
Adiponectin Gene Polymorphisms: A Case–Control Study on Their Role in Late-Onset Alzheimer’s Disease Risk
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Sample Collection and Processing
2.3. SNP Selection and Genotyping
2.4. Measurement of Plasma Adiponectin Levels
2.5. Statistical Analyses
3. Results
3.1. Characteristics of Study Subjects
3.2. Association of ADIPOQ Polymorphisms with LOAD Risk
3.3. Association of ADIPOQ Polymorphisms with Plasma Adiponectin Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alzheimer’s Association. 2022 Alzheimer’s disease facts and figures. Alzheimers Dement. 2022, 18, 700–789. [Google Scholar] [CrossRef]
- Andrade-Guerrero, J.; Santiago-Balmaseda, A.; Jeronimo-Aguilar, P.; Vargas-Rodríguez, I.; Cadena-Suárez, A.R.; Sánchez-Garibay, C.; Pozo-Molina, G.; Méndez-Catalá, C.F.; Cardenas-Aguayo, M.D.; Diaz-Cintra, S.; et al. Alzheimer’s Disease: An Updated Overview of Its Genetics. Int. J. Mol. Sci. 2023, 24, 3754. [Google Scholar] [CrossRef]
- Uddin, M.S.; Rahman, M.M.; Sufian, M.A.; Jeandet, P.; Ashraf, G.M.; Bin-Jumah, M.N.; Mousa, S.A.; Abdel-Daim, M.M.; Akhtar, M.F.; Saleem, A.; et al. Exploring the New Horizon of AdipoQ in Obesity-Related Alzheimer’s Dementia. Front. Physiol. 2021, 11, 567678. [Google Scholar] [CrossRef]
- Armstrong, R.A. Risk factors for Alzheimer’s disease. Folia Neuropathol. 2019, 57, 87–105. [Google Scholar] [CrossRef]
- Andrews, S.J.; Renton, A.E.; Fulton-Howard, B.; Podlesny-Drabiniok, A.; Marcora, E.; Goate, A.M. The complex genetic architecture of Alzheimer’s disease: Novel insights and future directions. eBioMedicine 2023, 90, 104511. [Google Scholar] [CrossRef]
- Wightman, D.P.; Jansen, I.E.; Savage, J.E.; Shadrin, A.A.; Bahrami, S.; Holland, D.; Rongve, A.; Børte, S.; Winsvold, B.S.; Drange, O.K.; et al. A genome-wide association study with 1,126,563 individuals identifies new risk loci for Alzheimer’s disease. Nat. Genet. 2021, 53, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Bellenguez, C.; Küçükali, F.; Jansen, I.E.; Kleineidam, L.; Moreno-Grau, S.; Amin, N.; Naj, A.C.; Campos-Martin, R.; Grenier-Boley, B.; Andrade, V.; et al. New insights into the genetic etiology of Alzheimer’s disease and related dementias. Nat. Genet. 2022, 54, 412–436. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.C.; Ramirez, A.; Grenier-Boley, B.; Bellenguez, C. Step by step: Towards a better understanding of the genetic architecture of Alzheimer’s disease. Mol. Psychiatry 2023, 28, 2716–2727. [Google Scholar] [CrossRef] [PubMed]
- Flores-Cordero, J.A.; Pérez-Pérez, A.; Jiménez-Cortegana, C.; Alba, G.; Flores-Barragán, A.; Sánchez-Margalet, V. Obesity as a Risk Factor for Dementia and Alzheimer’s Disease: The Role of Leptin. Int. J. Mol. Sci. 2022, 23, 5202. [Google Scholar] [CrossRef] [PubMed]
- Neto, A.; Fernandes, A.; Barateiro, A. The complex relationship between obesity and neurodegenerative diseases: An updated review. Front. Cell. Neurosci. 2023, 17, 1294420. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xu, F.; Wang, L.; Zhang, W.D.; Sun, C.Q.; Deng, H.W. Detecting potential causal relationship between multiple risk factors and Alzheimer’s disease using multivariable Mendelian randomization. Aging 2020, 12, 21747–21757. [Google Scholar] [CrossRef]
- Brenowitz, W.D.; Zimmerman, S.C.; Filshtein, T.J.; Yaffe, K.; Walter, S.; Hoffmann, T.J.; Jorgenson, E.; Whitmer, R.A.; Glymour, M.M. Extension of Mendelian Randomization to Identify Earliest Manifestations of Alzheimer Disease: Association of Genetic Risk Score for Alzheimer Disease with Lower Body Mass Index by Age 50 Years. Am. J. Epidemiol. 2021, 190, 2163–2171. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Wu, W.; Ma, W.; Huang, X.; Huang, T.; Peng, M.; Xu, A.; Lyu, J. Body mass index, genetic susceptibility, and Alzheimer’s disease: A longitudinal study based on 475,813 participants from the UK Biobank. J. Transl. Med. 2022, 20, 417. [Google Scholar] [CrossRef] [PubMed]
- Korologou-Linden, R.; Bhatta, L.; Brumpton, B.M.; Howe, L.D.; Millard, L.A.C.; Kolaric, K.; Ben-Shlomo, Y.; Williams, D.M.; Smith, G.D.; Anderson, E.L.; et al. The causes and consequences of Alzheimer’s disease: Phenome-wide evidence from Mendelian randomization. Nat. Commun. 2022, 13, 4726. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.Y.; Ou, Y.N.; Yang, Y.X.; Wang, Z.T.; Tan, L.; Yu, J.T. Associations of cardiovascular risk factors and lifestyle behaviors with neurodegenerative disease: A Mendelian randomization study. Transl. Psychiatry 2023, 13, 267. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Feng, J.; Jiang, S.; Guo, J.; Zhang, X.; Zhang, X.; Wang, C.; Ma, Y.; Dong, Z. Mendelian randomization analyses identify bidirectional causal relationships of obesity with psychiatric disorders. J. Affect. Disord. 2023, 339, 807–814. [Google Scholar] [CrossRef] [PubMed]
- European Alzheimer’s & Dementia Biobank Mendelian Randomization (EADB-MR) Collaboration; Luo, J.; Thomassen, J.Q.; Bellenguez, C.; Grenier-Boley, B.; de Rojas, I.; Castillo, A.; Parveen, K.; Küçükali, F.; Nicolas, A.; et al. Genetic Associations Between Modifiable Risk Factors and Alzheimer Disease. JAMA Netw. Open 2023, 6, e2313734. [Google Scholar] [CrossRef]
- Forny-Germano, L.; De Felice, F.G.; Vieira, M.N.D.N. The Role of Leptin and Adiponectin in Obesity-Associated Cognitive Decline and Alzheimer’s Disease. Front. Neurosci. 2019, 12, 1027. [Google Scholar] [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef]
