naidu, K.A.; Narasimhamurthy, K.; Vijayalakshmi, G. Bioactive Pigments of Monascus Purpureus Attributed to Antioxidant, HMG-CoA Reductase Inhibition and Anti-Atherogenic Functions. Front. Sustain. Food Syst. 2021, 5, 590427. [Google Scholar] [CrossRef]
- Liu, J.; Du, Y.; Ma, H.; Pei, X.; Li, M. Enhancement of Monascus Yellow Pigments Production by Activating the CAMP Signalling Pathway in Monascus Purpureus HJ11. Microb. Cell Fact. 2020, 19, 224. [Google Scholar] [CrossRef]
- Yang, T.; Liu, J.; Luo, F.; Lin, Q.; Rosol, T.J.; Deng, X. Anticancer Properties of Monascus Metabolites. Anti-Cancer Drugs 2014, 25, 735–744. [Google Scholar] [CrossRef]
- Kim, D.; Ku, S. Beneficial Effects of Monascus Sp. KCCM 10093 Pigments and Derivatives: A Mini Review. Molecules 2018, 23, 98. [Google Scholar] [CrossRef]
- Akihisa, T.; Tokuda, H.; Ukiya, M.; Kiyota, A.; Yasukawa, K.; Sakamoto, N.; Kimura, Y.; Suzuki, T.; Takayasu, J.; Nishino, H. Anti-Tumour-Initiating Effects of Monascin, an Azaphilonoid Pigment from the Extract of Monascus pilosus Fermented Rice (Red-Mold Rice). Chem. Biodivers. 2005, 2, 1305–1309. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, A.C.; Ogawa, C.Y.; De Camillis Rodrigues, L.; de Medeiros, L.S.; Veiga, T.A.M. Penicillium Genus as a Source for Anti-Leukemia Compounds: An Overview from 1984 to 2020. Leuk. Lymphoma 2021, 62, 2079–2093. [Google Scholar] [CrossRef] [PubMed]
- Leitão, A.L. Potential of Penicillium Species in the Bioremediation Field. Int. J. Environ. Res. Public Health 2009, 6, 1393–1417. [Google Scholar] [CrossRef]
- Ting, C.P.; Xu, G.; Zeng, X.; Maimone, T.J. Annulative Methods Enable a Total Synthesis of the Complex Meroterpene Berkeleyone A. J. Am. Chem. Soc. 2016, 138, 14868–14871. [Google Scholar] [CrossRef] [PubMed]
- Lou, J.; Fu, L.; Peng, Y.; Zhou, L. Metabolites from Alternaria Fungi and Their Bioactivities. Molecules 2013, 18, 5891–5935. [Google Scholar] [CrossRef]
- Weber, T.; Kim, H.U. The Secondary Metabolite Bioinformatics Portal: Computational Tools to Facilitate Synthetic Biology of Secondary Metabolite Production. Synth. Syst. Biotechnol. 2016, 1, 69–79. [Google Scholar] [CrossRef]
- Jansson, J.K.; Baker, E.S. A Multi-Omic Future for Microbiome Studies. Nat. Microbiol. 2016, 1, 16049. [Google Scholar] [CrossRef]
- Medema, M.H.; Fischbach, M.A. Computational Approaches to Natural Product Discovery. Nat. Chem. Biol. 2015, 11, 639–648. [Google Scholar] [CrossRef]
- Chen, R.; Wong, H.L.; Burns, B.P. New Approaches to Detect Biosynthetic Gene Clusters in the Environment. Medicines 2019, 6, 32. [Google Scholar] [CrossRef]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. AntiSMASH: Rapid Identification, Annotation and Analysis of Secondary Metabolite Biosynthesis Gene Clusters in Bacterial and Fungal Genome Sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef]
- Segura, V.; Descauses, M.C.; Charpentier, J.P.; Kinkel, K.; Mandin, C.; Ader, K.; Belmokhtar, N.; Boizot, N.; Buret, C.; Dejardin, A.; et al. A systems biology approach for identifying candidate genes involved in the natural variability of biomass yield and chemical properties in black poplar. In Proceedings of the IUFRO Genomics and Forest Tree Genetics, Arcachon, France, 30 May–3 June 2016; IUFRO Genomics and Forest Tree Genetics. Book of Abstract. p. 37. [Google Scholar]
- Pott, D.M.; Osorio, S.; Vallarino, J.G. From Central to Specialized Metabolism: An Overview of Some Secondary Compounds Derived from the Primary Metabolism for Their Role in Conferring Nutritional and Organoleptic Characteristics to Fruit. Front. Plant. Sci. 2019, 10, 835. [Google Scholar] [CrossRef] [PubMed]
- Gacek, A.; Strauss, J. The Chromatin Code of Fungal Secondary Metabolite Gene Clusters. Appl. Microbiol. Biotechnol. 2012, 95, 1389–1404. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal Secondary Metabolism—From Biochemistry to Genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Mortensen, U.H.; Vesth, T.; Andersen, M.R. Strategies to Establish the Link between Biosynthetic Gene Clusters and Secondary Metabolites. Fungal Genet. Biol. 2019, 130, 107–121. [Google Scholar] [CrossRef]
- Bills, G.F.; Gloer, J.B. Biologically Active Secondary Metabolites from the Fungi. Microbiol. Spectr. 2016, 4, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Baral, B.; Akhgari, A.; Metsä-Ketelä, M. Activation of Microbial Secondary Metabolic Pathways: Avenues and Challenges. Synt. Syst. Biotechnol. 2018, 3, 163–178. [Google Scholar] [CrossRef]
- Ye, J.-W.; Lin, Y.-N.; Yi, X.-Q.; Yu, Z.-X.; Liu, X.; Chen, G.-Q. Synthetic Biology of Extremophiles: A New Wave of Biomanufacturing. Trend. Biotechnol. 2022, 41, 342–357. [Google Scholar] [CrossRef]
- Theobald, S.; Vesth, T.C.; Rendsvig, J.K.; Nielsen, K.F.; Riley, R.; de Abreu, L.M.; Salamov, A.; Frisvad, J.C.; Larsen, T.O.; Andersen, M.R.; et al. Uncovering Secondary Metabolite Evolution and Biosynthesis Using Gene Cluster Networks and Genetic Dereplication. Sci. Rep. 2018, 8, 17957. [Google Scholar] [CrossRef]
- Skinnider, M.A.; Johnston, C.W.; Gunabalasingam, M.; Merwin, N.J.; Kieliszek, A.M.; MacLellan, R.J.; Li, H.; Ranieri, M.R.M.; Webster, A.L.H.; Cao, M.P.T.; et al. Comprehensive Prediction of Secondary Metabolite Structure and Biological Activity from Microbial Genome Sequences. Nat. Commun. 2020, 11, 6058. [Google Scholar] [CrossRef]
- Nielsen, J.C.; Nielsen, J. Development of Fungal Cell Factories for the Production of Secondary Metabolites: Linking Genomics and Metabolism. Synth. Syst. Biotechnol. 2017, 2, 5–12. [Google Scholar] [CrossRef]
