
Salt Acclimation of Cyanobacteria and Their Application in Biotechnology

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
Goal of the Review
2. Compatible Solutes–Universal & Useful
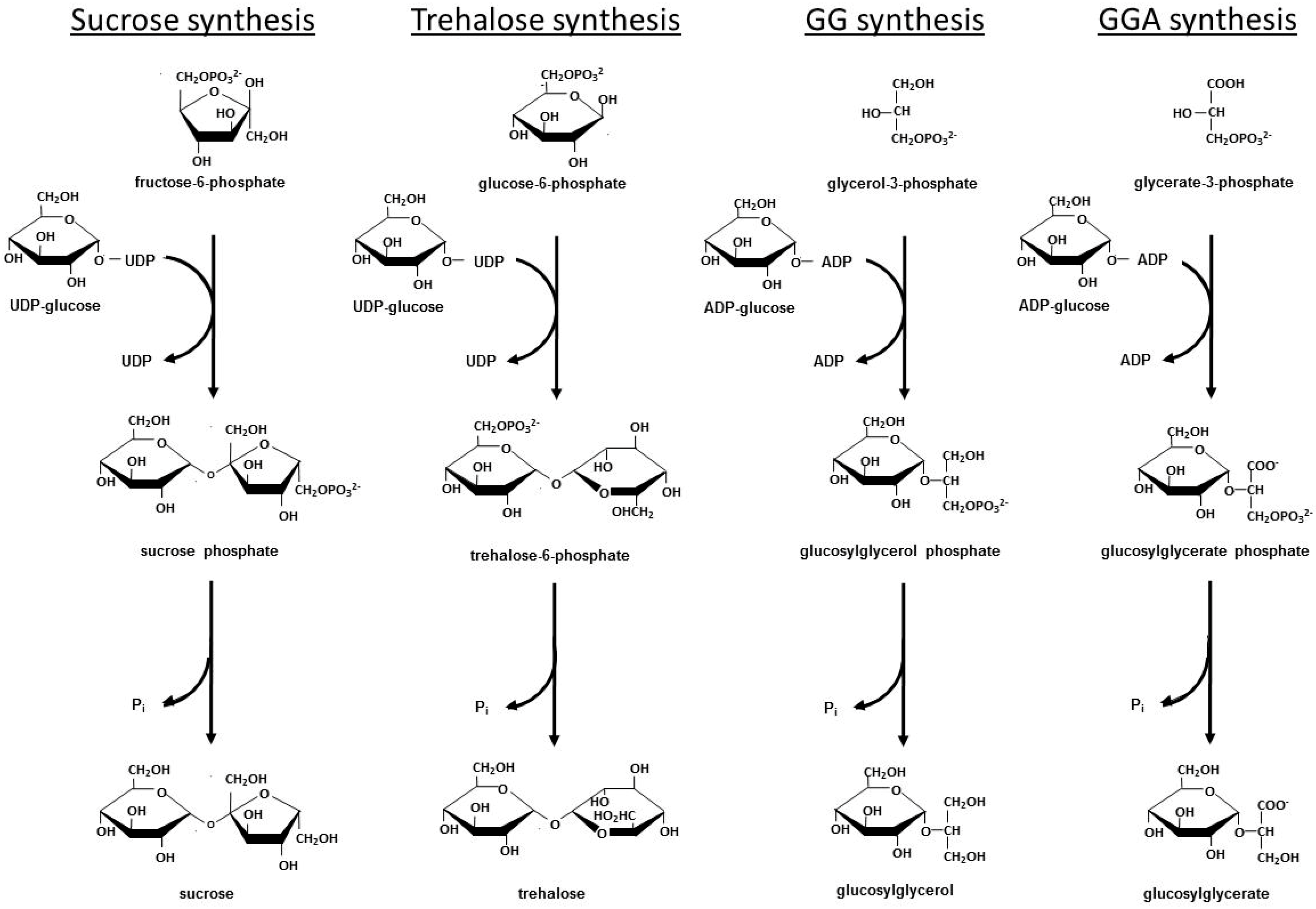
2.1. Insights Regarding Compatible Solute Synthesis

2.2. Compatible Solute Transport

3. Ion Transport
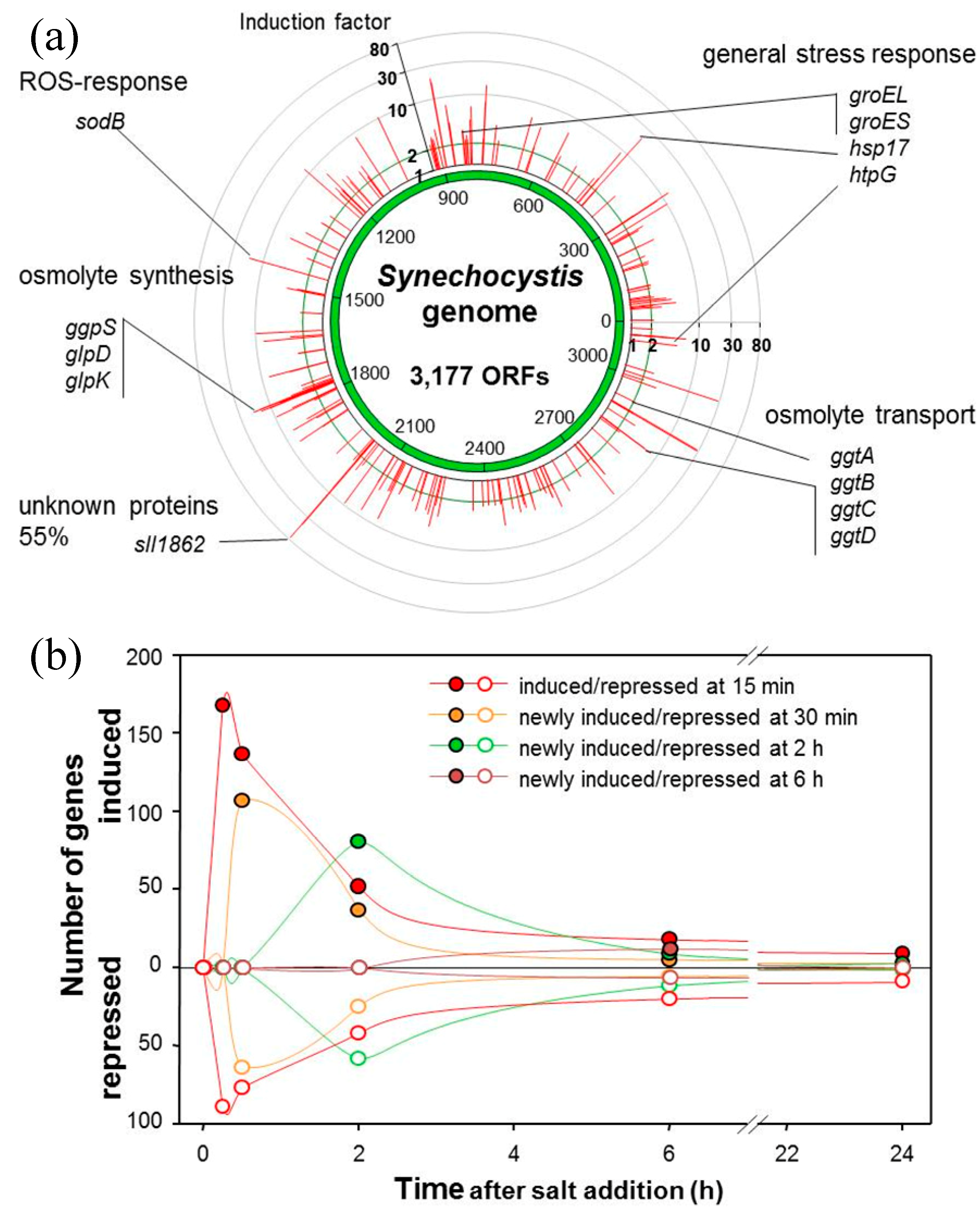
4. Salt Stress and “omics”

5. Sensing Salt Stress and Regulation of the Salt Acclimation Response
6. Salt Stress and Synthesis of Biotechnological Products
7. Conclusions
Acknowledgment
Author Contributions
Conflicts of Interest
References
- Tandeau de Marsac, N.T.; Houmard, J. Adaptation of cyanobacteria to environmental stimuli: New steps toward molecular mechanisms. FEMS Microbiol. Rev. 1993, 104, 119–190. [Google Scholar]
- Oren, A. Salts and brines. In The Ecology of Cyanobacteria; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, the Netherlands, 2000; pp. 281–306. [Google Scholar]
- Ladas, N.P.; Papageorigou, G.C. Cell turgor: A critical factor for the proliferation of cyanobacteria at unfavorable salinity. Photosynth Res. 2000, 65, 155–164. [Google Scholar] [PubMed]
- Galinski, E.A.; Trüper, H.G. Microbial behaviour in salt-stressed ecosystems. FEMS Microbiol. Rev. 1994, 15, 95–108. [Google Scholar] [CrossRef]
- Hagemann, M. Molecular biology of cyanobacterial salt acclimation. FEMS Microbiol. Rev. 2011, 35, 87–123. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. The genera Haloanaerobium, Halobacteroides and Sporohalobacter. In The Prokaryotes; A Handbook on the Biology of Bacteria: Ecophysiology, Isolation, Identification, Applications; Balows, A., Trüper, H.G., Dworkin, M., Harder, W., Schleifer, K.-H., Eds.; Springer: New York, NY, USA, 1993; pp. 1983–1900. [Google Scholar]
- Oren, A.; Mana, L. Amino acid composition of bulk protein and salt relationships of selected enzymes of Salinibacter ruber, an extremely halophilic bacterium. Extremophiles 2002, 6, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Bioenergetic aspects of halophilism. Microbiol. Mol. Biol. Rev. 1999, 63, 334–348. [Google Scholar] [PubMed]
- Müller, V.; Oren, A. Metabolism of chloride in halophilic prokaryotes. Extremophiles 2003, 7, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.D. Microbial water stress. Bacteriol. Rev. 1976, 40, 803–846. [Google Scholar] [PubMed]
- Bremer, E.; Krämer, R. Coping with osmotic challenges: osmoregulation through accumulation and release of compatible solute in bacteria. In Bacterial Stress Responses; Hengge-Aronis, R., Ed.; ASM Press: Washington, DC, USA, 2000; pp. 79–96. [Google Scholar]
- Wood, J. Bacterial osmoregulation: a paradigm for the study of cellular homeostasis. Annu. Rev. Microbiol. 2011, 65, 215–238. [Google Scholar] [CrossRef]
- Krämer, R. Bacterial stimulus perception and signal transduction: Response to osmotic stress. Chem. Rec. 2010, 10, 217–229. [Google Scholar] [CrossRef]
- Kempf, B.; Bremer, E. Uptake and synthsis of compatible solutes as microbial stress response to high-osmolality environment. Arch. Microbiol. 1998, 170, 319–330. [Google Scholar] [CrossRef]
- Potts, M. Desiccation tolerance of prokaryotes. Microbiol. Rev. 1994, 58, 755–805. [Google Scholar] [PubMed]
- Klähn, S.; Hagemann, M. Compatible solute biosynthesis in cyanobacteria. Environ. Microbiol. 2011, 13, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, M. Genomics of salt acclimation: synthesis of compatible solutes among cyanobacteria. In Genomics of Cyanobacteria; Chauvat, F., Cassier-Chauvat, C., Eds.; Academic Press: Waltham, MA, USA, 2013; Volum 65, pp. 27–55. [Google Scholar]
- Pade, N.; Hagemann, M. Cyanobacterial salt stress acclimation: genetic manipulation and regulation. In Stress Biology of Cyanobacteria: Molecular Mechanisms to Cellular Responses; Srivastava, A.K., Rai, A.N., Neilan, B.A., Eds.; CRC Press: London, UK, 2013; pp. 189–202. [Google Scholar]
- Arakawa, T.; Timasheff, S.N. The stabilization of proteins by osmolytes. Biophys. J. 1985, 47, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.M. Osmosensing by bacteria: Signals and membrane-based sensors. Micobiol. Mol. Biol. Rev. 1999, 63, 230–262. [Google Scholar]
