
Chemical Analysis and Biological Activities of Extracts Isolated from Symbiotic L. japonicus Plants
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material, Microbial Inoculation, and Growth Conditions
2.2. Testing Root Colonization by Microsymbionts
2.3. Treatments, Tissue Harvesting, and Extraction
- Non-inoculated plants grown at 21 °C (control sample “C1”)
- Non-inoculated plants grown at 24–25 °C (control sample “C2”)
- Plants inoculated with the Rhizobium bacterium M. loti (“R” sample)
- Plants inoculated with the Greece-isolated R. irregularis strain (“AMF” sample)
- Plants inoculated with the R. irregularis DAOM strain (“AMF-D” sample)
- Plants double-inoculated with Rhizobium and AMF (DAOM) (“R + D” sample)
2.4. Chemical Analysis of the Extracts
2.5. Cell Culture
2.6. Cell Viability Assay
2.7. Anti-Inflammatory Activity Assessment
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Lotus Japonicus Metabolite Profiling under Different Microbial Inoculation Treatments
3.2. Effects of L. japonicus Extracts on HEK-293 Cell Viability
3.3. Apoptotic Activities of L. japonicus Extracts
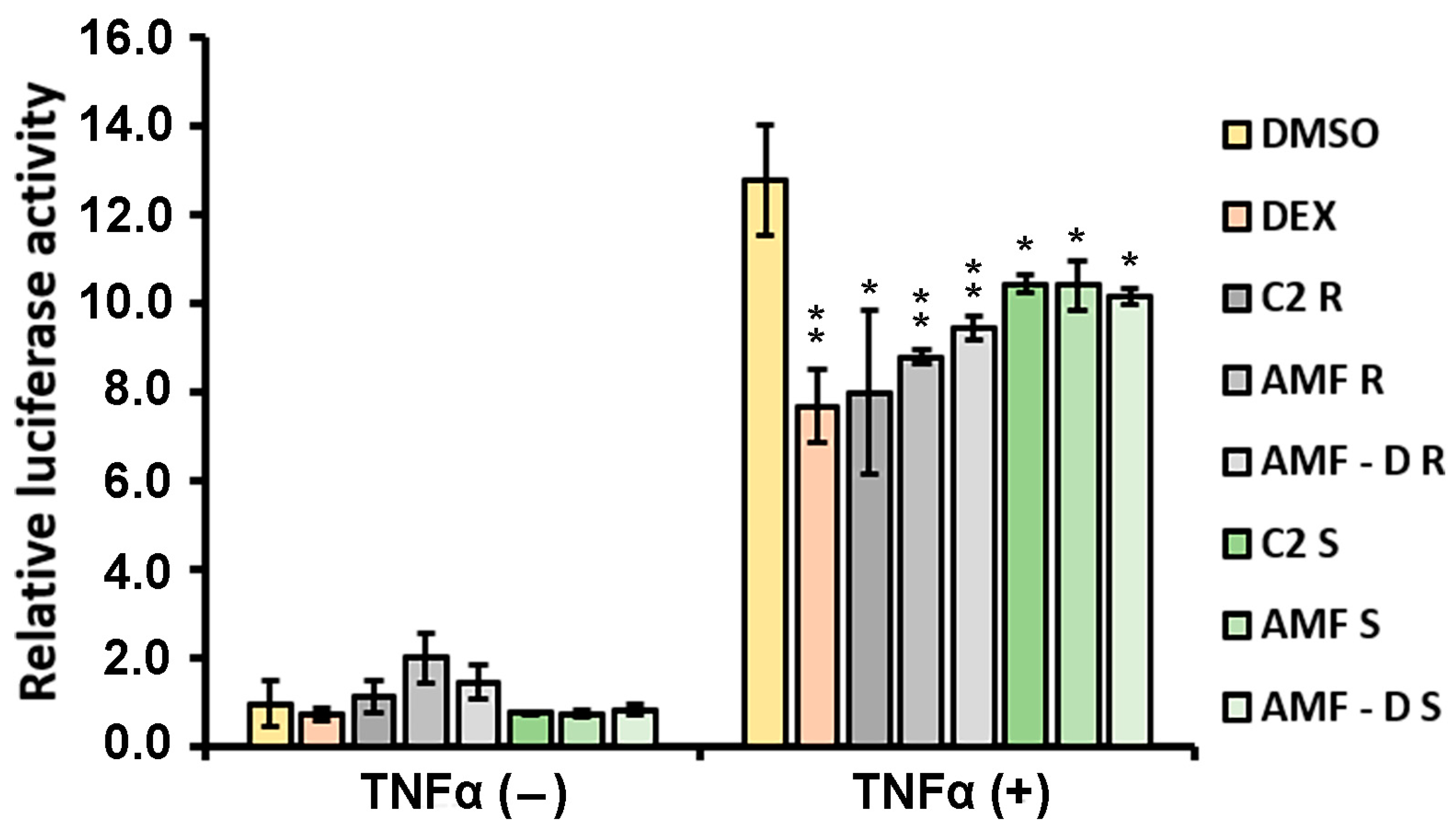
3.4. Anti-Inflammatory Properties of L. japonicus Extracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.-M. Plant Secondary Metabolite Biosynthesis and Transcriptional Regulation in Response to Biotic and Abiotic Stress Conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Hartmann, T. From Waste Products to Ecochemicals: Fifty Years Research of Plant Secondary Metabolism. Phytochemistry 2007, 68, 2831–2846. [Google Scholar] [CrossRef]
- Méteignier, L.-V.; Nützmann, H.-W.; Papon, N.; Osbourn, A.; Courdavault, V. Emerging Mechanistic Insights into the Regulation of Specialized Metabolism in Plants. Nat. Plants 2023, 9, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D. Mycorrhizal Symbiosis, 3rd ed.; Smith, S.E., Read, D., Eds.; Academic Press: London, UK, 2008; ISBN 978-0-12-370526-6. [Google Scholar]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cartabia, A.; Lalaymia, I.; Declerck, S. Arbuscular Mycorrhizal Fungi and Production of Secondary Metabolites in Medicinal Plants. Mycorrhiza 2022, 32, 221–256. [Google Scholar] [CrossRef]
- Amani Machiani, M.; Javanmard, A.; Habibi Machiani, R.; Sadeghpour, A. Arbuscular Mycorrhizal Fungi and Changes in Primary and Secondary Metabolites. Plants 2022, 11, 2183. [Google Scholar] [CrossRef]