- Kessler, S.C.; Chooi, Y.-H. Out for a RiPP: Challenges and Advances in Genome Mining of Ribosomal Peptides from Fungi. Nat. Prod. Rep. 2022, 39, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Hao, T.; Xie, Z.; Horsman, G.P.; Chen, Y. Genome Mining Unveils Widespread Natural Product Biosynthetic Capacity in Human Oral Microbe Streptococcus Mutans. Sci. Rep. 2016, 6, 37479. [Google Scholar] [CrossRef]
- Chen, Y.; Smanski, M.J.; Shen, B. Improvement of Secondary Metabolite Production in Streptomyces by Manipulating Pathway Regulation. Appl. Microb. Biotechnol. 2010, 86, 19–25. [Google Scholar] [CrossRef]
- Batista, T.M.; Hilario, H.O.; de Brito, G.A.M.; Moreira, R.G.; Furtado, C.; de Menezes, G.C.A.; Rosa, C.A.; Rosa, L.H.; Franco, G.R. Whole-Genome Sequencing of the Endemic Antarctic Fungus Antarctomyces pellizariae Reveals an Ice-Binding Protein, a Scarce Set of Secondary Metabolites Gene Clusters and Provides Insights on Thelebolales Phylogeny. Genomics 2020, 112, 2915–2921. [Google Scholar] [CrossRef] [PubMed]
- Pickens, L.B.; Tang, Y.; Chooi, Y.-H. Metabolic Engineering for the Production of Natural Products. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 211–236. [Google Scholar] [CrossRef]
- Mohimani, H.; Gurevich, A.; Mikheenko, A.; Garg, N.; Nothias, L.-F.; Ninomiya, A.; Takada, K.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of Peptidic Natural Products through Database Search of Mass Spectra. Nat. Chem. Biol. 2017, 13, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Mohimani, H.; Gurevich, A.; Shlemov, A.; Mikheenko, A.; Korobeynikov, A.; Cao, L.; Shcherbin, E.; Nothias, L.-F.; Dorrestein, P.C.; Pevzner, P.A. Dereplication of Microbial Metabolites through Database Search of Mass Spectra. Nat. Commun. 2018, 9, 4035. [Google Scholar] [CrossRef] [PubMed]
- Hodkinson, T.R.; Parnell, J.A. Reconstructing the Tree of Life: Taxonomy and Systematics of Species Rich Taxa; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Rangel, D.E.N.; Alder-Rangel, A.; Dadachova, E.; Finlay, R.D.; Kupiec, M.; Dijksterhuis, J.; Braga, G.U.L.; Corrochano, L.M.; Hallsworth, J.E. Fungal Stress Biology: A Preface to the Fungal Stress Responses Special Edition. Curr. Genet. 2015, 61, 231–238. [Google Scholar] [CrossRef]
- El-Elimat, T.; Ehrmann, B.; Cech, N.; Pearce, C.; Oberlies, N. De-Inventing the Wheel: Dereplication Tools for Natural Products Research. Planta Med. 2012, 78, PJ68. [Google Scholar] [CrossRef]
- Vivek-Ananth, R.P.; Sahoo, A.K.; Kumaravel, K.; Mohanraj, K.; Samal, A. MeFSAT: A Curated Natural Product Database Specific to Secondary Metabolites of Medicinal Fungi. RSC Adv. 2021, 11, 2596–2607. [Google Scholar] [CrossRef]
- Wolf, T.; Shelest, V.; Nath, N.; Shelest, E. CASSIS and SMIPS: Promoter-Based Prediction of Secondary Metabolite Gene Clusters in Eukaryotic Genomes. Bioinformatics 2016, 32, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Kim, H.U.; Medema, M.H.; Weber, T. Recent Development of AntiSMASH and Other Computational Approaches to Mine Secondary Metabolite Biosynthetic Gene Clusters. Brief. Bioinform. 2019, 20, 1103–1113. [Google Scholar] [CrossRef]
- Noar, R.D.; Daub, M.E. Bioinformatics Prediction of Polyketide Synthase Gene Clusters from Mycosphaerella Fijiensis. PLoS ONE 2016, 11, e0158471. [Google Scholar] [CrossRef]
- Kloosterman, A.M.; Medema, M.H.; van Wezel, G.P. Omics-Based Strategies to Discover Novel Classes of RiPP Natural Products. Curr. Opin. Biotechnol. 2021, 69, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Chaitankar, V.; Karakülah, G.; Ratnapriya, R.; Giuste, F.O.; Brooks, M.J.; Swaroop, A. Next Generation Sequencing Technology and Genomewide Data Analysis: Perspectives for Retinal Research. Prog. Retin. 2016, 55, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, B.R.; Khadayat, K.; Aryal, N.; Aryal, B.; Lamichhane, U.; Bhattarai, K.; Rana, N.; Regmi, B.P.; Adhikari, A.; Thapa, S.; et al. Untargeted Metabolomics of Streptomyces Species Isolated from Soils of Nepal. Processes 2022, 10, 1173. [Google Scholar] [CrossRef]
- Noar, R.D.; Daub, M.E. Transcriptome Sequencing of Mycosphaerella Fijiensis during Association with Musa Acuminata Reveals Candidate Pathogenicity Genes. BMC Genomics 2016, 17, 690. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.S.; Vetsigian, K.H. DesignSignatures: A Tool for Designing Primers That Yields Amplicons with Distinct Signatures. Bioinformatics 2016, 32, 1565–1567. [Google Scholar] [CrossRef]
- Rogozin, I.B.; Pavlov, Y.I.; Goncearenco, A.; De, S.; Lada, A.G.; Poliakov, E.; Panchenko, A.R.; Cooper, D.N. Mutational Signatures and Mutable Motifs in Cancer Genomes. Brief. Bioinform. 2017, 19, 1085–1101. [Google Scholar] [CrossRef]
- Disegha, G.C.; Akani, N.P. Bioinformatic Method for Fungi Identification. J. Biol. Gen. Res. 2019, 5, 1–5. [Google Scholar]
- Zierep, P.F.; Padilla, N.; Yonchev, D.G.; Telukunta, K.K.; Klementz, D.; Günther, S. SeMPI: A Genome-Based Secondary Metabolite Prediction and Identification Web Server. Nucleic Acids Res. 2017, 45, W64–W71. [Google Scholar] [CrossRef]
- Sousa, F.; Ferreira, D.; Reis, S.; Costa, P. Current Insights on Antifungal Therapy: Novel Nanotechnology Approaches for Drug Delivery Systems and New Drugs from Natural Sources. Pharmaceuticals 2020, 13, 248. [Google Scholar] [CrossRef] [PubMed]
- Guilhamon, P.; Lupien, M. SMuRF: A Novel Tool to Identify Regulatory Elements Enriched for Somatic Point Mutations. BMC Bioinform. 2018, 19, 454. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rodríguez, M.A.; Santander-Jiménez, S. Parallel Computing in Bioinformatics: A View from High-Performance, Heterogeneous, and Cloud Computing. J. Supercomput. 2019, 75, 3369–3373. [Google Scholar] [CrossRef]