- Poolman, B.; Glaasker, E. Regulation of compatible solute accumulation in bacteria. Mol. Microbiol. 1998, 29, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Reed, R.H.; Borowitzka, L.J.; Mackay, M.A.; Chudek, J.A.; Foster, R.; Warr, S.R.C.; Moore, D.J.; Stewart, W.D.P. Organic solute accumulation in osmotically stressed cyanobacteria. FEMS Microbiol. Lett. 1986, 39, 51–56. [Google Scholar] [CrossRef]
- Mackay, M.A.; Norton, R.S.; Borowitzka, L.J. Organic osmoregulatory solutes in cyanobacteria. J. Gen. Microbiol. 1984, 130, 2177–2191. [Google Scholar]
- Singh, A.K.; Chakarvathy, D.; Singh, T.P.K.; Singh, H.N. Evidence for a role for l-proline as a salinity protectant in the cyanobacterium Nostoc muscorum. Plant Cell Environ. 1996, 19, 490–494. [Google Scholar] [CrossRef]
- Fulda, S.; Huckauf, J.; Schoor, A.; Hagemann, M. Analysis of stress responses in the cyanobacterial strains Synechococcus sp. PCC 7942, Synechocystis sp. PCC 6803, and Synechococcus sp. PCC 7418: Osmolyte accumulation and stress proteins synthesis. J. Plant Physiol. 1999, 154, 240–249. [Google Scholar] [CrossRef]
- Klähn, S.; Steglich, C.; Hess, W.R.; Hagemann, M. Glucosylglycerate: A secondary compatible solute common to marine cyanobacteria from nitrogen-poor environments. Environ. Microbiol. 2010, 12, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Shih, P.M.; Wu, D.; Latifi, A.; Axen, S.D.; Fewer, D.P.; Talla, E.; Calteau, A.; Cai, F.; Tandeau de Marsac, N.; Rippka, R.; et al. Improving the coverage of the cyanobacterial phylum using diversity-driven genome sequencing. Proc. Natl. Acad. Sci. USA 2013, 110, 1053–1058. [Google Scholar] [CrossRef]
- Hewson, I.; Poretsky, R.S.; Beinart, R.A.; White, A.E.; Shi, T.; Bench, S.R.; Moisander, P.H.; Paerl, R.W.; Tripp, H.J.; Montoya, J.P.; Moran, M.A.; Zehr, J.P. In situ transcriptomic analysis of the globally important keystone N2-fixing taxon Crocosphaera watsonii. ISME J 2009, 3, 618–631. [Google Scholar] [CrossRef]
- Pade, N.; Compaore, J.; Klähn, S.; Stal, L.J.; Hagemann, M. The marine cyanobacterium Crocosphaera watsonii WH8501 synthesizes the compatible solute trehalose by a laterally acquired OtsAB fusion protein. Environ. Microbiol. 2012, 14, 1261–1271. [Google Scholar] [CrossRef]
- Bergman, B.; Sandh, G.; Lin, S.; Larsson, J.; Carpenter, E.J. Trichodesmium—a widespread marine cyanobacterium with unusual nitrogen fixation properties. FEMS Microbiol. Rev. 2013, 37, 286–302. [Google Scholar] [CrossRef] [PubMed]
- Salerno, G.L.; Porchia, A.C.; Vargas, W.A.; Abdian, P.L. Fructose-containing oligosaccharides: Novel compatible solutes in Anabaena cells exposed to salt stress. Plant Sci. 2004, 167, 1003–1008. [Google Scholar] [CrossRef]
- Fischer, D.; Geyer, A.; Loos, E. Occurrence of glucosylsucrose [α-d-glucopyranosyl-(1→2)-α-d-glucopyranosyl-(1→2)-β-d-fructofuranoside] and glucosylated homologues in cyanobacteria: Structural properties, cellular contents and possible function as thermoprotectants. FEBS J. 2006, 273, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Goh, F.; Barrow, K.D.; Burns, B.P.; Neilan, B.A. Identification and regulation of novel compatible solutes from hypersaline stromatolite-associated cyanobacteria. Arch. Microbiol. 2010, 192, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Borges, N.; Ramos, A.; Raven, N.D.H.; Sharp, R.J.; Santos, H. Comparative study of the thermostabilizing properties of mannosylglycerate and other compatible solutes on model enzymes. Extremophiles 2002, 6, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Hincha, D.K.; Hagemann, M. Stabilization of model membranes during drying by compatible solutes involved in the stress tolerance of plants and microorganisms. Biochem. J. 2004, 383, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Kandror, O.; DeLeon, A.; Goldberg, A.L. Trehalose synthesis is induced upon exposure of Escherichia coli to cold and is essential for viability at low temperatures. Proc. Natl. Acad. Sci. USA 2002, 99, 9727–9732. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.C.; Oh, S.J.; Seo, J.S.; Choi, W.B.; Song, S.I.; Kim, C.H.; Kim, Y.S.; Seo, H.S.; Choi, Y.D.; Nahm, B.H.; et al. Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth. Plant Physiol. 2003, 131, 516–524. [Google Scholar] [CrossRef]
