Effects of Dietary l-Glutamine Supplementation on the Intestinal Function and Muscle Growth of Piglets

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Experimental Design
2.2. Sample Collection
2.3. Intestinal Morphology Measurement
2.4. Determination of DAO Activity in Plasma
2.5. Assay of DNA, RNA, and Protein Contents in Intestinal Mucosa and Longissimus Dorsi Muscle
2.6. Determination of Amino Acid Contents in Intestinal Mucosa and Longissimus Dorsi Muscle
2.7. Quantitative Real-Time PCR Analyses for Gene Expression
2.8. Quantitative PCR Analyses for Colon Bacteria
2.9. Protein Immunoblot Analysis
2.10. Determination of the Antioxidant Capacity of Longissimus Dorsi Muscle
2.11. Statistical Analysis
3. Results
3.1. Determination the Activity of DAO in Plasma and Intestinal Morphology
3.2. The Contents of TP, RNA, and DNA in the Intestinal Mucosa of Piglets
3.3. Concentrations of Amino Acids in the Jejunum and Ileum
3.4. Intestinal Gene Expression and Colon Bacterial Abundance
3.5. Concentrations of Amino Acids in the Longissimus Dorsi Muscle
3.6. TP, RNA, and DNA in the Longissimus Dorsi Muscle and Muscle Gene Expression
3.7. Redox Status of Longissimus Dorsi Muscle
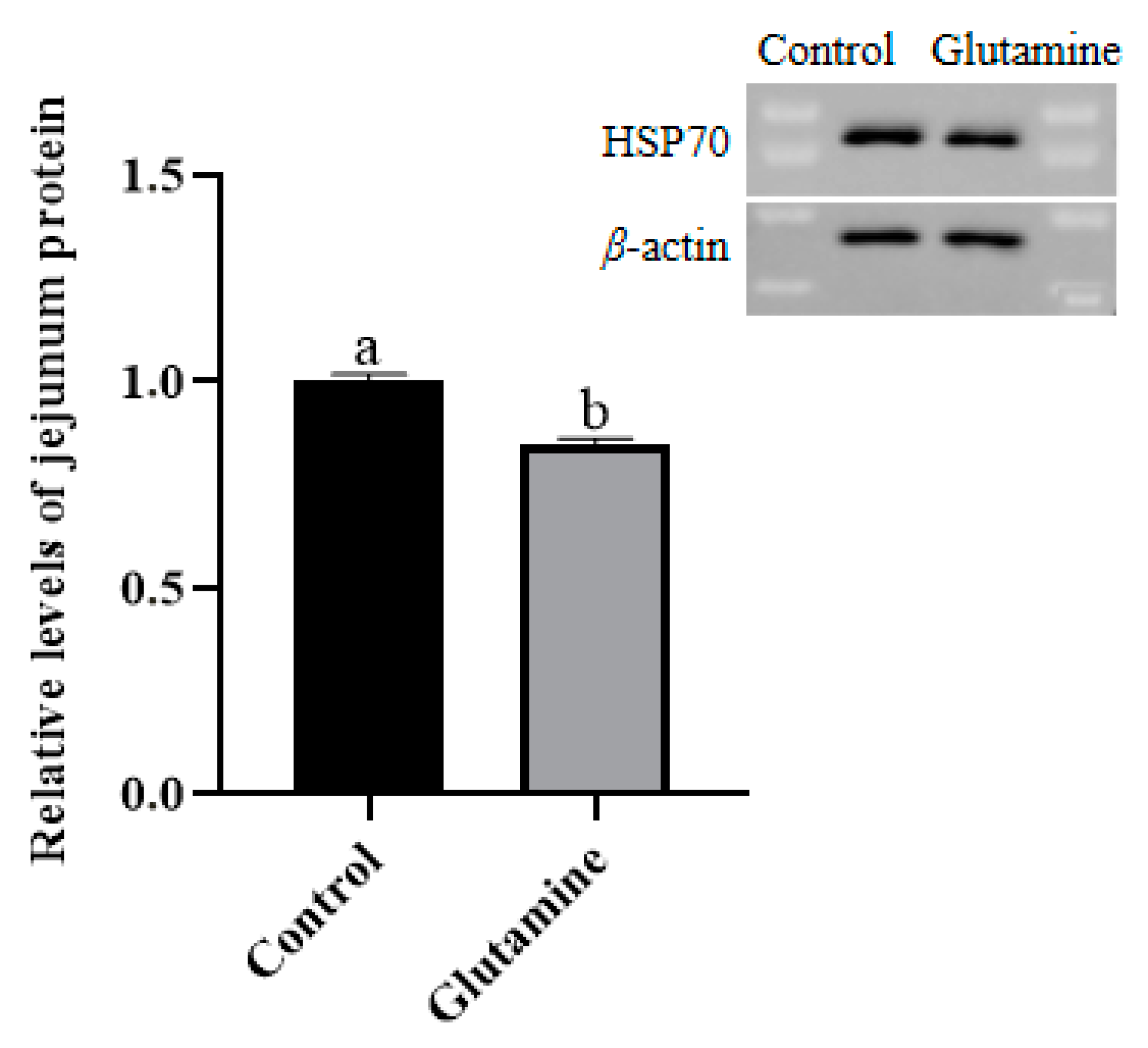
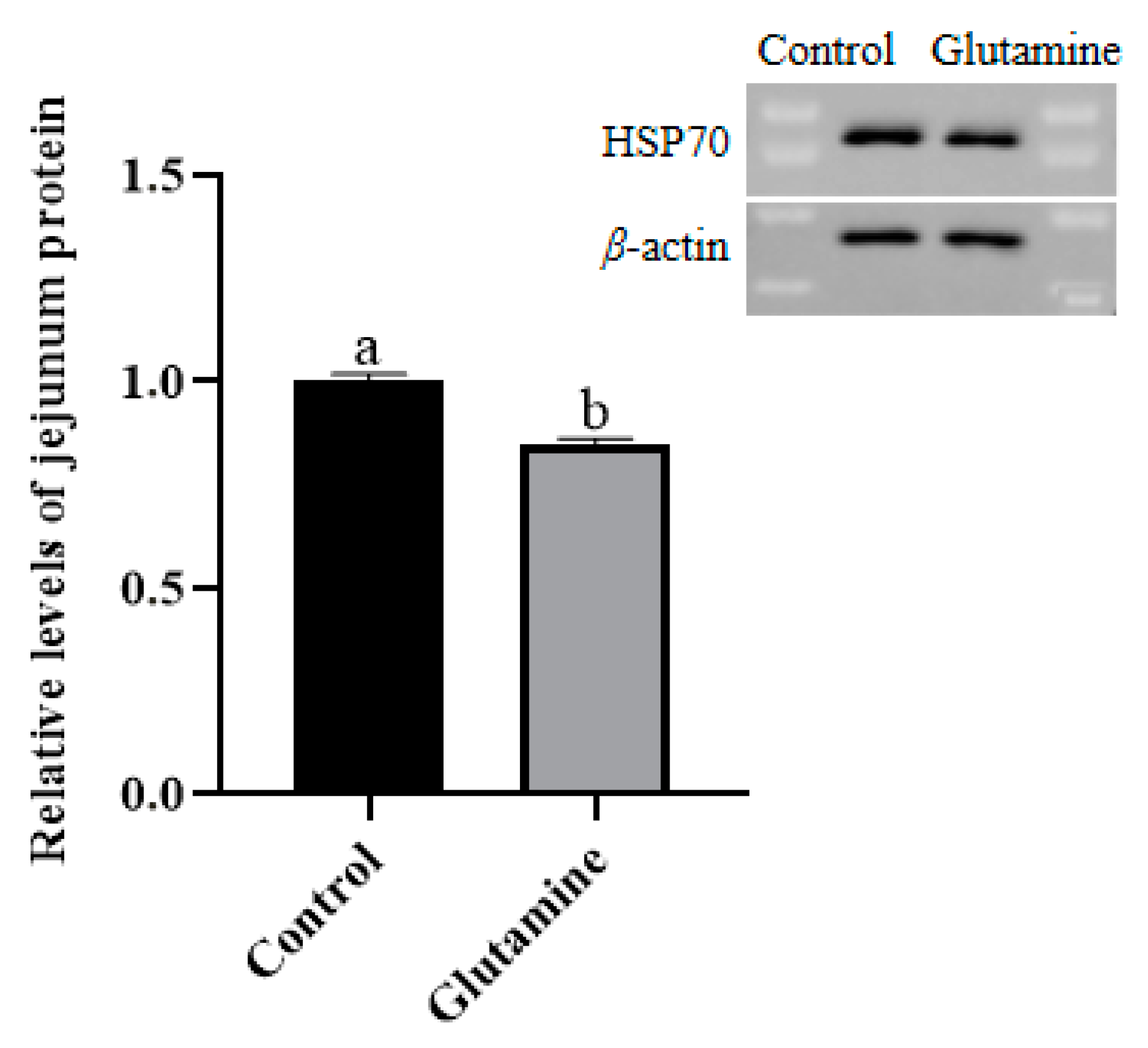
3.8. Effect of Gln Supplementation on the Abundance of the HSP70 Protein in the Jejunum Mucosa of Piglets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASCT2 | Sodium-dependent neutral amino acid transporter 2 |
| CAT | Catalase |
| DAO | Diamine oxidase |
| Gln | l-glutamine |
| GSTO2 | Glutathione-S-transferase omega-2 |
| H2O2 | Hydrogen peroxide |
| HSP70 | Heat shock protein-70 |
| IFIT | Interferon induced protein with tetratricopeptide repeats |
| ISGs | Interferon stimulated genes |
| MDA | Malondialdehyde |
| MSTN | Myostatin |
| mTOR | Mammalian target of rapamycin |
| MX1 | MX dynamin Like GTPase 1 |
| MX2 | MX dynamin like GTPase 2 |
| Myf5 | Myogenic Factor 5 |
| OAS1 | 2′-5′-Oligoadenylate Synthetase 1 |
| OASL | 2′-5′-Oligoadenylate Synthetase Like |
| PepT1 | Peptide transporter 1 |
| SLC7A6 | l-type amino acid transporter 2 |
| SLC7A7 | l-type amino acid transporter 1 |
| SLC6A19 | Sodium-independent amino acid transporter |
| SOD | Superoxide dismutase |
| TP | Total protein |
| VH | Villus height |
| VH/CD | Villus height/crypt depth ratio |
| VS | Villus surface area |
| VW | Villus width |
References
- Ji, F.J.; Wang, L.X.; Yang, H.S.; Hu, A.; Yin, Y.L. Review: The roles and functions of glutamine on intestinal health and performance of weaning pigs. Animal 2019, 13, 2727–2735. [Google Scholar] [CrossRef]
- Wu, G.; Morris, S.M., Jr. Arginine metabolism: Nitric oxide and beyond. Biochem. J. 1998, 336 Pt 1, 1–17. [Google Scholar] [CrossRef]
- Marc Rhoads, J.; Wu, G. Glutamine, arginine, and leucine signaling in the intestine. Amino Acids 2009, 37, 111–122. [Google Scholar] [CrossRef]
- Wu, G.; Fang, Y.Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione metabolism and its implications for health. J. Nutr. 2004, 1134, 489–492. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Dai, Z.; Li, D.; Wang, J.; Wu, Z. Amino acid nutrition in animals: Protein synthesis and beyond. Annu. Rev. Anim. Biosci. 2014, 2, 387–417. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.K.; Duan, J.L.; Yin, J.; Liu, G.; Cao, Z.; Xiong, X.; Chen, S.; Li, T.; Yin, Y.; Hou, Y.Q.; et al. Dietary L-glutamine supplementation modulates microbial community and activates innate immunity in the mouse intestine. Amino Acids 2014, 46, 2403–2413. [Google Scholar] [CrossRef]
