Chemomimesis and Molecular Darwinism in Action: From Abiotic Generation of Nucleobases to Nucleosides and RNA


 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Principles Underlying Progress towards Further Complexity
3. Principles for Systems Chemistry
4. One-Pot Initial Events under a Variety of Energy Sources: An Example
5. From Complex Mixtures to Pre-Genetic Materials
6. The Nucleobases are the Right Ones since the Beginning. The Case of AICA and fAICA
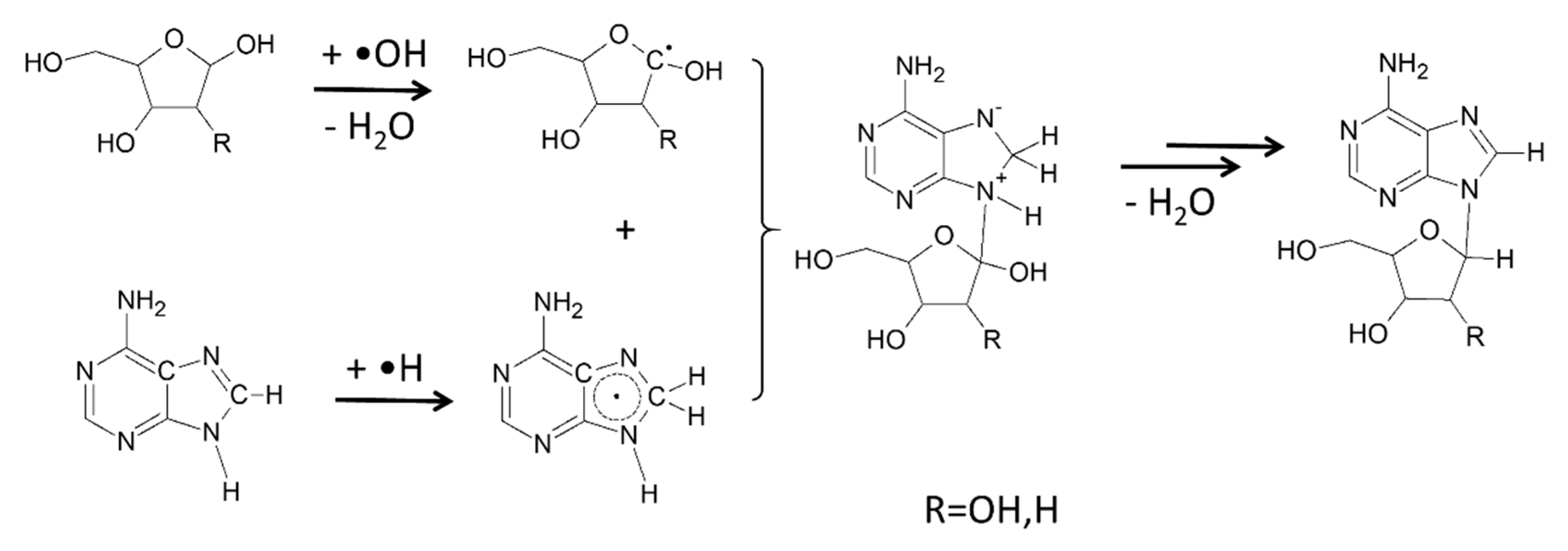
7. Focusing on Nucleosides
8. Regio- and Stereoselectivity of Nucleoside Formation is Conserved from the Beginning
9. Chemomimetic RNA
10. Exploring the Environment for the First Effective Phosphorylation Agents
11. Focusing on Differential Kinetic Stability
12. Focusing on Oligomerization
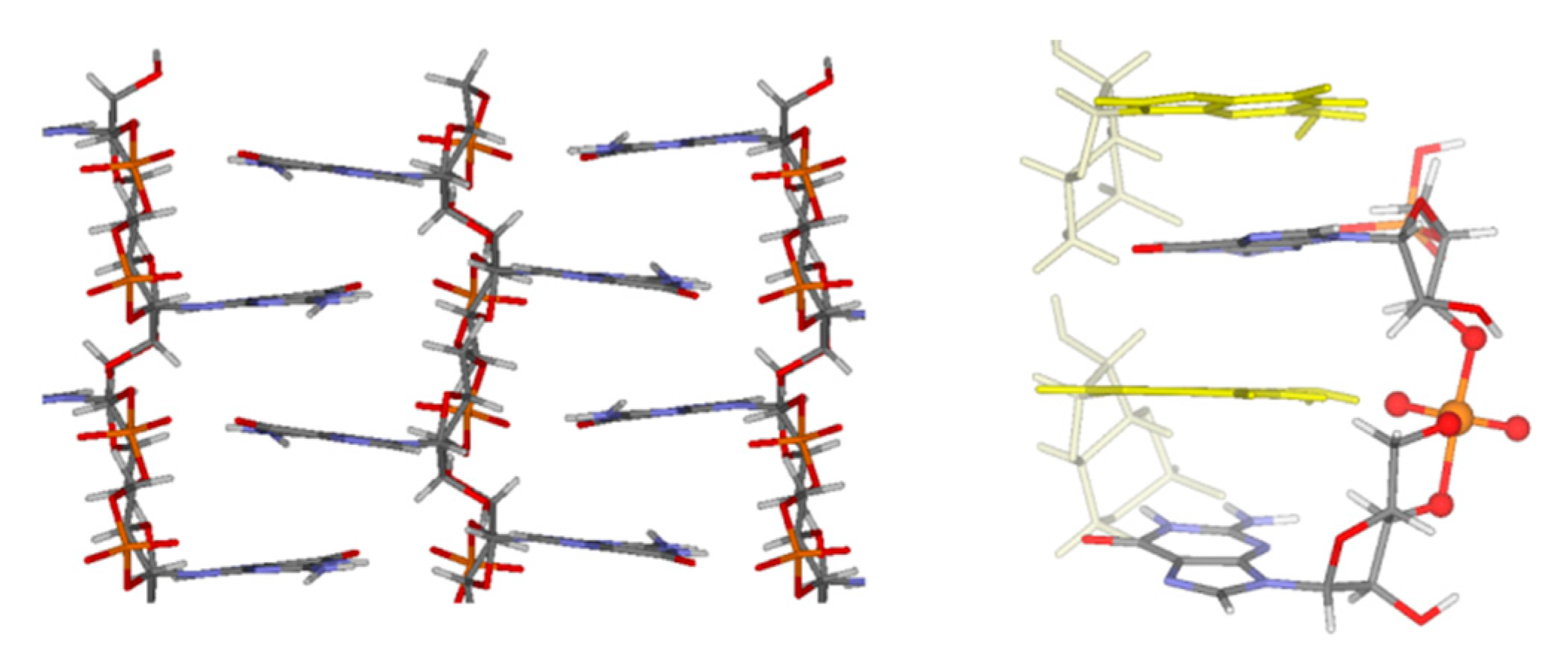
13. Chemomimesis of Cyclic Nucleotides
14. Concluding Remarks
Funding
Conflicts of Interest
References
- Miller, S.L.; Urey, H.C. Organic compound synthesis on the primitive earth. Science 1959, 130, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Chyba, C.; Sagan, C. Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: An inventory for the origins of life. Nature 1992, 355, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.A.; Ricardo, A.; Carrigan, M.A. Is there a common chemical model for life in the universe? Curr. Opin. Chem. Biol. 2004, 8, 672–689. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.A. Paradoxes in the origin of life. Orig. Life Evol. Biosph. 2014, 44, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Chyba, C.F.; Thomas, P.J.; Brookshaw, L.; Sagan, C. Cometary delivery of organic molecules to the early Earth. Science 1990, 249, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Kopplin, P.; Gabelica, Z.; Gougeon, R.D.; Fekete, A.; Kanawati, B.; Harir, M.; Gebefuegi, I.; Eckel, G.; Hertkorn, N. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl. Acad. Sci. USA 2010, 107, 2763–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, A.S.; Stern, J.C.; Elsila, J.E.; Glavin, D.P.; Dworkin, J.P. Understanding prebiotic chemistry through the analysis of extraterrestrial amino acids and nucleobases in meteorites. Chem. Soc. Rev. 2012, 41, 5459–5472. [Google Scholar] [CrossRef] [PubMed]
- James, E.G.; Padelis, P.P. Molecular and atomic line surveys of galaxies. I. The dense, star-forming gas phase as a beacon. Astrophys. J. 2012, 757, 156. [Google Scholar] [CrossRef]
- Carota, E.; Botta, G.; Rotelli, L.; Di Mauro, E.; Saladino, R. Current advances in prebiotic chemistry under space conditions. Curr. Org. Chem. 2015, 19, 1963–1979. [Google Scholar] [CrossRef]
- Rubin, R.H.; Swenson, G.W., Jr.; Benson, R.C.; Tigelaar, H.L.; Flygare, W.H. Microwave detection of interstellar formamide. Astrophys. J. 1971, 169, L39. [Google Scholar] [CrossRef]
- Ashkenasy, G.; Hermans, T.M.; Otto, S.; Taylor, A.F. Systems chemistry. Chem. Soc. Rev. 2017, 46, 2543–2554. [Google Scholar] [CrossRef] [PubMed]
- Stankiewicz, J.; Eckardt, L.H. Chembiogenesis 2005 and systems chemistry workshop. Angew. Chem. Int. Ed. 2006, 45, 342–344. [Google Scholar] [CrossRef]