- Diamant, S.; Rosenthal, D.; Azem, A.; Eliahu, N.; Ben-Zvi, A.P.; Goloubinoff, P. Dicarboxylic amino acids and glycine-betaine regulate chaperone-mediated protein-disaggregation under stress. Mol. Microbiol. 2003, 49, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Sawangwan, T.; Goedl, C.; Nidetzky, B. Glucosylglycerol and glucosylglycerate as enzyme stabilizers. Biotechnol. J. 2010, 5, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Luley-Goedel, C.; Nidetzky, B. Glycosides as compatible solutes: biosynthesis and applications. Nat. Prod. Rep. 2011, 28, 875–896. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, M.; Erdmann, N. Activation and pathway of glucosylglycerol biosynthesis in the cyanobacterium Synechocystis sp. PCC 6803. Microbiology 1994, 140, 1427–1431. [Google Scholar] [CrossRef]
- Hagemann, M.; Schoor, A.; Jeanjean, R.; Zuther, E.; Joset, F. The stpA gene form Synechocystis sp. strain PCC 6803 encodes the glucosylglycerol-phosphate phosphatase involved in cyanobacterial osmotic response to salt shock. J. Bacteriol. 1997, 179, 1727–1733. [Google Scholar] [PubMed]
- Marin, K.; Zuther, E.; Kerstan, T.; Kunert, A.; Hagemann, M. The ggpS gene from Synechocystis sp. strain PCC 6803 encoding glucosylglycerol-phosphate synthase is involved in osmolyte synthesis. J. Bacteriol. 1998, 180, 4843–4849. [Google Scholar]
- Porchia, A.C.; Salerno, G.L. Sucrose biosynthesis in a prokaryotic organism: Presence of two sucrose-phosphate synthases in Anabaena with remarkable differences compared with the plant enzymes. Proc. Natl. Acad. Sci. USA 1996, 93, 13600–13604. [Google Scholar] [CrossRef] [PubMed]
- Higo, A.; Katoh, H.; Ohmori, K.; Ikeuchi, M.; Ohmori, M. The role of a gene cluster for trehalose metabolism in dehydration tolerance of the filamentous cyanobacterium Anabaena sp. PCC 7120. Microbiology 2006, 152, 979–987. [Google Scholar] [CrossRef]
- Waditee, R.; Tanaka, Y.; Aoki, K.; Hibino, T.; Jikuya, H.; Takabe, T.; Takabe, T. Isolation and functional characterization of N-methyltransferase that catalyze betaine synthesis from glycine in a halotolerant photosynthetic organism Aphanothece halophytica. J. Biol. Chem. 2003, 278, 4932–4942. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Noël, G.M.; Cumino, A.C.; Kolman, M.A.; Salerno, G.L. First evidence of sucrose biosynthesis by single cyanobacterial bimodular proteins. FEBS Lett. 2013, 587, 1669–1674. [Google Scholar] [CrossRef]
- Kolman, M.A.; Torres, L.L.; Martin, M.L.; Salerno, G.L. Sucrose synthase in unicellular cyanobacteria and its relationship with salt and hypoxic stress. Planta 2011, 235, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Azua-Bustos, A.; Zúñiga, J.; Arenas-Fajardo, C.; Orellana, M.; Salas, L.; Rafael, V. Gloeocapsopsis AAB1, an extremely desiccation-tolerant cyanobacterium isolated from the Atacama Desert. Extremophiles 2014, 18, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Ikeda, Y.; Sakata, T.; Maeda, H. Cloning and analysis of the ggpS gene from Cyanobacteria Arthrospira spp. involved in the synthesis of an osmolyte glucosylglycerol. Biocontrol. Sci. 2011, 16, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Kothari, A.; Vaughn, M.; Garcia-Pichel, F. Comparative genomic analyses of the cyanobacterium, Lyngbya aestuarii BL J, a powerful hydrogen producer. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef]
- Mao, X.; Olman, V.; Stuart, R.; Paulsen, I.T.; Palenik, B.; Xu, Y. Computational prediction of the osmoregulation network in Synechococcus sp. WH8102. BMC Genomics 2010, 11. [Google Scholar] [CrossRef]
- Csonka, L.N.; Hanson, A.D. Prokaryotic osmoregulation: genetics and physiology. Annu. Rev. Microbiol. 1991, 45, 569–606. [Google Scholar] [CrossRef]