- Ren, W.K.; Yin, J.; Wu, M.; Liu, G.; Yang, G.; Xion, Y.; Su, D.; Wu, L.; Li, T.; Chen, S.; et al. Serum amino acids profile and the beneficial effects of L-arginine or L-glutamine supplementation in dextran sulfate sodium colitis. PLoS ONE 2014, 9, e88335. [Google Scholar] [CrossRef]
- Yi, D.; Hou, Y.; Wang, L.; Ouyang, W.; Long, M.; Zhao, D.; Ding, B.; Liu, Y.; Wu, G. L-Glutamine enhances enterocyte growth via activation of the mTOR signaling pathway independently of AMPK. Amino Acids 2015, 47, 65–78. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.L.; Zhang, J.; Wu, G.; Zhu, W.Y. Utilization of amino acids by bacteria from the pig small intestine. Amino Acids 2010, 39, 1201–1215. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.L.; Li, X.L.; Xi, P.B.; Zhang, J.; Wu, G.; Zhu, W.Y. Metabolism of select amino acids in bacteria from the pig small intestine. Amino Acids 2012, 42, 1597–1608. [Google Scholar] [CrossRef]
- Wu, G.; Meier, S.A.; Knabe, D.A. Dietary glutamine supplementation prevents jejunal atrophy in weaned pigs. J. Nutr. 1996, 126, 2578–2584. [Google Scholar] [CrossRef]
- Wang, J.; Chen, L.; Li, P.; Li, X.; Zhou, H.; Wang, F.; Li, D.; Yin, Y.; Wu, G. Gene expression is altered in piglet small intestine by weaning and dietary glutamine supplementation. J. Nutr. 2008, 138, 1025–1032. [Google Scholar] [CrossRef]
- Chamorro, S.; de Blas, C.; Grant, G.; Badiola, I.; Menoyo, D.; Carabaño, R. Effect of dietary supplementation with glutamine and a combination of glutamine-arginine on intestinal health in twenty-five-day-old weaned rabbits. J. Anim. Sci. 2010, 88, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Sawyer, M.B.; Field, C.J.; Dieleman, L.A.; Murray, D.; Baracos, V.E. Bolus oral glutamine protects rats against CPT-11-induced diarrhea and differentially activates cytoprotective mechanisms in host intestine but not tumor. J. Nutr. 2008, 138, 740–746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lu, T.; Han, L.; Zhao, L.; Niu, Y.; Chen, H. L-Glutamine supplementation alleviates constipation during late gestation of mini sows by modifying the microbiota composition in feces. Biomed Res. Int. 2017, 2017, 4862861. [Google Scholar] [CrossRef] [PubMed]
- Yi, D.; Li, B.; Hou, Y.; Wang, L.; Zhao, D.; Chen, H.; Wu, T.; Zhou, Y.; Ding, B.; Wu, G. Dietary supplementation with an amino acid blend enhances intestinal function in piglets. Amino Acids 2018, 50, 1089–1100. [Google Scholar] [CrossRef]
- Whiting, T.L.; Steele, G.G.; Wamnes, S.; Green, C. Evaluation of methods of rapid mass killing of segregated early weaned piglets. Can. Vet. J. 2011, 52, 753–758. [Google Scholar] [PubMed]
- Liu, P.; Piao, X.S.; Kim, S.W.; Wang, L.; Shen, Y.B.; Lee, H.S.; Li, S.Y. Effects of chito-oligosaccharide supplementation on the growth performance, nutrient digestibility, intestinal morphology, and fecal shedding of Escherichia coli and Lactobacillus in weaning pigs. J. Anim. Sci. 2008, 86, 2609–2618. [Google Scholar] [CrossRef]