- Kindermann, M.; Stahl, I.; Reimold, M.; Pankau, W.M.; von Kiedrowski, G. Systems chemistry: Kinetic and computational analysis of a nearly exponential organic replicator. Angew. Chem. Int. Ed. 2005, 44, 6750–6755. [Google Scholar] [CrossRef] [PubMed]
- Sadownik, J.W.; Mattia, E.; Nowak, P.; Otto, S. Diversification of self-replicating molecules. Nat. Chem. 2016, 8, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Wicken, J.S. An organismic critique of molecular Darwinism. J. Theor. Biol. 1985, 117, 545–561. [Google Scholar] [CrossRef]
- Arber, W. Molecular Darwinism: The contingency of spontaneous genetic variation. Genome Biol. Evol. 2011, 3, 1090–1092. [Google Scholar] [CrossRef] [PubMed]
- Küppers, B.-O. Information and the Origin of Life; MIT Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Higgs, P.G. Chemical evolution and the evolutionary definition of life. J. Mol. Evol. 2017, 84, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Eschenmoser, A.; Loewenthal, E. Chemistry of potentially prebiological natural products. Chem. Soc. Rev. 1992, 21, 1–16. [Google Scholar] [CrossRef]
- Menor-Salván, C.; Marín-Yaseli, M.R. A new route for the prebiotic synthesis of nucleobases and hydantoins in water/ice solutions involving the photochemistry of acetylene. Chem. Eur. J. 2013, 19, 6488–6497. [Google Scholar] [CrossRef] [PubMed]
- Pereto, J. Out of fuzzy chemistry: From prebiotic chemistry to metabolic networks. Chem. Soc. Rev. 2012, 41, 5394–5403. [Google Scholar] [CrossRef] [PubMed]
- Airapetian, V.S.; Glocer, A.; Gronoff, G.; Hebrard, E.; Danchi, W. Prebiotic chemistry and atmospheric warming of early Earth by an active young sun. Nat. Geosci. 2016, 9, 452–455. [Google Scholar] [CrossRef]
- Saladino, R.; Botta, G.; Bizzarri, B.M.; Di Mauro, E.; Garcia Ruiz, J.M. A global scale scenario for prebiotic chemistry: Silica-based self-assembled mineral structures and formamide. Biochemistry 2016, 55, 2806–2811. [Google Scholar] [CrossRef] [PubMed]
- Mulkidjanian, A.Y.; Bychkov, A.Y.; Dibrova, D.V.; Galperin, M.Y.; Koonin, E.V. Origin of first cells at terrestrial, anoxic geothermal fields. Proc. Natl. Acad. Sci. USA 2012, 109, E821–E830. [Google Scholar] [CrossRef] [PubMed]
- Djokic, T.; van Kranendonk, M.J.; Campbell, K.A.; Walter, M.R.; Ward, C.R. Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits. Nat. Commun. 2017, 8, 15263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferus, M.; Pietrucci, F.; Saitta, A.M.; Knížek, A.; Kubelík, P.; Ivanek, O.; Shestivska, V.; Civiš, S. Formation of nucleobases in a Miller–Urey reducing atmosphere. Proc. Natl. Acad. Sci. USA 2017, 114, 4306–4311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferus, M.; Nesvorný, D.; Šponer, J.; Kubelík, P.; Michalčíková, R.; Shestivská, V.; Šponer, J.E.; Civiš, S. High-energy chemistry of formamide: A unified mechanism of nucleobase formation. Proc. Natl. Acad. Sci. USA 2015, 112, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Crestini, C.; Ciciriello, F.; Costanzo, G.; Di Mauro, E. Formamide chemistry and the origin of informational polymers. Chem. Biodivers. 2007, 4, 694–720. [Google Scholar] [CrossRef] [PubMed]
