
AKT, p-AKT, ERK1/2 and p-ERK1/2 in Mural Granulosa Cells Are Not Correlated to Different Ovarian Stimulation Protocols in Patients Undergoing Assisted Reproductive Treatment


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Patients
2.2. Ovarian Stimulation
2.3. Granulosa Cells Preparation
2.4. SDS-PAGE and Western Blot
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Report of the Meeting on the Prevention of Infertility at the Primary Health Care Level 12–16 December 1983, Geneva; World Health Organization: Geneva, Switzerland, 1984.
- Alteri, A.; Arroyo, G.; Baccino, G.; Craciunas, L.; De Geyter, C.; Ebner, T.; Koleva, M.; Kordic, K.; McHeik, S.; Mertes, H.; et al. ESHRE guideline: Number of embryos to transfer during IVF/ICSIdagger. Hum. Reprod. 2024, 39, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Bromer, J.G.; Seli, E. Assessment of embryo viability in assisted reproductive technology: Shortcomings of current approaches and the emerging role of metabolomics. Curr. Opin. Obstet. Gynecol. 2008, 20, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Herrero, J.; Meseguer, M. Selection of high potential embryos using time-lapse imaging: The era of morphokinetics. Fertil. Steril. 2013, 99, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Serhal, P.F.; Ranieri, D.M.; Kinis, A.; Marchant, S.; Davies, M.; Khadum, I.M. Oocyte morphology predicts outcome of intracytoplasmic sperm injection. Hum. Reprod. 1997, 12, 1267–1270. [Google Scholar] [CrossRef] [PubMed]
- Ruvolo, G.; Fattouh, R.R.; Bosco, L.; Brucculeri, A.M.; Cittadini, E. New molecular markers for the evaluation of gamete quality. J. Assist. Reprod. Genet. 2013, 30, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Kidder, G.M.; Vanderhyden, B.C. Bidirectional communication between oocytes and follicle cells: Ensuring oocyte developmental competence. Can. J. Physiol. Pharmacol. 2010, 88, 399–413. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente, R.; Eppig, J.J. Transcriptional activity of the mouse oocyte genome: Companion granulosa cells modulate transcription and chromatin remodeling. Dev. Biol. 2001, 229, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Canipari, R. Oocyte-granulosa cell interactions. Hum. Reprod. Update 2000, 6, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Chronowska, E. High-throughput analysis of ovarian granulosa cell transcriptome. Biomed. Res. Int. 2014, 2014, 213570. [Google Scholar] [CrossRef]
- Uyar, A.; Torrealday, S.; Seli, E. Cumulus and granulosa cell markers of oocyte and embryo quality. Fertil. Steril. 2013, 99, 979–997. [Google Scholar] [CrossRef]