- Hampson, D.J. Alterations in piglet small intestinal structure at weaning. Res. Vet. Sci. 1986, 40, 32–40. [Google Scholar] [CrossRef]
- Hou, Y.; Yao, K.; Wang, L.; Ding, B.; Fu, D.; Liu, Y.; Zhu, H.; Liu, J.; Li, Y.; Kang, P.; et al. Effects of α-ketoglutarate on energy status in the intestinal mucosa of weaned piglets chronically challenged with lipopolysaccharide. Br. J. Nutr. 2011, 106, 357–363. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, L.; Zhang, W.; Yang, Z.; Ding, B.; Zhu, H.; Liu, Y.; Qiu, Y.; Yin, Y.; Wu, G. Protective effects of N-acetylcysteine on intestinal functions of piglets challenged with lipopolysaccharide. Amino Acids 2012, 43, 1233–1242. [Google Scholar] [CrossRef]
- Johnson, L.R.; Chandler, A.M. RNA and DNA of gastric and duodenal mucosa in antrectomized and gastrin-treated rats. Am. J. Physiol. 1973, 224, 937–940. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Xie, C.; Zhang, S.; Zhang, G.; Zhang, F.; Chu, L.; Qiao, S. Estimation of the optimal ratio of standardized ileal digestible threonine to lysine for finishing barrows fed low crude protein diets. Asian-Australas. J. Anim. Sci. 2013, 59, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, L.; Yi, D.; Ding, B.; Yang, Z.; Li, J.; Chen, X.; Qiu, Y.; Wu, G. N-acetylcysteine reduces inflammation in the small intestine by regulating redox, EGF and TLR4 signaling. Amino Acids 2013, 45, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Yi, D.; Hou, Y.; Wang, L.; Long, M.; Hu, S.; Mei, H.; Yan, L.; Hu, C.A.; Wu, G. N-acetylcysteine stimulates protein synthesis in enterocytes independently of glutathione synthesis. Amino Acids 2016, 48, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Ott, S.J.; Musfeldt, M.; Ullmann, U.; Hampe, J.; Schreiber, S. Quantification of intestinal bacterial populations by real-time PCR with a universal primer set and minor groove binder probes: A global approach to the enteric flora. J. Clin. Microbiol. 2004, 42, 2566–2572. [Google Scholar] [CrossRef] [PubMed]
- Meurens, F.; Berri, M.; Auray, G.; Melo, S.; Levast, B.; Virlogeux-Payant, I.; Chevaleyre, C.; Gerdts, V.; Salmon, H. Early immune response following Salmonella enterica subspecies enterica serovar Typhimurium infection in porcine jejunal gut loops. Vet. Res. 2009, 40, 5. [Google Scholar] [CrossRef]
- Nygard, A.B.; Jørgensen, C.B.; Cirera, S.; Fredholm, M. Selection of reference genes for gene expression studies in pig tissues using SYBR green qPCR. BMC. Mol. Bio. 2007, 8, 67. [Google Scholar] [CrossRef]
- Tako, E.; Glahn, R.P.; Welch, R.M.; Lei, X.; Yasuda, K.; Miller, D.D. Dietary inulin affects the expression of intestinal enterocyte iron transporters, receptors and storage protein and alters the microbiota in the pig intestine. Br. J. Nutr. 2008, 99, 472–480. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Methods. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Castillo, M.; Martín-Orúe, S.M.; Manzanilla, E.G.; Badiola, I.; Martín, M.; Gasa, J. Quantification of total bacteria, enterobacteria and lactobacilli populations in pig digesta by real-time PCR. Vet. Microbiol. 2006, 114, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, L.; Ding, B.; Liu, Y.; Zhu, H.; Liu, J.; Li, Y.; Wu, X.; Yin, Y.; Wu, G. Dietary alpha-ketoglutarate supplementation ameliorates intestinal injury in lipopolysaccharide-challenged piglets. Amino Acids 2010, 39, 555–564. [Google Scholar] [CrossRef]