- Lei, X.; Qiu, S.; Yang, G.; Wu, Q. Adiponectin and metabolic cardiovascular diseases: Therapeutic opportunities and challenges. Genes Dis. 2022, 10, 1525–1536. [Google Scholar] [CrossRef]
- Chen, R.; Shu, Y.; Zeng, Y. Links between Adiponectin and Dementia: From Risk Factors to Pathophysiology. Front. Aging Neurosci. 2020, 11, 356. [Google Scholar] [CrossRef] [PubMed]
- Dezonne, R.S.; Pereira, C.M.; de Moraes Martins, C.J.; de Abreu, V.G.; Francischetti, E.A. Adiponectin, the adiponectin paradox, and Alzheimer’s Disease: Is this association biologically plausible? Metab. Brain Dis. 2023, 38, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Bloemer, J.; Pinky, P.D.; Govindarajulu, M.; Hong, H.; Judd, R.; Amin, R.H.; Moore, T.; Dhanasekaran, M.; Reed, M.N.; Suppiramaniam, V. Role of Adiponectin in Central Nervous System Disorders. Neural Plast. 2018, 2018, 4593530. [Google Scholar] [CrossRef] [PubMed]
- Letra, L.; Rodrigues, T.; Matafome, P.; Santana, I.; Seiça, R. Adiponectin and sporadic Alzheimer’s disease: Clinical and molecular links. Front. Neuroendocrinol. 2019, 52, 1–11. [Google Scholar] [CrossRef]
- Ng, R.C.; Jian, M.; Ma, O.K.; Bunting, M.; Kwan, J.S.; Zhou, G.J.; Senthilkumar, K.; Iyaswamy, A.; Chan, P.K.; Li, M.; et al. Chronic oral administration of adipoRon reverses cognitive impairments and ameliorates neuropathology in an Alzheimer’s disease mouse model. Mol. Psychiatry 2021, 26, 5669–5689. [Google Scholar] [CrossRef]
- Rizzo, M.R.; Fasano, R.; Paolisso, G. Adiponectin and Cognitive Decline. Int. J. Mol. Sci. 2020, 21, 2010. [Google Scholar] [CrossRef]
- García-García, I.; Fernández-Andújar, M.; Narváez, M.; García-Casares, N. Assessing adipokines as potential biomarkers of dementia, Alzheimer’s disease, and mild cognitive impairment: A systematic review and meta-analysis. Obes. Rev. 2023, 24, e13573. [Google Scholar] [CrossRef]
- Wennberg, A.M.; Gustafson, D.; Hagen, C.E.; Roberts, R.O.; Knopman, D.; Jack, C.; Petersen, R.C.; Mielke, M.M. Serum Adiponectin Levels, Neuroimaging, and Cognition in the Mayo Clinic Study of Aging. J. Alzheimers Dis. 2016, 53, 573–581. [Google Scholar] [CrossRef]
- Kim, J.W.; Byun, M.S.; Yi, D.; Kim, M.J.; Jung, J.H.; Kong, N.; Jung, G.; Ahn, H.; Lee, J.Y.; Kang, K.M.; et al. Serum Adiponectin and In Vivo Brain Amyloid Deposition in Cognitively Normal Older Adults: A Cohort Study. Aging Dis. 2023, 14, 904–918. [Google Scholar] [CrossRef]
- Waragai, M.; Ho, G.; Takamatsu, Y.; Wada, R.; Sugama, S.; Takenouchi, T.; Masliah, E.; Hashimoto, M. Adiponectin Paradox in Alzheimer’s Disease; Relevance to Amyloidogenic Evolvability? Front. Endocrinol. 2020, 11, 108. [Google Scholar] [CrossRef]
- Waragai, M.; Ho, G.; Takamatsu, Y.; Wada, R.; Sugama, S.; Takenouchi, T.; Masliah, E.; Hashimoto, M. Adiponectin Paradox as a Therapeutic Target in Alzheimer’s Disease. J. Alzheimer’s Dis. 2020, 76, 1249–1253. [Google Scholar] [CrossRef]
- Comuzzie, A.G.; Funahashi, T.; Sonnenberg, G.; Martin, L.J.; Jacob, H.J.; Black, A.E.; Maas, D.; Takahashi, M.; Kihara, S.; Tanaka, S.; et al. The genetic basis of plasma variation in adiponectin, a global endophenotype for obesity and the metabolic syndrome. J. Clin. Endocrinol. Metab. 2001, 86, 4321–4325. [Google Scholar] [CrossRef]
- Vasseur, F.; Helbecque, N.; Dina, C.; Lobbens, S.; Delannoy, V.; Gaget, S.; Boutin, P.; Vaxillaire, M.; Leprêtre, F.; Dupont, S.; et al. Single-nucleotide polymorphism haplotypes in the both proximal promoter and exon 3 of the APM1 gene modulate adipocyte-secreted adiponectin hormone levels and contribute to the genetic risk for type 2 diabetes in French Caucasians. Hum. Mol. Genet. 2002, 11, 2607–2614. [Google Scholar] [CrossRef]
- Lindsay, R.S.; Funahashi, T.; Krakoff, J.; Matsuzawa, Y.; Tanaka, S.; Kobes, S.; Bennett, P.H.; Tataranni, P.A.; Knowler, W.C.; Hanson, R.L. Genome-wide linkage analysis of serum adiponectin in the Pima Indian population. Diabetes 2003, 52, 2419–2425. [Google Scholar] [CrossRef]
- Ling, H.; Waterworth, D.M.; Stirnadel, H.A.; Pollin, T.I.; Barter, P.J.; Kesäniemi, Y.A.; Mahley, R.W.; McPherson, R.; Waeber, G.; Bersot, T.P.; et al. Genome-wide linkage and association analyses to identify genes influencing adiponectin levels: The GEMS Study. Obesity 2009, 17, 737–744. [Google Scholar] [CrossRef]
- Heid, I.M.; Henneman, P.; Hicks, A.; Coassin, S.; Winkler, T.; Aulchenko, Y.S.; Fuchsberger, C.; Song, K.; Hivert, M.F.; Waterworth, D.M.; et al. Clear detection of ADIPOQ locus as the major gene for plasma adiponectin: Results of genome-wide association analyses including 4659 European individuals. Atherosclerosis 2010, 208, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Henneman, P.; Aulchenko, Y.S.; Frants, R.R.; Zorkoltseva, I.V.; Zillikens, M.C.; Frolich, M.; Oostra, B.A.; van Dijk, K.W.; van Duijn, C.M. Genetic architecture of plasma adiponectin overlaps with the genetics of metabolic syndrome-related traits. Diabetes Care 2010, 33, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.B.; Waterworth, D.; O’Rahilly, S.; Hivert, M.F.; Loos, R.J.; Perry, J.R.; Tanaka, T.; Timpson, N.J.; Semple, R.K.; Soranzo, N.; et al. A genome-wide association study reveals variants in ARL15 that influence adiponectin levels. PLoS Genet. 2009, 5, e1000768. [Google Scholar] [CrossRef]
