
Long-Term Changes in Cyanobacteria Populations in Lake Kinneret (Sea of Galilee), Israel: An Eco-Physiological Outlook


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Study Site
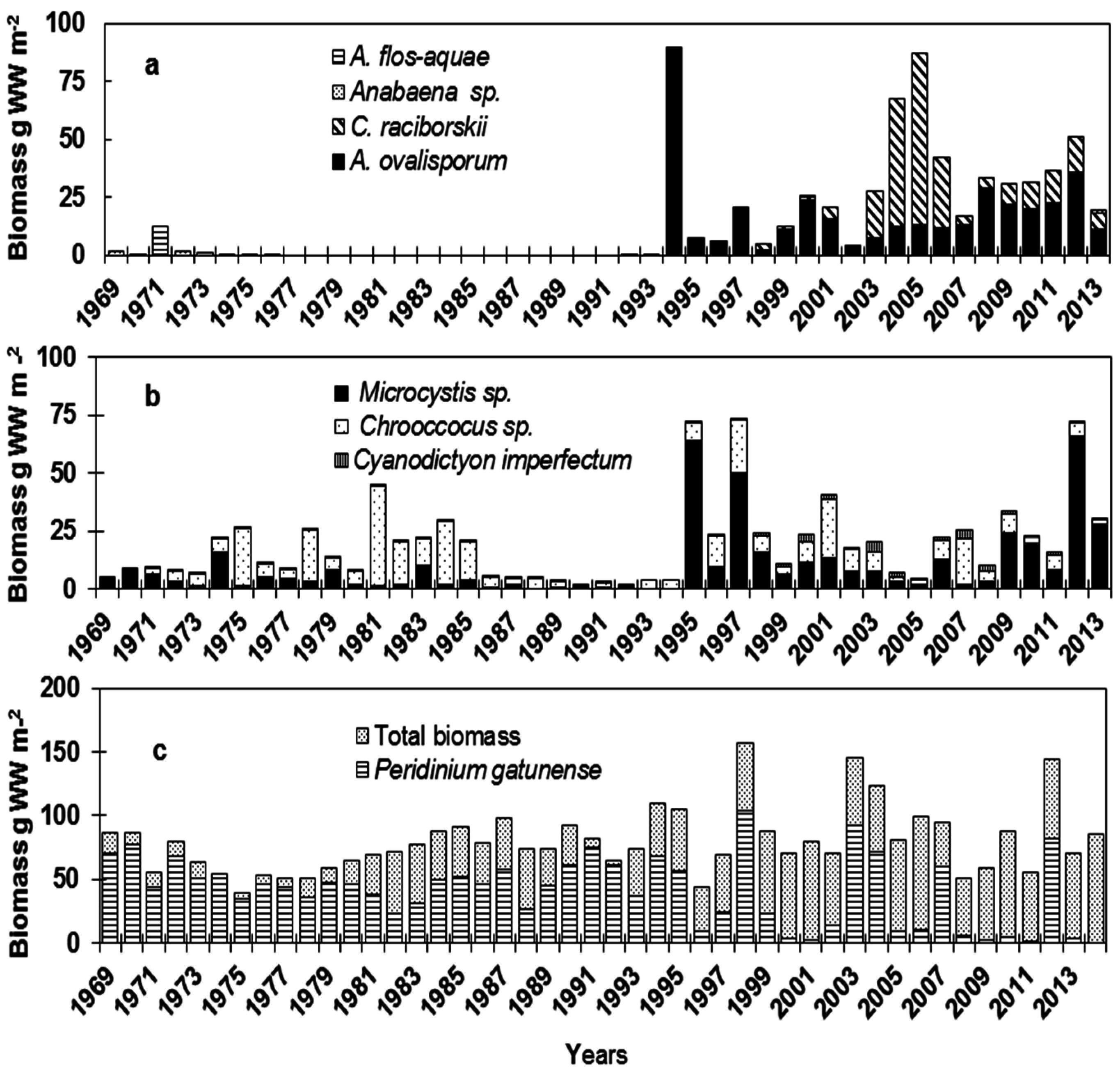
3. The Players

3.1. Nostocales
3.2. Chroococcales
3.3. Total and Peridinium gatunense Biomass
4. Long-Term Variations in Environmental Conditions
Temperature and Wind
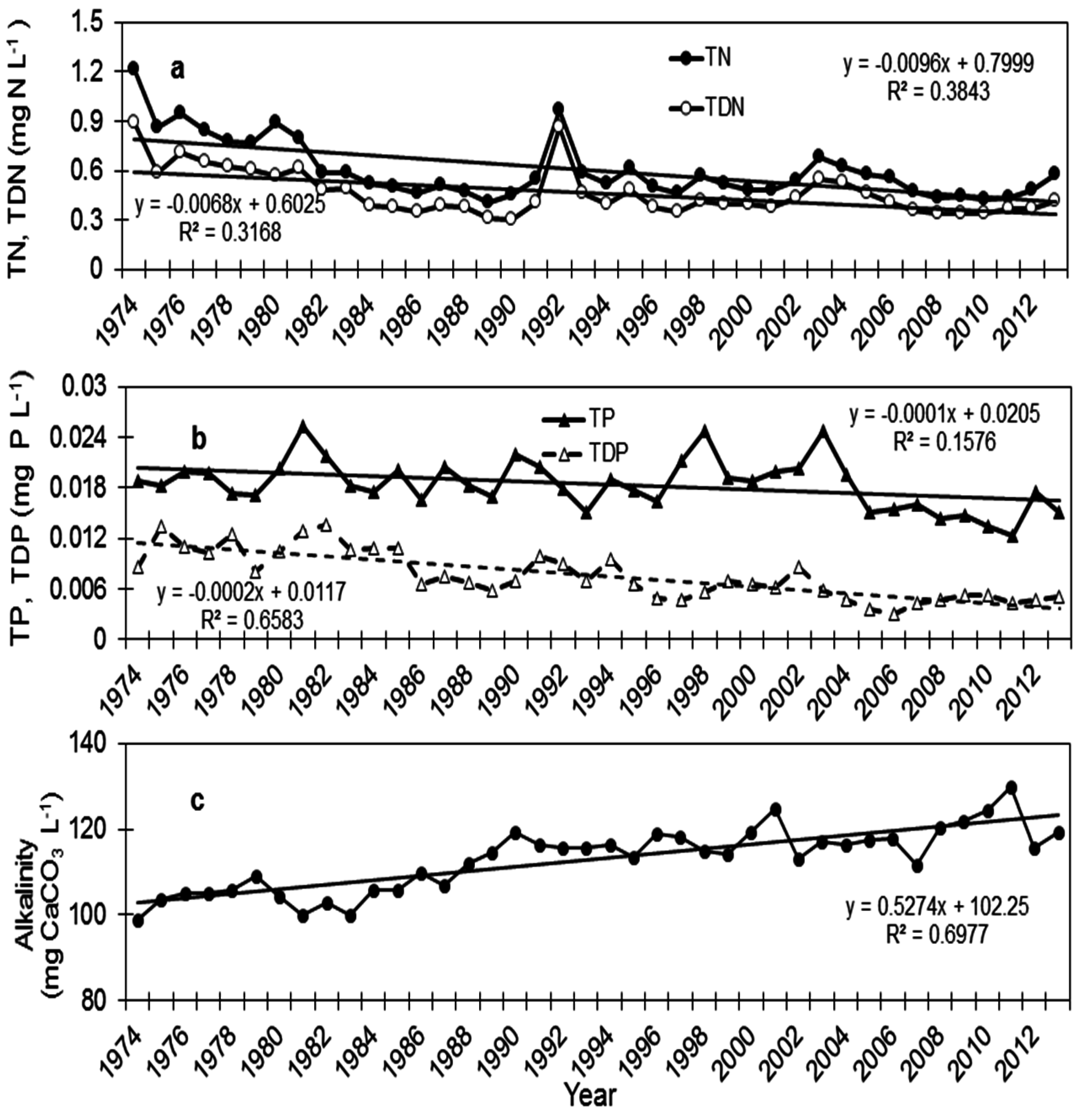
5. Variations in Nutrients Status
5.1. Total Nitrogen and Total Dissolved Nitrogen
5.2. Total Phosphorus and Total Dissolved Phosphorus

5.3. Salinity
5.4. Alkalinity, pH
6. Physiological Response of Cyanobacteria Populations to the Changing Environment
7. Biotic Interactions
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oliver, R.L.; Ganf, G.G. Freshwater blooms. In The ecology of Cyanobacteria: Their Diversity in Time and Space; Whitton, B.A., Potts, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 149–194. [Google Scholar]
- Huisman, J.; Matthijs, H.C.; Visser, P.M. Harmful Cyanobacteria; Springer: Dordrecht, The Netherlands, 2005; Volume 3. [Google Scholar]
- Paerl, H.W.; Gardner, W.S.; McCarthy, M.J.; Peierls, B.L.; Wilhelm, S.W. Algal blooms: Noteworthy nitrogen. Science 2014, 346, 175–175. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.F.; Hamilton, D.P.; Barnes, G.E. Recent occurrence of Cylindrospermopsis raciborskii in Waikato lakes of New Zealand. New Zeal J. Mar. Freshw. Res. 2003, 37, 829–836. [Google Scholar] [CrossRef]
- Wiedner, C.; Rücker, J.; Brüggemann, R.; Nixdorf, B. Climate change affects timing and size of populations of an invasive cyanobacterium in temperate regions. Oecologia 2007, 152, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Mehnert, G.; Leunert, F.; Cires, S.; Johnk, K.D.; Rucker, J.; Nixdorf, B.; Wiedner, C. Competitiveness of invasive and native cyanobacteria from temperate freshwaters under various light and temperature conditions. J. Plankton Res. 2010, 32, 1009–1021. [Google Scholar] [CrossRef]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, P.B.; Ley, L.M.; Dean, S.; Pick, F.R. The occurrence of the cyanobacterium Cylindrospermopsis raciborskii in Constance Lake: An exotic cyanoprokaryote new to Canada. Phycologia 2005, 44, 17–25. [Google Scholar] [CrossRef]