- Ziegler, C.; Bremer, E.; Krämer, R. The BCCT family of carriers: from physiology to crystal structure. Mol. Microbiol. 2010, 78, 13–34. [Google Scholar] [PubMed]
- Pittelkow, M.; Bremer, E. Cellular adjustment of Bacillus subtilis and other bacilli to fluctuating salinities. In Halophiles and Hypersaline Environments: Current Research and Future Trends; Ventosa, A., Ohren, A., Ma, Y., Eds.; Springer: Berlin, Germany, 2011; pp. 275–302. [Google Scholar]
- Lucht, J.M.; Bremer, E. Adaptation of Escherichia coli to high osmolarity environments: Osmoregulation of the high-affinity glycine betaine transport system ProU. FEMS Microbiol. Rev. 1995, 14, 3–20. [Google Scholar] [CrossRef]
- Moore, D.J.; Reed, R.H.; Stewart, W.D.P. A glycine betaine transport system in Aphanothece halophytica and other glycine betaine-synthesizing cyanobacteria. Arch. Microbiol. 1987, 147, 399–405. [Google Scholar] [CrossRef]
- Laloknam, S.; Tanaka, K.; Buaboocha, T.; Waditee, R.; Incharoensakdi, A.; Hibino, T.; Tanaka, Y.; Takabe, T. Halotolerant cyanobacterium Aphanothece halophytica contains a betaine transporter active at alkaline pH and high salinity. Appl. Environ. Microbiol. 2006, 72, 6018–6026. [Google Scholar] [CrossRef] [PubMed]
- Mikkat, S.; Hagemann, M.; Schoor, A. Active transport of glucosylglycerol is involved in salt adaptation of the cyanobacterium Synechocystis sp. PCC 6803. Arch. Microbiol. 1996, 142, 1725–1732. [Google Scholar] [CrossRef]
- Mikkat, S.; Effmert, U.; Hagemann, M. Uptake and use of the osmoprotective compounds trehalose, glucosylglycerol, and sucrose by the cyanobacterium Synechocystis sp. PCC6803. Arch. Microbiol. 1997, 167, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, M.; Richter, S.; Mikkat, S. The ggtA gene encodes a subunit of the transport system for the osmoprotective compound glucosylglycerol in Synechocystis sp. strain PCC 6803. J. Bacteriol. 1997, 179, 714–720. [Google Scholar] [PubMed]
- Mikkat, S.; Hagemann, M. Molecular analysis of the ggtBCD gene cluster of Synechocystis sp. Strain PCC6803 encoding subunits of an ABC transporter for osmoprotective compounds. Arch. Microbiol 2000, 174, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Kopf, M.; Klähn, S.; Pade, N.; Weingärtner, C.; Hagemann, M.; Voß, B.; Hess, W.R. Comparative genome analysis of the closely related Synechocystis strains PCC 6714 and PCC 6803. DNA Res. 2014, 21, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Ferjani, A.; Mustardy, L.; Sulpice, R.; Marin, K.; Suzuki, I.; Hagemann, M.; Murata, N. Glucosylglycerol, a compatible solute, sustains cell division under salt stress. Plant Physiol. 2003, 131, 1628–1637. [Google Scholar] [CrossRef] [PubMed]
- Horn, C.; Jenewein, S.; Sohn-Bösser, L.; Bremer, E.; Schmitt, L. Biochemical and structural analysis of the Bacillus subtilis ABC transporter OpuA and its isolated subunits. J. Mol. Microbiol. Biotechnol. 2005, 10, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Ducat, D.C.; Avelar-Rivas, J.A.; Way, J.C.; Silver, P.A. Rerouting carbon flux to enhance photosynthetic productivity. Appl. Environ. Microbiol. 2012, 78, 2660–2668. [Google Scholar] [CrossRef] [PubMed]
- Roder, A.; Hoffmann, E.; Hagemann, M.; Berg, G. Synthesis of the compatible solutes glucosylglycerol and trehalose by salt-stressed cells of Stenotrophomonas strains. FEMS Microbiol. Lett. 2005, 243, 219–226. [Google Scholar] [CrossRef]
- Alavi, P.; Starcher, M.R.; Zachow, C.; Müller, H.; Berg, G. Root-microbe systems: The effect and mode of interaction of stress protecting agent (SPA) Stenotrophomonas rhizophila DSM14405T. Front. Plant Sci. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Reed, R.H.; Warr, S.R.C.; Richardson, D.L.; Moore, D.J.; Stewart, W.D.P. Multiphasic osmotic adjustment in a euryhaline cyanobacterium. FEMS Microbiol. Lett. 1985, 28, 225–229. [Google Scholar] [CrossRef]
- Marin, K.; Kanesaki, Y.; Los, D.A.; Murata, N.; Suzuki, I.; Hagemann, M. Gene expression profiling reflects physiological processes in salt acclimation of Synechocystis sp. Strain PCC 6803. Plant Physiol. 2004, 136, 3290–3300. [Google Scholar] [CrossRef]