- Jee, S.H.; Sull, J.W.; Lee, J.E.; Shin, C.; Park, J.; Kimm, H.; Cho, E.Y.; Shin, E.S.; Yun, J.E.; Park, J.W.; et al. Adiponectin concentrations: A genome-wide association study. Am. J. Hum. Genet. 2010, 87, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, Y.; Lange, E.M.; Croteau-Chonka, D.C.; Kuzawa, C.W.; McDade, T.W.; Qin, L.; Curocichin, G.; Borja, J.B.; Lange, L.A.; et al. Genome-wide association study for adiponectin levels in Filipino women identifies CDH13 and a novel uncommon haplotype at KNG1-ADIPOQ. Hum. Mol. Genet. 2010, 19, 4955–4964. [Google Scholar] [CrossRef] [PubMed]
- Dastani, Z.; Hivert, M.F.; Timpson, N.; Perry, J.R.; Yuan, X.; Scott, R.A.; Henneman, P.; Heid, I.M.; Kizer, J.R.; Lyytikäinen, L.P.; et al. Novel loci for adiponectin levels and their influence on type 2 diabetes and metabolic traits: A multi-ethnic meta-analysis of 45,891 individuals. PLoS Genet. 2012, 8, e1002607. [Google Scholar] [CrossRef]
- Reilly, D.; Hao, K.; Jensen, M.K.; Girman, C.J.; Rimm, E.B. Use of systems biology approaches to analysis of genome-wide association studies of myocardial infarction and blood cholesterol in the nurses’ health study and health professionals’ follow-up study. PLoS ONE 2013, 8, e85369. [Google Scholar] [CrossRef]
- Wu, Y.; Gao, H.; Li, H.; Tabara, Y.; Nakatochi, M.; Chiu, Y.F.; Park, E.J.; Wen, W.; Adair, L.S.; Borja, J.B.; et al. A meta-analysis of genome-wide association studies for adiponectin levels in East Asians identifies a novel locus near WDR11-FGFR2. Hum. Mol. Genet. 2014, 23, 1108–1119. [Google Scholar] [CrossRef]
- Li, W.D.; Jiao, H.; Wang, K.; Yang, F.; Grant, S.F.; Hakonarson, H.; Ahima, R.; Arlen Price, R. Pathway-Based Genome-wide Association Studies Reveal That the Rac1 Pathway Is Associated with Plasma Adiponectin Levels. Sci. Rep. 2015, 5, 13422. [Google Scholar] [CrossRef] [PubMed]
- Suhre, K.; Arnold, M.; Bhagwat, A.M.; Cotton, R.J.; Engelke, R.; Raffler, J.; Sarwath, H.; Thareja, G.; Wahl, A.; DeLisle, R.K.; et al. Connecting genetic risk to disease end points through the human blood plasma proteome. Nat. Commun. 2017, 8, 14357. [Google Scholar] [CrossRef]
- Hivert, M.F.; Scholtens, D.M.; Allard, C.; Nodzenski, M.; Bouchard, L.; Brisson, D.; Lowe, L.P.; McDowell, I.; Reddy, T.; Dastani, Z.; et al. Genetic determinants of adiponectin regulation revealed by pregnancy. Obesity 2017, 25, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Spracklen, C.N.; Karaderi, T.; Yaghootkar, H.; Schurmann, C.; Fine, R.S.; Kutalik, Z.; Preuss, M.H.; Lu, Y.; Wittemans, L.B.L.; Adair, L.S.; et al. Exome-Derived Adiponectin-Associated Variants Implicate Obesity and Lipid Biology. Am. J. Hum. Genet. 2019, 105, 15–28. [Google Scholar] [CrossRef]
- Spracklen, C.N.; Iyengar, A.K.; Vadlamudi, S.; Raulerson, C.K.; Jackson, A.U.; Brotman, S.M.; Wu, Y.; Cannon, M.E.; Davis, J.P.; Crain, A.T.; et al. Adiponectin GWAS loci harboring extensive allelic heterogeneity exhibit distinct molecular consequences. PLoS Genet. 2020, 16, e1009019. [Google Scholar] [CrossRef] [PubMed]
- Pietzner, M.; Wheeler, E.; Carrasco-Zanini, J.; Cortes, A.; Koprulu, M.; Wörheide, M.A.; Oerton, E.; Cook, J.; Stewart, I.D.; Kerrison, N.D.; et al. Mapping the proteo-genomic convergence of human diseases. Science 2021, 374, eabj1541. [Google Scholar] [CrossRef]
- Thareja, G.; Belkadi, A.; Arnold, M.; Albagha, O.M.E.; Graumann, J.; Schmidt, F.; Grallert, H.; Peters, A.; Gieger, C.; The Qatar Genome Program Research Consortium; et al. Differences and commonalities in the genetic architecture of protein quantitative trait loci in European and Arab populations. Hum. Mol. Genet. 2023, 32, 907–916. [Google Scholar] [CrossRef]
- Sarsani, V.; Brotman, S.M.; Xianyong, Y.; Fernandes Silva, L.; Laakso, M.; Spracklen, C.N. A cross-ancestry genome-wide meta-analysis, fine-mapping, and gene prioritization approach to characterize the genetic architecture of adiponectin. HGG Adv. 2024, 5, 100252. [Google Scholar] [CrossRef]
- Enns, J.E.; Taylor, C.G.; Zahradka, P. Variations in Adipokine Genes AdipoQ, Lep, and LepR are Associated with Risk for Obesity-Related Metabolic Disease: The Modulatory Role of Gene-Nutrient Interactions. J. Obes. 2011, 2011, 168659. [Google Scholar] [CrossRef]
- Breitfeld, J.; Stumvoll, M.; Kovacs, P. Genetics of adiponectin. Biochimie 2012, 94, 2157–2163. [Google Scholar] [CrossRef]
- Menzaghi, C.; Trischitta, V.; Doria, A. Genetic influences of adiponectin on insulin resistance, type 2 diabetes, and cardiovascular disease. Diabetes 2007, 56, 1198–1209. [Google Scholar] [CrossRef]
- Heid, I.M.; Wagner, S.A.; Gohlke, H.; Iglseder, B.; Mueller, J.C.; Cip, P.; Ladurner, G.; Reiter, R.; Stadlmayr, A.; Mackevics, V.; et al. Genetic architecture of the APM1 gene and its influence on adiponectin plasma levels and parameters of the metabolic syndrome in 1,727 healthy Caucasians. Diabetes 2006, 55, 375–384. [Google Scholar] [CrossRef]
- Hivert, M.F.; Manning, A.K.; McAteer, J.B.; Florez, J.C.; Dupuis, J.; Fox, C.S.; O’Donnell, C.J.; Cupples, L.A.; Meigs, J.B. Common variants in the adiponectin gene (ADIPOQ) associated with plasma adiponectin levels, type 2 diabetes, and diabetes-related quantitative traits: The Framingham Offspring Study. Diabetes 2008, 57, 3353–3359. [Google Scholar] [CrossRef] [PubMed]
