Sustainability of a Compartmentalized Host-Parasite Replicator System under Periodic Washout-Mixing Cycles


Abstract
:1. Introduction
2. Materials and Methods
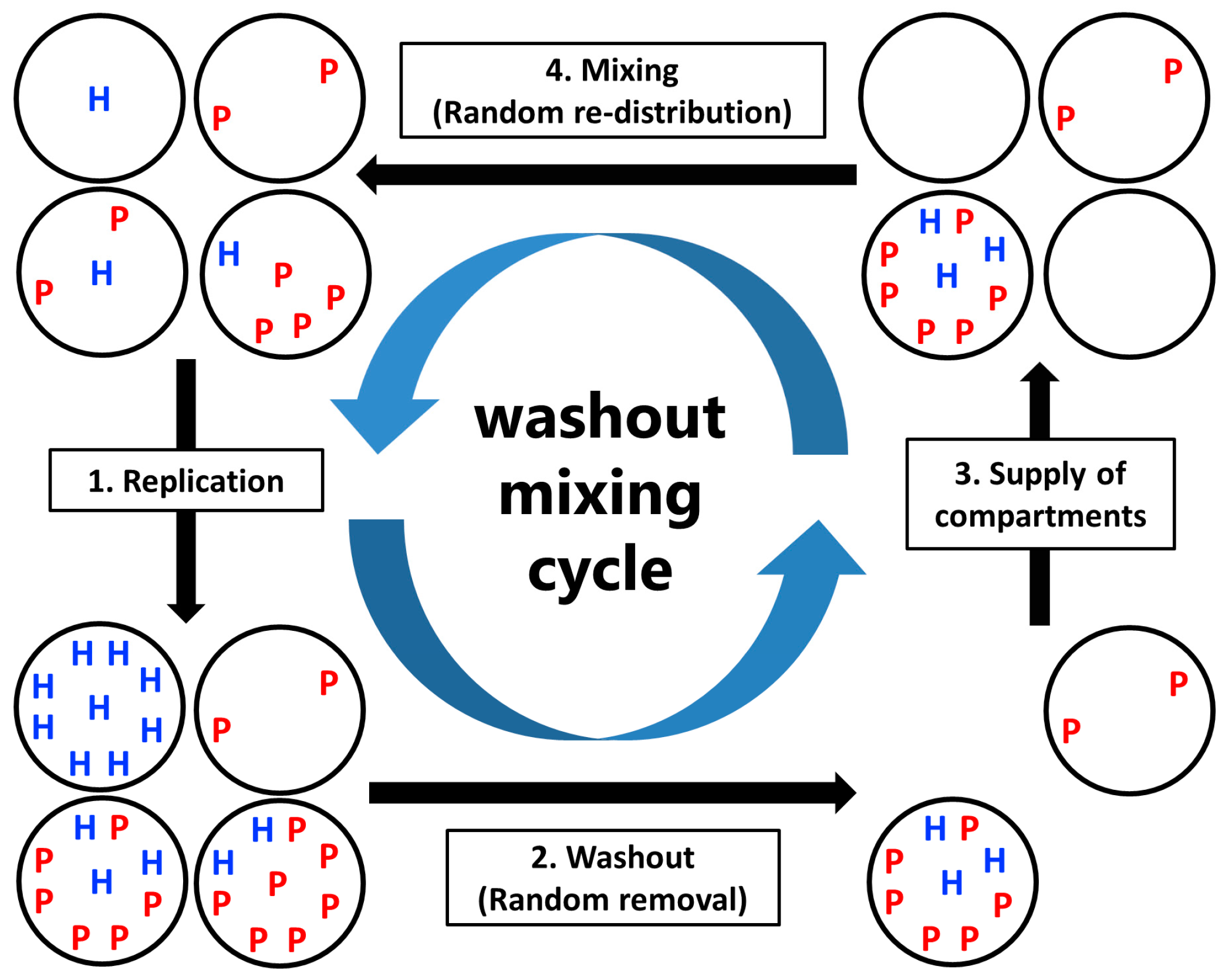
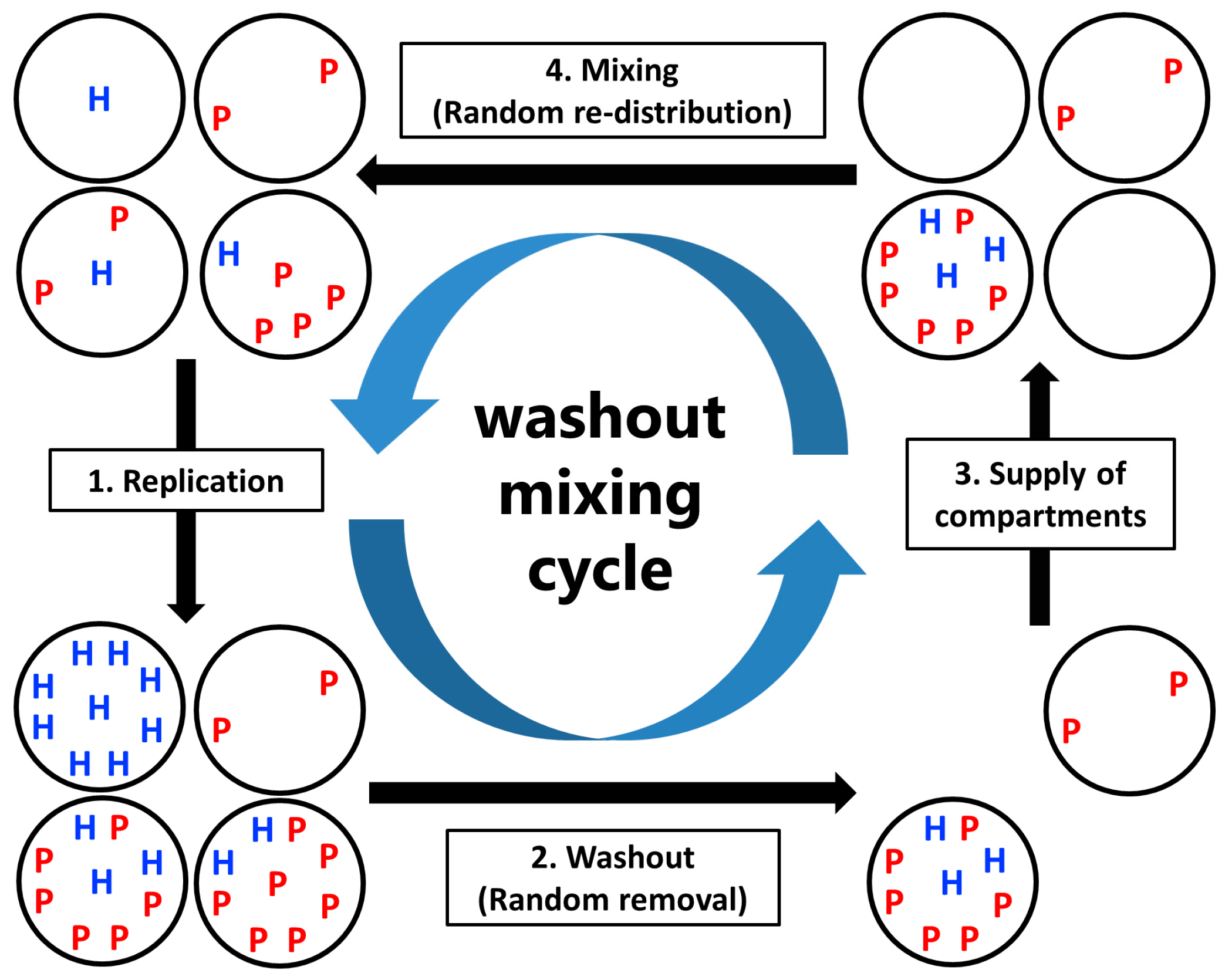
Model: CHPR System under Periodic Washout-Mixing Cycles
- (1)
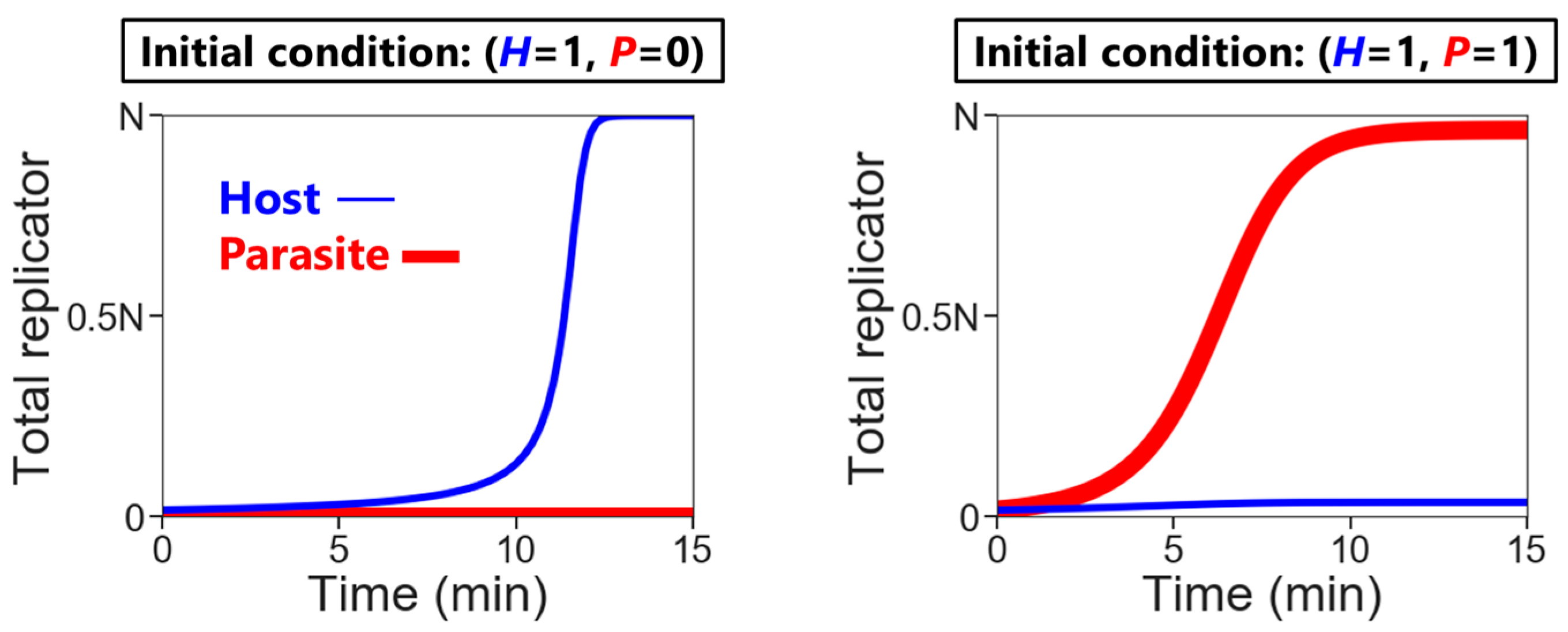
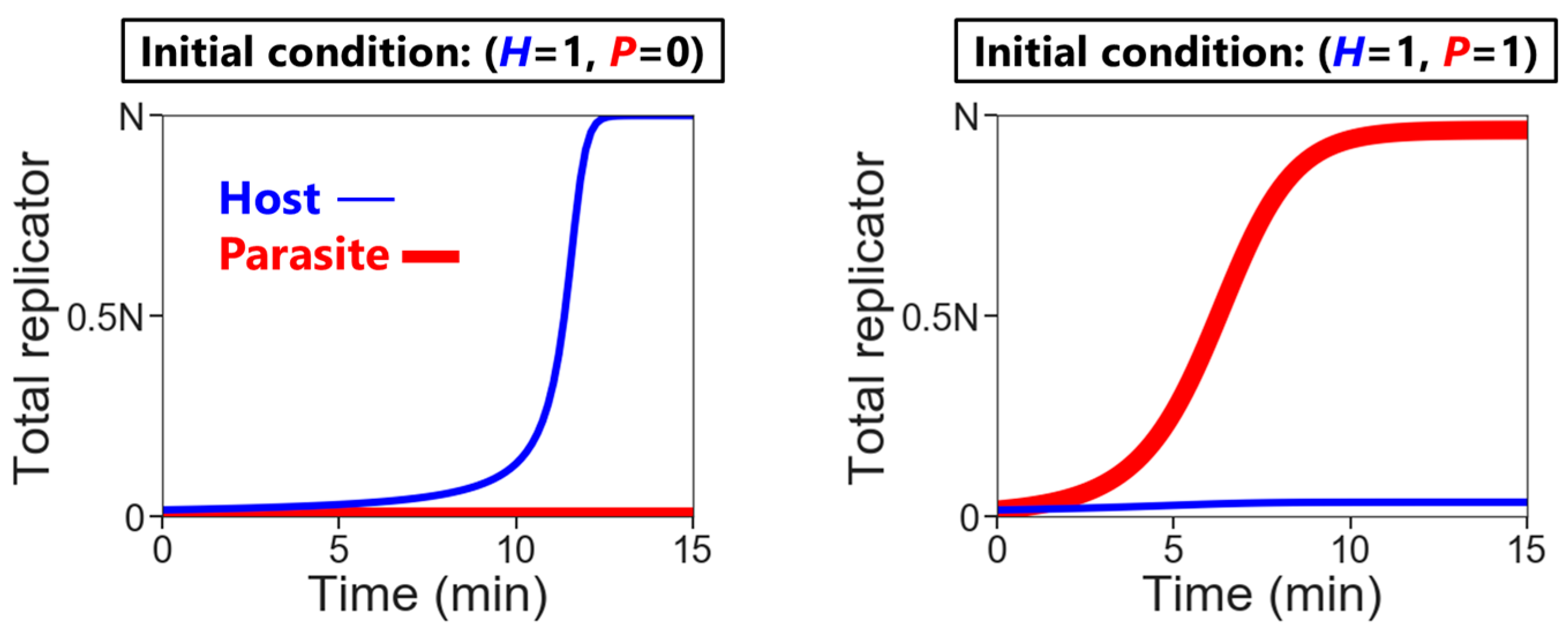
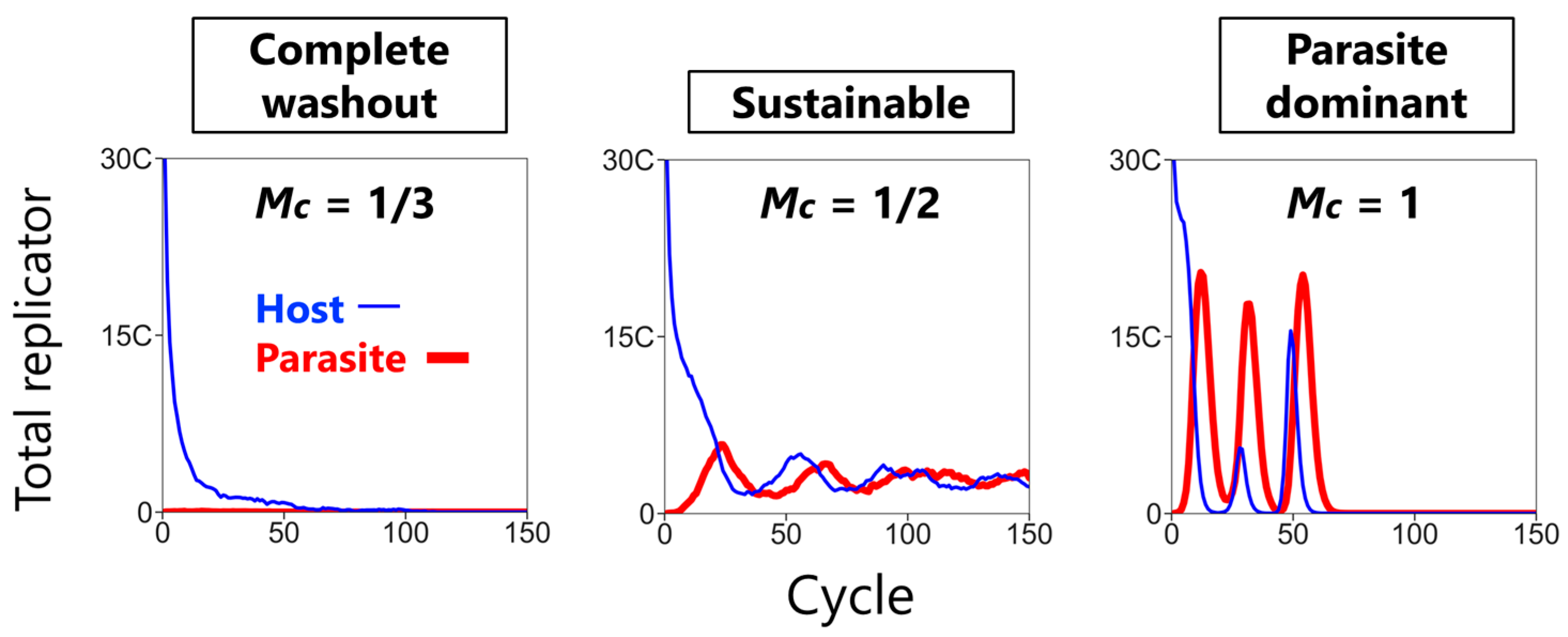
- Replication: The hosts replicate by themselves, while the parasites replicate depending on the hosts. The replication reactions occur for a fixed period of time in each compartment, according to the differential