- Blumwald, E.; Mehlhorn, R.J.; Packer, L. Studies of osmoregulation in salt adaptation of cyanobacteria with ESR spin-probe techniques. Proc. Natl. Acad. Sci. USA 1983, 80, 2599–2602. [Google Scholar] [CrossRef] [PubMed]
- Shapiguzov, A.; Lyukevich, A.A.; Allakhverdiev, S.I.; Sergeyenko, T.V.; Suzuki, I.; Murata, N.; Los, D.A. Osmotic shrinkage of cells of Synechocystis sp. PCC 6803 by water efflux via aquaporins regulates osmostress-inducible gene expression. Microbiology 2005, 151, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, N.; Kobayashi, H.; Katoh, H.; Ogawa, T.; Futatsugi, L.; Nakamura, T.; Bakker, E.P.; Uozumi, N. Na+-dependent K+ uptake Ktr system from the cyanobacterium Synechocystis sp. PCC 6803 and its role in the early phases of cell adaptation to hyperosmotic shock. J. Biol. Chem. 2004, 279, 54952–54962. [Google Scholar] [CrossRef] [PubMed]
- Nanatani, K.; Shijuku, T.; Takano, Y.; Zulkifli, L.; Yamazaki, T.; Tominaga, A.; Souma, S.; Onai, K.; Morishita, M.; Ishiura, M.; et al. Comparative analysis of kdp and ktr mutants reveals distinct doles of the potassium transporters in the model cyanobacterium Synechocystis sp. PCC 6803. J. Bacteriol. 2014. [Google Scholar] [CrossRef]
- Schubert, H.; Fulda, S.; Hagemann, M. Effects of adaptation to different salt concentrations on photosynthesis and pigmentation of the cyanobacterium Synechocystis sp. PCC 6803. J. Plant Physiol. 1993, 142, 291–295. [Google Scholar] [CrossRef]
- Hagemann, M.; Fulda, S.; Schubert, H. DNA, RNA and protein synthesis in the cyanobacterium Synechocystis sp. PCC 6803 adapted to different salt concentrations. Curr. Microbiol. 1994, 28, 201–207. [Google Scholar] [CrossRef]
- Elanskaya, I.V.; Karandashova, I.V.; Bogachev, A.V.; Hagemann, M. Functional analysis of the Na+/H+ antiporter encoding genes of the cyanobacterium Synechocystis PCC 6803. Biochemistry (Moscow) 2002, 67, 432–440. [Google Scholar] [CrossRef]
- Wang, H.L.; Postier, B.L.; Burnap, R.L. Polymerase chain reaction-based mutageneses identify key transporters belonging to multigene families involved in Na+ and pH homeostasis of Synechocystis sp. PCC 6803. Mol. Microbiol. 2002, 44, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Tsunekawa, K.; Shijuku, T.; Hayashimoto, M.; Kojima, Y.; Onai, K.; Morishita, M.; Ishiura, M.; Kuroda, T.; Nakamura, T.; Kobayashi, H.; et al. Identification and characterization of the Na+/H+ antiporter Nhas3 from the thylakoid membrane of Synechocystis sp. PCC 6803. J. Biol. Chem. 2009, 284, 16513–16521. [Google Scholar] [CrossRef]
- Blanco-Rivero, A.; Leganés, F.; Fernández-Valiente, E.; Calle, P.; Fernández-Piñas, F. mrpA, a gene with roles in resistance to Na+ and adaptation to alkaline pH in the cyanobacterium Anabaena sp. PCC7120. Microbiology 2005, 151, 1671–1682. [Google Scholar] [CrossRef] [PubMed]
- Fukaya, F.; Promden, W.; Hibino, T.; Tanaka, Y.; Nakamura, T.; Takabe, T. An Mrp-like cluster in the halotolerant cyanobacterium Aphanothece halophytica functions as a Na+/H+ antiporter. Appl. Environ. Microbiol. 2009, 75, 6626–6629. [Google Scholar] [CrossRef]
- Soontharapirakkul, K.; Promden, W.; Yamada, N.; Kageyama, H.; Incharoensakdi, A.; Iwamoto-Kihara, A.; Takabe, T. Halotolerant cyanobacterium Aphanothece halophytica contains an Na+-dependent F1F0-ATP synthase with a potential role in salt-stress tolerance. J. Biol. Chem. 2011, 286, 10169–10176. [Google Scholar] [CrossRef] [PubMed]
- Jayaram, H.; Robertson, J.L.; Wu, F.; Williams, C.; Miller, C. Structure of a slow CLC Cl−/H+ antiporter from a cyanobacterium. Biochemistry 2011, 50, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.; Esper, B.; Karandashova, I.; Teuber, M.; Elanskaya, I.; Rögner, M.; Hagemann, M. Potassium uptake in the unicellular cyanobacterium Synechocystis sp. strain PCC 6803 mainly depends on a Ktr-like system encoded by slr1509 (ntpJ). FEBS Lett. 2003, 548, 53–58. [Google Scholar] [CrossRef]