- Schweiger, R.; Müller, C. Leaf Metabolome in Arbuscular Mycorrhizal Symbiosis. Curr. Opin. Plant Biol. 2015, 26, 120–126. [Google Scholar] [CrossRef]
- Chaudhary, V.; Kapoor, R.; Bhatnagar, A.K. Effectiveness of Two Arbuscular Mycorrhizal Fungi on Concentrations of Essential Oil and Artemisinin in Three Accessions of Artemisia annua L. Appl. Soil. Ecol. 2008, 40, 174–181. [Google Scholar] [CrossRef]
- Domokos, E.; Jakab-Farkas, L.; Darkó, B.; Bíró-Janka, B.; Mara, G.; Albert, C.; Balog, A. Increase in Artemisia annua Plant Biomass Artemisinin Content and Guaiacol Peroxidase Activity Using the Arbuscular Mycorrhizal Fungus Rhizophagus irregularis. Front. Plant Sci. 2018, 9, 478. [Google Scholar] [CrossRef]
- Domokos, E.; Bíró-Janka, B.; Bálint, J.; Molnár, K.; Fazakas, C.; Jakab-Farkas, L.; Domokos, J.; Albert, C.; Mara, G.; Balog, A. Arbuscular Mycorrhizal Fungus Rhizophagus irregularis Influences Artemisia annua Plant Parameters and Artemisinin Content under Different Soil Types and Cultivation Methods. Microorganisms 2020, 8, 899. [Google Scholar] [CrossRef]
- Sailo, G.L.; Bagyaraj, D.J. Influence of Different AM-Fungi on the Growth, Nutrition and Forskolin Content of Coleus forskohlii. Mycol. Res. 2005, 109, 795–798. [Google Scholar] [CrossRef]
- Singh, R.; Soni, S.K.; Kalra, A. Synergy between Glomus fasciculatum and a Beneficial Pseudomonas in Reducing Root Diseases and Improving Yield and Forskolin Content in Coleus forskohlii Briq. under Organic Field Conditions. Mycorrhiza 2013, 23, 35–44. [Google Scholar] [CrossRef]
- Bulut, M.; Wendenburg, R.; Bitocchi, E.; Bellucci, E.; Kroc, M.; Gioia, T.; Susek, K.; Papa, R.; Fernie, A.R.; Alseekh, S. A Comprehensive Metabolomics and Lipidomics Atlas for the Legumes Common Bean, Chickpea, Lentil and Lupin. Plant J. 2023, 116, 1152–1171. [Google Scholar] [CrossRef] [PubMed]
- Serventi, L.; Cai, X.; Chen, R.; Dilrukshi, N.; Su, J.; Tuange, R.P.N.; Ham, E.E. Anticancer Properties of Aqueous Extracts from Leguminosae. Nutraceuticals 2022, 2, 323–334. [Google Scholar] [CrossRef]
- Sánchez-Chino, X.; Jiménez-Martínez, C.; Dávila-Ortiz, G.; Álvarez-González, I.; Madrigal-Bujaidar, E. Nutrient and Nonnutrient Components of Legumes, and Its Chemopreventive Activity: A Review. Nutr. Cancer 2015, 67, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Guang, C.; Chen, J.; Sang, S.; Cheng, S. Biological Functionality of Soyasaponins and Soyasapogenols. J. Agric. Food Chem. 2014, 62, 8247–8255. [Google Scholar] [CrossRef]