- Kosten, S.; Huszar, V.L.; Bécares, E.; Costa, L.S.; Donk, E.; Hansson, L.A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N. Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Change Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Sukenik, A.; Hadas, O.; Kaplan, A.; Quesada, A. Invasion of Nostocales (cyanobacteria) to subtropical and temperate freshwater lakes—physiological, regional, and global driving forces. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef]
- Padisák, J. Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, an expanding, highly adaptive cyanobacterium: Worldwide distribution and review of its ecology. Archiv Für Hydrobiologie Supplementband Monographische Beitrage 1997, 107, 563–593. [Google Scholar]
- Anderson, J.N.; Jeppesen, E.; Søndergaard, M. Ecological effects of reduced nutrient loading (oligotrophication) on lakes: An introduction. Freshw. Biol. 2005, 50, 1589–1593. [Google Scholar] [CrossRef]
- Moss, B.; Barker, T.; Stephen, D.; Williams, A.E.; Balayla, D.J.; Beklioglu, M.; Carvalho, L. Consequences of reduced nutrient loading on a lake system in a lowland catchment: Deviations from the norm? Freshw. Biol. 2005, 50, 1687–1705. [Google Scholar] [CrossRef]
- Dokulil, M.T.; Teubner, K. Do phytoplankton communities correctly track trophic changes? An assessment using directly measured and palaeolimnological data. Freshw. Biol. 2005, 50, 1594–1604. [Google Scholar] [CrossRef]
- Eckert, W.; Nishri, A. The Phosphorus Cycle. In Lake Kinneret: Ecology and Management; Zohary, T., Sukenik, A., Berman, T., Nishri, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 347–363. [Google Scholar]
- Berman, T.; Pollingher, U. Annual and seasonal variations of phytoplankton, chlorophyll and photosynthesis in Lake Kinneret. Limnol. Oceanogr. 1974, 19, 31–54. [Google Scholar] [CrossRef]
- Zohary, T. Changes to the phytoplankton assemblage of Lake Kinneret after decades of a predictable, repetitive pattern. Freshw. Biol. 2004, 49, 1355–1371. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Zohary, T.; Yacobi, Y.; Alster, A.; Fishbein, T.; Lippman, S.; Tibor, G. Phytoplankton. In Lake Kinneret: Ecology and Management; Zohary, T., Sukenik, A., Berman, T., Nishri, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 213–226. [Google Scholar]
- Alster, A.; Kaplan-Levy, R.; Sukenik, A.; Zohary, T. Morphology and phylogeny of a non-toxic invasive Cylindrospermopsis raciborskii from a Mediterranean Lake. Hydrobiologia 2010, 639, 115–128. [Google Scholar] [CrossRef]
- Hadas, O.; Pinkas, R.; Malinsky-Rushansky, N.; Nishri, A.; Kaplan, A.; Rimmer, A.; Sukenik, A. Appearance and establishment of diazotrophic cyanobacteria in Lake Kinneret, Israel. Freshw. Biol. 2012, 57, 1214–1227. [Google Scholar] [CrossRef]