- Kua, J.; Thrush, K.L. HCN, formamidic acid, and formamide in aqueous solution: A free energy map. J. Phys. Chem. B 2016, 120, 8175–8185. [Google Scholar] [CrossRef] [PubMed]
- Adande, G.R.; Woolf, N.J.; Ziurys, L.M. Observations of interstellar formamide: Availability of a prebiotic precursor in the galactic habitable zone. Astrobiology 2013, 13, 439–453. [Google Scholar] [CrossRef] [PubMed]
- López-Sepulcre, A.; Jaber, A.A.; Mendoza, E.; Lefloch, B.; Ceccarelli, C.; Vastel, C.; Bachiller, R.; Cernicharo, J.; Codella, C.; Kahane, C.; et al. Shedding light on the formation of the pre-biotic molecule formamide with ASAI. Mon. Notices R. Astron. Soc. 2015, 449, 2438–2458. [Google Scholar] [CrossRef] [Green Version]
- Biver, N.; Bockelée-Morvan, D.; Debout, V.; Crovisier, J.; Boissier, J.; Lis, D.C.; Dello Russo, N.; Moreno, R.; Colom, P.; Paubert, G.; et al. Complex organic molecules in comets C/2012 F6 (Lemmon) and C/2013 R1 (Lovejoy): Detection of ethylene glycol and formamide. Astron. Astrophys. 2014, 566, L5. [Google Scholar] [CrossRef]
- Kröcher, O.; Elsener, M.; Jacob, E. A model gas study of ammonium formate, methanamide and guanidinium formate as alternative ammonia precursor compounds for the selective catalytic reduction of nitrogen oxides in diesel exhaust gas. Appl. Catal. B 2009, 88, 66–82. [Google Scholar] [CrossRef]
- Saladino, R.; Botta, G.; Pino, S.; Costanzo, G.; Di Mauro, E. Genetics first or metabolism first? The formamide clue. Chem. Soc. Rev. 2012, 41, 5526–5565. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Crestini, C.; Pino, S.; Costanzo, G.; Di Mauro, E. Formamide and the origin of life. Phys. Life Rev. 2012, 9, 84–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saladino, R.; Crestini, C.; Costanzo, G.; Negri, R.; Di Mauro, E. A possible prebiotic synthesis of purine, adenine, cytosine, and 4(3H)-pyrimidone from formamide: Implications for the origin of life. Bioorg. Med. Chem. 2001, 9, 1249–1253. [Google Scholar] [CrossRef]
- Saladino, R.; Šponer, J.E.; Šponer, J.; Di Mauro, E. Rewarming the primordial soup: Revisitations and rediscoveries in prebiotic chemistry. ChemBioChem 2018, 19, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Niether, D.; Afanasenkau, D.; Dhont, J.K.G.; Wiegand, S. Accumulation of formamide in hydrothermal pores to form prebiotic nucleobases. Proc. Natl. Acad. Sci. USA 2016, 113, 4272–4277. [Google Scholar] [CrossRef] [PubMed]
- Niether, D.; Wiegand, S. Heuristic approach to understanding the accumulation process in hydrothermal pores. Entropy 2017, 19, 33. [Google Scholar] [CrossRef]
- Šponer, J.E.; Šponer, J.; Nováková, O.; Brabec, V.; Šedo, O.; Zdráhal, Z.; Costanzo, G.; Pino, S.; Saladino, R.; Di Mauro, E. Emergence of the first catalytic oligonucleotides in a formamide-based origin scenario. Chem. Eur. J. 2016, 22, 3572–3586. [Google Scholar] [CrossRef] [PubMed]