- Ranner, J.L.; Schalk, S.; Martyniak, C.; Parniske, M.; Gutjahr, C.; Stark, T.D.; Dawid, C. Primary and Secondary Metabolites in Lotus japonicus. J. Agric. Food Chem. 2023, 71, 11277–11303. [Google Scholar] [CrossRef]
- Forslund, K.; Morant, M.; Jørgensen, B.; Olsen, C.E.; Asamizu, E.; Sato, S.; Tabata, S.; Bak, S. Biosynthesis of the Nitrile Glucosides Rhodiocyanoside A and D and the Cyanogenic Glucosides Lotaustralin and Linamarin in Lotus japonicus. Plant Physiol. 2004, 135, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Saito, S.; Motawia, M.S.; Olsen, C.E.; Møller, B.L.; Bak, S. Biosynthesis of Rhodiocyanosides in Lotus japonicus: Rhodiocyanoside A Is Synthesized from (Z)-2-Methylbutanaloxime via 2-Methyl-2-Butenenitrile. Phytochemistry 2012, 77, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Tsikou, D.; Tsiknia, M.; Nikolaou, C.N.; Ehaliotis, C.; Papadopoulou, K.K. The Effect of Rhizophagus irregularis and Mesorhizobium Loti Co-Inoculation on Lotus japonicus. J. Exp. Mol. Biol. 2019, 20, 1–6. [Google Scholar]
- Hewitt, E.J. Sand and Water Culture Methods Used in the Study of Plant Nutrition. In Commonwealth Bureau of Horticulture and Plantation Crops. East Malling, England. Technical Communications, 2nd ed.; Commonwealth Agricultural Bureaux: Farnham Royal, UK, 1966. [Google Scholar]
- Hoagland, D.R. The Water-Culture Method for Growing Plants without Soil; Arnon, D.I., Ed.; College of Agriculture, University of California: Berkeley, CA, USA, 1950. [Google Scholar]
- Vierheilig, H.; Coughlan, A.P.; Wyss, U.; Piché, Y. Ink and Vinegar, a Simple Staining Technique for Arbuscular-Mycorrhizal Fungi. Appl. Environ. Microbiol. 1998, 64, 5004–5007. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Kalousi, F.D.; Pollastro, F.; Christodoulou, E.C.; Karra, A.G.; Tsialtas, I.; Georgantopoulos, A.; Salamone, S.; Psarra, A.-M.G. Apoptotic, Anti-Inflammatory Activities and Interference with the Glucocorticoid Receptor Signaling of Fractions from Pistacia lentiscus L. Var. Chia Leaves. Plants 2022, 11, 934. [Google Scholar] [CrossRef]
- Bunim, J.J. A Decade of Anti-Inflammatory Steroids, from Cortisone to Dexamethasone. Summary. Ann. N. Y Acad. Sci. 1959, 82, 1012–1013. [Google Scholar] [CrossRef]