equations,andwhere H and P are the numbers of the hosts and the parasites in each compartment, respectively, and N is the amount of nutrients. Here, we assumed the second-order reactions based on the idea that replication occurs via the host (e.g., a replicase ribozyme) binding to a template (i.e., the host itself or the parasite). Reaction constants kH and kP used here were determined, based on the realistic values obtained from our previous RNA replication experiments [30]. We assumed here that the parasite replicates much faster than the host (i.e., kP is much larger than kH). Therefore, the hosts replicate better when the compartment does not contain any parasites (Figure 2, left), while in the presence of the parasites, the parasites replicate predominantly (Figure 2, right). Parasite generation stochastically occurs before replication. If a compartment has the hosts, the probability of an appearance of one molecule of the parasite is
- (2)
- Washout: A fixed number C × (1 − 1/W) compartments are randomly removed from the population.
- (3)
- Supply of compartments: Compartments filled with nutrients but without the replicators are replenished to maintain the population size of compartments C.
- (4)
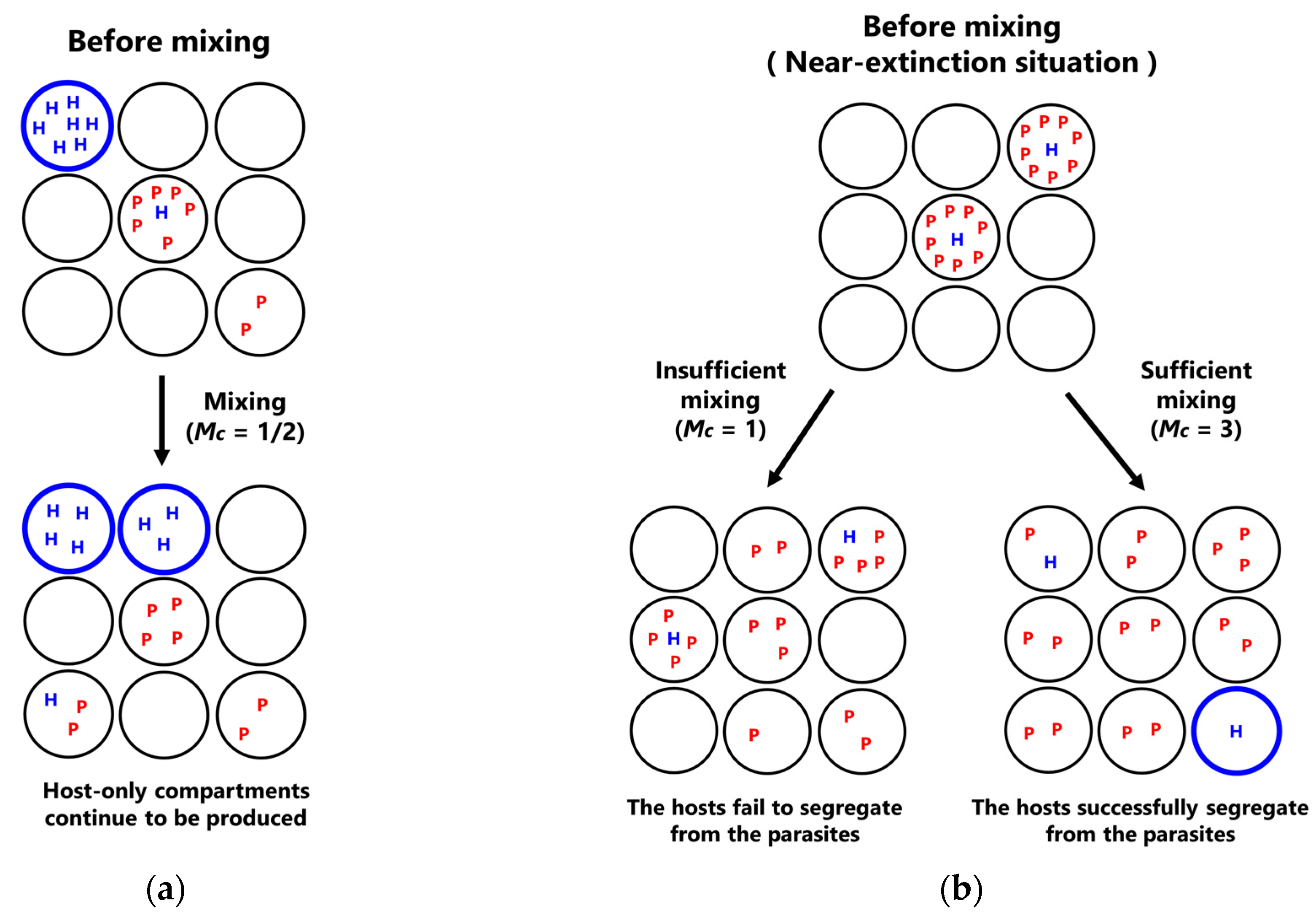
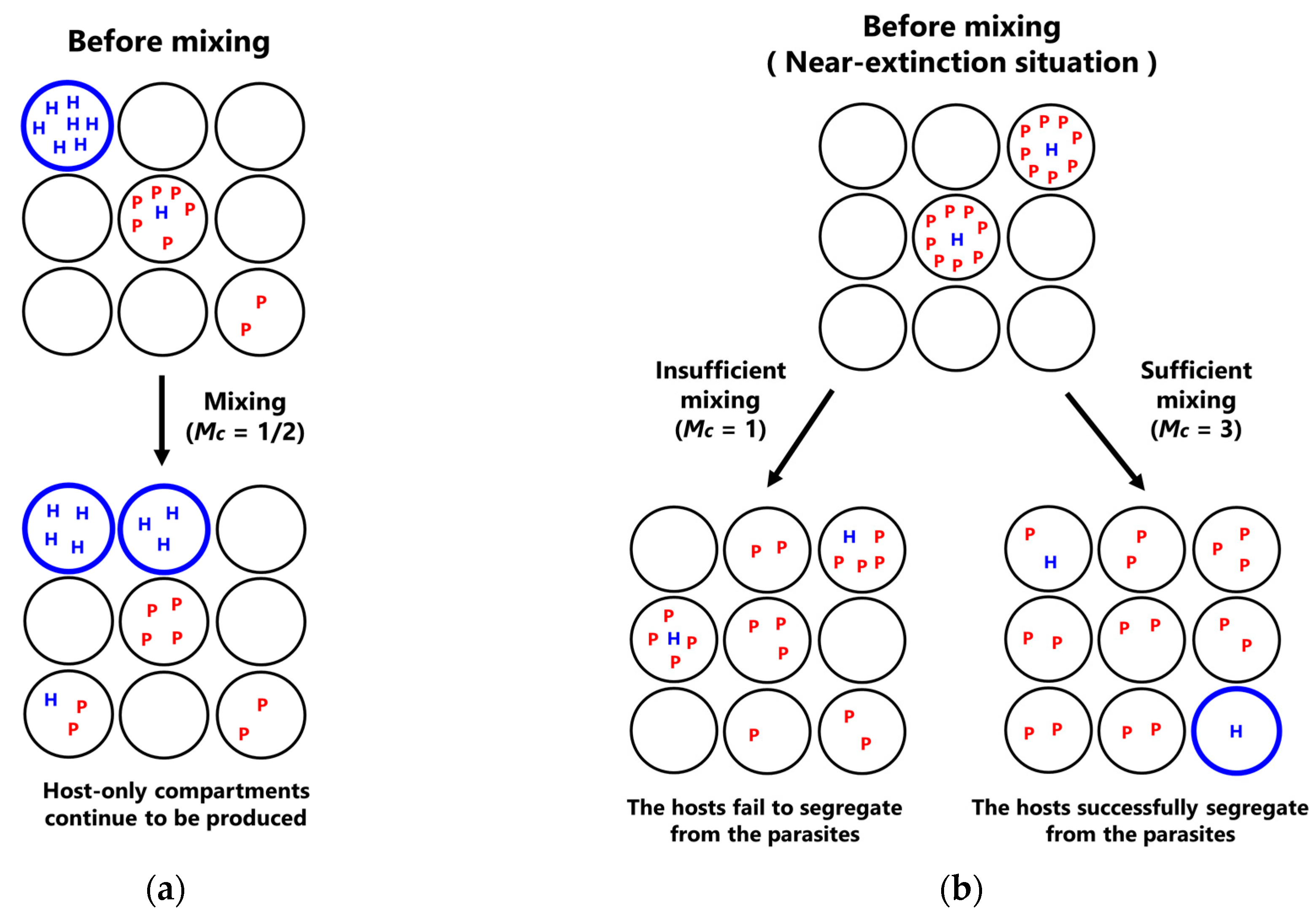
- Mixing: Replicators in compartments are re-distributed through a fixed number of random fusion–fission events among compartments. In a single fusion-fission event, two compartments are randomly chosen, and the number of replicators from each compartment are summed and re-distributed to each compartment, according to the binomial distribution (p = 0.5). Note that the numbers of the hosts and the parasites after re-distribution become integers by omitting the fractional-part of the summed number of the hosts and the parasites. The fusion-fission events repeatedly occur according to a fixed parameter M (mixing intensity).