- Pollingher, U.; Hadas, O.; Yacobi, Y.Z.; Zohary, T.; Berman, T. Aphanizomenon ovalisporum (Forti) in Lake Kinneret, Israel. J. Plankton Res. 1998, 20, 1321–1339. [Google Scholar] [CrossRef]
- Hadas, O.; Pinkas, R.; Delphine, E.; Vardi, A.; Kaplan, A.; Sukenik, A. Limnological and ecophysiological aspects of Aphanizomenon ovalisporum bloom in Lake Kinneret, Israel. J. Plankton Res. 1999, 21, 1439–1453. [Google Scholar] [CrossRef]
- Sukenik, A.; Hadas, O.; Kaplan, A. Cyanobacteria. In Lake Kinneret: Ecology and Management; Zohary, T., Sukenik, A., Berman, T., Nishri, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 213–226. [Google Scholar]
- Gkelis, S.; Moustaka-Gouni, M.; Sivonen, K.; Lanaras, T. First report of the cyanobacterium Aphanizomenon ovalisporum Forti in two Greek lakes and cyanotoxin occurrence. J. Plankton Res. 2005, 27, 1295–1300. [Google Scholar] [CrossRef]
- Messineo, V.; Melchiorre, S.; Di Corcia, A.; Gallo, P.; Bruno, M. Seasonal succession of Cylindrospermopsis raciborskii and Aphanizomenon ovalisporum blooms with cylindrospermopsin occurrence in the volcanic Lake Albano, Central Italy. Environ. Toxicol. 2010, 25, 18–27. [Google Scholar] [PubMed]
- Hickel, B.; Pollingher, U. Mass development of an iron precipitating cyanophyte (Cyanodictyon imperfectum) in a subtropical lake (Lake Kinneret, Israel). Phycologia 1988, 27, 291–294. [Google Scholar] [CrossRef]
- Serruya, C.; Edelstein, M.; Pollingher, U.; Serruya, S. Lake Kinneret sediments: Nutrient composition of the pore water and mud water exchanges. Limnol. Oceanogr. 1974, 19, 489–508. [Google Scholar] [CrossRef]
- Sukenik, A.; Eshkol, R.; Livne, A.; Hadas, O.; Rom, M.; Tchernov, D.; Vardi, A.; Kaplan, A. Inhibition of growth and photosynthesis of the dinoflagellate Peridinium gatunense by Microcystis sp.(cyanobacteria): A novel allelopathic mechanism. Limnol. Oceanogr. 2002, 47, 1656–1663. [Google Scholar] [CrossRef]
- Vardi, A.; Schatz, D.; Beeri, K.; Motro, U.; Sukenik, A.; Levine, A.; Kaplan, A. Dinoflagellate-cyanobacterium communication may determine the composition of phytoplankton assemblage in a mesotrophic lake. Curr. Biol. 2002, 12, 1767–1772. [Google Scholar] [CrossRef] [PubMed]
- Israel Oceanographic & Limnological Research. Available online: http://kinneret.ocean.org.il/nitur_grp.aspx (accessed on 2 February 2015).