- Siitonen, N.; Pulkkinen, L.; Lindström, J.; Kolehmainen, M.; Eriksson, J.G.; Venojärvi, M.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Tuomilehto, J.; Uusitupa, M. Association of ADIPOQ gene variants with body weight, type 2 diabetes and serum adiponectin concentrations: The Finnish Diabetes Prevention Study. BMC Med. Genet. 2011, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Warren, L.L.; Li, L.; Nelson, M.R.; Ehm, M.G.; Shen, J.; Fraser, D.J.; Aponte, J.L.; Nangle, K.L.; Slater, A.J.; Woollard, P.M.; et al. Deep resequencing unveils genetic architecture of ADIPOQ and identifies a novel low-frequency variant strongly associated with adiponectin variation. Diabetes 2012, 61, 1297–1301. [Google Scholar] [CrossRef]
- Mather, K.J.; Christophi, C.A.; Jablonski, K.A.; Knowler, W.C.; Goldberg, R.B.; Kahn, S.E.; Spector, T.; Dastani, Z.; Waterworth, D.; Richards, J.B.; et al. Common variants in genes encoding adiponectin (ADIPOQ) and its receptors (ADIPOR1/2), adiponectin concentrations, and diabetes incidence in the Diabetes Prevention Program. Diabet. Med. 2012, 29, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Peters, K.E.; Beilby, J.; Cadby, G.; Warrington, N.M.; Bruce, D.G.; Davis, W.A.; Davis, T.M.; Wiltshire, S.; Knuiman, M.; McQuillan, B.M.; et al. A comprehensive investigation of variants in genes encoding adiponectin (ADIPOQ) and its receptors (ADIPOR1/R2), and their association with serum adiponectin, type 2 diabetes, insulin resistance and the metabolic syndrome. BMC Med. Genet. 2013, 14, 15. [Google Scholar] [CrossRef] [PubMed]
- Sandy An, S.; Palmer, N.D.; Hanley, A.J.; Ziegler, J.T.; Mark Brown, W.; Freedman, B.I.; Register, T.C.; Rotter, J.I.; Guo, X.; Ida Chen, Y.D.; et al. Genetic analysis of adiponectin variation and its association with type 2 diabetes in African Americans. Obesity 2013, 21, E721–E729. [Google Scholar] [CrossRef] [PubMed]
- An, S.S.; Palmer, N.D.; Hanley, A.J.; Ziegler, J.T.; Brown, W.M.; Haffner, S.M.; Norris, J.M.; Rotter, J.I.; Guo, X.; Chen, Y.D.; et al. Estimating the contributions of rare and common genetic variations and clinical measures to a model trait: Adiponectin. Genet. Epidemiol. 2013, 37, 13–24. [Google Scholar] [CrossRef]
- Su, M.; Jia, A.; He, Y.; Song, Y. Associations of the Polymorphisms in ADIPOQ with Circulating Levels of Adiponectin and Lipids: A Meta-Analysis. Horm. Metab. Res. 2021, 53, 541–561. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Y.; Luo, Z. Effect of Adiponectin Variant on Lipid Profile and Plasma Adiponectin Levels: A Multicenter Systematic Review and Meta-Analysis. Cardiovasc. Ther. 2022, 2022, 4395266. [Google Scholar] [CrossRef]
- Li, W.; Yu, Z.; Hou, D.; Zhou, L.; Deng, Y.; Tian, M.; Feng, X. Relationship between Adiponectin Gene Polymorphisms and Late-Onset Alzheimer’s Disease. PLoS ONE 2015, 10, e0125186. [Google Scholar] [CrossRef]
- Yin, H.; Zhang, Y.; Hua, L.; Li, J.; Zeng, Z.; Yang, X.; Gong, B.; Geng, S.; Liu, Y.; Zhang, H.; et al. Relationship of polymorphisms and haplotype in interleukin-16 and adiponectin gene with late-onset Alzheimer’s disease risk. Oncotarget 2017, 8, 79157–79164. [Google Scholar] [CrossRef] [PubMed]
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E.M. Clinical diagnosis of Alzheimer’s disease: Report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984, 34, 939–944. [Google Scholar] [CrossRef]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]
- Sherry, S.T.; Ward, M.H.; Kholodov, M.; Baker, J.; Phan, L.; Smigielski, E.M.; Sirotkin, K. dbSNP: The NCBI database of genetic variation. Nucleic Acids Res. 2001, 29, 308–311. [Google Scholar] [CrossRef]
- Kyriakou, T.; Collins, L.J.; Spencer-Jones, N.J.; Malcolm, C.; Wang, X.; Snieder, H.; Swaminathan, R.; Burling, K.A.; Hart, D.J.; Spector, T.D.; et al. Adiponectin gene ADIPOQ SNP associations with serum adiponectin in two female populations and effects of SNPs on promoter activity. J. Hum. Genet. 2008, 53, 718–727. [Google Scholar] [CrossRef]
- Laumen, H.; Saningong, A.D.; Heid, I.M.; Hess, J.; Herder, C.; Claussnitzer, M.; Baumert, J.; Lamina, C.; Rathmann, W.; Sedlmeier, E.M.; et al. Functional characterization of promoter variants of the adiponectin gene complemented by epidemiological data. Diabetes 2009, 58, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Wassel, C.L.; Pankow, J.S.; Jacobs, D.R., Jr.; Steffes, M.W.; Li, N.; Schreiner, P.J. Variants in the adiponectin gene and serum adiponectin: The Coronary Artery Development in Young Adults (CARDIA) Study. Obesity 2010, 18, 2333–2338. [Google Scholar] [CrossRef] [PubMed]
- An, S.S.; Hanley, A.J.; Ziegler, J.T.; Brown, W.M.; Haffner, S.M.; Norris, J.M.; Rotter, J.I.; Guo, X.; Chen, Y.D.; Wagenknecht, L.E.; et al. Association between ADIPOQ SNPs with plasma adiponectin and glucose homeostasis and adiposity phenotypes in the IRAS Family Study. Mol. Genet. Metab. 2012, 107, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Ramya, K.; Ayyappa, K.A.; Ghosh, S.; Mohan, V.; Radha, V. Genetic association of ADIPOQ gene variants with type 2 diabetes, obesity and serum adiponectin levels in south Indian population. Gene 2013, 532, 253–262. [Google Scholar] [CrossRef]
- Rohde, K.; Keller, M.; Horstmann, A.; Liu, X.; Eichelmann, F.; Stumvoll, M.; Villringer, A.; Kovacs, P.; Tönjes, A.; Böttcher, Y. Role of genetic variants in ADIPOQ in human eating behaviour. Genes Nutr. 2015, 10, 449. [Google Scholar] [CrossRef]
- Phan, L.; Jin, Y.; Zhang, H.; Qiang, W.; Shekhtman, E.; Shao, D.; Revoe, D.; Villamarin, R.; Ivanchenko, E.; Kimura, M.; et al. ALFA: Allele Frequency Aggregator. 2020. Available online: www.ncbi.nlm.nih.gov/snp/docs/gsr/alfa/ (accessed on 10 January 2021).