3. Results
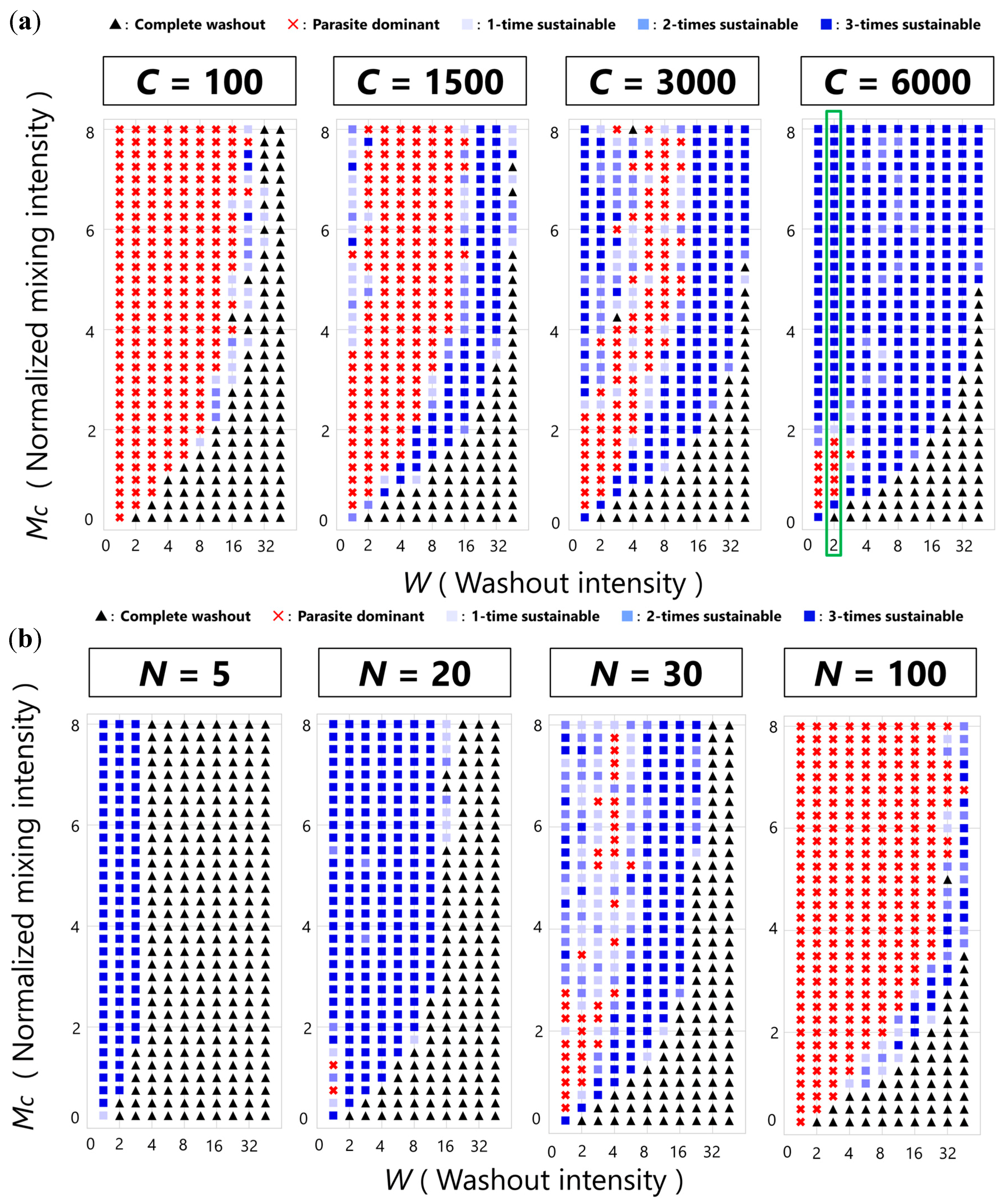
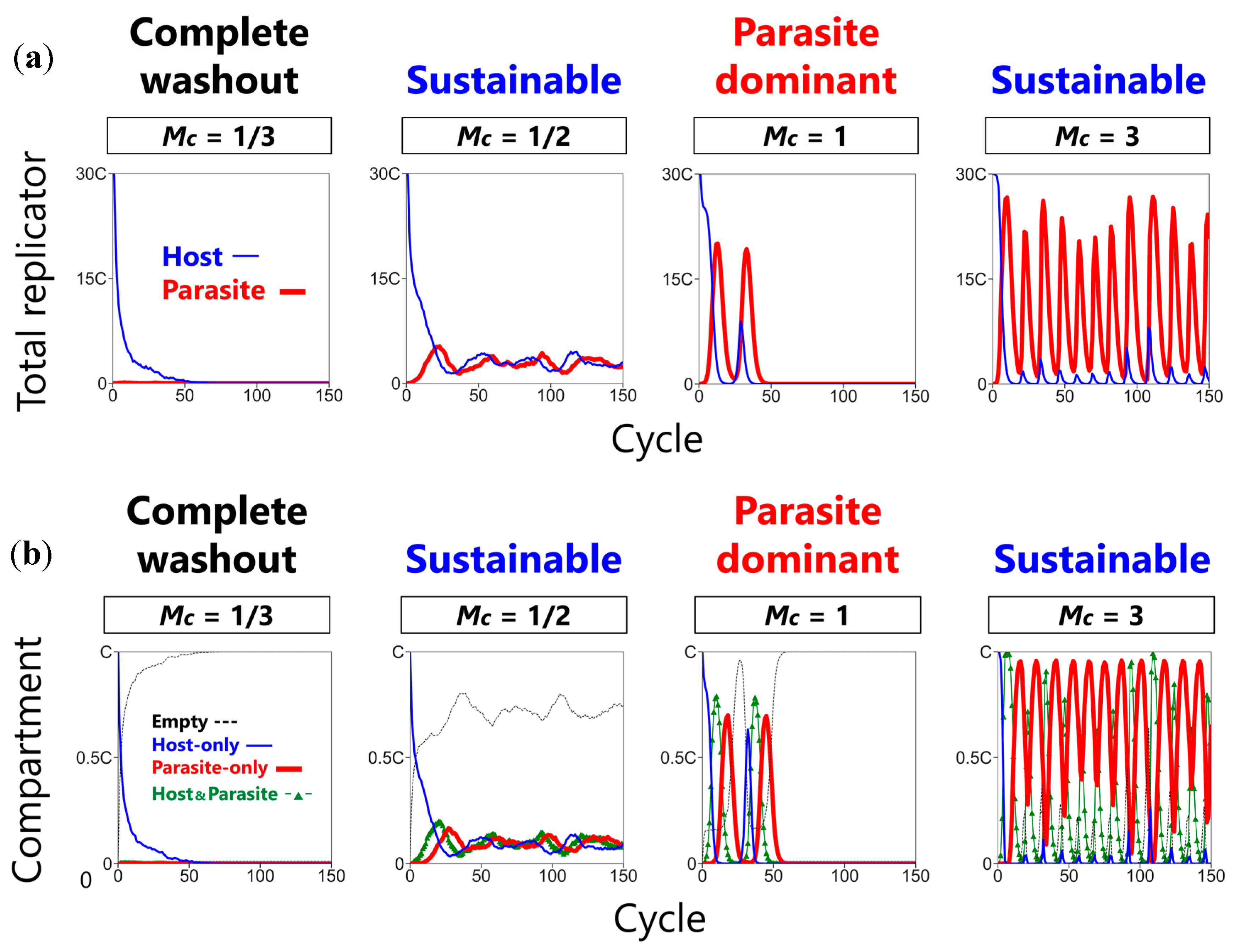
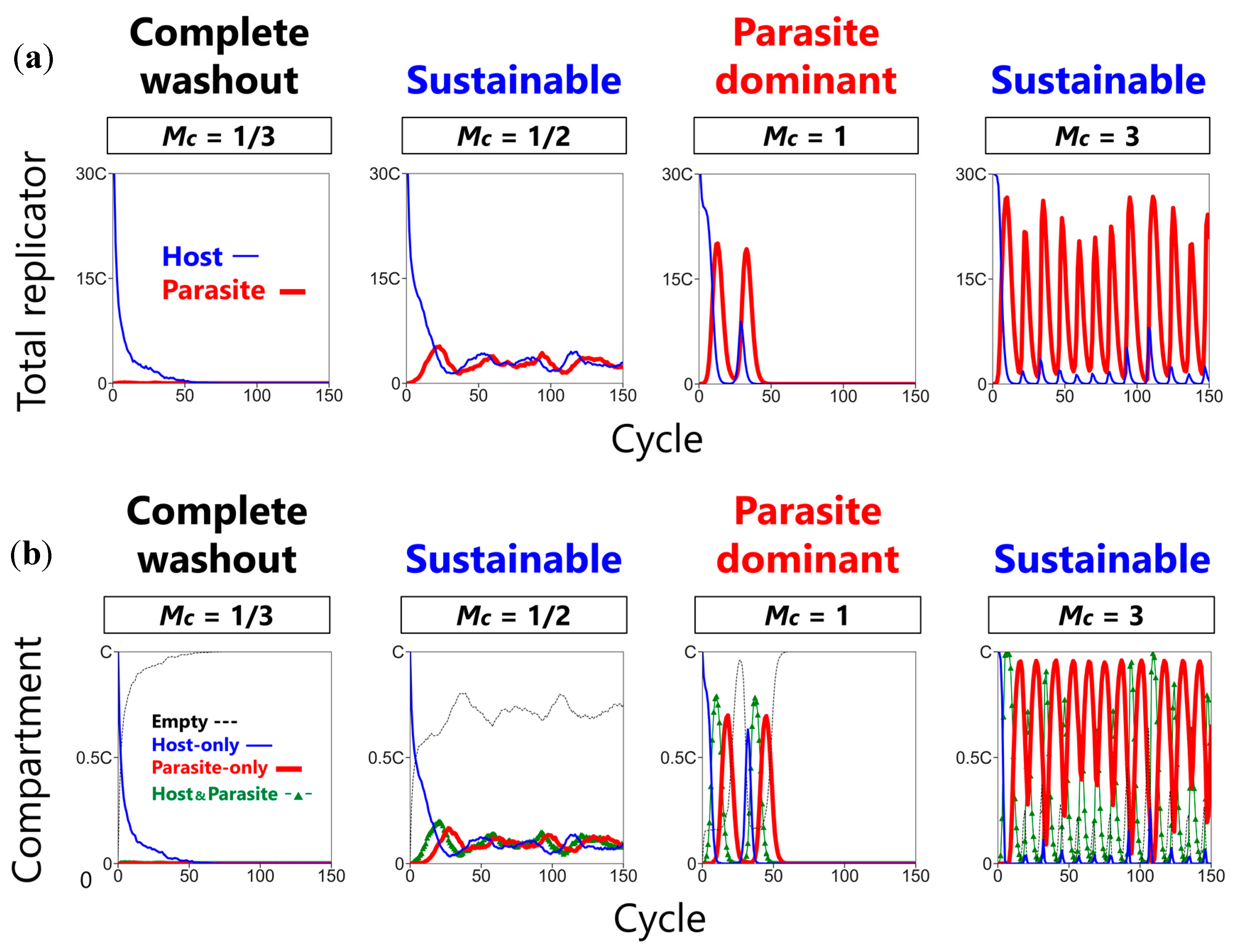
3.1. Three Phases of the CHPR System
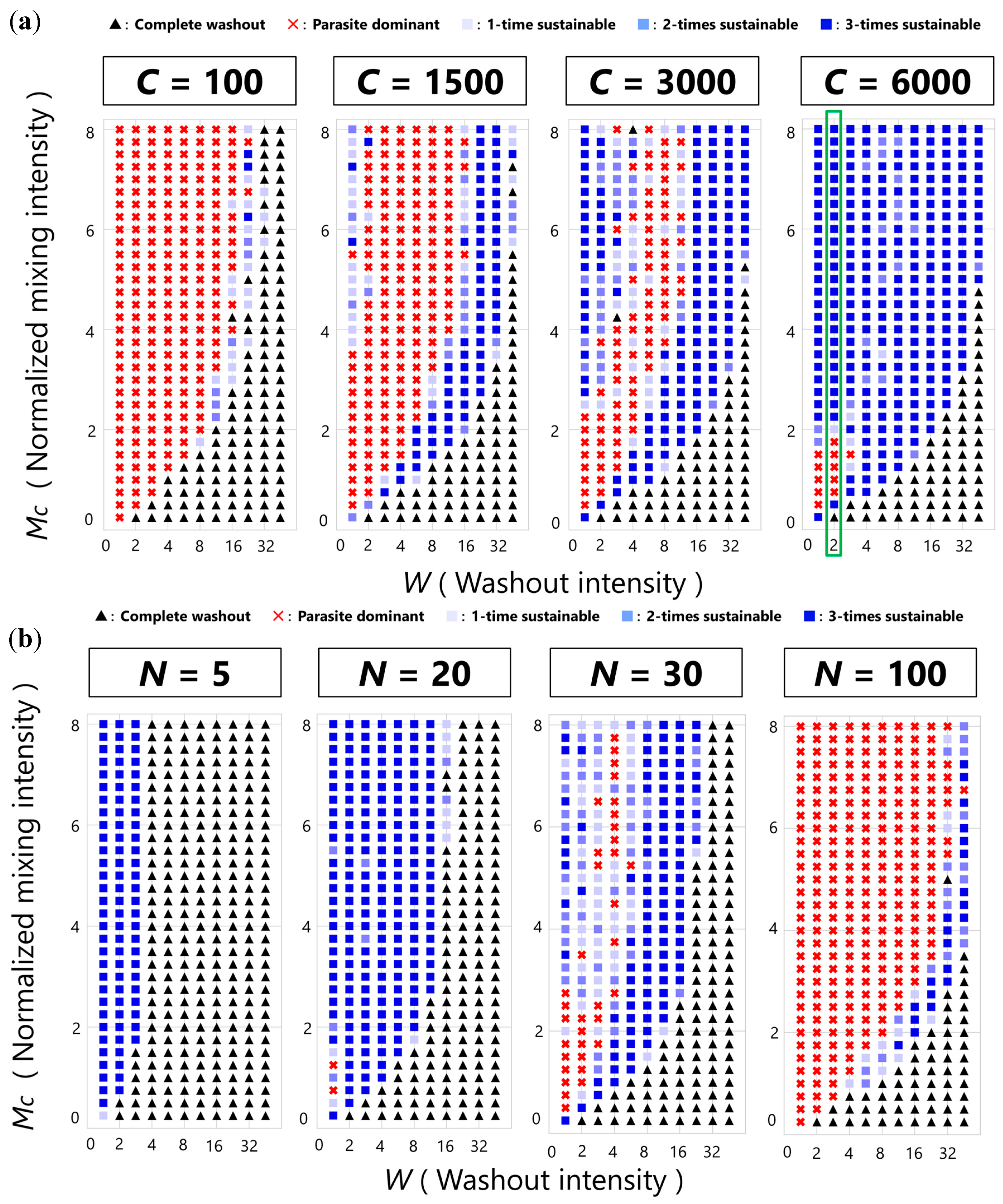
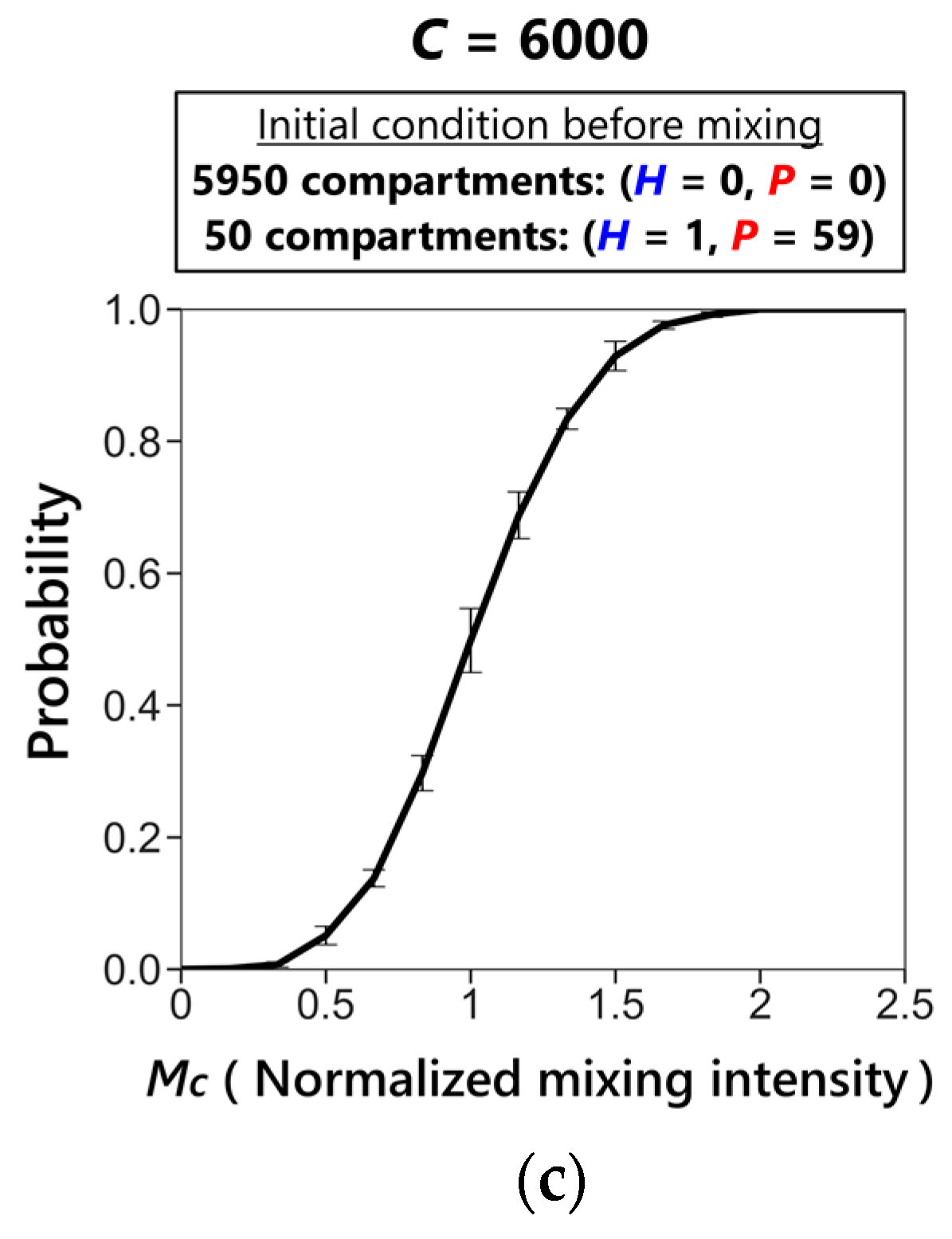
3.2. Sustainability of the CHPR System
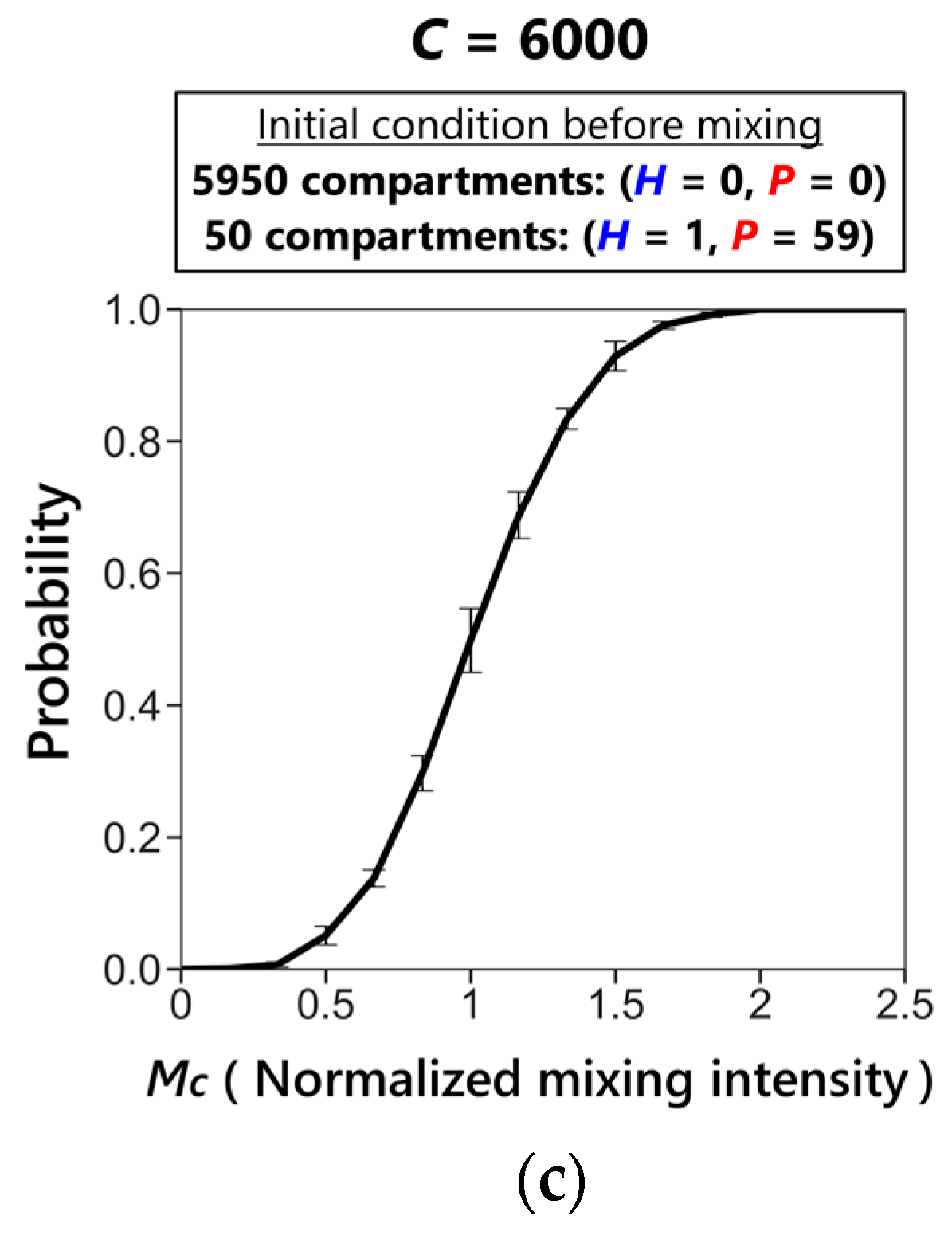
3.3. Phase Transition between Sustainable and Extinct Regions
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gilbert, W. Origin of life: The RNA world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Orgel, L.E. Molecular replication. Nature 1992, 358, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Joyce, G.F. The antiquity of RNA-based evolution. Nature 2002, 418, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Yarus, M. Life from an RNA World: The Ancestor within; Harvard University Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Luisi, P.L. The Emergence of Life: From Chemical Origins to Synthetic Biology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar]
- Kun, Á.; Szilágyi, A.; Könnyu, B.; Boza, G.; Zachar, I.; Szathmáry, E. The dynamics of the RNA world: Insights and challenges. Ann. N. Y. Acad. Sci. 2015, 1341, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Szilágyi, A.; Zachar, I.; Scheuring, I.; Kun, Á.; Könnyű, B.; Czárán, T. Ecology and Evolution in the RNA World Dynamics and Stability of Prebiotic Replicator Systems. Life 2017, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Eigen, M.; Schuster, P. The hypercycle. Naturwissenschaften 1978, 65, 7–41. [Google Scholar] [CrossRef]
- Takeuchi, N.; Hogeweg, P. Evolutionary dynamics of RNA-like replicator systems: A bioinformatic approach to the origin of life. Phys. Life Rev. 2012, 9, 219–263. [Google Scholar] [CrossRef] [PubMed]
- Eugene, V.K.; Yuri, I.W.; Mikhail, I.K. Inevitability of the emergence and persistence of genetic parasites caused by evolutionary instability of parasite-free states. Biol. Direct 2017, 12, 1–12. [Google Scholar]