- Liu, C.-W.; Murray, J.D. The Role of Flavonoids in Nodulation Host-Range Specificity: An Update. Plants 2016, 5, 33. [Google Scholar] [CrossRef]
- Tian, B.; Pei, Y.; Huang, W.; Ding, J.; Siemann, E. Increasing Flavonoid Concentrations in Root Exudates Enhance Associations between Arbuscular Mycorrhizal Fungi and an Invasive Plant. ISME J. 2021, 15, 1919–1930. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Siemann, E.; Tian, B.; Ding, J. Root Flavonoids Are Related to Enhanced AMF Colonization of an Invasive Tree. AoB Plants 2020, 12, plaa002. [Google Scholar] [CrossRef]
- Veitch, N.C. Isoflavonoids of the Leguminosae. Nat. Prod. Rep. 2007, 24, 417–464. [Google Scholar] [CrossRef]
- Vitale, D.C.; Piazza, C.; Melilli, B.; Drago, F.; Salomone, S. Isoflavones: Estrogenic Activity, Biological Effect and Bioavailability. Eur. J. Drug Metab. Pharmacokinet. 2013, 38, 15–25. [Google Scholar] [CrossRef]
- Maleki, S.J.; Crespo, J.F.; Cabanillas, B. Anti-Inflammatory Effects of Flavonoids. Food Chem. 2019, 299, 125124. [Google Scholar] [CrossRef]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Al-Mana, F.A.; Mahmoud, E.A.; Zin El-Abedin, T.K.A.; Mattar, M.A.; Ekiert, H. Phenolic Compounds of Catalpa speciosa, Taxus cuspidate, and Magnolia acuminata Have Antioxidant and Anticancer Activity. Molecules 2019, 24, 412. [Google Scholar] [CrossRef]
- Kim, K.M.; Heo, D.R.; Kim, Y.-A.; Lee, J.; Kim, N.S.; Bang, O.-S. Coniferaldehyde Inhibits LPS-Induced Apoptosis through the PKC α/β II/Nrf-2/HO-1 Dependent Pathway in RAW264.7 Macrophage Cells. Environ. Toxicol. Pharmacol. 2016, 48, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.-J.; Lu, L.; Yang, J.-J.; Wang, X.-X.; Su, G.; Wang, Z.-L.; Chen, G.-H.; Sun, H.-M.; Wang, M.-Y.; Yang, Y. Lariciresinol Induces Apoptosis in HepG2 Cells via Mitochondrial-Mediated Apoptosis Pathway. Eur. J. Pharmacol. 2018, 821, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ulanowska, M.; Olas, B. Biological Properties and Prospects for the Application of Eugenol—A Review. Int. J. Mol. Sci. 2021, 22, 3671. [Google Scholar] [CrossRef] [PubMed]
- Mlala, S.; Oyedeji, A.O.; Gondwe, M.; Oyedeji, O.O. Ursolic Acid and Its Derivatives as Bioactive Agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef] [PubMed]