- Bada, J.L.; Chalmers, J.H.; Cleaves, H.J. Is formamide a geochemically plausible prebiotic solvent? Phys. Chem. Chem. Phys. 2016, 18, 20085–20090. [Google Scholar] [CrossRef] [PubMed]
- Adam, Z.R.; Hongo, Y.; Cleaves, H.J.; Yi, R.; Fahrenbach, A.C.; Yoda, I.; Aono, M. Estimating the capacity for production of formamide by radioactive minerals on the prebiotic Earth. Sci. Rep. 2018, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Saitta, A.M.; Saija, F. Miller experiments in atomistic computer simulations. Proc. Natl. Acad. Sci. USA 2014, 111, 13768–13773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saladino, R.; Crestini, C.; Cossetti, C.; Di Mauro, E.; Deamer, D. Catalytic effects of Murchison material: Prebiotic synthesis and degradation of RNA precursors. Orig. Life Evol. Biosph. 2011, 41, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Botta, G.; Delfino, M.; Di Mauro, E. Meteorites as catalysts for prebiotic chemistry. Chem. Eur. J. 2013, 19, 16916–16922. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Carota, E.; Botta, G.; Kapralov, M.; Timoshenko, G.N.; Rozanov, A.Y.; Krasavin, E.; Di Mauro, E. Meteorite-catalyzed syntheses of nucleosides and of other prebiotic compounds from formamide under proton irradiation. Proc. Natl. Acad. Sci. USA 2015, 112, E2746–E2755. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Carota, E.; Botta, G.; Kapralov, M.; Timoshenko, G.N.; Rozanov, A.; Krasavin, E.; Di Mauro, E. First evidence on the role of heavy ion irradiation of meteorites and formamide in the origin of biomolecules. Orig. Life Evol. Biosph. 2016, 46, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Botta, L.; Saladino, R.; Bizzarri, B.M.; Cobucci-Ponzano, B.; Iacono, R.; Avino, R.; Caliro, S.; Carandente, A.; Lorenzini, F.; Tortora, A.; et al. Formamide-based prebiotic chemistry in the Phlegrean fields. Adv. Space Res. 2017. [Google Scholar] [CrossRef]
- Rotelli, L.; Trigo-Rodríguez, J.M.; Moyano-Cambero, C.E.; Carota, E.; Botta, L.; Di Mauro, E.; Saladino, R. The key role of meteorites in the formation of relevant prebiotic molecules in a formamide/water environment. Sci. Rep. 2016, 6, 38888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferus, M.; Michalčíková, R.; Shestivská, V.; Šponer, J.; Šponer, J.E.; Civiš, S. High-energy chemistry of formamide: A simpler way for nucleobase formation. J. Phys. Chem. A 2014, 118, 719–736. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, B.M.; Botta, L.; Perez-Valverde, M.I.; Saladino, R.; Di Mauro, E.; Garcia Ruiz, J.M. Silica metal-oxide vesicles catalyze comprehensive prebiotic chemistry. Chem. Eur. J. 2018. [Google Scholar] [CrossRef]
- Ferus, M.; Kubelík, P.; Knížek, A.; Pastorek, A.; Sutherland, J.; Civiš, S. High energy radical chemistry formation of HCN-rich atmospheres on early Earth. Sci. Rep. 2017, 7, 6275. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Jeilani, Y.A.; Hung, H.M.; Nguyen, M.T. Radical pathways for the prebiotic formation of pyrimidine bases from formamide. J. Phys. Chem. A 2015, 119, 8871–8883. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.A.; Kim, H.-J.; Carrigan, M.A. Asphalt, water, and the prebiotic synthesis of ribose, ribonucleosides, and RNA. Acc. Chem. Res. 2012, 45, 2025–2034. [Google Scholar] [CrossRef] [PubMed]