- Sajulga, R.; Easterly, C.; Riffle, M.; Mesuere, B.; Muth, T.; Mehta, S.; Kumar, P.; Johnson, J.; Gruening, B.A.; Schiebenhoefer, H.; et al. Survey of Metaproteomics Software Tools for Functional Microbiome Analysis. PLoS ONE 2020, 15, e0241503. [Google Scholar] [CrossRef]
- Teer, J.K. An Improved Understanding of Cancer Genomics through Massively Parallel Sequencing. Transl. Cancer Res. 2014, 3, 243. [Google Scholar] [CrossRef]
- Khella, C.A.; Mehta, G.A.; Mehta, R.N.; Gatza, M.L. Recent Advances in Integrative Multi-Omics Research in Breast and Ovarian Cancer. JPM 2021, 11, 149. [Google Scholar] [CrossRef]
- Hassanpour, S.H.; Dehghani, M. Review of Cancer from Perspective of Molecular. J. Cancer Res. Pract. 2017, 4, 127–129. [Google Scholar] [CrossRef]
- Medema, M.H.; Paalvast, Y.; Nguyen, D.D.; Melnik, A.; Dorrestein, P.C.; Takano, E.; Breitling, R. Pep2Path: Automated Mass Spectrometry-Guided Genome Mining of Peptidic Natural Products. PLoS Comput. Biol. 2014, 10, e1003822. [Google Scholar] [CrossRef]
- Chakraborty, P. Gene Cluster from Plant to Microbes: Their Role in Genome Architecture, Organism’s Development, Specialized Metabolism and Drug Discovery. Biochimie 2022, 193, 1–15. [Google Scholar] [CrossRef]
- Taylor, A.M.; Shih, J.; Ha, G.; Gao, G.F.; Zhang, X.; Berger, A.C.; Schumacher, S.E.; Wang, C.; Hu, H.; Liu, J.; et al. Genomic and Functional Approaches to Understanding Cancer Aneuploidy. Cancer Cell 2018, 33, 676–689.e3. [Google Scholar] [CrossRef]
- Raphael, B.J.; Dobson, J.R.; Oesper, L.; Vandin, F. Identifying Driver Mutations in Sequenced Cancer Genomes: Computational Approaches to Enable Precision Medicine. Genome Med. 2014, 6, 5. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Wendl, M.C.; McMichael, J.F.; Raphael, B.J. Expanding the Computational Toolbox for Mining Cancer Genomes. Nat. Rev. Genet. 2014, 15, 556–570. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska, A.; Rudzki, M.; Rudzki, S.; Lewandowski, T.; Laskowska, B. Environmental Risk Factors for Cancer—Review Paper. Ann. Agric. Environ. Med. 2019, 26, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mohs, R.C.; Greig, N.H. Drug Discovery and Development: Role of Basic Biological Research. TRCI 2017, 3, 651–657. [Google Scholar] [CrossRef]
- Fang, Z.-Z.; Gonzalez, F.J. LC–MS-Based Metabolomics: An Update. Arch. Toxicol. 2014, 88, 1491–1502. [Google Scholar] [CrossRef]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-Omics Approaches to Disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- Kind, T.; Fiehn, O. Advances in Structure Elucidation of Small Molecules Using Mass Spectrometry. Bioanal. Rev. 2010, 2, 23–60. [Google Scholar] [CrossRef]
- Ziemert, N.; Alanjary, M.; Weber, T. The Evolution of Genome Mining in Microbes—A Review. Nat. Prod. Rep. 2016, 33, 988–1005. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. AntiSMASH 5.0: Updates to the Secondary Metabolite Genome Mining Pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Epstein, S.C.; Charkoudian, L.K.; Medema, M.H. A Standardized Workflow for Submitting Data to the Minimum Information about a Biosynthetic Gene Cluster (MIBiG) Repository: Prospects for Research-Based Educational Experiences. Stand. Genom. Sci. 2018, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- Dejong, C.A.; Chen, G.M.; Li, H.; Johnston, C.W.; Edwards, M.R.; Rees, P.N.; Skinnider, M.A.; Webster, A.L.H.; Magarvey, N.A. Polyketide and Nonribosomal Peptide Retro-Biosynthesis and Global Gene Cluster Matching. Nat. Chem. Biol. 2016, 12, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Morales, P.; Kopp, J.F.; Martínez-Guerrero, C.; Yáñez-Guerra, L.A.; Selem-Mojica, N.; Ramos-Aboites, H.; Feldmann, J.; Barona-Gómez, F. Phylogenomic Analysis of Natural Products Biosynthetic Gene Clusters Allows Discovery of Arseno-Organic Metabolites in Model Streptomycetes. Genome Biol. Evol. 2016, 8, 1906–1916. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zheng, Z.; Zou, H.; Li, N.; Wu, M. Characterization of the Secondary Metabolite Biosynthetic Gene Clusters in Archaea. Comput. Biol. Chem. 2019, 78, 165–169. [Google Scholar] [CrossRef]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Wieland Brown, L.C.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into Secondary Metabolism from a Global Analysis of Prokaryotic Biosynthetic Gene Clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef]
- Skinnider, M.A.; Dejong, C.A.; Rees, P.N.; Johnston, C.W.; Li, H.; Webster, A.L.H.; Wyatt, M.A.; Magarvey, N.A. Genomes to Natural Products PRediction Informatics for Secondary Metabolomes (PRISM). Nucleic Acids Res. 2015, 43, 9645–9662. [Google Scholar] [CrossRef]
- Hughes, J.; Rees, S.; Kalindjian, S.; Philpott, K. Principles of Early Drug Discovery: Principles of Early Drug Discovery. Brit. J. Pharmacol. 2011, 162, 1239–1249. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Plemenitaš, A.; Oren, A. Strategies of Adaptation of Microorganisms of the Three Domains of Life to High Salt Concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef]
- Ren, H.; Shi, C.; Zhao, H. Computational Tools for Discovering and Engineering Natural Product Biosynthetic Pathways. iScience 2020, 23, 100795. [Google Scholar] [CrossRef]
- Noman, E.; Al-Shaibani, M.M.; Bakhrebah, M.A.; Almoheer, R.; Al-Sahari, M.; Al-Gheethi, A.; Radin Mohamed, R.M.S.; Almulaiky, Y.Q.; Abdulaal, W.H. Potential of Anti-Cancer Activity of Secondary Metabolic Products from Marine Fungi. J. Fungi 2021, 7, 436. [Google Scholar] [CrossRef]
- Metpally, R.; Reddy, B. Comparative Proteome Analysis of Psychrophilic versus Mesophilic Bacterial Species: Insights into the Molecular Basis of Cold Adaptation of Proteins. BMC Genom. 2009, 10, 11. [Google Scholar] [CrossRef]