- Tay, K.-C.; Tan, L.T.-H.; Chan, C.K.; Hong, S.L.; Chan, K.-G.; Yap, W.H.; Pusparajah, P.; Lee, L.-H.; Goh, B.-H. Formononetin: A Review of Its Anticancer Potentials and Mechanisms. Front. Pharmacol. 2019, 10, 820. [Google Scholar] [CrossRef] [PubMed]
- Madadi, E.; Mazloum-Ravasan, S.; Yu, J.S.; Ha, J.W.; Hamishehkar, H.; Kim, K.H. Therapeutic Application of Betalains: A Review. Plants 2020, 9, 1219. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wang, W.; He, Q.; Wu, Y.; Lu, Z.; Sun, J.; Liu, Z.; Shao, Y.; Wang, A. Oleic Acid Induces Apoptosis and Autophagy in the Treatment of Tongue Squamous Cell Carcinomas. Sci. Rep. 2017, 7, 11277. [Google Scholar] [CrossRef]
- Palomino, O.M.; Giordani, V.; Chowen, J.; Alfonso, S.F.; Goya, L. Physiological Doses of Oleic and Palmitic Acids Protect Human Endothelial Cells from Oxidative Stress. Molecules 2022, 27, 5217. [Google Scholar] [CrossRef]
- Vandewalle, J.; Luypaert, A.; De Bosscher, K.; Libert, C. Therapeutic Mechanisms of Glucocorticoids. Trends Endocrinol. Metab. 2018, 29, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, S.; Kamal, A.; Khan, F.; Jamali, K.S.; Saify, Z.S. Gallic and Vanillic Acid Suppress Inflammation and Promote Myelination in an In Vitro Mouse Model of Neurodegeneration. Mol. Biol. Rep. 2019, 46, 997–1011. [Google Scholar] [CrossRef] [PubMed]
- Sozen, E.; Demirel, T.; Ozer, N.K. Vitamin E: Regulatory Role in the Cardiovascular System. IUBMB Life 2019, 71, 507–515. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Metabolites | Shoot Extracts | Root Extracts | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| * Control | AMF | AMF-D | R | R + D | Control | AMF | AMF-D | R | R + D | |
| Phenolics | ||||||||||
| vanillic acid | 1 | 0.4 | 1.1 | 1.2 | 0.9 | 1 | 0.5 | 0.2 | 3.0 | 1.5 |
| apigenin 7-glucoside | 1 | 0.4 | 0.3 | 5.4 | 3.4 | 1 | 1.9 | 2.1 | 0.4 | 2.0 |
| cyanidin3-rytinoside | 1 | 0.4 | 0.1 | 2.4 | 1.7 | 1 | 1.3 | 0.7 | 0.6 | 6.8 |
| hydroxycaffeic acid | 1 | 0.6 | 1.1 | 3.4 | 1.7 | 1 | 0.0 | 0.4 | 10.7 | 7.4 |
| isoorientin | 1 | 0.6 | 0.5 | 3.2 | 0.8 | 1 | 0.9 | 0.8 | 1.2 | 0.9 |
| isorhoifolin | 1 | 0.3 | 0.3 | 5.1 | 2.9 | 1 | 0.5 | 0.2 | 4.6 | 3.1 |
| lariciresinol | 1 | 0.6 | 0.6 | 0.6 | 1.0 | 1 | 1.3 | 2.5 | 1.7 | 5.3 |
| luteolin-5-glucoside | 1 | 0.6 | 0.5 | 3.2 | 0.8 | 1 | 0.9 | 0.8 | 1.2 | 0.9 |
| quercetin-3-rutinoside | 1 | 0.2 | 0.3 | 4.7 | 1.3 | N/A | N/A | N/A | N/A | N/A |