- Benner, S.A.; Kim, H.-J. The case for a martian origin for Earth life. In Instruments, Methods, and Missions for Astrobiology XVII; Proceedings of SPIE 9606; SPIE Press: San Diego, CA, USA, 2015. [Google Scholar] [CrossRef]
- He, C.; Lin, G.; Upton, K.T.; Imanaka, H.; Smith, M.A. Structural investigation of HCN polymer isotopomers by solution-state multidimensional NMR. J. Phys. Chem. A 2012, 116, 4751–4759. [Google Scholar] [CrossRef] [PubMed]
- Minard, R.D.; Hatcher, P.G.; Gourley, R.C.; Matthews, C.N. Structural investigations of hydrogen cyanide polymers: New insights using TMAH thermochemolysis/GC-MS. Orig. Life Evol. Biosph. 1998, 28, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Ricardo, A.; Carrigan, M.A.; Olcott, A.N.; Benner, S.A. Borate minerals stabilize ribose. Science 2004, 303, 196. [Google Scholar] [CrossRef] [PubMed]
- Joyce, G.F. Foreword. In Origins of Life: The Central Concepts; Deamer, D.W., Ed.; Jones and Bartlett: Boston, MA, USA, 1994. [Google Scholar]
- Trifonov, E.N. Vocabulary of definitions of life suggests a definition. J. Biomol. Struct. Dyn. 2011, 29, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Lane, N.; Martin, W.F. The origin of membrane bioenergetics. Cell 2012, 151, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Adamala, K.; Szostak, J.W. Competition between model protocells driven by an encapsulated catalyst. Nat. Chem. 2013, 5, 495–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, T.F.; Szostak, J.W. Exploding vesicles. J. Syst. Chem. 2011, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Oró, J.; Kimball, A.P. Synthesis of purines under possible primitive earth conditions. I. Adenine from hydrogen cyanide. Arch. Biochem. Biophys. 1961, 94, 217–227. [Google Scholar] [CrossRef]
- Saladino, R.; Brucato, J.R.; De Sio, A.; Botta, G.; Pace, E.; Gambicorti, L. Photochemical synthesis of citric acid cycle intermediates based on titanium dioxide. Astrobiology 2011, 11, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Neri, V.; Crestini, C.; Costanzo, G.; Graciotti, M.; Di Mauro, E. Synthesis and degradation of nucleic acid components by formamide and iron sulfur minerals. J. Am. Chem. Soc. 2008, 130, 15512–15518. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Barontini, M.; Cossetti, C.; Di Mauro, E.; Crestini, C. The effects of borate minerals on the synthesis of nucleic acid bases, amino acids and biogenic carboxylic acids from formamide. Orig. Life Evol. Biosph. 2011, 41, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Neri, V.; Crestini, C.; Costanzo, G.; Graciotti, M.; Di Mauro, E. The role of the formamide/zirconia system in the synthesis of nucleobases and biogenic carboxylic acid derivatives. J. Mol. Evol. 2010, 71, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Bizzarri, B.M.; Botta, L.; Šponer, J.; Šponer, J.E.; Georgelin, T.; Jaber, M.; Rigaud, B.; Kapralov, M.; Timoshenko, G.N.; et al. Proton irradiation: A key to the challenge of N-glycosidic bond formation in a prebiotic context. Sci. Rep. 2017, 7, 14709. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, R.A.; Orgel, L.E. Studies in prebiotic synthesis. V. Synthesis and photoanomerization of pyrimidine nucleosides. J. Mol. Biol. 1970, 47, 531–543. [Google Scholar] [CrossRef]