- Ambroziak, M.; Kolanowska, M.; Bartoszewicz, Z.; Budaj, A. Adiponectin gene variants and decreased adiponectin plasma levels are associated with the risk of myocardial infarction in young age. Gene 2018, 642, 498–504. [Google Scholar] [CrossRef]
- Li, Z.; Peng, X.; Wu, Y.; Xia, Y.; Liu, X.; Zhang, Q. The influence of adiponectin gene polymorphism on the pioglitazone response in the Chinese with type 2 diabetes. Diabetes Obes. Metab. 2008, 10, 794–802. [Google Scholar] [CrossRef]
- Zadjali, F.; Al-Yahyaee, S.; Hassan, M.O.; Albarwani, S.; Bayoumi, R.A. Association of adiponectin promoter variants with traits and clusters of metabolic syndrome in Arabs: Family-based study. Gene 2013, 527, 663–669. [Google Scholar] [CrossRef]
- Hussain, M.K.; Deli, F.A.; Algenabi, A.H.A.; Abdul-Rudha, K.H. Adiponectin gene polymorphisms as a predictor for development of type 2 diabetes mellitus in Iraqi population. Gene 2018, 662, 118–122. [Google Scholar] [CrossRef]
- Palit, S.P.; Patel, R.; Jadeja, S.D.; Rathwa, N.; Mahajan, A.; Ramachandran, A.V.; Dhar, M.K.; Sharma, S.; Begum, R. A genetic analysis identifies a haplotype at adiponectin locus: Association with obesity and type 2 diabetes. Sci. Rep. 2020, 10, 2904. [Google Scholar] [CrossRef]
- Ye, Y.; Pu, D.; Liu, J.; Li, F.; Cui, Y.; Wu, J. Adiponectin gene polymorphisms may not be associated with idiopathic premature ovarian failure. Gene 2013, 518, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, S.; Chen, Y.; Liu, H.; Lan, C.; Chen, X.; Chi, S.; Chen, S.; Zhang, W. APM1 gene variants −11377C/G and 4545G/C are associated respectively with obesity and with non-obesity in Chinese type 2 diabetes. Diabetes Res. Clin. Pract. 2009, 84, 205–210. [Google Scholar] [CrossRef]
- Durmanova, V.; Parnicka, Z.; Javor, J.; Minarik, G.; Vrazda, L.; Vaseckova, B.; Gmitterova, K.; Kralova, M.; Pecenak, J.; Filipcik, P.; et al. A Novel Association of Polymorphism in the ITGA4 Gene Encoding the VLA-4 α4 Subunit with Increased Risk of Alzheimer’s Disease. Mediat. Inflamm. 2018, 2018, 7623823. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A web tool for the analysis of association studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef]
- Francischetti, E.A.; Dezonne, R.S.; Pereira, C.M.; de Moraes Martins, C.J.; Celoria, B.M.J.; de Oliveira, P.A.C.; de Abreu, V.G. Insights Into the Controversial Aspects of Adiponectin in Cardiometabolic Disorders. Horm. Metab. Res. 2020, 52, 695–707. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Boutin, P.; Mori, Y.; Tobe, K.; Dina, C.; Yasuda, K.; Yamauchi, T.; Otabe, S.; Okada, T.; Eto, K.; et al. Genetic variation in the gene encoding adiponectin is associated with an increased risk of type 2 diabetes in the Japanese population. Diabetes 2002, 51, 536–540. [Google Scholar] [CrossRef]
- Menzaghi, C.; Ercolino, T.; Salvemini, L.; Coco, A.; Kim, S.H.; Fini, G.; Doria, A.; Trischitta, V. Multigenic control of serum adiponectin levels: Evidence for a role of the APM1 gene and a locus on 14q13. Physiol. Genom. 2004, 19, 170–174. [Google Scholar] [CrossRef]
- Woo, J.G.; Dolan, L.M.; Deka, R.; Kaushal, R.D.; Shen, Y.; Pal, P.; Daniels, S.R.; Martin, L.J. Interactions between noncontiguous haplotypes in the adiponectin gene ACDC are associated with plasma adiponectin. Diabetes 2006, 55, 523–529. [Google Scholar] [CrossRef]
- Ong, K.L.; Li, M.; Tso, A.W.; Xu, A.; Cherny, S.S.; Sham, P.C.; Tse, H.F.; Lam, T.H.; Cheung, B.M.; Lam, K.S.; et al. Association of genetic variants in the adiponectin gene with adiponectin level and hypertension in Hong Kong Chinese. Eur. J. Endocrinol. 2010, 163, 251–257. [Google Scholar] [CrossRef]
- Vasseur, F.; Helbecque, N.; Lobbens, S.; Vasseur-Delannoy, V.; Dina, C.; Clément, K.; Boutin, P.; Kadowaki, T.; Scherer, P.E.; Froguel, P. Hypoadiponectinaemia and high risk of type 2 diabetes are associated with adiponectin-encoding (ACDC) gene promoter variants in morbid obesity: Evidence for a role of ACDC in diabesity. Diabetologia 2005, 48, 892–899. [Google Scholar] [CrossRef]
- Tankó, L.B.; Siddiq, A.; Lecoeur, C.; Larsen, P.J.; Christiansen, C.; Walley, A.; Froguel, P. ACDC/adiponectin and PPAR-gamma gene polymorphisms: Implications for features of obesity. Obes. Res. 2005, 13, 2113–2121. [Google Scholar] [CrossRef]
- Guo, X.; Saad, M.F.; Langefeld, C.D.; Williams, A.H.; Cui, J.; Taylor, K.D.; Norris, J.M.; Jinagouda, S.; Darwin, C.H.; Mitchell, B.D.; et al. Genome-wide linkage of plasma adiponectin reveals a major locus on chromosome 3q distinct from the adiponectin structural gene: The IRAS family study. Diabetes 2006, 55, 1723–1730. [Google Scholar] [CrossRef]
- Schwarz, P.E.; Towers, G.W.; Fischer, S.; Govindarajalu, S.; Schulze, J.; Bornstein, S.R.; Hanefeld, M.; Vasseur, F. Hypoadiponectinemia is associated with progression toward type 2 diabetes and genetic variation in the ADIPOQ gene promoter. Diabetes Care 2006, 29, 1645–1650. [Google Scholar] [CrossRef]