- Bresch, C.; Niesert, U.; Harnasch, D. Hypercycles, parasites and packages. J. Theor. Biol. 1980, 85, 399–405. [Google Scholar] [CrossRef]
- Niesert, U.; Harnasch, D.; Bresch, C. Origin of life between Scylla and Charybdis. J. Mol. Evol. 1981, 17, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Szathmáry, E.; Demeter, L. Group selection of early replicators and the origin of life. J. Theor. Biol. 1987, 128, 463–486. [Google Scholar] [CrossRef]
- Grey, D.; Hutson, V.; Szathmary, E. A re-examination of the stochastic corrector model. Proc. R. Soc. Lond. B Biol. Sci. 1995, 262, 29–35. [Google Scholar] [CrossRef]
- Fontanari, J.F.; Santos, M.; Szathmáry, E. Coexistence and error propagation in pre-biotic vesicle models: A group selection approach. J. Theor. Biol. 2006, 239, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, N.; Hogeweg, P. Multilevel selection in models of prebiotic evolution II: A direct comparison of compartmentalization and spatial self-organization. PLoS Comput. Biol. 2009, 5, e1000542. [Google Scholar] [CrossRef] [PubMed]
- Markvoort, A.J.; Sinai, S.; Nowak, M.A. Computer simulations of cellular group selection reveal mechanism for sustaining cooperation. J. Theor. Biol. 2014, 357, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Hubai, A.G.; Kun, Á. Maximal gene number maintainable by stochastic correction—The second error threshold. J. Theor. Biol. 2016, 405, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, N.; Hogeweg, P.; Kaneko, K. The origin of a primordial genome through spontaneous symmetry breaking. Nat. Commun. 2017, 8, 250. [Google Scholar] [CrossRef] [PubMed]
- Károlyi, G.; Péntek, Á.; Scheuring, I.; Tél, T.; Toroczkai, Z. Chaotic flow: The physics of species coexistence. Proc. Natl. Acad. Sci. USA 2000, 97, 13661–13665. [Google Scholar] [CrossRef] [PubMed]