- Rimmer, A.; Gal, G.; Opher, T.; Lechinsky, Y.; Yacobi, Y.Z. Mechanisms of long-term variations in the thermal structure of a warm lake. Limnol. Oceanogr. 2011, 56, 974–988. [Google Scholar] [CrossRef]
- Ostrovsky, I.; Rimmer, A.; Yacobi, Y.Z.; Nishri, A.; Sukenik, A.; Hadas, O.; Zohary, T. Long-term changes in the Lake Kinneret ecosystem: The effects of climate change and anthropogenic factors. In Climatic Change and Global Warming of Inland Waters: Impacts and Mitigation for Ecosystems and Societies; John Wiley & Sons: New York, NY, USA, 2013; pp. 271–293. [Google Scholar]
- Markel, D.; Shamir, U.; Green, P. Operational management of Lake Kinneret and its watershed. In Lake Kinneret: Ecology and Management; Zohary, T., Sukenik, A., Berman, T., Nishri, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 541–560. [Google Scholar]
- Hambright, K.; Zohary, T. Lakes Hula and Agmon: Destruction and creation of wetland ecosystems in northern Israel. Wetl. Ecol. Manag. 1998, 6, 83–89. [Google Scholar] [CrossRef]
- Saaroni, H.; Ziv, B.; Osetinsky, I.; Alpert, P. Factors governing the interannual variation and the long-term trend of the 850 hPa temperature over Israel. Q. J. R. Meteorol. Soc. 2010, 136, 305–318. [Google Scholar]
- Rimmer, A.; Aota, Y.; Kumagai, M.; Eckert, W. Chemical stratification in thermally stratified lakes: A chloride mass balance model. Limnol. Oceanogr. 2005, 50, 147–157. [Google Scholar] [CrossRef]
- Rimmer, A. Systems hydrology models for the Upper Catchments of the Jordan River and Lake Kinneret, Israel. Isr. J. Earth Sci. 2007, 56, 1–17. [Google Scholar] [CrossRef]
- Kaplan, A.; Volokita, M.; Zenvirth, D.; Reinhold, L. An essential role for sodium in the bicarbonate transporting system of the cyanobacterium Anabaena variabilis. FEBS Lett. 1984, 176, 166–168. [Google Scholar] [CrossRef]
- Nishri, A.; Stiller, M. Dissolved Inorganic Carbon (DIC). In Lake Kinneret: Ecology and Management; Zohary, T., Sukenik, A., Berman, T., Nishri, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 397–415. [Google Scholar]
- Kaplan, A.; Reinhold, L. CO2 concentrating mechanisms in photosynthetic microorganisms. Annu. Rev. Plant Biol. 1999, 50, 539–570. [Google Scholar] [CrossRef]
- Gophen, M.; Smith, V.H.; Nishri, A.; Threlkeld, S.T. Nitrogen deficiency, phosphorus sufficiency, and the invasion of Lake Kinneret, Israel, by the N2-fixing cyanobacterium Aphanizomenon ovalisporum. Aquat. Sci. 1999, 61, 293–306. [Google Scholar]