- El-Elimat, T.; Figueroa, M.; Ehrmann, B.M.; Cech, N.B.; Pearce, C.J.; Oberlies, N.H. High-Resolution MS, MS/MS, and UV Database of Fungal Secondary Metabolites as a Dereplication Protocol for Bioactive Natural Products. J. Nat. Prod. 2013, 76, 1709–1716. [Google Scholar] [CrossRef]
- Jaspars, M.; De Pascale, D.; Andersen, J.H.; Reyes, F.; Crawford, A.D.; Ianora, A. The Marine Biodiscovery Pipeline and Ocean Medicines of Tomorrow. J. Mar. Biol. Ass. 2016, 96, 151–158. [Google Scholar] [CrossRef]
- Umemura, M.; Koike, H.; Nagano, N.; Ishii, T.; Kawano, J.; Yamane, N.; Kozone, I.; Horimoto, K.; Shin-ya, K.; Asai, K.; et al. MIDDAS-M: Motif-Independent De Novo Detection of Secondary Metabolite Gene Clusters through the Integration of Genome Sequencing and Transcriptome Data. PLoS ONE 2013, 8, e84028. [Google Scholar] [CrossRef]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; de los Santos, E.L.C.; Kim, H.U.; Nave, M.; et al. AntiSMASH 4.0—Improvements in Chemistry Prediction and Gene Cluster Boundary Identification. Nucleic Acids Res. 2017, 45, W36–W41. [Google Scholar] [CrossRef] [PubMed]
- Talkal, R.; Tikariha, H.; Purohit, H. An Approach to In Silico Dissection of Bacterial Intelligence Through Selective Genomic Tools. Indian J. Microbiol. 2018, 58, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Khater, S.; Anand, S.; Mohanty, D. In Silico Methods for Linking Genes and Secondary Metabolites: The Way Forward. Synt. Syst. Biotechnol. 2016, 1, 80–88. [Google Scholar] [CrossRef]
- Lakhani, J.; Khunteta, A.; Chowdhary, A.; Harwani, D. In Silico Detection Tools to Identify Fungal Secondary Metabolites and Their Biosynthetic Gene Clusters. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2020; pp. 23–35. ISBN 978-0-12-821008-6. [Google Scholar]
- Vesth, T.C.; Brandl, J.; Andersen, M.R. FunGeneClusterS: Predicting Fungal Gene Clusters from Genome and Transcriptome Data. Synt. Syst. Biotechnol. 2016, 1, 122–129. [Google Scholar] [CrossRef]
- Agrawal, P.; Mohanty, D. A Machine Learning-Based Method for Prediction of Macrocyclization Patterns of Polyketides and Non-Ribosomal Peptides. Bioinformatics 2021, 37, 603–611. [Google Scholar] [CrossRef]
- Mohamed, A.; Nguyen, C.H.; Mamitsuka, H. Current Status and Prospects of Computational Resources for Natural Product Dereplication: A review. Brief. Bioinform. 2016, 17, 309–321. [Google Scholar] [CrossRef]
- Li, H.D.; Xu, Y.; Zhu, X.; Liu, Q.; Omenn, G.S.; Wang, J. A knowledge-Integrated Clustering Approach Based on Expression Profiles of Gene Sets. J. Bioinform. Comput. Biol. 2020, 18, 2040009. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, K.; Chen, I.-M.A.; Chu, K.; Ratner, A.; Seshadri, R.; Kyrpides, N.C.; Ivanova, N.N.; Mouncey, N.J. IMG-ABC v.5.0: An Update to the IMG/Atlas of Biosynthetic Gene Clusters Knowledgebase. Nucleic Acids Res. 2019, 48, D422–D430. [Google Scholar] [CrossRef] [PubMed]
- Greco, C.; Keller, N.P.; Rokas, A. Unearthing Fungal Chemodiversity and Prospects for Drug Discovery. Curr. Opin. Microbiol. 2019, 51, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Weber, T.; Medema, M.H. BiG-FAM: The Biosynthetic Gene Cluster Families Database. Nucleic Acids Res. 2021, 49, D490–D497. [Google Scholar] [CrossRef]
- Aleti, G. Secondary Metabolite Biosynthetic Gene Diversity–Bacillales Non-Ribosomal Peptides and Polyketides in Plant-Microbe Interactions. Ph.D. Thesis, Austrian Institute of Technology, Seibersdorf, Austria, 2016. [Google Scholar]
- Röttig, M.; Medema, M.H.; Blin, K.; Weber, T.; Rausch, C.; Kohlbacher, O. NRPSpredictor2—A Web Server for Predicting NRPS Adenylation Domain Specificity. Nucleic Acids Res. 2011, 39, W362–W367. [Google Scholar] [CrossRef]
- Bragin, E.; Chatzimichali, E.A.; Wright, C.F.; Hurles, M.E.; Firth, H.V.; Bevan, A.P.; Swaminathan, G.J. DECIPHER: Database for the Interpretation of Phenotype-Linked Plausibly Pathogenic Sequence and Copy-Number Variation. Nucleic Acids Res. 2014, 42, D993–D1000. [Google Scholar] [CrossRef]
- Maiti, P.; Sharma, P.; Nand, M.; Bhatt, I.D.; Ramakrishnan, M.A.; Mathpal, S.; Joshi, T.; Pant, R.; Mahmud, S.; Simal-Gandara, J.; et al. Integrated Machine Learning and Chemoinformatics-Based Screening of Mycotic Compounds against Kinesin Spindle ProteinEg5 for Lung Cancer Therapy. Molecules 2022, 27, 1639. [Google Scholar] [CrossRef]
- Boddy, C.N. Bioinformatics Tools for Genome Mining of Polyketide and Non-Ribosomal Peptides. JIMB 2014, 41, 443–450. [Google Scholar] [CrossRef]
- Juan, H.-F.; Huang, H.-C. Bioinformatics: Microarray Data Clustering and Functional Classification. Methods Mol. Biol. 2007, 382, 405–416. [Google Scholar]
- Emam, H.E.; El-Shahat, M.; Allayeh, A.K.; Ahmed, H.B. Functionalized starch for formulation of graphitic carbon nanodots as viricidal/anticancer laborers. Biocatal. Agric. Biotechnol. 2023, 47, 102577. [Google Scholar] [CrossRef]
- Ahmed, H.B.; El-Shahat, M.; Allayeh, A.K.; Emam, H.E. Maillard Reaction for Nucleation of Polymer Quantum Dots from Chitosan-Clucose Conjugate: Antagonistic for Cancer and Viral Diseases. Int. J. Biol. Macromol. 2023, 224, 858–870. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of the selected novel cytotoxic compounds from extremophilic fungi.

Figure 2.
Steps of antiSMASH and SMURF algorithms’ analysis, from the input (sequenced DNA) to output files.
Figure 2.
Steps of antiSMASH and SMURF algorithms’ analysis, from the input (sequenced DNA) to output files.

Figure 3.
The stages of early clinical trials leading to cancer drug discovery.


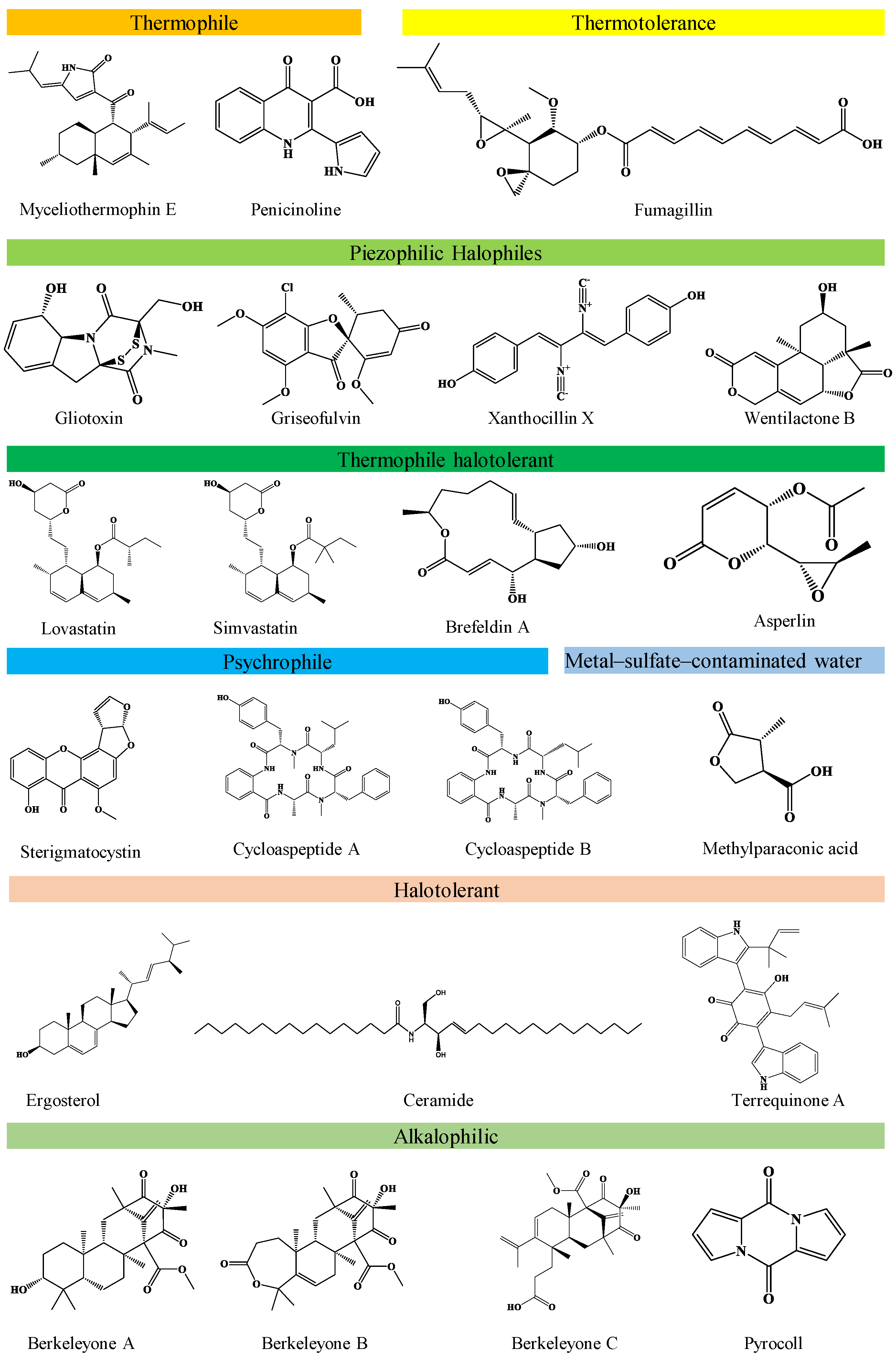{kind=link}
{kind=link}
{kind=link}
Table 1.
Investigated antitumour properties and target cell lines of fungi that proliferate, grow, and develop under extreme temperature conditions.
Table 1.