- Ballal, A.; Basu, B.; Apte, S.K. The Kdp-ATPase system and its regulation. J. Biosci. 2007, 32, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Katoh, H.; Asthana, R.K.; Ohmori, M. Gene expression in the cyanobacterium Anabaena sp. PCC7120 under desiccation. Microb. Ecol. 2004, 47, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Kanesaki, Y.; Suzuki, I.; Allakhverdiev, S.I.; Mikami, K.; Murata, N. Salt stress and hyperosmotic stress regulate the expression of different sets of genes in Synechocystis sp. PCC 6803. Biochem. Biophys. Res. Commun. 2002, 290, 339–348. [Google Scholar] [CrossRef]
- Marin, K.; Suzuki, I.; Yamaguchi, K.; Ribbeck, K.; Yamamoto, H.; Kanesaki, Y.; Hagemann, M.; Murata, N. Identification of histidine kinases that act as sensors in the perception of salt stress in Synechocystis sp. PCC 6803. Proc. Natl. Acad. Sci. USA 2003, 100, 9061–9066. [Google Scholar] [CrossRef] [PubMed]
- Mitschke, J.; Georg, J.; Scholz, I.; Sharma, C.M.; Dienst, D.; Bantscheff, J.; Voss, B.; Steglich, C.; Wilde, A.; Vogel, J.; Hess, W.R. An experimentally anchored map of transcriptional start sites in the model cyanobacterium Synechocystis sp. PCC6803. Proc. Natl Acad Sci USA 2011, 108, 2124–2129. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.; Bryant, D.A. Synechococcus sp. strain PCC 7002 transcriptome: acclimation to temperature, salinity, oxidative stress, and mixotrophic growth conditions. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef]
- Schubert, H.; Hagemann, M. Salt effect on 77 K fluorescence and photosynthesis in the cyanobacterium Synechocystis spec. PCC 6803. FEMS Microbiol. Lett. 1990, 71, 169–172. [Google Scholar] [CrossRef]
- Qiao, J.; Huang, S.; Te, R.; Wang, J.; Chen, L.; Zhang, W. Integrated proteomic and transcriptomic analysis reveals novel genes and regulatory mechanisms involved in salt stress responses in Synechocystis sp. PCC 6803. Appl. Microbiol. Biotechnol. 2013, 97, 8253–8264. [Google Scholar] [CrossRef] [PubMed]
- Billis, K.; Billini, M.; Tripp, H.J.; Kyrpides, N.C.; Mavromatis, K. Comparative transcriptomics between Synechococcus PCC 7942 and Synechocystis PCC 6803 provide insights into mechanisms of stress acclimation. PLoS One 2014, 9, e109738. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Huang, F.; Nilsson, F.; Hagemann, M.; Norling, B. Proteomics of Synechocystis sp. strain PCC 6803 identification of periplasmic proteins in cells grown at low and high salt concentrations. Eur. J. Biochem. 2000, 267, 5900–5907. [Google Scholar] [CrossRef]
- Huang, F.; Fulda, S.; Hagemann, M.; Norling, B. Proteomic screening of salt-stress-induced changes in plasma membranes of Synechocystis sp. strain PCC 6803. Proteomics 2006, 6, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Mikkat, S.; Huang, F.; Huckauf, J.; Marin, K.; Norling, B.; Hagemann, M. Proteome analysis of salt stress response in the cyanobacterium Synechocystis sp. strain PCC 6803. Proteomics 2006, 6, 2733–2745. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, Y.; Chen, W.; Ding, L.; Li, P.; Zhao, X.; Wang, X.; Li, A.; Bao, Q. Identification of differentially expressed proteins of Arthrospira (Spirulina) platensis-YZ under salt-stress conditions by proteomics and qRT-PCR analysis. Proteome Sci. 2013, 11. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Shao, M.; Chen, L.; Wang, J.; Wu, G.; Tian, X.; Liu, J.; Huang, S.; Zhang, W. Systematic characterization of hypothetical proteins in Synechocystis sp. PCC 6803 reveals proteins functionally relevant to stress responses. Gene 2013, 512, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Rai, S.; Agrawal, C.; Shrivastava, A.K.; Singh, P.K.; Rai, L.C. Comparative proteomics unveils cross species variations in Anabaena under salt stress. J. Proteomics 2014, 98, 254–270. [Google Scholar] [CrossRef] [PubMed]