- Emanuelli, I.P.; Costa, C.B.; Rafagnin Marinho, L.S.; Seneda, M.M.; Meirelles, F.V. Cumulus-oocyte interactions and programmed cell death in bovine embryos produced in vitro. Theriogenology 2019, 126, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.L.; Redmer, D.A.; Reynolds, L.P.; Bilski, J.J.; Grazul-Bilska, A.T. Gap junctional intercellular communication of bovine granulosa and thecal cells from antral follicles: Effects of luteinizing hormone and follicle-stimulating hormone. Endocrine 2002, 18, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Scott, R., 3rd; Zhang, M.; Seli, E. Metabolism of the oocyte and the preimplantation embryo: Implications for assisted reproduction. Curr. Opin. Obstet. Gynecol. 2018, 30, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.P.; Ferreira, M.C.F.; Silveira, C.O.; Campos, J.R.; Borges, I.T.; Baeta, P.G.; Silva, F.H.S.; Reis, F.M.; Del Puerto, H.L. Clinical correlation of apoptosis in human granulosa cells-A review. Cell Biol. Int. 2018, 42, 1276–1281. [Google Scholar] [CrossRef] [PubMed]
- Varras, M.; Griva, T.; Kalles, V.; Akrivis, C.; Paparisteidis, N. Markers of stem cells in human ovarian granulosa cells: Is there a clinical significance in ART? J. Ovarian Res. 2012, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Bosco, L.; Chiarelli, R.; Roccheri, M.C.; Matranga, D.; Ruvolo, G. Relationship between apoptosis and survival molecules in human cumulus cells as markers of oocyte competence. Zygote 2017, 25, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Ruvolo, G.; Roccheri, M.C.; Luparello, C.; Matranga, D.; Ferrigno, A.; Bosco, L. DNA fragmentation index, pAKT and pERK1/2 in cumulus cells are related to oocyte competence in patients undergoing in vitro fertilization programme. Zygote 2019, 27, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, K.; Saito, H.; Saito, T.; Ito, M.; Ohta, N.; Takahashi, T.; Hiroi, M. The incidence of apoptotic bodies in membrana granulosa can predict prognosis of ova from patients participating in in vitro fertilization programs. Fertil. Steril. 1997, 68, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Nakahara, K.; Saito, H.; Saito, T.; Ito, M.; Ohta, N.; Takahashi, T.; Hiroi, M. Ovarian fecundity in patients with endometriosis can be estimated by the incidence of apoptotic bodies. Fertil. Steril. 1998, 69, 931–935. [Google Scholar] [CrossRef]
- Sadraie, S.H.; Saito, H.; Kaneko, T.; Saito, T.; Hiroi, M. Effects of aging on ovarian fecundity in terms of the incidence of apoptotic granulosa cells. J. Assist. Reprod. Genet. 2000, 17, 168–173. [Google Scholar] [CrossRef]
- Corn, C.M.; Hauser-Kronberger, C.; Moser, M.; Tews, G.; Ebner, T. Predictive value of cumulus cell apoptosis with regard to blastocyst development of corresponding gametes. Fertil. Steril. 2005, 84, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.S.; Cui, W. Proliferation, survival and metabolism: The role of PI3K/AKT/mTOR signalling in pluripotency and cell fate determination. Development 2016, 143, 3050–3060. [Google Scholar] [CrossRef] [PubMed]