- Bouatia-Naji, N.; Meyre, D.; Lobbens, S.; Séron, K.; Fumeron, F.; Balkau, B.; Heude, B.; Jouret, B.; Scherer, P.E.; Dina, C.; et al. ACDC/adiponectin polymorphisms are associated with severe childhood and adult obesity. Diabetes 2006, 55, 545–550. [Google Scholar] [CrossRef]
- Petrone, A.; Zavarella, S.; Caiazzo, A.; Leto, G.; Spoletini, M.; Potenziani, S.; Osborn, J.; Vania, A.; Buzzetti, R. The promoter region of the adiponectin gene is a determinant in modulating insulin sensitivity in childhood obesity. Obesity 2006, 14, 1498–1504. [Google Scholar] [CrossRef]
- Jang, Y.; Chae, J.S.; Koh, S.J.; Hyun, Y.J.; Kim, J.Y.; Jeong, Y.J.; Park, S.; Ahn, C.M.; Lee, J.H. The influence of the adiponectin gene on adiponectin concentrations and parameters of metabolic syndrome in non-diabetic Korean women. Clin. Chim. Acta 2008, 391, 85–90. [Google Scholar] [CrossRef]
- Tong, G.; Wang, N.; Leng, J.; Tong, X.; Shen, Y.; Yang, J.; Ye, X.; Zhou, L.; Zhou, Y. Common variants in adiponectin gene are associated with coronary artery disease and angiographical severity of coronary atherosclerosis in type 2 diabetes. Cardiovasc. Diabetol. 2013, 12, 67. [Google Scholar] [CrossRef]
- Smetnev, S.; Klimushina, M.; Kutsenko, V.; Kiseleva, A.; Gumanova, N.; Kots, A.; Skirko, O.; Ershova, A.; Yarovaya, E.; Metelskaya, V.; et al. Associations of SNPs of the ADIPOQ Gene with Serum Adiponectin Levels, Unstable Angina, and Coronary Artery Disease. Biomolecules 2019, 9, 537. [Google Scholar] [CrossRef]
- Cox, A.J.; Lambird, J.E.; An, S.S.; Register, T.C.; Langefeld, C.D.; Carr, J.J.; Freedman, B.I.; Bowden, D.W. Variants in adiponectin signaling pathway genes show little association with subclinical CVD in the diabetes heart study. Obesity 2013, 21, E456–E462. [Google Scholar] [CrossRef]
- Pollin, T.I.; Tanner, K.; O’connell, J.R.; Ott, S.H.; Damcott, C.M.; Shuldiner, A.R.; McLenithan, J.C.; Mitchell, B.D. Linkage of plasma adiponectin levels to 3q27 explained by association with variation in the APM1 gene. Diabetes 2005, 54, 268–274. [Google Scholar] [CrossRef]
- González-Sánchez, J.L.; Zabena, C.A.; Martínez-Larrad, M.T.; Fernández-Pérez, C.; Pérez-Barba, M.; Laakso, M.; Serrano-Ríos, M. An SNP in the adiponectin gene is associated with decreased serum adiponectin levels and risk for impaired glucose tolerance. Obes. Res. 2005, 13, 807–812. [Google Scholar] [CrossRef]
- Mackevics, V.; Heid, I.M.; Wagner, S.A.; Cip, P.; Doppelmayr, H.; Lejnieks, A.; Gohlke, H.; Ladurner, G.; Illig, T.; Iglseder, B.; et al. The adiponectin gene is associated with adiponectin levels but not with characteristics of the insulin resistance syndrome in healthy Caucasians. Eur. J. Hum. Genet. 2006, 14, 349–356. [Google Scholar] [CrossRef]
- Zhang, D.; Ma, J.; Brismar, K.; Efendic, S.; Gu, H.F. A single nucleotide polymorphism alters the sequence of SP1 binding site in the adiponectin promoter region and is associated with diabetic nephropathy among type 1 diabetic patients in the Genetics of Kidneys in Diabetes Study. J. Diabetes Complicat. 2009, 23, 265–272. [Google Scholar] [CrossRef]
- Lee, B.P.; Lloyd-Laney, H.O.; Locke, J.M.; McCulloch, L.J.; Knight, B.; Yaghootkar, H.; Cory, G.; Kos, K.; Frayling, T.M.; Harries, L.W. Functional characterisation of ADIPOQ variants using individuals recruited by genotype. Mol. Cell. Endocrinol. 2016, 428, 49–57. [Google Scholar] [CrossRef]
- Jin, T.; Huang, W.; Cao, F.; Yu, X.; Ying, Z.; Guo, S.; Cheng, Y.; Xu, C. Causal association between adiponectin and the risk of Alzheimer’s disease: A Mendelian randomization study. Front. Neurol. 2022, 13, 1038975. [Google Scholar] [CrossRef]
- Horgusluoglu, E.; Neff, R.; Song, W.M.; Wang, M.; Wang, Q.; Arnold, M.; Krumsiek, J.; Galindo-Prieto, B.; Ming, C.; Nho, K.; et al. Integrative metabolomics-genomics approach reveals key metabolic pathways and regulators of Alzheimer’s disease. Alzheimers Dement. 2022, 18, 1260–1278. [Google Scholar] [CrossRef]
- Narayana Swamy, A.; Valasala, H.; Kamma, S. In silico Evaluation of Nonsynonymous Single Nucleotide Polymorphisms in the ADIPOQ Gene Associated with Diabetes, Obesity, and Inflammation. Avicenna J. Med. Biotechnol. 2015, 7, 121–127. [Google Scholar]
- Takahashi, M.; Arita, Y.; Yamagata, K.; Matsukawa, Y.; Okutomi, K.; Horie, M.; Shimomura, I.; Hotta, K.; Kuriyama, H.; Kihara, S.; et al. Genomic structure and mutations in adipose-specific gene, adiponectin. Int. J. Obes. Relat. Metab. Disord. 2000, 24, 861–868. [Google Scholar] [CrossRef]
- Waki, H.; Yamauchi, T.; Kamon, J.; Ito, Y.; Uchida, S.; Kita, S.; Hara, K.; Hada, Y.; Vasseur, F.; Froguel, P.; et al. Impaired multimerization of human adiponectin mutants associated with diabetes. Molecular structure and multimer formation of adiponectin. J. Biol. Chem. 2003, 278, 40352–40363. [Google Scholar] [CrossRef]