- Yao, K.; Yin, Y.; Li, X.; Xi, P.; Wang, J.; Lei, J.; Hou, Y.; Wu, G. Alpha-ketoglutarate inhibits glutamine degradation and enhances protein synthesis in intestinal porcine epithelial cells. Amino Acids 2012, 42, 2491–2500. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, K. Review of a histological intestinal approach to assessing the intestinal function in chickens and pigs. Anim. Sci. J. 2007, 78, 356–370. [Google Scholar] [CrossRef]
- Luk, G.D.; Bayless, T.M.; Baylin, S.B. Plasma postheparin diamine oxidase. Sensitive provocative test for quantitating length of acute intestinal mucosal injury in the rat. J. Clin. Investig. 1983, 71, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.C.; Liu, Y.; Zhu, L.; Rhoads, J.M. Arginine and citrulline protect intestinal cell monolayer tight junctions from hypoxia-induced injury in piglets. Pediatr. Res. 2012, 72, 576–582. [Google Scholar] [CrossRef]
- Beaufrère, A.M.; Neveux, N.; Patureau Mirand, P.; Buffière, C.; Marceau, G.; Sapin, V.; Cynober, L.; Meydinal-Denis, D. Long-term intermittent glutamine supplementation repairs intestinal damage (structure and functional mass) with advanced age: Assessment with plasma citrulline in a rodent model. J. Nutr. Health Aging 2014, 18, 814–819. [Google Scholar] [CrossRef]
- Wu, G. Intestinal mucosal amino acid catabolism. J. Nutr. 1998, 128, 1249–1252. [Google Scholar] [CrossRef]
- Gleeson, M. Dosing and efficacy of glutamine supplementation in human exercise and sport training. J. Nutr. 2008, 138, 2045S–2049S. [Google Scholar] [CrossRef]
- Hamilton, L.E.; Zigo, M.; Mao, J.; Xu, W.; Sutovsky, P.; O’Flaherty, C.; Oko, R. GSTO2 Isoforms Participate in the Oxidative Regulation of the Plasmalemma in Eutherian Spermatozoa during Capacitation. Antioxidants 2019, 8, 601. [Google Scholar] [CrossRef] [PubMed]
- Sepponen, K.; Pösö, A.R. The inducible form of heat shock protein 70 in the serum, colon and small intestine of the pig: Comparison to conventional stress markers. Vet. J. 2006, 171, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Zhang, X.H.; Li, X.M.; Zhou, Y.M.; Li, W.; Huang, X.X.; Zhang, L.L.; Wang, T. Intestinal growth and morphology is associated with the increase in heat shock protein 70 expression in weaning piglets through supplementation with glutamine. J. Anim. Sci. 2011, 89, 3634–3642. [Google Scholar] [CrossRef] [PubMed]
- Al-Aqil, A.; Zulkifli, I. Changes in heat shock protein 70 expression and blood characteristics in transported broiler chickens as affected by housing and early age feed restriction. Poult. Sci. 2009, 88, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Li, M.M.H. All about the RNA: Interferon-Stimulated Genes that interfere with viral RNA processes. Front. Immunol. 2020, 9, 605024. [Google Scholar] [CrossRef] [PubMed]
- Pidugu, V.K.; Pidugu, H.B.; Wu, M.M.; Liu, C.J.; Lee, T.C. Emerging functions of human IFIT proteins in cancer. Front. Mol. Biosci. 2019, 19, 148. [Google Scholar] [CrossRef]
- Ashley, C.L.; Abendroth, A.; McSharry, B.P.; Slobedman, B. Interferon-Independent Upregulation of Interferon-Stimulated Genes during Human Cytomegalovirus Infection is Dependent on IRF3 Expression. Viruses 2019, 11, 246. [Google Scholar] [CrossRef]