- Powner, M.W.; Gerland, B.; Sutherland, J.D. Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions. Nature 2009, 459, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Powner, M.W.; Sutherland, J.D. Phosphate-mediated interconversion of ribo- and arabino-configured prebiotic nucleotide intermediates. Angew. Chem. Int. Ed. 2010, 49, 4641–4643. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Tsanakopoulou, M.; Magnani, C.J.; Szabla, R.; Šponer, J.E.; Šponer, J.; Góra, R.W.; Sutherland, J.D. A prebiotically plausible synthesis of pyrimidine β-ribonucleosides and their phosphate derivatives involving photoanomerization. Nat. Chem. 2017, 9, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Thoma, I.; Deutsch, A.; Gehrke, T.; Mayer, P.; Zipse, H.; Carell, T. A high-yielding, strictly regioselective prebiotic purine nucleoside formation pathway. Science 2016, 352, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.; Strazewski, P. Bringing prebiotic nucleosides and nucleotides down to Earth. Angew. Chem. Int. Ed. 2016, 55, 13930–13933. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Crestini, C.; Busiello, V.; Ciciriello, F.; Costanzo, G.; Di Mauro, E. Differential stability of 3′- and 5′-phosphoester bonds in deoxy monomers and oligomers. J. Biol. Chem. 2005, 280, 35658–35669. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Crestini, C.; Ciciriello, F.; Di Mauro, E.; Costanzo, G. Origin of informational polymers: Differential stability of phosphoester bonds in ribomonomers and ribooligomers. J. Biol. Chem. 2006, 281, 5790–5796. [Google Scholar] [CrossRef] [PubMed]
- Ciciriello, F.; Costanzo, G.; Pino, S.; Crestini, C.; Saladino, R.; Di Mauro, E. Molecular complexity favors the evolution of ribopolymers. Biochemistry 2008, 47, 2732–2742. [Google Scholar] [CrossRef] [PubMed]
- Georgiadis, M.M.; Singh, I.; Kellett, W.F.; Hoshika, S.; Benner, S.A.; Richards, N.G.J. Structural basis for a six nucleotide genetic alphabet. J. Am. Chem. Soc. 2015, 137, 6947–6955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benner, S.A. Understanding nucleic acids using synthetic chemistry. Acc. Chem. Res. 2004, 37, 784–797. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, P.E. Peptide nucleic acids and the origin of life. Chem. Biodivers. 2007, 4, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Eschenmoser, A. Towards a chemical etiology of nucleic acid structure. Orig. Life Evol. Biosph. 1997, 27, 535–553. [Google Scholar] [CrossRef] [PubMed]
- Westheimer, F.H. Why nature chose phosphates. Science 1987, 235, 1173–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, A.; Lohrmann, R.; Orgel, L.E. Phosphorylation with inorganic phosphates at moderate temperatures. Science 1967, 157, 952. [Google Scholar] [CrossRef] [PubMed]