- Iwashima, Y.; Katsuya, T.; Ishikawa, K.; Ouchi, N.; Ohishi, M.; Sugimoto, K.; Fu, Y.; Motone, M.; Yamamoto, K.; Matsuo, A.; et al. Hypoadiponectinemia is an independent risk factor for hypertension. Hypertension 2004, 43, 1318–1323. [Google Scholar] [CrossRef]
- Jungtrakoon, P.; Plengvidhya, N.; Tangjittipokin, W.; Chimnaronk, S.; Salaemae, W.; Chongjaroen, N.; Chanprasert, K.; Sujjitjoon, J.; Srisawat, C.; Yenchitsomanus, P.T. Novel adiponectin variants identified in type 2 diabetic patients reveal multimerization and secretion defects. PLoS ONE 2011, 6, e26792. [Google Scholar] [CrossRef]
- Paik, H.; Lee, J.; Jeong, C.S.; Park, J.S.; Lee, J.H.; Rappoport, N.; Kim, Y.; Sohn, H.Y.; Jo, C.; Kim, J.; et al. Identification of a pleiotropic effect of ADIPOQ on cardiac dysfunction and Alzheimer’s disease based on genetic evidence and health care records. Transl. Psychiatry 2022, 12, 389. [Google Scholar] [CrossRef]


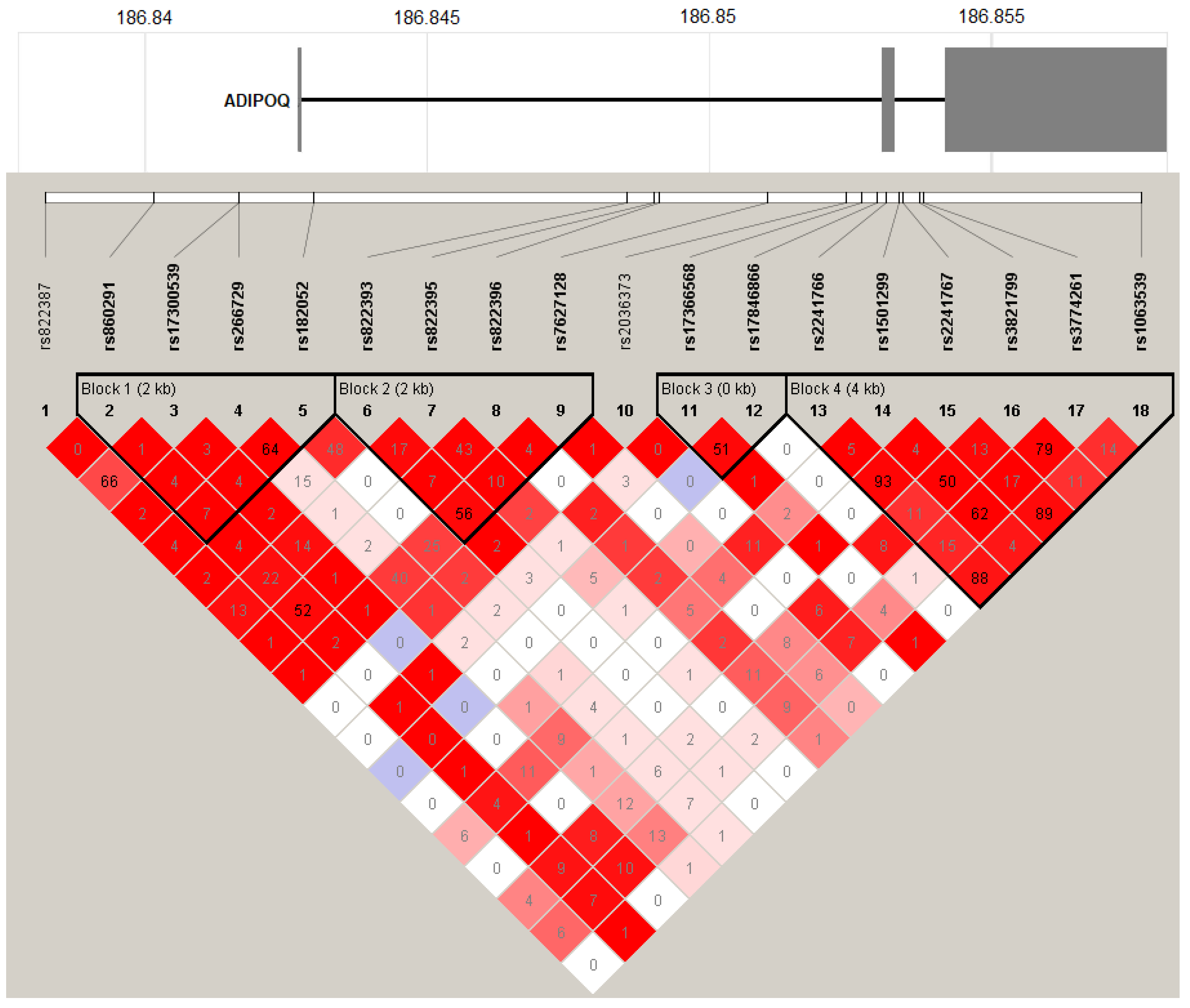{kind=link}
| Parameter | LOAD (n = 385) | Controls (n = 533) | p-Value |
|---|---|---|---|
| Age at examination (years) | 77.73 ± 6.36 | 76.35 ± 7.68 | 0.0031 |
| Age of onset (years) | 75.06 ± 6.39 | - | - |
| Sex (female/male) | 243 (63.12%)/142 (36.88%) | 320 (60.04%)/213 (39.96%) | 0.34 |
| MoCA score | 16.15 ± 5.82 | 27.53 ± 1.40 | <0.0001 |
| BMI (kg/m2) | 26.57 ± 4.53 | 27.77 ± 3.99 | <0.0001 |
| Hypertension (yes/no) | 277 (71.95%)/108 (28.05%) | 355 (66.60%)/178 (33.40%) | 0.085 |
| T2DM (yes/no) | 93 (24.16%)/292 (75.84%) | 72 (13.51%)/461 (86.49%) | <0.0001 |
| APOE ε4 positivity (yes/no) | 195 (50.65%)/190 (49.35%) | 101 (18.95%)/432 (81.05%) | <0.0001 |
| dbSNP | Alleles a | Position b | RefSeqGene | Gene Region | Aliases |
|---|---|---|---|---|---|
| rs822387 | T>C | chr3:186838248 | NG_021140.1:g.575T>C | 5′-flanking region | −14,811 T/C |
| rs860291 | C>T | chr3:186840168 | NG_021140.1:g.2495T>C | 5′-flanking region | −12,823 C/T; −12,891 C/T |
| rs17300539 | G>A | chr3:186841671 | NG_021140.1:g.3998G>A | Promoter | −11,388G/A; −11,391 G/A |
| rs266729 | C>G | chr3:186841685 | NG_021140.1:g.4012C>G | Promoter | −11,377 C/G; −11,365 C/G |
| rs182052 | G>A | chr3:186842993 | NG_021140.1:g.5320G>A | Intron 1 | −10,066 G/A; −10,068 G/A |
| rs822393 | C>T | chr3:186848537 | NG_021140.1:g.10864C>T | Intron 1 | −4522 C/T |
| rs822395 | A>C | chr3:186849018 | NG_021140.1:g.11345C>A | Intron 1 | −4034 A/C; −4041 A/C |
| rs822396 | A>G | chr3:186849088 | NG_021140.1:g.11415G>A | Intron 1 | −3964 A/G; −3971 A/G |
| rs7627128 | C>A | chr3:186851010 | NG_021140.1:g.13337C>A | Intron 1 | −2049 C/A |
| rs2036373 | T>G | chr3:186852402 | NG_021140.1:g.14729T>G | Intron 1 | −657 T/G |
| rs17366568 | G>A | chr3:186852664 | NG_021140.1:g.14991G>A | Intron 1 | −395 G/A |
| rs17846866 | T>G | chr3:186852957 | NG_021140.1:g.15284T>G | Intron 1 | −102 T/G |
| rs2241766 | T>G | chr3:186853103 | NG_021140.1:g.15430T>G | Exon 2 | +45 T/G; Gly15Gly; T94G |
| rs1501299 | G>T | chr3:186853334 | NG_021140.1:g.15661G>T | Intron 2 | +276 G/T |
| rs2241767 | A>G | chr3:186853407 | NG_021140.1:g.15734A>G | Intron 2 | +349 A/G |
| rs3821799 | C>T | chr3:186853697 | NG_021140.1:g.16024T>C | Intron 2 | +639 C/T |
| rs3774261 | G>A | chr3:186853770 | NG_021140.1:g.16097A>G | Intron 2 | +712 G/A |
| rs1063539 | G>C | chr3:186857603 | NG_021140.1:g.19930G>C | Exon 3/3′ UTR | +4545 G/C |
| ADIPOQ SNPs | LOAD (n = 385) | Controls (n = 533) | Logistic Regression Analysis (Log-Additive Model) | |||
|---|---|---|---|---|---|---|
| Genotypes a | MAF | Genotypes a | MAF | p-Value | OR (95% CI) | |
| rs822387 T>C | 336/46/3 | 0.0675 | 463/65/5 | 0.0704 | 0.64 | 0.91 (0.62–1.34) |
| rs860291 C>T | 301/80/4 | 0.1143 | 425/105/3 | 0.1041 | 0.54 | 1.11 (0.80–1.53) |
| rs17300539 G>A | 335/49/1 | 0.0662 | 457/70/6 | 0.0769 | 0.32 | 0.82 (0.56–1.21) |
| rs266729 C>G | 206/147/32 | 0.2740 | 279/211/43 | 0.2786 | 0.66 | 0.95 (0.76–1.18) |
| rs182052 G>A | 151/175/59 | 0.3805 | 212/252/69 | 0.3659 | 0.72 | 1.04 (0.85–1.27) |
| rs822393 C>T | 213/139/33 | 0.2662 | 296/205/32 | 0.2523 | 0.36 | 1.11 (0.89–1.39) |
| rs822395 A>C | 170/167/48 | 0.3416 | 240/229/64 | 0.3349 | 0.86 | 0.98 (0.80–1.21) |
| rs822396 A>G | 256/115/14 | 0.1857 | 358/159/16 | 0.1792 | 0.80 | 0.97 (0.75–1.25) |
| rs7627128 C>A | 269/106/10 | 0.1636 | 363/153/17 | 0.1754 | 0.59 | 0.93 (0.71–1.21) |
| rs2036373 T>G | 343/42/0 | 0.0545 | 475/57/1 | 0.0553 | 0.62 | 1.12 (0.72–1.75) |
| rs17366568 G>A | 302/79/4 | 0.1130 | 401/127/5 | 0.1285 | 0.47 | 0.89 (0.65–1.22) |
| rs17846866 T>G | 329/56/0 | 0.0727 | 464/67/2 | 0.0666 | 0.48 | 1.15 (0.77–1.72) |
| rs2241766 T>G | 304/77/4 | 0.1104 | 427/100/6 | 0.1051 | 0.66 | 1.08 (0.78–1.48) |