Investigated antitumour properties and target cell lines of fungi that proliferate, grow, and develop under extreme temperature conditions.
| Species | Type of Extreme Fungi | Compounds | References | |||
|---|---|---|---|---|---|---|
| Living and Thriving Habitats | Metabolites with Antitumour Properties | Effective Dose | Examples of Tissues or Cells Affected by Metabolites | |||
| Penicillium algidum | Psychrophile | Soil, Zackenberg, Greenland | Cyclic nitropeptide Psychrophilin A (nitropeptide) Psychrophilin D (nitropeptide) | 10.1 μg/mL | Cytotoxicity against P388 murine leukaemia cell lines | [73,89,93] |
| Cycloaspeptide A, B | ˃12 μg/mL | Antitumour activity in general | ||||
| Friedmanniomyces endolithicus | Psychrophile | Antarcticrocky deserts | Melanin pigments | – | Cytotoxicity against HFB4 cell line | [94] |
| Aspergillus sp. | Psychrophile | Alga-derived, North Sea, Germany | Cyclotripeptide Sterigmatocystin Psychrophilin E–F | 4.4–28.5 μM | Antitumour activity in general Cytotoxicity against HCT-8 (colorectal), BEL-7402, BGC-823, A-549, and A–2780, HCT116 cell line | [73,95] |
| Aspergillus clavatus C2WU | Thermophile | Hydrothermal vents crab Kueishantao, Taiwan | Clavatustide A, B | 15 μg/mL | HCC cell lines (Bel-7402, HepG2, and SMMC-7721), suppress HHC cell proliferation by inducing G1 arrest and suppressing the G1/S transition in a dose-dependent manner | [96] |
| Mucor strictus | Psychrophile | Shandong, China | Ergosterol | 37–78 μM | HeLa and MCF-7 cell lines | [97] |
| Myceliophthora thermophila | Thermophile | Fumaroles soil, Taipei, Taiwan | Myceliothermophins A–E | 0.26–1.05 μg/mL | A549, Hep3B, MCF-7, and HepG2cell lines | [26,98] |
| Malbranchea sulfurea | Thermophile | Hot spring, Taiwan | Malbranpyrroles A–F | 3–11 µM | Stop G0/G1 phase in all MCF-7 and HepG2 cells cancer cell lines | [99] |
| Aspergillus terreus | Thermophile halotolerant | Fumaroles soil, Hot spring, Taiwan | Terrein | 0.8 ± 0.3 μg/m | Suppressed the activity of ABCG2 transporters, activating the caspase-7 signalling pathway, and induced apoptosis by inhibiting the Akt pathway | [37,42,92,100] |
| L–asparaginase | 3.79–12.6 µg/mL | Cytotoxicity against HCT-116, Hep-G2, and MCF-7 cell lines | ||||
| Lovastatin | 0.6–15.6 μM | Cytotoxicity against lung carcinomas (DLRP, H1299), breast carcinomas (MCF-7, SKBR-3, BT474A, MDA-MB-453), and malignant melanomas (HT144, SK-MEL-28, M14) | ||||
| Asperlin | 1.1–110 nM | Inhibits breast cancer by induction of apoptosis and MCF-7; cytotoxicity against MCF-7 cell line | ||||
| Thelebolus sp. IITKGP-BT12 | Psychrophilic Halophilic | Sabinar saline wetland in Spain | Exopolysaccharide (EPS) | 275.42 μg/mL | Anti-proliferative activity against B16–F0 cells, with mediated apoptosis | [75,101] |
| Oidiodendron truncatum GW3–13 | Psychrophile | Soil, Great Wall station (Chinese Antarctic station), Antarctica | Piperazines | 0.3–28 nM | Antitumour activity in general | [77,102] |
| Oidioperazines A–D (Epipolythiodioxopiperazines) | >10.1 μg/mL | Cytotoxicity against HCT-8, BEL-7402, BGC-823 (gastric), A-549, and A-2780 cell lines | ||||
| Chetracins B–C (Epipolythiodioxopiperazines) | 0.003–0.028 μg/mL | |||||
| Chetracin D (Diketopiperazines) | 0.14 ± 0.03–1.83 ± 0.07 μg/mL | |||||
| Melinacidin IV, T988,T988 A | 3–28 nM | Cytotoxicity against HCT-8, BEL-7402 -BGC-823 (gastric), A-549, and A-2780 | ||||
| Penicillium sp. PR19 N–1 | Psychrophile | Marine sludge, Prydz Bay, Antarctica | Eremophilane–type sesquiterpenes | 5.2–85.8 μM | Cytotoxicity against HL-60 and A-549 cell lines | [30,42] |
| Chloro–eremophilane sesquiterpenes | 11.8–12.2 μM | |||||
| Aspergillus sp. | Psychrophile | Alga-derived, North Sea, Germany | Cyclotripeptide Sterigmatocystin | 4.4–28.5 μM | Antitumour activity in general | [42] |
| Trichoderma velutinum | Psychrophile | Soil, Himalayan, India | Lipopeptaibols | 2–7 μM | Antitumour activity in general | [42] |
| Penicillium rubrum | Thermophile/ acidiophile | Damp, water-damaged buildings or potato dextrose broth (pH 2.7) | Berkazaphilones A–C | 10–100 μg/mL | Solid tumour cancers and activation of caspase-1 in prostate cancer and ovarian cancer | [103] |
| Octadienoic acid derivatives | ||||||
| Berkedienoic acid | ||||||
| Berkedienolactone | ||||||
| Azaphilone | ||||||
| Vermistatin | ||||||
| Dihydrovermistatin | ||||||
| Penisimplicissin | ||||||
| Aldehyde | ||||||
| Methyl paraconic acid | ||||||
| Penicillium sp. | Thermophile | Ghamiqa hot springs, Saudi Arabia (temperature range from 45°C to 65 °C) | 3-(furan 12-carboxylic acid)-6-(methoxycarbonyl)-4-hydroxy-4-methyl-4 | – | Cytotoxicity against the lymphoma human cancer cell line HTB-176 | [37,42,90,103,104] |
| 5-dihydro-2H-pyran 1 3α-methyl-7-hydroxy-5-carboxylic acid methyl ester-1-indanone 2 | ||||||
| Piezophilic Halophiles | Deep-water sediment sample that was collected at a depth of 5115 m in the East Pacific | Ethyl Acetate (Etoac) | 2.6 µM | Inhibitor of acute myeloid leukaemia (AML) cells | ||
| Austinol | 2–7.6 μM | Cytotoxicity against HTB-176 cell line | ||||
| Emodin (1,3,8-trihydroxy-6-methylanthraquinone) | ||||||
| 2-methyl-penicinoline 5 | 11.3 µM | Inhibitor of HepG2 cell lines | ||||
| Penicinoline (γ-pyridone alkaloids) | 11.8–12.2 µM | |||||
| Methylpenicinoline γ-pyridone alkaloids | 11.3–13.2 µM | |||||
| Mycophenolic acid | 0.5–1 μM | Inhibitor of human inosine 5′-monophosphate dehydrogenase (IMPDH) | ||||
| Fudecadione A–B | 12.6–24 μM | Cytotoxicity against NCI-H187, MCF-7, and KB cell lines | ||||
| Ligerin | 117 nM | Inhibitor of POS1 cell lines | ||||
| Aspergillus fumigatus | Thermotolerant | Soil and decaying organic matter, Conidia surviving at 70 °C | Fumagillin | 8–40 µM | Inhibitor of B16, F10, A549, and L132 Cell line | [91] |
| Gliotoxin | 0.54–2.7 µM | |||||
| Penicillium chrysogenum | Thermotolerant | Damp or water-damaged buildings | Chloctanspirone A | 9.2, 39.7 μM | Cytotoxicity against HL-60 and A-549 cell lines | [37] |
Table 2.
Investigated antitumour properties and target cell lines of piezophilic or barophilic (high pressures) fungi metabolites.
Table 2.