- Benbrook, D.M.; Masamha, C.P. The pro-survival function of Akt kinase can be overridden or altered to contribute to induction of apoptosis. Curr. Cancer Drug Targets 2011, 11, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Sinha, D.; Bannergee, S.; Schwartz, J.H.; Lieberthal, W.; Levine, J.S. Inhibition of ligand-independent ERK1/2 activity in kidney proximal tubular cells deprived of soluble survival factors up-regulates Akt and prevents apoptosis. J. Biol. Chem. 2004, 279, 10962–10972. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.; Iwase, A.; Ando, H.; Kurotsuchi, S.; Harata, T.; Kikkawa, F. PTEN and Akt expression during growth of human ovarian follicles. J. Assist. Reprod. Genet. 2007, 24, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.L.; Cowan, R.G.; Harman, R.M.; Quirk, S.M. Cell cycle progression and activation of Akt kinase are required for insulin-like growth factor I-mediated suppression of apoptosis in granulosa cells. Mol. Endocrinol. 2004, 18, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Chang, Y.; Wei, L.; Chen, J.; Li, J.; Goldsmith, S.; Silber, S.; Liang, X. Apoptosis of mural granulosa cells is increased in women with diminished ovarian reserve. J. Assist. Reprod. Genet. 2019, 36, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Jancar, N.; Kopitar, A.N.; Ihan, A.; Virant Klun, I.; Bokal, E.V. Effect of apoptosis and reactive oxygen species production in human granulosa cells on oocyte fertilization and blastocyst development. J. Assist. Reprod. Genet. 2007, 24, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Ruvolo, G.; Bosco, L.; Pane, A.; Morici, G.; Cittadini, E.; Roccheri, M.C. Lower apoptosis rate in human cumulus cells after administration of recombinant luteinizing hormone to women undergoing ovarian stimulation for in vitro fertilization procedures. Fertil. Steril. 2007, 87, 542–546. [Google Scholar] [CrossRef]
- Han, Z.B.; Lan, G.C.; Wu, Y.G.; Han, D.; Feng, W.G.; Wang, J.Z.; Tan, J.H. Interactive effects of granulosa cell apoptosis, follicle size, cumulus-oocyte complex morphology, and cumulus expansion on the developmental competence of goat oocytes: A study using the well-in-drop culture system. Reproduction 2006, 132, 749–758. [Google Scholar] [CrossRef]
- Jiang, J.Y.; Xiong, H.; Cao, M.; Xia, X.; Sirard, M.A.; Tsang, B.K. Mural granulosa cell gene expression associated with oocyte developmental competence. J. Ovarian Res. 2010, 3, 6. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Ran, J.; Zhu, S.; Quan, S.; Ye, B.; Yu, A.; Kang, Y.; Lin, Y. Effect of GOLPH3 on cumulus granulosa cell apoptosis and ICSI pregnancy outcomes. Sci. Rep. 2017, 7, 7863. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, L.A.; Egbert, J.R. Regulation of Mammalian Oocyte Meiosis by Intercellular Communication within the Ovarian Follicle. Annu. Rev. Physiol. 2017, 79, 237–260. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group | A | B | p Value |