- Luisi, P.L.; Walde, P.; Oberholzer, T. Lipid vesicles as possible intermediates in the origin of life. Curr. Opin. Colloid Interface Sci. 1999, 4, 33–39. [Google Scholar] [CrossRef]
- Hanczyc, M.M.; Szostak, J.W. Replicating vesicles as models of primitive cell growth and division. Curr. Opin. Chem. Biol. 2004, 8, 660–664. [Google Scholar] [CrossRef] [PubMed]
- Oparin, A.I. The Origin of Life on the Earth, 3rd ed.; Academic Press: New York, NY, USA, 1957. [Google Scholar]
- Frankel, E.A.; Bevilacqua, P.C.; Keating, C.D. Polyamine/nucleotide coacervates provide strong compartmentalization of Mg2+, nucleotides, and RNA. Langmuir 2016, 32, 2041–2049. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Martin, W. On the origin of genomes and cells within inorganic compartments. Trends Genet. 2005, 21, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Branciamore, S.; Gallori, E.; Szathmáry, E.; Czárán, T. The origin of life: Chemical evolution of a metabolic system in a mineral honeycomb? J. Mol. Evol. 2009, 69, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobson, C.M.; Ellison, G.B.; Tuck, A.F.; Vaida, V. Atmospheric aerosols as prebiotic chemical reactors. Proc. Natl. Acad. Sci. USA 2000, 97, 11864–11868. [Google Scholar] [CrossRef] [PubMed]
- Trevors, J.T.; Pollack, G.H. Hypothesis: The origin of life in a hydrogel environment. Prog. Biophys. Mol. Biol. 2005, 89, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bansho, Y.; Ichihashi, N.; Kazuta, Y.; Matsuura, T.; Suzuki, H.; Yomo, T. Importance of parasite RNA species repression for prolonged translation-coupled RNA self-replication. Chem. Biol. 2012, 19, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Bansho, Y.; Furubayashi, T.; Ichihashi, N.; Yomo, T. Host–parasite oscillation dynamics and evolution in a compartmentalized RNA replication system. Proc. Natl. Acad. Sci. USA 2016, 113, 4045–4050. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, S.; Kun, Á.; Ryckelynck, M.; Coldren, F.; Szilágyi, A.; Jossinet, F.; Rick, C.; Nghe, P.; Szathmáry, E.; Griffiths, A.D. Transient compartmentalization of RNA replicators prevents extinction due to parasites. Science 2016, 354, 1293–1296. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.S. A theory of group selection. Proc. Natl. Acad. Sci. USA 1975, 72, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Rinehart, J.S. Fluctuations in geyser activity caused by variations in Earth tidal forces, barometric pressure, and tectonic stresses. J. Geophys. Res. Biogeosci. 1972, 77, 342–350. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Default |
|---|---|
| Population size of compartments (C) | Variable (fixed in a run) |
| Mixing intensity (M) | Variable (fixed in a run) |
| Normalized mixing intensity (Mc = M/C) | Variable (fixed in a run) |
| Washout intensity (W) | Variable (fixed in a run) |
| Nutrient amount (N) | Variable (fixed in a run) |
| Rate constant of the host (kH) | 0.091 per minute |
| Rate constant of the parasite (kP) | 0.480 per minute |
| Parasite generation rate | 0.0002 |
| Reaction time | 15 min |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furubayashi, T.; Ichihashi, N. Sustainability of a Compartmentalized Host-Parasite Replicator System under Periodic Washout-Mixing Cycles. Life 2018, 8, 3. https://doi.org/10.3390/life8010003
Furubayashi T, Ichihashi N. Sustainability of a Compartmentalized Host-Parasite Replicator System under Periodic Washout-Mixing Cycles. Life. 2018; 8(1):3. https://doi.org/10.3390/life8010003
Chicago/Turabian StyleFurubayashi, Taro, and Norikazu Ichihashi. 2018. "Sustainability of a Compartmentalized Host-Parasite Replicator System under Periodic Washout-Mixing Cycles" Life 8, no. 1: 3. https://doi.org/10.3390/life8010003