| gamma tocopherol | 1 | 0.0 | 1.0 | 7.2 | 1.1 | 1 | 0.1 | 0.2 | 2.3 | 2.3 |
| m-coumaric acid | 1 | 0.6 | 0.3 | 1.8 | 1.0 | 1 | 0.9 | 0.9 | 19.5 | 8.2 |
| maachiain | 1 | 0.1 | 0.3 | 0.4 | 0.4 | 1 | 0.2 | 0.2 | 0.5 | 11.3 |
| glycinol | 1 | 1.3 | 1.2 | 1.2 | 1.2 | 1 | 1.0 | 0.9 | 1.1 | 1.0 |
| phaseollin | 1 | 0.6 | 3.5 | 1.8 | 1 | 0.1 | 0.0 | 0.4 | 2.8 | |
| vestitol | 1 | 0.0 | 0.4 | 0.1 | 0.2 | 1 | 0.1 | 0.1 | 16.8 | 4.5 |
| sophorol | 1 | 0.0 | 0.6 | 0.4 | 1.5 | 1 | 0.1 | 0.1 | 0.3 | 8.5 |
| (6aR,11aR)-3,9-Dihydroxypterocarpan | 1 | 0.2 | 0.3 | 0.7 | 0.2 | 1 | 1.0 | 0.2 | 6.6 | 2.4 |
| 2′-Hydroxyformononetin | 1 | 0.1 | 0.3 | 0.4 | 0.4 | 1 | 0.2 | 0.2 | 0.5 | 11.3 |
| astragalin | 1 | 0.6 | 0.5 | 3.2 | 0.8 | 1 | 0.9 | 0.8 | 1.2 | 0.9 |
| chalconaringenin | 1 | 1.3 | 1.2 | 1.2 | 1.2 | 1 | 1.0 | 0.9 | 1.1 | 1.0 |
| delphinidin | 1 | 0.1 | 0.1 | 0.1 | 0.1 | 1 | 0.3 | 0.1 | 0.1 | 0.0 |
| eugenol | 1 | 60.9 | 72.2 | 80.8 | 79.1 | 1 | 1.0 | 1.2 | 2.2 | 1.6 |
| formononetin | 1 | 0.0 | 0.4 | 0.2 | 0.3 | 1 | 0.3 | 0.0 | 2.8 | 1.6 |
| genistein | 1 | 0.2 | 0.1 | 0.1 | 0.1 | 1 | 0.4 | 0.2 | 0.6 | 0.4 |
| genistin | 1 | 0.7 | 0.5 | 0.0 | 2.1 | 1 | 1.9 | 2.1 | 1.7 | 2.0 |
| homogentistic acid | 1 | 0.4 | 1.1 | 1.2 | 0.9 | 1 | 0.5 | 0.2 | 3.0 | 1.5 |
| leucopelargonidin | 1 | 5.2 | 4.6 | 4.4 | 4.3 | 1 | 1.0 | 1.1 | 1.0 | 1.0 |
| lotaustralin | 1 | 0.3 | 0.7 | 1.0 | 1.8 | 1 | 0.4 | 0.1 | 8.0 | 9.5 |
| pelargonidin | 1 | 0.2 | 0.1 | 0.1 | 0.1 | 1 | 0.4 | 0.2 | 0.6 | 0.0 |
| phenylalcetaldehyde | 1 | 0.6 | 0.6 | 6.2 | 1.3 | 1 | 0.1 | 0.0 | 12.6 | 5.8 |
| sinapaldehyde | 1 | 0.8 | 0.9 | 0.4 | 0.8 | 1 | 1.0 | 1.0 | 1.0 | 0.6 |
| vestitone | 1 | 0.8 | 0.7 | 0.6 | 0.7 | 1 | 0.9 | 0.8 | 1.0 | 0.9 |
| coniferyl acetate | 1 | 1.2 | 1.1 | 1.2 | 1.1 | 1 | 1.0 | 1.0 | 1.1 | 1.1 |
| astragalin | 1 | 0.6 | 0.5 | 3.2 | 0.8 | 1 | 0.9 | 0.8 | 1.2 | 0.9 |
| caffeic acid hexoside | 1 | 0.6 | 1.1 | 3.4 | 1.7 | 1 | 0.0 | 0.4 | 10.7 | 7.4 |
| coniferaldehyde | 1 | 0.5 | 0.4 | 3.1 | 1.6 | 1 | 0.9 | 0.1 | 3.2 | 4.5 |
| diosmetin | 1 | 0.0 | 0.6 | 0.4 | 1.5 | 1 | 0.1 | 0.1 | 0.3 | 8.5 |
| eriodictyol | N/A | N/A | N/A | 1 | N/A | N/A | N/A | N/A | 5.3 | N/A |
| N-containing compounts | ||||||||||
| caffeine | 1 | 0.5 | 0.4 | 1.4 | 2.2 | 1 | 0.3 | 0.3 | 2.1 | 3.5 |
| cinchonidine | 1 | 1.1 | 1.0 | 1.2 | 1.0 | 1 | 1.1 | 0.4 | 0.1 | 1.1 |