Investigated antitumour properties and target cell lines of piezophilic or barophilic (high pressures) fungi metabolites.
| Species | Type of Extreme Fungi | Compounds | References | |||
|---|---|---|---|---|---|---|
| Living and Thriving Habitats | Name of Metabolites with Antitumour Properties | Effective Dose | Examples of Tissues or Cells Affected by Metabolites | |||
| Dichotomomyces cejpii F31-1 | Piezophilic Halophiles | Hainan Sanya National Coral Reef Reserve, China. | Dichotomocej A–D | 35–39.1 μM | Cytotoxicity against rhabdomyosarcoma (RD) and HCT116 cell line | [42,117,118,119] |
| Penicillium sp. SCSIO Ind16F01 | Piezophilic Halophiles | Deep water, deep-sea sediment, Indian Ocean | Chromone (Epiremisporine B–C) | 15.8–16.6 μM | Cytotoxicity against SGC7901, MCF-7, and K562 cell lines | |
| Penicillium purpurogenum G59 | Piezophilic Halophiles | Deep water, deep-sea sediment Indian Ocean | Chromone (Epiremisporine B, B1), Chromone (Isoconiochaetone C), Remisporine B, Coniochaetone A, Methyl 8-hydroxy-6-methyl-9-oxo-9H-xanthene-1-carboxylate | 54.7–144.3 μM | Cytotoxicity against K562, HL-60, HeLa, and BGC-823 cancer cell lines at 100 μg/mL | |
| Aspergillus sp. SCSIO Ind09F01 | Piezophilic Halophiles | Deep-sea China | Diketopiperazine | 0.015–95.4 μM | Cytotoxicity against H460, SF–268, MCF-7, HT-29, and CHO-K1 cell lines | [107] |
| Penicillium sp. F23-2 | Piezophilic Halophiles | Deep-sea sediment (depth 5080 m) | Meleagrins D–E | 5–<100–200 μg/mL | Cytotoxicity and Pro-apoptoticagainst A-549, HL-60, BEL-7402, and MOLT-4 cell lines | [42,120] |
| Penicillium sp. | Piezophilic Halophiles | Deep-sea sediment, Arabian Sea | Conidiogenones B–G (diterpenes) | 0.038–6.7 μM | Cytotoxicity against A-549, A-549, and HL-60 cell lines | [42,116,121,122] |
| Roquefortine groups (C, F–G), Xanthocillin X, Chrysogine | 1.8–6.7 μM | |||||
| Brevione I–H | 7.44–28.4 μM | Cytotoxicity against HL-60, A-549, MCF-7 and HEP3B cell lines | ||||
| Peniphenones A | 5.9 to 9.3 μg/mL | |||||
| Peniphenones D | ||||||
| Deep-sea sediment (depth 5115 m) | Breviane spiroditerpenoids Breviones I–K | 7.44–32.5 µM | Cytotoxicity against MCF-7 and A549 cell lines | |||
| Aspergillus dimorphicus SD317 | Piezophilic Halophiles | South China Sea sediment, (depth 2038 m) | Wentilactone A–B | Low micromolar dose | Cytotoxicity against HGC-27, HeLa, A549 and HT29 cell lines | [93] |
| Aspergillus versicolor | Piezophilic Halophiles | Deep-sea sediment, Bohai Sea, China | Brevianamide | 8.63 µM | Antitumour activity in general | [93,95] |
| Pacific Ocean, China (depth 2326 m) | Emodin (2-(dimethoxymethyl)-1-hydroxyanthracene-9,10-dione) | 3.9–62 μg | Moderate inhibitory activity against MRSA-CGMCC 1.12409 and MRSA ATCC-43300MIC | |||
| Penicillium brevicompactum DFFSCS025 | Piezophilic Halophiles | Deep-sea sediment, Hainan, China | Mycochromenic acid, Gliotoxin | 5.6–13.71 μM | Cytotoxicity against HCT116 cell lines | [42,104] |
| Aspergillus SCSIO Ind09F01 | Piezophilic Halophiles | Deep-sea sediment | Asperethers A | 0.015–95.4 μM and < 0.03–0.894 μM | Cytotoxicity against A-549, K562, and Huh-7 cell lines | [42] |
| Aspergillus wentii SD–310 | Piezophilic Halophiles | Deep-sea South China Sea (depth 2038 m) | Asperethers A, E | Micromolar range | Antitumour activity in general | [109] |
| Aspergillus westerdijkiae SCSIO 05233 | Piezophilic Halophiles | Deep-sea hypersaline sediments, South China Sea (Depth 4593 m) | Circumdatin F–G | 2.5–11.3 μM | Cytotoxicity against K562, HL60, myelogenous, and promyelocytic leukaemia cell lines | [109] |
| Acaromyces ingoldii FS121 | Piezophilic Halophiles | South China Sea (Depth 3415 m) | Naphtha- [2,3-b] Pyrandione analogue, Acaromyester A, Thiazole analogue | 100 μM range | Cytotoxicity against MCF-7, NCI-H460, SF-268, and HepG-2 | [110,112] |
| Penicillium brevicompactum DFFSCS025 | Piezophilic Halophiles | South China Sea deep-sea sediment | Brevianamide | 15.6 μM | Cytotoxicity against HCT116 cell lines | [93] |
Table 3.
Investigated Antitumour properties and target cell lines of acidophilic or alkalophilic fungi metabolites.
Table 3.
Investigated Antitumour properties and target cell lines of acidophilic or alkalophilic fungi metabolites.
| Species | Type of Extreme Fungi | Compounds | References | |||
|---|---|---|---|---|---|---|
| Living and Thriving Habitats | Name of Metabolites with Antitumour Properties | Effective Dose | Examples of Tissues or Cells Affected by Metabolites | |||
| Pithomyces sp. | Acidophiles, Piezophiles and metal tolerance | Surface and deep-sea water (a 1500 ft deep) Berkeley Pit Lake with high pH (2.7), North America | Aromatic compounds (not named) | 0.4–4.61 μg/mL | Cytotoxicity against A549, SK-OV-3, SK-MEL-2, XF-498, HCT-15 | [143] |
| Penicillium rubrum | Acidophiles | Surface water in Berkeley Pit Lake, North America | Berkeleyones A–D | 2.7–37.8 μM | Cytotoxicity against interleukin 1-β production and caspase-1 enzyme signalling in THP-1 cell lines | [65,138,139,142] |
| Meroterpenes | 0.37–0.64 µM | Cytotoxicity against MMP-3, NCI-H460, IL-1β RPMI-8226 cell, RPMI-8226, SR, RPMI-8226 cell lines as caspase-1 inhibitors | ||||
| Berkedienolactone | 10–100 μg/mL | |||||
| ‘Octadienoic acid (berkedienoic derivatives) | 5.67 μM | |||||
| Sesquiterpene | 300 nM–30 μM | Moderate caspase-1-inhibiting activity and strong cytotoxicity against MMP-3 activity | ||||
| Vermistatin | 2.0–200 μg/mL | Inhibitors of caspase-1, production of IL-1β in THP-1 and leukaemia cell lines | ||||
| Penisimplicissin | 10–100 μg/mL | |||||
| Dihydrovermistatin | 20 μg/mL | |||||
| Methylparaconic acid | – | |||||
| Berkazaphilone B–C (Polyketoids) | 2.0–20 μg/mL | Antitumour activity in general | ||||
| Cysteine protease | 0.0052–34 µg/mL | Suppressing the caspase-1 and MMP-3 | ||||
| Penicillium sp. | Acidophiles | Surface water or algae-associated, in Berkeley Pit Lake, North America | Berkeleytrione (Hybrid–polyketideterpenoid) | 6.40 μM | Suppressing the caspase-1 and matrix Metalloproteinase-3 (MMP-3), with moderate activity against lung cancer cell lines | [42,99,126,128,134,136,141,144,145,146] |
| Sesquiterpene | 300 nM–30 μM | Moderate caspase-1-inhibiting activity and strong activity against MMP-3 cell lines | ||||
| Berkeleyacetal C, Drimane sesquiterpene, Berkelic acid Berkeleydione | Micromolar to millimolar range | MMP-3 and caspase-1 inhibitor activity in NCI–DTP 60 tumour cell lines; suppressing metalloproteinase-3 (MMP-3) and the cysteine protease caspase-1 (Casp-1), and cytotoxic activity against NCI-H460 cell lines | ||||
| Aspergillus fumigatus KMC901 | Acidophiles | Contaminated acid mine Gangneung, South Korea | Diketopiperazine | 0.24–0.9 μM | Cytotoxicity against DU145, AGS, A549, and HCT-116 cell lines | [147] |
| Penicillium clavigerum | Acidophiles | Water sample in Berkeley Pit Lake, North America | Phomopsolides A–F, Phomopsolide C | >100–150 μM | Suppressing MMP-3 and caspase-1 | [134] |
| Pleurostomophora sp. | Acidophiles | Water sample, Berkeley Pit Lake, North America | Berkchaetoazaphilones B (Azaphilones) | 1.1–10 μM | Inhibited the TNFα, IL-6, and IL-1β production in inflammation assay and significant cytotoxicity activity against Y79 and LOX-IMVI cell lines | [148] |
| Penicillium solitum | Acidophiles | Water sample, Berkeley Pit Lake, North America | Berkedrimanes A–B (Sesquiterpene lactones), Tricarboxylic acid, Berkeleyacetals C | Micromolar range | Suppressing caspase-1 and caspase-3 in the micromolar range and mitigated the production of IL-1β by intact inflammasomes at low micromolar concentrations | [42,134,149] |
| Emericellopsis alkalina | Alkali-tolerant or Alkalophilic | Alkaline soil samples of Zheltyr Lake, Russia | Emericellipsins A–E | 2.30 ± 0.30–8.00 ± 1.04 μM | Cytotoxicity against HMO2, HCT-116, MCF 7, K-562, B16, MDA-MB-231, HepG2, etc. | [124,125] |
| Streptomyces sp. AK 409 | Alkali-tolerant or Alkalophilic | Water samples of high Ph | Pyrocoll | 0.65±0.09 μg/mL | Antitumor activity against HMO2, MCF 7, HepG2, etc. | [42,125,150] |
| Emericellipsin A | 2.8 and <0.5 μM | HepG2 and HeLa cell lines | ||||
Table 4.