| rs1501299 G>T | 184/167/34 | 0.3052 | 270/215/48 | 0.2917 | 0.24 | 1.14 (0.92–1.41) |
| rs2241767 A>G | 306/77/2 | 0.1052 | 430/97/6 | 0.1022 | 0.79 | 1.05 (0.75–1.46) |
| rs3821799 C>T | 112/191/82 | 0.4610 | 168/251/114 | 0.4493 | 0.28 | 1.12 (0.92–1.36) |
| rs3774261 G>A | 134/186/65 | 0.4104 | 202/242/89 | 0.3940 | 0.21 | 1.14 (0.93–1.39) |
| rs1063539 G>C | 302/80/3 | 0.1117 | 423/104/6 | 0.1088 | 0.84 | 1.03 (0.75–1.43) |
| ADIPOQ Haplotypes | LOAD (n = 385) | Controls (n = 533) | Logistic Regression Analysis | |
|---|---|---|---|---|
| EHF | EHF | p-Value | OR (95% CI) | |
| Block 1 (rs860291–rs17300539–rs266729–rs182052) | ||||
| CGCG | 0.4390 | 0.4531 | Reference | |
| CGGA | 0.2740 | 0.2786 | 0.87 | 0.98 (0.77–1.24) |
| TGCG | 0.1143 | 0.1041 | 0.55 | 1.11 (0.79–1.56) |
| CGCA | 0.1065 | 0.0872 | 0.25 | 1.23 (0.87–1.74) |
| CACG | 0.0662 | 0.0769 | 0.43 | 0.85 (0.57–1.27) |
| Block 2 (rs822393–rs822395–rs822396–rs7627128) | ||||
| CAAC | 0.3926 | 0.4116 | Reference | |
| CCGC | 0.1857 | 0.1781 | 0.87 | 1.02 (0.77–1.36) |
| TAAA | 0.1622 | 0.1713 | 0.95 | 0.99 (0.74–1.32) |
| CCAC | 0.1540 | 0.1549 | 0.82 | 1.04 (0.77–1.40) |
| TAAC | 0.1022 | 0.0781 | 0.062 | 1.42 (0.98–2.05) |
| Block 3 (rs17366568–rs17846866) | ||||
| GT | 0.8857 | 0.8705 | Reference | |
| AG | 0.0714 | 0.0656 | 0.57 | 1.12 (0.75–1.68) |
| AT | 0.0416 | 0.0629 | 0.083 | 0.66 (0.42–1.05) |
| Block 4 (rs2241766–rs1501299–rs2241767–rs3821799–rs3774261–rs1063539) | ||||
| TGACGG | 0.5237 | 0.5413 | Reference | |
| TTATAG | 0.2997 | 0.2917 | 0.24 | 1.15 (0.91–1.44) |
| GGGTAC | 0.0993 | 0.1004 | 0.69 | 1.07 (0.76–1.51) |
| TGATGG | 0.0537 | 0.0553 | 0.75 | 1.08 (0.68–1.69) |
| ADIPOQ SNPs | Adiponectin Levels (μg/mL) a | Genetic Model | padj-Value b | Δ (95% CI) b | ||
|---|---|---|---|---|---|---|
| 1/1 | 1/2 | 2/2 | ||||
| rs822387 T>C | 10.14 ± 5.36 (136) | 11.95 ± 5.86 (18) | 13.39 ± 0.46 (2) | A | 0.14 | 1.61 (−0.50–3.72) |
| rs860291 C>T | 10.18 ± 5.11 (126) | 11.23 ± 6.68 (29) | 12.41 ± 0.00 (1) | A | 0.50 | 0.67 (−1.29–2.64) |
| rs17300539 G>A | 10.19 ± 5.39 (138) | 11.89 ± 5.65 (17) | 13.07 ± 0.00 (1) | A | 0.20 | 1.55 (−0.80–3.89) |
| rs266729 C>G | 11.05 ± 5.72 (80) | 10.04 ± 5.16 (62) | 8.18 ± 4.08 (14) | R | 0.12 | −2.26 (−5.09–0.56) |
| rs182052 G>A | 11.66 ± 5.76 (55) | 9.79 ± 5.48 (73) | 9.46 ± 4.05 (28) | D | 0.075 | −1.56 (−3.26–0.14) |
| rs822393 C>T | 11.09 ± 5.64 (81) | 9.72 ± 5.44 (57) | 9.33 ± 3.86 (18) | D | 0.10 | −1.36 (−2.97–0.26) |
| rs822395 A>C | 9.68 ± 4.95 (75) | 10.89 ± 5.68 (64) | 11.65 ± 6.16 (17) | D | 0.048 | 1.64 (0.03–3.25) |
| rs822396 A>G | 10.30 ± 5.13 (104) | 10.55 ± 6.15 (47) | 10.90 ± 4.71 (5) | D | 0.83 | 0.19 (−1.54–1.91) |
| rs7627128 C>A | 10.77 ± 5.56 (107) | 9.66 ± 5.25 (44) | 8.72 ± 2.12 (5) | A | 0.33 | −0.76 (−2.27–0.75) |
| rs2036373 T>G | 10.07 ± 5.17 (140) | 13.18 ± 6.76 (16) | - | - | 0.043 | 2.80 (0.12–5.49) |
| rs17366568 G>A | 10.27 ± 5.19 (125) | 10.97 ± 6.45 (28) | 10.06 ± 5.51 (3) | D | 0.20 | 1.36 (−0.70–3.43) |
| rs17846866 T>G | 10.28 ± 5.20 (135) | 11.08 ± 6.70 (21) | - | - | 0.18 | 1.65 (−0.76–4.07) |
| rs2241766 T>G | 10.56 ± 5.25 (127) | 9.24 ± 5.79 (28) | 21.38 ± 0.00 (1) | O | 0.18 | −1.45 (−3.58–0.67) |
| rs1501299 G>T | 10.41 ± 5.06 (73) | 10.30 ± 5.82 (69) | 10.72 ± 5.43 (14) | D | 0.99 | −0.01 (−1.66–1.65) |
| rs2241767 A>G | 10.54 ± 5.29 (125) | 9.80 ± 5.94 (31) | - | - | 0.35 | −0.98 (−3.03–1.07) |
| rs3821799 C>T | 10.42 ± 4.63 (47) | 10.50 ± 5.86 (76) | 10.10 ± 5.52 (33) | R | 0.60 | −0.54 (−2.54–1.46) |
| rs3774261 G>A | 10.26 ± 4.47 (54) | 10.78 ± 6.11 (76) | 9.52 ± 5.08 (26) | R | 0.22 | −1.38 (−3.56–0.80) |
| rs1063539 G>C | 10.54 ± 5.29 (125) | 9.41 ± 5.63 (30) | 21.38 ± 0.00 (1) | O | 0.20 | −1.36 (−3.43–0.71) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javor, J.; Ďurmanová, V.; Klučková, K.; Párnická, Z.; Radošinská, D.; Šutovský, S.; Vašečková, B.; Režnáková, V.; Králová, M.; Gmitterová, K.; et al. Adiponectin Gene Polymorphisms: A Case–Control Study on Their Role in Late-Onset Alzheimer’s Disease Risk. Life 2024, 14, 346. https://doi.org/10.3390/life14030346
Javor J, Ďurmanová V, Klučková K, Párnická Z, Radošinská D, Šutovský S, Vašečková B, Režnáková V, Králová M, Gmitterová K, et al. Adiponectin Gene Polymorphisms: A Case–Control Study on Their Role in Late-Onset Alzheimer’s Disease Risk. Life. 2024; 14(3):346. https://doi.org/10.3390/life14030346
Chicago/Turabian StyleJavor, Juraj, Vladimíra Ďurmanová, Kristína Klučková, Zuzana Párnická, Dominika Radošinská, Stanislav Šutovský, Barbora Vašečková, Veronika Režnáková, Mária Králová, Karin Gmitterová, and et al. 2024. "Adiponectin Gene Polymorphisms: A Case–Control Study on Their Role in Late-Onset Alzheimer’s Disease Risk" Life 14, no. 3: 346. https://doi.org/10.3390/life14030346