| theobromine | 1 | 0.8 | 0.1 | 0.2 | 0.8 | 1 | 0.2 | 0.3 | 0.8 | 2.2 |
| portulacaxanthin | 1 | 4.0 | 9.0 | 26.2 | 0.5 | 1 | 0.6 | 2.4 | 8.4 | 1.2 |
| Terpenes | ||||||||||
| oleanolic acid | 1 | 0.3 | 0.6 | 2.4 | 0.9 | 1 | 0.1 | 0.0 | 1.8 | 0.7 |
| ursolic acid | 1 | 0.3 | 0.6 | 2.4 | 0.9 | 1 | 0.1 | 0.0 | 1.8 | 0.7 |
| menthone | 1 | 0.4 | 0.7 | 2.0 | 1.1 | 1 | 0.7 | 0.8 | 0.9 | 0.8 |
| Sterols | ||||||||||
| desmosterol | 1 | 1.2 | 1.3 | 0.7 | 0.6 | 1 | 1.2 | 1.0 | 0.6 | 0.5 |
| Fatty acids | ||||||||||
| myristoleic | 1 | 0.4 | 0.5 | 0.6 | 1.8 | 1 | 0.7 | 0.7 | 1.3 | 8.0 |
| octadecanoic acid | 1 | 0.6 | 0.8 | 0.7 | 0.7 | 1 | 1.1 | 0.8 | 0.8 | 1.8 |
| oleic acid | 1 | 0.6 | 0.5 | 0.8 | 0.6 | 1 | 0.7 | 1.1 | 2.9 | 2.0 |
| palmitoleic acid | 1 | 0.7 | 0.7 | 0.6 | 1.4 | 1 | 1.1 | 1.0 | 1.4 | 6.9 |
| pentadecanoic acid | 1 | 0.8 | 0.8 | 0.6 | 1.2 | 1 | 0.9 | 0.7 | 1.1 | 2.8 |
| glycerol tributanoate | 1 | 0.5 | 0.5 | 0.7 | 1.0 | 1 | 1.0 | 2.6 | 2.3 | 5.9 |
| Metabolites | Root/Shoot Ratios | ||||
|---|---|---|---|---|---|
| * Control | AMF | AMF-D | R | R + D | |
| Phenolics | |||||
| vanillic acid | 10.4 | 14.4 | 2.1 | 27.2 | 17.0 |
| apigenin 7-glucoside | 0.3 | 1.8 | 2.5 | 0.0 | 0.2 |
| cyanidin3-rytinoside | 0.1 | 0.5 | 0.9 | 0.0 | 0.6 |
| hydroxycaffeic acid | 0.3 | 0.0 | 0.1 | 0.9 | 1.3 |
| isoorientin | 0.5 | 0.8 | 0.9 | 0.2 | 0.6 |
| isorhoifolin | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| lariciresinol | 0.7 | 1.6 | 2.7 | 1.9 | 3.4 |
| luteolin-5-glucoside | 0.5 | 0.8 | 0.9 | 0.2 | 0.6 |
| quercetin-3-rutinoside | Shoot sp. | Shoot sp. | Shoot sp. | Shoot sp. | Shoot sp. |
| gamma tocopherol | 20.0 | 123.1 | 5.0 | 6.4 | 42.0 |
| m-coumaric acid | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| maachiain | 1.0 | 1.8 | 0.5 | 1.3 | 26.1 |
| glycinol | 1.1 | 0.9 | 0.9 | 1.0 | 1.0 |
| phaseollin | 94.4 | 14.3 | 10.1 | 149.3 | |
| vestitol | 0.0 | 0.0 | 0.0 | 0.5 | 0.1 |
| sophorol | 0.7 | 1.5 | 0.1 | 0.5 | 4.0 |
| (6aR,11aR)-3,9-Dihydroxypterocarpan | 1.1 | 4.5 | 0.7 | 10.9 | 10.2 |
| 2′-Hydroxyformononetin | 1.0 | 1.8 | 0.5 | 1.3 | 26.1 |
| astragalin | 0.5 | 0.8 | 0.9 | 0.2 | 0.6 |
| chalconaringenin | 1.1 | 0.9 | 0.9 | 1.0 | 1.0 |
| delphinidin | 0.3 | 1.2 | 0.4 | 0.2 | 0.1 |
| eugenol | 49.0 | 0.8 | 0.8 | 1.3 | 1.0 |
| formononetin | 0.2 | 1.6 | 0.0 | 2.5 | 0.9 |
| genistein | 0.3 | 0.6 | 0.9 | 2.4 | 1.5 |