- Pingale, K.D.; Kanade, G.D.; Karpe, Y.A. Hepatitis E virus polymerase binds to IFIT1 to protect the viral RNA from IFIT1-mediated translation inhibition. J. Gen. Virol. 2019, 100, 471–483. [Google Scholar] [CrossRef]
- Osmanyan, A.K.; Ghazi Harsini, S.; Mahdavi, R.; Fisinin, V.I.; Arkhipova, A.L.; Glazko, T.T.; Kovalchuk, S.N.; Kosovsky, G.Y. Intestinal amino acid and peptide transporters in broiler are modulated by dietary amino acids and protein. Amino Acids 2018, 50, 353–357. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, C.; Chen, K.; Zhao, X.; Geng, Z. Effect of L-theanine on growth performance, intestinal development and health, and peptide and amino acid transporters expression of broilers. J. Sci. Food Agric. 2020, 100, 1718–1725. [Google Scholar] [CrossRef]
- Liu, Y.; Kong, X.; Li, F.; Tan, B.; Li, Y.; Duan, Y.; Yin, Y.; He, J.; Hu, C.; Blachier, F.; et al. Co-dependence of genotype and dietary protein intake to affect expression on amino acid/peptide transporters in porcine skeletal muscle. Amino Acids 2016, 48, 75–90. [Google Scholar] [CrossRef]
- Wang, C.Y.; Liu, S.; Xie, X.N.; Tan, Z.R. Regulation profile of the intestinal peptide transporter 1 (PepT1). Drug Des. Dev. Ther. 2017, 8, 3511–3517. [Google Scholar] [CrossRef]
- Spanier, B.; Rohm, F. Proton coupled oligopeptide transporter 1 (PepT1) function, regulation, and influence on the intestinal homeostasis. Compr. Physiol. 2018, 8, 843–869. [Google Scholar] [PubMed]
- Dai, Z.L.; Wu, G.; Zhu, W.Y. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front. Biosci. 2011, 16, 1768–1786. [Google Scholar] [CrossRef]
- Perna, S.; Alalwan, T.A.; Alaali, Z.; Alnashaba, T.; Gasparri, C.; Infantino, V.; Hammad, L.; Riva, A.; Petrangolini, G.; Allegrini, P.; et al. The role of Glutamine in the complex interaction between gut microbiota and health: A narrative review. Int. J. Mol. Sci. 2019, 20, 5232. [Google Scholar] [CrossRef] [PubMed]
- Preedy, V.R.; Peters, T.J. Changes in protein, RNA and DNA and rates of protein synthesis in muscle-containing tissues of the mature rat in response to ethanol feeding: A comparative study of heart, small intestine and gastrocnemius muscle. Alcohol Alcohol. 1990, 25, 489–498. [Google Scholar]
- Guay, F.; Trottier, N.L. Muscle growth and plasma concentrations of amino acids, insulin-like growth factor-I, and insulin in growing pigs fed reduced-protein diets. J. Anim. Sci. 2006, 84, 3010–3019. [Google Scholar] [CrossRef] [PubMed]
- Goron, A.; Lamarche, F.; Blanchet, S.; Delangle, P.; Schlattner, U.; Fontaine, E.; Moinard, C. Citrulline stimulates muscle protein synthesis, by reallocating ATP consumption to muscle protein synthesis. J. Cachexia Sarcopenia Muscle 2019, 10, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wu, G. Roles of dietary glycine, proline, and hydroxyproline in collagen synthesis and animal growth. Amino Acids 2018, 50, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Xu, W.D.; Chen, A.G. Effect of Gly-L-Gln on growth performance and intestinal absorption ability in weaned piglets. China J. Anim. Sci. 2005, 41, 6–8. [Google Scholar]
- He, J.; Feng, G.D.; Ao, X.; Li, Y.F.; Qian, H.X.; Liu, J.B.; Bai, G.Y.; He, Z.Z. Effects of L-glutamine on growth performance, antioxidant ability, immunity and expression of genes related to intestinal health in weanling pigs. Livest. Sci. 2016, 189, 102–109. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Items | Content | Items | Content |