|---|---|---|---|
| Patient, n | 18 | 14 | p = 0.480 |
| Total oocytes, n | 56 | 45 | p = 0.562 |
| MII oocytes, n | 54 | 43 | p = 0.917 |
| Nr. fertilized oocytes, n (%) | 50 (93) | 39 (91) | p = 0.830 |
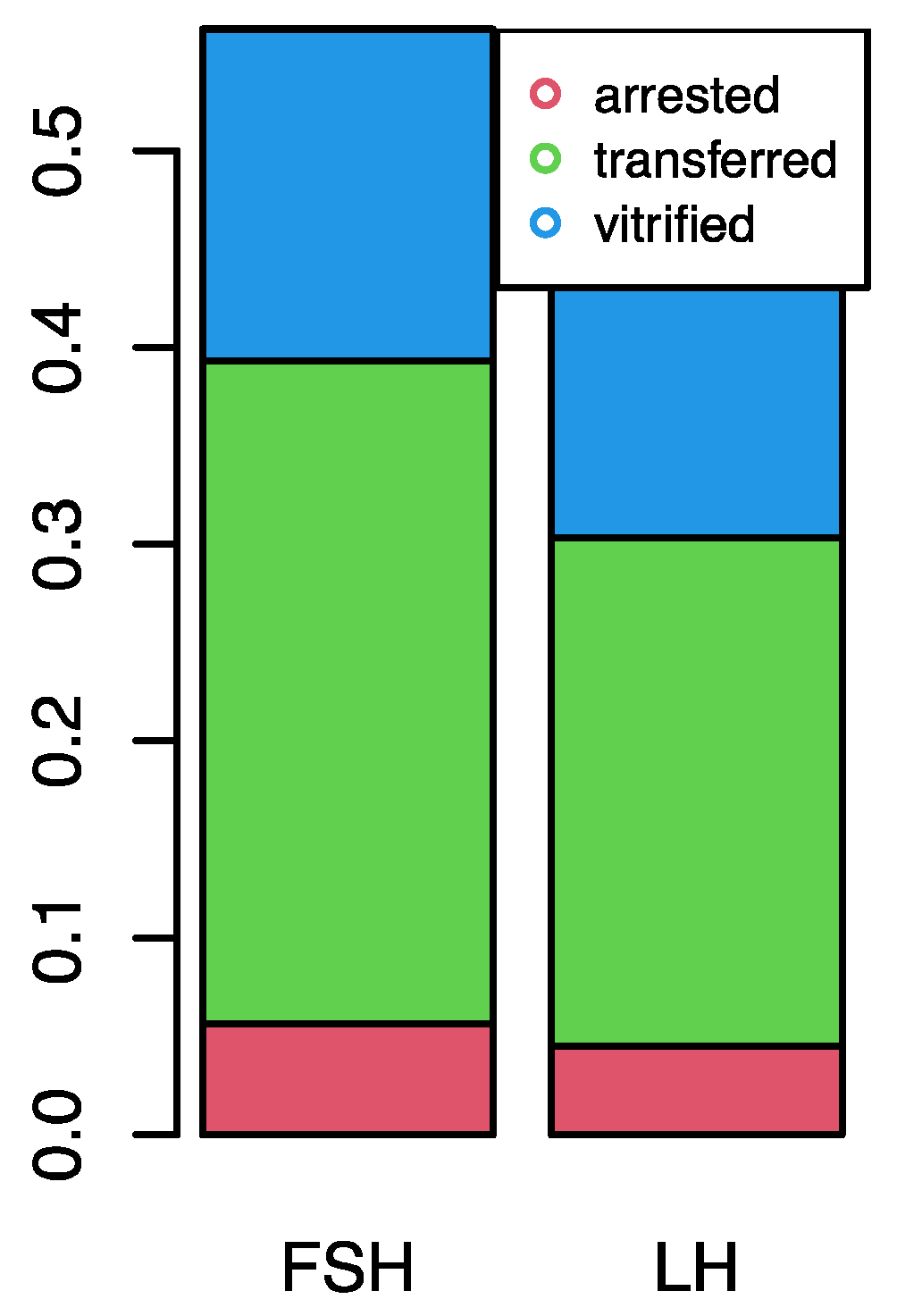
| Nr. arrested embryos, n (%) | 5 (10) | 4 (10) | p = 0.982 |
| Nr. transferred embryos, n (%) | 30 (60) | 23 (59) | p = 0.956 |
| Nr. vitrified embryos, n (%) | 15 (30) | 12 (31) | p = 0.965 |
| Nr. clinical pregnancies, n (%) | 6 (33) | 6 (43) | p = 0.581 |
| Implantation rate, % | 36 | 42 | p = 0.722 |
| Group A r-FSH (n = 18) (MII Oocytes = 54) | Group B r-FSH + r-LH (n = 14) (MII Oocytes = 43) | p-Value § | |
|---|---|---|---|
| Median (Q1–Q3) | Median (Q1–Q3) | ||
| AKT | 4.08 (1.55–10.20) | 2.51 (1.49–9.61) | 0.683 |
| p-AKT | 5.22 (3.13–11.67) | 3.96 (1.73–11.92) | 0.410 |
| ERK 1/2 | 3.92 (2.02–10.32) | 3.05 (1.82–10.59) | 0.750 |
| p-ERK1/2 | 5.14 (2.30–11.16) | 6.59 (2.96–12.45) | 0.743 |
| Arrested (n =9) Median (Q1–Q3) | Transferred (n = 53) Median (Q1–Q3) | p-Value § | |
|---|---|---|---|
| AKT | 5.22 (3.55–11.25) | 3.45 (1.49–8.76) | 0.350 |
| p-AKT | 7.54 (3.73–12.24) | 4.63 (1.98–11.05) | 0.876 |
| ERK1/2 | 4.75 (2.67–7.09) | 3.06 (1.93–10.40) | 0.369 |
| p-ERK1/2 | 4.02 (0.73–5.85) | 4.93 (2.83–10.37) | 0.424 |
| AdjOR (95% CI) | p-Value | |
|---|---|---|
| AKT | 1.03 (0.95–1.12) | 0.429 |
| p-AKT | 0.98 (0.96–1.01) | 0.249 |
| ERK1/2 | 0.98 (0.94–1.02) | 0.290 |
| p-ERK1/2 | 0.98 (0.95–1.01) | 0.150 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruvolo, G.; Matranga, D.; Barreca, M.M.; Bosco, L. AKT, p-AKT, ERK1/2 and p-ERK1/2 in Mural Granulosa Cells Are Not Correlated to Different Ovarian Stimulation Protocols in Patients Undergoing Assisted Reproductive Treatment. Life 2024, 14, 554. https://doi.org/10.3390/life14050554
Ruvolo G, Matranga D, Barreca MM, Bosco L. AKT, p-AKT, ERK1/2 and p-ERK1/2 in Mural Granulosa Cells Are Not Correlated to Different Ovarian Stimulation Protocols in Patients Undergoing Assisted Reproductive Treatment. Life. 2024; 14(5):554. https://doi.org/10.3390/life14050554
Chicago/Turabian StyleRuvolo, Giovanni, Domenica Matranga, Maria Magdalena Barreca, and Liana Bosco. 2024. "AKT, p-AKT, ERK1/2 and p-ERK1/2 in Mural Granulosa Cells Are Not Correlated to Different Ovarian Stimulation Protocols in Patients Undergoing Assisted Reproductive Treatment" Life 14, no. 5: 554. https://doi.org/10.3390/life14050554