- Hadas, O.; Pinkas, R.; Malinsky-Rushansky, N.; Shalev-Alon, G.; Delphine, E.; Berner, T.; Sukenik, A.; Kaplan, A. Physiological variables determined under laboratory conditions may explain the bloom of Aphanizomenon ovalisporum in Lake Kinneret. Eur. J. Phycol. 2002, 37, 259–267. [Google Scholar] [CrossRef]
- Holland, D.P.; Pantorno, A.; Orr, P.T.; Stojkovic, S.; Beardall, J. The impacts of a high CO2 environment on a bicarbonate user: The cyanobacterium Cylindrospermopsis raciborskii. Water Res. 2012, 46, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Bar-Yosef, Y.; Murik, O.; Sukenik, A.; Hadas, O.; Kaplan, A. Multiannual variations in phytoplankton populations: What distinguished the blooms of Aphanizomenon ovalisporum in Lake Kinneret in 2010 from 2009? Environ. Microbiol. Rep. 2012, 4, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Bar-Yosef, Y.; Sukenik, A.; Hadas, O.; Viner-Mozzini, Y.; Kaplan, A. Enslavement in the water body by toxic Aphanizomenon ovalisporum, inducing alkaline phosphatase in phytoplanktons. Curr. Biol. 2010, 20, 1557–1561. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Kong, F.; Shi, X.; Cao, H. Morphological response of Microcystis aeruginosa to grazing by different sorts of zooplankton. Hydrobiologia 2006, 563, 225–230. [Google Scholar] [CrossRef]
- Deng, J.; Qin, B.; Paerl, H.W.; Zhang, Y.; Wu, P.; Ma, J.; Chen, Y. Effects of nutrients, temperature and their interactions on spring phytoplankton community succession in Lake Taihu, China. PloS One 2014, 9. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hadas, O.; Kaplan, A.; Sukenik, A. Long-Term Changes in Cyanobacteria Populations in Lake Kinneret (Sea of Galilee), Israel: An Eco-Physiological Outlook. Life 2015, 5, 418-431. https://doi.org/10.3390/life5010418
Hadas O, Kaplan A, Sukenik A. Long-Term Changes in Cyanobacteria Populations in Lake Kinneret (Sea of Galilee), Israel: An Eco-Physiological Outlook. Life. 2015; 5(1):418-431. https://doi.org/10.3390/life5010418
Chicago/Turabian StyleHadas, Ora, Aaron Kaplan, and Assaf Sukenik. 2015. "Long-Term Changes in Cyanobacteria Populations in Lake Kinneret (Sea of Galilee), Israel: An Eco-Physiological Outlook" Life 5, no. 1: 418-431. https://doi.org/10.3390/life5010418
APA StyleHadas, O., Kaplan, A., & Sukenik, A. (2015). Long-Term Changes in Cyanobacteria Populations in Lake Kinneret (Sea of Galilee), Israel: An Eco-Physiological Outlook. Life, 5(1), 418-431. https://doi.org/10.3390/life5010418