|---|---|---|---|
| Ingredient | Nutrient level | ||
| Corn | 38.4 | DE (MJ/kg) | 14.27 |
| Soybean meal | 16 | CP | 18.54 |
| Flour | 12 | Lys | 1.5 |
| Whey powder (low protein) | 8 | Met | 0.42 |
| Soybean protein concentrate | 5 | Cys | 0.29 |
| Wheat middling | 5 | Thr | 0.93 |
| Fish meal | 4.5 | Trp | 0.23 |
| Glucose | 3 | Ca | 0.75 |
| CaHPO4 | 1.33 | AP | 0.49 |
| Limestone | 0.37 | TP | 0.68 |
| NaCl | 0.25 | Na | 0.31 |
| Plant oil | 3.95 | CF | 6.26 |
| Premix * | 1 | NaCl | 0.61 |
| Lys | 0.64 | ||
| Met | 0.13 | ||
| Thr | 0.21 | ||
| Choline | 0.12 | ||
| Mildew preventive | 0.1 |
| Items | Primer Sequence (5′-3′) |
|---|---|
| Total bacteria count | (F) CGGTCCAGACTCCTACGGG |
| (R) TTACCGCGGCTGCTGGCAC | |
| Enterococcus genus | (F) CCCTTATTGTTAGTTGCCATCATT |
| (R) ACTCGTTGTACTTCCCATTGT | |
| Enterobacterium family | (F) CATTGACGTTACCCGCAGAAGAAGC |
| (R) CTCTACGAGACTCAAGCTTGC | |
| Clostridium coccoides | (F) AATGACGGTACCTGACTAA |
| (R) CTTTGAGTTTCATTCTTGCGAA | |
| Lactobacillus genus | (F) TCGCGTC(C/T)GGTGTGAAAG |
| (R) CCACATCCAGC(A/G)TCCAC | |
| Internal reference | (F) CAGAAATGGGAATGGAAAGTTG |
| (R) CCATTGGTCAGGTCATTCAATACA | |
| ASCT2 | (F) GCCAGCAAGATTGTGGAGAT |
| (R) GAGCTGGATGAGGTTCCAAA | |
| GSTO2 | (F) GCCTTGAGATGTGGGAGAGAA |
| (R) AAGATGGTGTTCTGATAGCCAAGA | |
| IFIT1 | (F) GCTAAACCAAACACCGCAGAA |
| (R) GGAACTCAATCTCCTCCAAGACC | |
| IFIT2 | (F) CAGAAGGCGGCAGAGAATG |
| (R) ACACAGAGGCAGGCGAGATAG | |
| IFIT3 | (F) GCATTTTCCAGCCAGCATC |
| (R) TCTGTTCCTTTCCTTTCCTTCCT | |
| IFIT5 | (F) CAGAAAATACAGCCATCCACCA |
| (R) AGGGCACTTAAACTCTGCACATC | |
| IFN-α | (F) ACTCCATCCTGGCTGTGAGGAAAT |
| (R) ATCTCATGACTTCTGCCCTGACGA | |
| IFN-β | (F) AGCAGATCTTCGGCATTCTC |
| (R) GTCATCCATCTGCCCATCAA | |
| MSTN | (F) GAAGTCAAGGTAACAGACACACCAA |
| (R) GCAATAATCCAGTCCCATCCA | |
| MX1 | (F) AGTGCGGCTGTTTACCAAG |
| (R) TTCACAAACCCTGGCAACTC | |
| MX2 | (F) CGCATTCTTTCACTCGCATC |
| (R) CCTCAACCCACCAACTCACA | |
| Myf5 | (F) CCACGACTAACCCCAACCA |
| (R) TTTTCCACCTGCTCCCTCA | |
| OAS1 | (F) TGGTGGTGGAGACACACACA |
| (R) CCAACCAGAGACCCATCCA | |
| OASL | (F) GGCACCCCTGTTTTCCTCT |
| (R) AGCACCGCTTTTGGATGG | |
| PepT1 | (F) ATTCTCAGGCTCCTTCCAACA |
| (R) GCAACCCCGCAAACAGA | |
| SLC7A6 | (F) CTGCCGCCTGCATGTGT |
| (R) TGTGCCCCACTTGACATAGG | |
| SLC7A7 | (F) TTTGGTTCCCAAGGTTGCA |
| (R) GCAGCTTCCTGGCATTGC | |
| SLC6A19 | (F) CGAGTACCCGTACCTGATGGA |
| (R) TGCGTAGAAGGGCGAAGAA |
| Items | Control Group | Gln Group | p-Value |
|---|---|---|---|
| Plasma | |||
| DAO (mmol/L) | 15.36 ± 2.49 a | 9.38 ± 3.54 b | 0.002 |
| Duodenum | |||
| Villus height (μm) | 282 ± 24 b | 349 ± 13 a | <0.01 |
| Crypt depth (μm) | 119 ± 7 | 121 ± 7 | 0.574 |
| Villus height/crypt depth | 2.45 ± 0.16 b | 2.89 ± 0.23 a | <0.01 |
| Villus width (μm) | 140 ± 8 b | 161 ± 5 a | <0.01 |
| Villus surface area (μm2) | 32,431 ± 3438 b | 44,744 ± 3273 a | <0.01 |
| Jejunum | |||
| Villus height (μm) | 260 ± 21 b | 311 ± 31 a | 0.002 |
| Crypt depth (μm) | 99 ± 7 | 103 ± 16 | 0.564 |
| Villus height/crypt depth | 2.60 ± 0.14 b | 3.06 ± 0.36 a | 0.006 |
| Villus width (μm) | 104 ± 5 b | 124 ± 8 a | <0.01 |