- Rai, S.; Singh, S.; Shrivastava, A.K.; Rai, L.C. Salt and UV-B induced changes in Anabaena PCC 7120: Physiological, proteomic and bioinformatic perspectives. Photosynth. Res. 2013, 118, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Poolman, B.; Spitzer, J.J.; Wood, J.M. Bacterial osmosensing: roles of membrane structure and electrostatics in lipid-protein and protein-protein interactions. Biochim. Biophys. Acta 2004, 1666, 88–104. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.F.; Stirnberg, M.; Roenneke, B.; Marin, K. A novel mechanism of osmosensing, a salt-dependent protein-nucleic acid interaction in the cyanobacterium Synechocystis species PCC 6803. J. Biol Chem 2011, 286, 3235–3241. [Google Scholar] [CrossRef]
- Marin, K.; Huckauf, J.; Fulda, S.; Hagemann, M. Salt-dependent expression of glucosylglycerol-phosphate synthase, involved in osmolyte synthesis in the cyanobacterium Synechocystis sp. strain PCC 6803. J. Bacteriol. 2002, 184, 2870–2877. [Google Scholar] [CrossRef] [PubMed]
- Klähn, S.; Höhne, A.; Simon, E.; Hagemann, M. The gene ssl3076 encodes a protein mediating the salt-induced expression of ggpS for the biosynthesis of the compatible solute glucosylglycerol in Synechocystis sp. strain PCC 6803. J. Bacteriol. 2010, 192, 4403–4412. [Google Scholar] [CrossRef] [PubMed]
- Gralla, J.D.; Vargas, D.R. Potassium glutamate as a transcriptional inhibitor during bacterial osmoregulation. EMBO J. 2006, 25, 1515–1521. [Google Scholar] [PubMed]
- Nikkinen, H.L.; Hakkila, K.; Gunnelius, L.; Huokko, T.; Pollari, M.; Tyystjärvi, T. The SigB σ factor regulates multiple salt acclimation responses of the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2012, 158, 514–523. [Google Scholar] [CrossRef] [PubMed]
- Pollari, M.; Gunnelius, L.; Tuominen, I.; Ruotsalainen, V.; Tyystjärvi, E.; Salminen, T.; Tyystjärvi, T. Characterization of single and double inactivation strains reveals new physiological roles for group 2 sigma factors in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2008, 147, 1994–2005. [Google Scholar] [CrossRef] [PubMed]
- Tyystjärvi, T.; Huokko, T.; Rantamäki, S.; Tyystjärvi, E. Impact of different group 2 sigma factors on light use efficiency and high salt stress in the cyanobacterium Synechocystis sp. PCC 6803. PLoS One 2013, 8, e63020. [Google Scholar] [CrossRef] [PubMed]
- Shoumskaya, M.A.; Paithoonrangsarid, K.; Kanesaki, Y.; Los, D.A.; Zinchenko, V.V.; Tanticharoen, M.; Suzuki, I.; Murata, N. Identical Hik-Rre systems are involved in perception and transduction of salt signals and hyperosmotic signals but regulate the expression of individual genes to different extents in Synechocystis. J. Biol. Chem. 2005, 280, 21531–21538. [Google Scholar]
- Li, T.; Yang, H.M.; Cui, S.X.; Suzuki, I.; Zhang, L.F.; Li, L.; Bo, T.T.; Wang, J.; Murata, N.; Huang, F. Proteomic study of the impact of Hik33 mutation in Synechocystis sp. PCC 6803 under normal and salt stress conditions. J. Proteome Res. 2012, 11, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.H.; Black, T.A.; Jäger, K.; Panoff, J.M.; Wolk, C.P. Regulation of an osmoticum-responsive gene in Anabaena sp. strain PCC 7120. J. Bacteriol. 1998, 180, 6332–6337. [Google Scholar]
- Ehira, S.; Kimura, S.; Miyazaki, S.; Ohmori, M. Sucrose synthesis in the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120 is controlled by the two-component response regulator OrrA. Appl. Environ. Microbiol. 2014, 80, 5672–5679. [Google Scholar] [CrossRef]
- Chen, L.; Wu, L.; Zhu, Y.; Song, Z.; Wang, J.; Zhang, W. An orphan two-component response regulator Slr1588 involves salt tolerance by directly regulating synthesis of compatible solutes in photosynthetic Synechocystis sp. PCC 6803. Mol. Biosyst. 2014, 10, 1765–1774. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Zhang, X.; Chi, X.; Guan, X.; Li, Y.; Qin, S.; Shao, H.B. Serine/threonine protein kinase SpkG is a candidate for high salt resistance in the unicellular cyanobacterium Synechocystis sp. PCC 6803. PLoS One 2011, 6, e18718. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.K.; Latifi, A.; Chen, W.L.; Zhang, C.C. The inositol monophosphatase All2917 (IMPA1) is involved in osmotic adaptation in Anabaena sp. PCC7120. Environ. Microbiol. Rep. 2012, 4, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Algenol Biofuels. Available online: http://www.algenol.com/ (accessed on 23 December 2014).