| genistin | 0.2 | 0.6 | 0.8 | 56.9 | 0.2 |
| homogentistic acid | 10.4 | 14.4 | 2.1 | 27.2 | 17.0 |
| leucopelargonidin | 4.7 | 0.9 | 1.1 | 1.1 | 1.1 |
| lotaustralin | 0.1 | 0.1 | 0.0 | 0.6 | 0.4 |
| pelargonidin | 0.3 | 0.6 | 0.9 | 2.4 | 0.1 |
| phenylalcetaldehyde | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 |
| sinapaldehyde | 0.8 | 1.0 | 1.0 | 2.0 | 0.6 |
| vestitone | 0.8 | 0.9 | 1.0 | 1.3 | 1.1 |
| coniferyl acetate | 1.1 | 0.9 | 1.0 | 1.1 | 1.1 |
| astragalin | 0.5 | 0.8 | 0.9 | 0.2 | 0.6 |
| caffeic acid hexoside | 0.3 | 0.0 | 0.1 | 0.9 | 1.3 |
| coniferaldehyde | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| diosmetin | 0.7 | 1.5 | 0.1 | 0.5 | 4.0 |
| eriodictyol | - | - | - | 5.4 | - |
| N-containing compounts | |||||
| caffeine | 0.5 | 0.3 | 0.3 | 0.8 | 0.8 |
| cinchonidine | 0.9 | 1.0 | 0.3 | 0.1 | 1.0 |
| theobromine | 0.6 | 0.1 | 1.2 | 2.2 | 1.6 |
| portulacaxanthin | 0.5 | 0.1 | 0.1 | 0.1 | 1.2 |
| Terpenes | |||||
| oleanolic acid | 1.9 | 0.6 | 0.0 | 1.4 | 1.6 |
| ursolic acid | 1.9 | 0.6 | 0.0 | 1.4 | 1.6 |
| menthone | 0.6 | 1.1 | 0.6 | 0.3 | 0.4 |
| Sterols | |||||
| desmosterol | 1.3 | 1.4 | 1.1 | 1.2 | 1.0 |
| Fatty acids | |||||
| myristoleic | 0.9 | 1.5 | 1.2 | 2.0 | 3.9 |
| octadecanoic acid | 0.9 | 1.6 | 1.0 | 1.0 | 2.3 |
| oleic acid | 1.0 | 1.1 | 2.0 | 3.5 | 3.2 |
| palmitoleic acid | 0.9 | 1.4 | 1.3 | 1.9 | 4.4 |
| pentadecanoic acid | 1.3 | 1.4 | 1.1 | 2.3 | 3.2 |
| glycerol tributanoate | 0.5 | 1.2 | 2.8 | 1.7 | 3.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalousi, F.D.; Tsakos, M.; Nikolaou, C.N.; Georgantopoulos, A.; Psarra, A.-M.G.; Tsikou, D. Chemical Analysis and Biological Activities of Extracts Isolated from Symbiotic L. japonicus Plants. Life 2024, 14, 189. https://doi.org/10.3390/life14020189
Kalousi FD, Tsakos M, Nikolaou CN, Georgantopoulos A, Psarra A-MG, Tsikou D. Chemical Analysis and Biological Activities of Extracts Isolated from Symbiotic L. japonicus Plants. Life. 2024; 14(2):189. https://doi.org/10.3390/life14020189
Chicago/Turabian StyleKalousi, Foteini D., Michail Tsakos, Christina N. Nikolaou, Achilleas Georgantopoulos, Anna-Maria G. Psarra, and Daniela Tsikou. 2024. "Chemical Analysis and Biological Activities of Extracts Isolated from Symbiotic L. japonicus Plants" Life 14, no. 2: 189. https://doi.org/10.3390/life14020189