| Villus surface area (μm2) | 20,481 ± 3519 b | 26,809 ± 3987 a | 0.005 |
| Ileum | |||
| Villus height (μm) | 231 ± 16 b | 290 ± 22 a | <0.01 |
| Crypt depth (μm) | 110 ± 12 | 114 ± 16 | 0.631 |
| Villus height/crypt depth | 2.18 ± 0.10 b | 2.59 ± 0.30 a | 0.002 |
| Villus width (μm) | 106 ± 6 b | 120 ± 10 a | 0.005 |
| Villus surface area (μm2) | 18,462 ± 1742 b | 25,847 ± 3745 a | <0.01 |
| Items | Control Group | Gln Group | p-Value |
|---|---|---|---|
| Duodenum | |||
| TP (mg/g) | 67.20 ± 7.10 | 66.00 ± 3.90 | 0.694 |
| RNA (mg/g) | 2.51 ± 0.32 | 2.58 ± 0.34 | 0.522 |
| DNA (mg/g) | 0.36 ± 0.07 | 0.40 ± 0.07 | 0.234 |
| RNA/DNA | 7.27 ± 1.66 | 6.91 ± 1.84 | 0.256 |
| TP/DNA | 196.64 ± 45.10 | 175.37 ± 35.97 | 0.716 |
| Jejunum | |||
| TP (mg/g) | 51.50 ± 7.60 b | 62.70 ± 6.10 a | 0.006 |
| RNA (mg/g) | 1.70 ± 0.33 | 1.75 ± 0.24 | 0.744 |
| DNA (mg/g) | 0.21 ± 0.02 | 0.19 ± 0.02 | 0.162 |
| RNA/DNA | 7.79 ± 1.04 b | 8.80 ± 0.46 a | <0.01 |
| TP/DNA | 255.12 ± 61.68 b | 319.59 ± 52.72 a | 0.001 |
| Ileum | |||
| TP (mg/g) | 31.90 ± 6.70 b | 58.90 ± 7.30 a | <0.01 |
| RNA (mg/g) | 2.24 ± 0.36 | 2.22 ± 0.30 | 0.709 |
| DNA (mg/g) | 0.18 ± 0.026 a | 0.14 ± 0.01 b | 0.001 |
| RNA/DNA | 11.49 ± 2.67 b | 15.25 ± 2.85 a | 0.020 |
| TP/DNA | 166.67 ± 30.90 b | 402.96 ± 61.66 a | <0.01 |
| Amino Acids (μg/g) | Control Group | Gln Group | p-Value |
|---|---|---|---|
| Jejunum | |||
| 1-Methylhistidine | 4.081 ± 1.896 | 4.103 ± 2.893 | 0.986 |
| Arginine | 216.335 ± 61.647 | 210.695 ± 66.219 | 0.863 |
| Carnosine | 6.582 ± 1.502 a | 4.55 ± 1.153 b | 0.009 |
| Citrulline | 94.349 ± 26.045 b | 160.174 ± 47.633 a | 0.004 |
| Cystine | 2.489 ± 0.649 | 4.731 ± 3.406 | 0.089 |
| Glutamine | 833.294 ± 133.851 a | 638.748 ± 108.339 b | 0.006 |
| Hydroxyproline | 112.917 ± 5.053 | 110.441 ± 15.041 | 0.666 |
| Ornithine | 45.68 ± 7.7 | 43.54 ± 5.49 | 0.533 |
| β-Alanine | 60.883 ± 8 | 74.941 ± 21.413 | 0.104 |
| β-Aminoisobutyric acid | 17.703 ± 6.117 | 20.05 ± 23.795 | 0.791 |
| Ileum | |||
| 1-Methylhistidine | 2.956 ± 0.463 a | 1.869 ± 0.566 b | 0.001 |
| Arginine | 187.077 ± 19.474 | 184.844 ± 39.183 | 0.887 |
| Carnosine | 7.565 ± 2.58 | 7.607 ± 2.256 | 0.973 |
| Citrulline | 62.951 ± 10.771 b | 87.347 ± 22.556 a | 0.015 |
| Cystine | 2.939 ± 0.613 | 3.598 ± 1.069 | 0.152 |
| Glutamine | 983.157 ± 186.976 a | 679.397 ± 140.461 b | 0.003 |
| Hydroxyproline | 167.301 ± 17.16 a | 142.792 ± 7.786 b | 0.002 |
| Ornithine | 110.647 ± 18.887 | 120.151 ± 52.105 | 0.635 |
| β-Alanine | 67.41 ± 16.363 b | 86.473 ± 17.62 a | 0.042 |
| β-Aminoisobutyric acid | 2.676 ± 0.613 a | 1.425 ± 0.133 b | <0.01 |
| Items | Control Group | Gln Group | p-Value |
|---|---|---|---|
| Duodenum | |||
| GSTO2 | 1.000 ± 0.213 b | 3.749 ± 0.913 a | <0.01 |
| IFN-β | 1.000 ± 0.119 b | 2.684 ± 0.649 a | <0.01 |
| Jejunum | |||
| GSTO2 | 1.000 ± 0.257 | 1.254 ± 0.274 | 0.077 |
| IFIT1 | 1.000 ± 0.281 a | 0.532 ± 0.138 b | 0.001 |
| IFIT2 | 1.000 ± 0.248 a | 0.613 ± 0.163 b | 0.002 |
| IFIT3 | 1.000 ± 0.163 a | 0.661 ± 0.139 b | 0.001 |
| IFIT5 | 1.000 ± 0.119 a | 0.659 ± 0.158 b | <0.01 |
| IFN-α | 1.000 ± 0.245 b | 1.286 ± 0.272 a | 0.044 |
| MX1 | 1.000 ± 0.143 a | 0.464 ± 0.106 b | <0.01 |
| MX2 | 1.000 ± 0.265 a | 0.600 ± 0.155 b | 0.002 |
| OAS1 | 1.000 ± 0.272 a | 0.670 ± 0.175 b | 0.012 |
| OASL | 1.000 ± 0.281 a | 0.446 ± 0.111 b | <0.01 |