Investigated Antitumour properties and target cell lines of halophilic and halotolerant fungi metabolites.
Table 4.
Investigated Antitumour properties and target cell lines of halophilic and halotolerant fungi metabolites.
| Species | Type of Extreme Fungi | Compounds | References | |||
|---|---|---|---|---|---|---|
| Living and Thriving Habitats | Name of Metabolites with Antitumour Properties | Effective Dose | Examples of Tissues or Cells Affected by Metabolites | |||
| Aspergillus sp. | Halotolerant | Sonoran Desert, Arizona | Terrequinone | 0.6–1.2 mM | Cytotoxicity against MCF-7, SF-268, and NCI–H460 cell lines | [29,37,42,107,176] |
| 11-methoxycurvularin, 11-hydroxycurvularin, Dehydrocurvularin, Penicillic acid, 13-deoxy-12(13)-dehydroterrefuranone | 1.1–2.5 μM | Cytotoxicity against MIA Pa Ca-2, MCF-7, SF-268, NCI–H460, and MIA Pa Ca-2 cell lines | ||||
| Aspochalasins | 0.6–1.2 mM | Antitumour activity in general | ||||
| Ergosterol | 3.3 μM | Selective activity against RKO cells | ||||
| Cytochalasin E | 37–78 μM | Cytotoxicity RKO, A-549, and BEL-7402, All A549, Hela, BEL-7402, and RKO cell lines. | ||||
| Rosellichalasin | – | |||||
| Aspergillus sp. F1 | Halotolerant | Solar saltern, Shandong, China | Aspochalasins A, C–D, TMC-169, Qunine | 37–78 μM | Cytotoxicity against MCF-7, SF-268, and NCIH60 cell lines | [42] |
| Aspergillus variecolor B-17 | Halophilic, Halotolerant, Xerophilic | Jilantai salt field, Inner Mongolia, China | Variecolortides A–C (Alkaloids) | 61, 69, and 71 μM | Cytotoxicity against K-56 2 human leukaemia cell line | |
| Indole-3-ethenamide | 3.027 μM | Cytotoxicity against P388, A549, HL-60, and BEL-7402 cell activity against DPPH cell lines | [42,176,177] | |||
| 2-heptyl-3,6-dihydroxy-5-(3-methyl-2-butenyl) ben-zaldehyde and 2-(3E,5E-heptadienyl)-3,6-di-hydroxy-5-(3-methyl-2-bu-tenyl) benzaldehyde | 9.99–203 μmol | |||||
| Tetrahydroauroglaucin (As-pergin) | 3–30.5 μM | Cytotoxicity against human U251 and glioma U87MG cell lines | ||||
| Flavoglauci | – | |||||
| Variecolorquinones A, B | 1.3 and 3.7 μM | Antitumour activity in general | ||||
| Myrothecium sp. GS-1 | Halophilic | Saline soil, Gansu, China | Ceramide(N-acetyl-3, 5, 11, 18-tetrahydroxyoctadecyl-2-amine) | 63.6 μM | Cytotoxicity against PC-3, HL-60, and MCF-7 cell lines | [42,64,178] |
| Variecolorins A–K | 3.3 μM | |||||
| Aspergillus versicolor KR87 | Halophilic | Soil fungi from the salt desert Little Rann of Kutch, India | Carnosol, Isouvaretin, Ginsenoside Rg5, Boesenbergin A | 6.98–12.8 g/mL | Antitumour activity in general | [179] |
| Tegafur | 0.59–1.38 μg/mL | |||||
| Procarbazine | 0.15 mM | |||||
| Penicillium chrysogenum or Penicillium rubens | Halotolerant | Subtropical, salted, and damp or water-damaged buildings | Chloctanspirone A and B | 9.2 and 39.7 μM | Cytotoxicity against HL-60 and lung cancer and A-549 cell lines | [122,163] |
| Aspergillus flocculosus PT05-1 | Halotolerant | Putian saltern of Fujian Province of China | (22R,23S) -epoxy-3b,11a,14b,16b-tetrahydroxyergosta-5,7-dien-12–one, 6- (1H-pyrrol-2-yl) hexa-1,3,5-trienyl-4-methoxy-2H-pyran-2-one, Ergosteroid 1, 7-nor-ergosterolide, 3b-hydroxyergosta-8,24(28) -dien-7-one | 12–18 μM | Cytotoxicity against HL-60 and BEL-7402 cell lines | [180] |
| Aspergillus sclerotiorum PT06-1 | Halotolerant | Salt field, Fujian, China | Ceramide (N-acetyl-3, 5,11, 18-tetrahydroxyoctadecyl-2-amine) | 63.6 μM | Cytotoxicity against PC-3, HL-60, and MCF-7 cell lines | [37,42] |
| Asperphenamate | 92.3–97.9 μM | Cytotoxicity against T47D, MDA-MB-231, and HL-60 cell lines | ||||
| Aspergillus repens strain K42 | Xerophilic | Rice sample collected in Korea | Anthraquinones2-(dimethoxymethyl)-1-hydroxyanthracene-9,10-dione | 3.9–62 μg/mL | Inhibitory activity against MRSA CGMCC 1.12409 and MRSA ATCC 43300 | [42,181] |
| Alkaloids | ||||||
| Chaetomium globosum | Xerophilic Endophytic | Sonoran Desert, Arizona | Globosumones A–B, Orsellinic acid esters | 3.4–9.5 μM | Cytotoxicity against SF-268, MCP-7, MIA Pa Ca-2, and NCI-H460 cell lines | [42,182] |
| Aspergillus flavipes | Xerophilic endophytic | Sonoran Desert, Arizona | Asperphenamate | 3.0 and 18.3 μM | Moderate cytotoxic activity against several cancer cell lines | [42,183,184] |
| Aspochalasins A, C–D, TMC-169 | 3.4–9.5 μM | Cytotoxicity against MCF-7, SF-268, and NCIH60 cell lines | ||||
| Flavichalasines A–F | 9.6–26.6 μM | Cytotoxicity against HL60, NB4, 231, HEP-3B, HCT116, and RKO cell lines | ||||
| Cladosporium cladosporioides | Xerophilic Psychrophilic | Saprotroph occurring as a secondary infection on decaying or necrotic parts of plants | Perylenequinones | 1 μM/L | Induce apoptosis when Hce-8693 cells were incubated with photoactivated EA, HA, and HB | [42] |
| Cladochromes F, G | 0.05–50 μM | Antitumour activity in general | ||||
| Monascus sp. | Halotolerant Xerophilic Halotolerant | Non-sterile dried red fermented rice sample | Monaphilone A, B Ankaflavin Canthaxanthin Monascorubramin Rubropunctatin | 55.3–77.6 μM | Cytotoxicity against Hep G2, Hep-2, and A549 human cancer cell lineS | [185,186,187] |
| Aureobasidium pullulans | Halotolerant | Salt farm soil | Pullulan | 32.2–63.1 μg liamocins/mL | Cytotoxicity against liver cancer cells | [167,169,171] |
| Soluble glucans | 174.29 μg/mL | |||||
Table 5.