- Chisti, Y. Constraints to commercialization of algal fuels. J. Biotechnol. 2013, 167, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Guschina, I.A.; Harwood, J.L. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res. 2006, 45, 160–186. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.K.; Dhar, D.W. Microalgae as second generation biofuel. A review. Agron. Sustain. Dev. 2011, 31, 605–629. [Google Scholar] [CrossRef]
- Bernstein, H.C.; Konopka, A.; Melnicki, M.R.; Hill, E.A.; Kucek, L.A.; Zhang, S.; Shen, G.; Bryant, D.A.; Beliaev, A.S. Effect of mono- and dichromatic light quality on growth rates and photosynthetic performance of Synechococcus sp. PCC 7002. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, D.; Momot, D.; Brasg, I.A.; Ananyev, G.; Lenz, O.; Bryant, D.A.; Dismukes, G.C. Boosting autofermentation rates and product yields with sodium stress cycling: application to production of renewable fuels by cyanobacteria. Appl. Environ. Microbiol. 2010, 76, 6455–6462. [Google Scholar] [CrossRef] [PubMed]
- Guerra, L.T.; Xu, Y.; Bennette, N.; McNeely, K.; Bryant, D.A.; Dismukes, G.C. Natural osmolytes are much less effective substrates than glycogen for catabolic energy production in the marine cyanobacterium Synechococcus sp. strain PCC 7002. J. Biotechnol. 2013, 166, 65–75. [Google Scholar] [CrossRef] [PubMed]
- bitop AG Extremolytes. Available online: http://www.bitop.de/cms/website.php?id=/en/index/extremolyte.htm (accessed on 23 December 2014).
- Kovacevic, V.; Wesseler, J. Cost-effectiveness analysis of algae energy production in the EU. Energy Policy 2010, 38, 5749–5757. [Google Scholar] [CrossRef]
- Du, W.; Liang, F.; Duan, Y.; Tan, X.; Lu, X. Exploring the photosynthetic production capacity of sucrose by cyanobacteria. Metab. Eng. 2013, 19, 17–25. [Google Scholar] [PubMed]
- Reed, R.H.; Warr, S.R.C.; Kerby, N.W.; Stewart, W.D.P. Osmotic shock-induced release of low molecular weight metabolites from free-living and immobilized cyanobacteria. Enzyme Microb. Technol. 1986, 8, 101–104. [Google Scholar]
- Fulda, S.; Hagemann, M.; Libbert, E. Release of glucosylglycerol from the cyanobacterium Synechocystis spec. SAG 92.79 by hypoosmotic shock. Arch. Microbiol. 1990, 153, 405–408. [Google Scholar] [CrossRef]
- Erdmann, N.; Zuther, E.; Abarzua, S. Comparative studies on the photoproduction of nonhydrogenous resources by cyanobacteria. Curr. Microbiol. 1992, 25, 83–88. [Google Scholar] [CrossRef]
- Takagi, M.; Karseno; Yoshida, T. Effect of salt concentration on intracellular accumulation of lipids and triacylglyceride in marine microalgae Dunaliella cells. J. Biosci. Bioeng. 2006, 101, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; Khozin-Goldberg, I.; Cohen, Z.; Boussiba, S. The effect of light, salinity, and nitrogen availability on lipid production by Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2011, 90, 1429–1441. [Google Scholar] [CrossRef] [PubMed]
- Waditee, R.; Bhuiyan, M.N.; Rai, V.; Aoki, K.; Tanaka, Y.; Hibino, T.; Suzuki, S.; Takano, J.; Jagendorf, A.T.; Takabe, T.; Takabe, T. Genes for direct methylation of glycine provide high levels of glycinebetaine and abiotic-stress tolerance in Synechococcus and Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 1318–1323. [Google Scholar] [CrossRef] [PubMed]
- Klähn, S.; Marquardt, D.M.; Rollwitz, I.; Hagemann, M. Expression of the ggpPS gene for glucosylglycerol biosynthesis from Azotobacter vinelandii improves the salt tolerance of Arabidopsis thaliana. J. Exp. Bot. 2009, 60, 1679–1689. [Google Scholar] [CrossRef]
- Sievers, N.; Muders, K.; Henneberg, M.; Klähn, S.; Effmert, M.; Junghans, H.; Hagemann, M. Establishing glucosylglycerol synthesis in potato (Solanum tuberosum L. cv. Albatros) by expression of the ggpPS gene from Azotobacter vinelandii. J. Plant Sci. Mol. Breed. 2013. [Google Scholar] [CrossRef]
- Kaczmarzyk, D.; Anfelt, J.; Särnegrim, A.; Hudson, E.P. Overexpression of sigma factor SigB improves temperature and butanol tolerance of Synechocystis sp. PCC6803. J. Biotechnol. 2014, 182–183, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Miyao, K.; Tamoi, M.; Kanaboshi, H.; Miyasaka, H.; Shigeoka, S. Molecular characterization of glutathione peroxidase-like protein in halotolerant Chlamydomonas sp. W80. Physiol. Plant. 2003, 117, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, K.; Miyao, K.; Gaber, A.; Takeda, T.; Kanaboshi, H.; Miyasaka, H.; Shigeoka, S. Enhancement of stress tolerance in transgenic tobacco plants overexpressing Chlamydomonas glutathione peroxidase in chloroplasts or cytosol. Plant J. 2004, 37, 21–33. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pade, N.; Hagemann, M. Salt Acclimation of Cyanobacteria and Their Application in Biotechnology. Life 2015, 5, 25-49. https://doi.org/10.3390/life5010025
Pade N, Hagemann M. Salt Acclimation of Cyanobacteria and Their Application in Biotechnology. Life. 2015; 5(1):25-49. https://doi.org/10.3390/life5010025
Chicago/Turabian StylePade, Nadin, and Martin Hagemann. 2015. "Salt Acclimation of Cyanobacteria and Their Application in Biotechnology" Life 5, no. 1: 25-49. https://doi.org/10.3390/life5010025
APA StylePade, N., & Hagemann, M. (2015). Salt Acclimation of Cyanobacteria and Their Application in Biotechnology. Life, 5(1), 25-49. https://doi.org/10.3390/life5010025