| SLC7A6 | 1.000 ± 0.154 b | 1.227 ± 0.236 a | 0.039 |
| Ileum | |||
| GSTO2 | 1.000 ± 0.118 b | 2.005 ± 0.399 a | <0.01 |
| IFIT1 | 1.000 ± 0.224 a | 0.607 ± 0.135 b | 0.001 |
| IFN-α | 1.000 ± 0.169 b | 1.321 ± 0.275 a | 0.014 |
| IFN-β | 1.000 ± 0.101 b | 1.392 ± 0.293 a | 0.003 |
| MX1 | 1.000 ± 0.161 a | 0.712 ± 0.148 b | 0.002 |
| SLC7A7 | 1.000 ± 0.235 b | 1.291 ± 0.251 a | 0.031 |
| Colon bacteria | |||
| Clostridium coccoides | 1.000 ± 0.154 a | 0.335 ± 0.059 b | <0.01 |
| Enterobacterium family | 1.000 ± 0.136 a | 0.354 ± 0.044 b | <0.01 |
| Enterococcus genus | 1.000 ± 0.246 a | 0.728 ± 0.181 b | 0.024 |
| Lactobacillus genus | 1.000 ± 0.242 b | 2.450 ± 0.449 a | <0.01 |
| Amino Acids | Control Group | Gln Group | p-Value |
|---|---|---|---|
| 1-Methylhistidine (μg/g) | 3.955 ± 1.209 | 3.743 ± 1.437 | 0.755 |
| Arginine (μg/g) | 56.966 ± 8.866 b | 107.416 ± 52.704 a | 0.018 |
| Carnosine (ng/g) | 13.65 ± 0.916 | 14.044 ± 1.411 | 0.519 |
| Citrulline (μg/g) | 57.191 ± 11.348 b | 82.69 ± 14.84 a | 0.002 |
| Glutamine (ng/g) | 2.075 ± 0.328 a | 1.629 ± 0.233 b | 0.007 |
| Hydroxyproline (μg/g) | 94.618 ± 12.03 b | 124.028 ± 23.401 a | 0.007 |
| Ornithine (μg/g) | 155.918 ± 33.51 b | 222.05 ± 59.788 a | 0.016 |
| β-Alanine (ng/g) | 0.962 ± 0.107 | 0.857 ± 0.225 | 0.254 |
| β-Aminoisobutyric acid (μg/g) | 14.926 ± 5.312 | 16.039 ± 3.385 | 0.625 |
| Items | Control Group | Gln Group | p-Value |
|---|---|---|---|
| TP (mg/g) | 29.1 ± 2.2 b | 37.4 ± 5.1 a | 0.001 |
| RNA (mg/g) | 0.060 ± 0.017 a | 0.035 ± 0.007 b | <0.01 |
| DNA (mg/g) | 0.336 ± 0.021 b | 0.386 ± 0.049 a | 0.003 |
| RNA/DNA | 0.170 ± 0.042 a | 0.091 ± 0.020 b | <0.01 |
| TP/DNA | 87.771 ± 5.219 b | 100.078 ± 11.917 a | <0.01 |
| Genes | |||
| ASCT2 | 1.000 ± 0.212 | 0.847 ± 0.179 | 0.140 |
| SLC6A19 | 1.000 ± 0.230 | 1.102 ± 0.185 | 0.343 |
| MSTN | 1.000 ± 0.202 | 0.964 ± 0.217 | 0.738 |
| Myf5 | 1.000 ± 0.164 | 1.087 ± 0.187 | 0.339 |
| PepT1 | 1.00 ± 0.176 b | 1.569 ± 0.408 a | 0.003 |
| SLC7A7 | 1.00 ± 0.189 | 0.878 ± 0.197 | 0.228 |
| SLC7A6 | 1.000 ± 0.250 | 1.065 ± 0.249 | 0.611 |
| Items | Control Group | Gln Group | p-Value |
|---|---|---|---|
| CAT (U/mg protein) | 1.11 ± 0.33 | 1.09 ± 0.30 | 0.910 |
| MDA (nmol/mg protein) | 0.65 ± 0.15 a | 0.41 ± 0.20 b | 0.020 |
| H2O2 (μmol/mg protein) | 1.59 ± 0.53 a | 1.11 ± 0.25 b | 0.009 |
| SOD (U/mg protein) | 73.1 ± 12.03 b | 83.8 ± 7.11 a | 0.048 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Shen, M.; Liu, J.; Zhang, Y.; Zhu, Z.; Li, B.; Guo, S.; Yi, D.; Ding, B.; Wu, T.; et al. Effects of Dietary l-Glutamine Supplementation on the Intestinal Function and Muscle Growth of Piglets. Life 2024, 14, 405. https://doi.org/10.3390/life14030405
Wang L, Shen M, Liu J, Zhang Y, Zhu Z, Li B, Guo S, Yi D, Ding B, Wu T, et al. Effects of Dietary l-Glutamine Supplementation on the Intestinal Function and Muscle Growth of Piglets. Life. 2024; 14(3):405. https://doi.org/10.3390/life14030405
Chicago/Turabian StyleWang, Lei, Meng Shen, Jiale Liu, Yanyan Zhang, Zhekun Zhu, Baocheng Li, Shuangshuang Guo, Dan Yi, Binying Ding, Tao Wu, and et al. 2024. "Effects of Dietary l-Glutamine Supplementation on the Intestinal Function and Muscle Growth of Piglets" Life 14, no. 3: 405. https://doi.org/10.3390/life14030405