Investigated Antitumour properties and target cell lines of other contaminated environments’ fungal metabolites.
Table 5.
Investigated Antitumour properties and target cell lines of other contaminated environments’ fungal metabolites.
| Species | Type of Extreme Fungi | Compounds | References | |||
|---|---|---|---|---|---|---|
| Living and Thriving Habitats | Name of Metabolites with Antitumour Properties | Effective Dose | Examples of Tissues or Cells Affected by Metabolites | |||
| Penicillium simplicissimum | Heavy metal-tolerant Cd (II) | Decaying vegetation | Dihydrovermistatin | 8 µM | Plays a role in regulating inflammation and apoptosis. Inhibition of the signal transduction enzyme caspase-1, activity against 60 human cell lines | [188,189] |
| Penicillium rubrum | Metal-sulphate-contaminated water | Berkeley Pit Lake, a depth of 270 m abandoned open-pit copper mine Butte, Montana | Aldehyde | – | Signal transduction enzyme caspase-1 or because of their structural similarity to these inhibitors and ability to inhibit interleukin-1β production by inflammasomes in induced THP-1 cells | [42,99,104,134] |
| Methylparaconic acid | ||||||
| Vermistatin analogue | 0.28–33.9 µM | |||||
| Penisimplicissin | 0.33–0.66 mM | |||||
| Acid mine waste lake in Montana | Berkazaphilones A–C | 20 μg/mL | Cytotoxicity to leukaemia cancer cell lines through inhibition of caspase-1 | |||
| Azaphilone-type polyketides | Exhibited potent cytotoxic effects against human retinoblastoma, leukaemia, and melanoma cell lines | |||||
| Red pigment berkchaetorubramine | – | |||||
| Alternaria sp. ZJ9-6B | Extreme temperatures and UV-radiation tolerant | With Mangrove (Aegiceras corniculatum) populations South China Sea | Alterporriol K, M | 13.1–29.1 μM | Cytotoxicity against MDA-MB-435 and MCF-7 cell lines | [42,190] |
| Resveratrol | ||||||
| Porritoxin | ||||||
| Penicillium chrysogenum or Penicillium rubens | Thermophile | Subtropical regions, damp or water-damaged buildings | Chloctanspirone A, B | 9.2–39.7 μM | Cytotoxicity against leukaemia HL-60 and A-549 cell lines | [163] |
| Alternaria tenuis Sg17-1 | Extreme temperature- and UV-radiation-tolerant | Damp or water-damaged buildings | Isocoumarins B, F | 0.002–0.02 mM | Cytotoxicity against A375-S2 and Hela cell lines | [191] |
| Sg17-1-4 | ||||||
| Abortiporus biennis | Cu2O- and/or Al2O3-tolerant | Roots or wood chips of deciduous trees, Asia, America, and Europe | Laccases | 6.7, 12.5, 9.2 mM | Cytotoxicity against Hep G2 and MCF-7 cells and inhibitory activity against HIV-1 RT | [99,143] |
| Isocoumarins B, F | ||||||
| Pithomyces sp. | Acidophiles, piezophiles, and metal ions tolerance | Berkeley Pit, North America (depth of 1500 ft) | Aromatic compounds, Pitholides A–D | – | Cytotoxicity against brine shrimp cancer cell lines | [42,123] |
Table 6.
Computational, “Omics”, and bioinformatic tools and databases for identification of metabolic gene clusters related to drug discovery of extremophiles and extremotolerant fungi.
Table 6.
Computational, “Omics”, and bioinformatic tools and databases for identification of metabolic gene clusters related to drug discovery of extremophiles and extremotolerant fungi.
| Name of Tools | System Requirements | Inputs | Functions, Outputs, and Accessibility Approaches | References |
|---|---|---|---|---|
| AntiSMASH and FungiSMASH’ |
|
|
| [38,148,241,242,243,244,245,246,247,248,249] |
| SMuRF |
|
|
| [213,214,242,243,250,251,252] |
| CLUSEAN/Cluster Sequence Analyzer |
|
|
| [120,244,247] |
| GNP/PRISM |
|
|
| [74,248,253,254,255,256,257,258,259,260] |
| DEREPLICATOR+ |
|
|
| [99,246,250,261,262,263,264,265,266,267,268,269] |
| SBSPKS |
|
|
| [214,254,255] |
| ClusterMine 360 |
|
|
| [270] |
| IMG–ABC |
|
|
| [271] |
| MIBiG |
|
|
| [272,273] |
| C–Hunter |
|
|
| [274] |
| NRPS Predictor |
|
|
| [259,260,261,275] |
| ClustScan Professional |
|
|
| [254,255,256,257,258,276] |
| DECIPHERⓇ |
|
|
| [276] |
| MeFSAT | Any Web browser in any OS |
|
| [277] |
| NaPDoS® | Any Web browser in any OS |
|
| [278,279] |
| NP.searcher |
|
| Coordinates for NRPS/PKS
| [164] |
| CASSIS/SMIPS |
|
|
| [262,263,264] |
| MIDDAS-M |
|
|
| [262,265] |
| FunGeneClusterS |
|
|
| [266,267] |
| SeMPI |
|
|
| [108,268] |
| GRAPE |
|
|
| [250,255,268] |
| GARLIC |
|
|
| [220,250] |
| SeMPI |
|
|
| [268] |
| SEARCH-PKS/NRPS-PKS/SBSPKS |
|
|
| [192] |
| ClusterFinder |
| Input File: Example_Input.Txt: Column Description include:
|
| [16] |
| Pep2Path |
|
|
| [237,247,248] |
| GNP/PRISMR |
|
|
| [248,253,274] |
| EvoMiningN |
|
|
| [274] |
| Java Treeview |
| Cross-platform rewrite extensions to the file format that allows the results of additional analysis |
| [279] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kiss, A.; Hariri Akbari, F.; Marchev, A.; Papp, V.; Mirmazloum, I. The Cytotoxic Properties of Extreme Fungi’s Bioactive Components—An Updated Metabolic and Omics Overview. Life 2023, 13, 1623. https://doi.org/10.3390/life13081623
AMA Style
Kiss A, Hariri Akbari F, Marchev A, Papp V, Mirmazloum I. The Cytotoxic Properties of Extreme Fungi’s Bioactive Components—An Updated Metabolic and Omics Overview. Life. 2023; 13(8):1623. https://doi.org/10.3390/life13081623
Chicago/Turabian StyleKiss, Attila, Farhad Hariri Akbari, Andrey Marchev, Viktor Papp, and Iman Mirmazloum. 2023. "The Cytotoxic Properties of Extreme Fungi’s Bioactive Components—An Updated Metabolic and Omics Overview" Life 13, no. 8: 1623. https://doi.org/10.3390/life13081623
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.