- Lohrmann, R.; Orgel, L.E. Prebiotic synthesis: Phosphorylation in aqueous solution. Science 1968, 161, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Osterberg, R.; Orgel, L.E.; Lohrmann, R. Further studies of urea-catalyzed phosphorylation reactions. J. Mol. Evol. 1973, 2, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Lohrmann, R.; Orgel, L.E. Urea-inorganic phosphate mixtures as prebiotic phosphorylating agents. Science 1971, 171, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.W. Prebiotic phosphorus chemistry reconsidered. Orig. Life Evol. Biosph. 1997, 27, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, Y.; Watanabe, H.; Saitoh, M.; Namba, T. Volcanic production of polyphosphates and its relevance to prebiotic evolution. Nature 1991, 352, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Schoffstall, A.M. Prebiotic phosphorylation of nucleosides in formamide. Orig. Life 1976, 7, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Schoffstall, A.M.; Barto, R.; Ramos, D. Nucleoside and deoxynucleoside phosphorylation in formamide solutions. Orig. Life Evol. Biosph. 1982, 12, 143–151. [Google Scholar] [CrossRef]
- Schoffstall, A.M.; Laing, E. Phosphorylation mechanisms in chemical evolution. Orig. Life Evol. Biosph. 1985, 15, 141–150. [Google Scholar] [CrossRef]
- Schoffstall, A.M.; Mahone, S.M. Formate ester formation in amide solutions. Orig. Life Evol. Biosph. 1988, 18, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Saladino, R.; Crestini, C.; Ciciriello, F.; Di Mauro, E. Nucleoside phosphorylation by phosphate minerals. J. Biol. Chem. 2007, 282, 16729–16735. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Crestini, C.; Ciciriello, F.; Pino, S.; Costanzo, G.; Di Mauro, E. From formamide to RNA: The roles of formamide and water in the evolution of chemical information. Res. Microbiol. 2009, 160, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Burcar, B.; Pasek, M.; Gull, M.; Cafferty, B.J.; Velasco, F.; Hud, N.V.; Menor-Salván, C. Darwin’s warm little pond: A one-pot reaction for prebiotic phosphorylation and the mobilization of phosphate from minerals in a urea-based solvent. Angew. Chem. Int. Ed. 2016, 55, 13249–13253. [Google Scholar] [CrossRef] [PubMed]
- Gibard, C.; Bhowmik, S.; Karki, M.; Kim, E.-K.; Krishnamurthy, R. Phosphorylation, oligomerization and self-assembly in water under potential prebiotic conditions. Nat. Chem. 2017, 10, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Pasek, M.A.; Gull, M.; Herschy, B. Phosphorylation on the early Earth. Chem. Geol. 2017, 475, 149–170. [Google Scholar] [CrossRef]
- Hazen, R.M. Paleomineralogy of the Hadean Eon: A preliminary species list. Am. J. Sci. 2013, 313, 807–843. [Google Scholar] [CrossRef]
- Shapiro, R.; Kang, S. Uncatalyzed hydrolysis of deoxyuridine, thymidine, and 5-bromodeoxyuridine. Biochemistry 1969, 8, 1806–1810. [Google Scholar] [CrossRef] [PubMed]
- Garrett, E.R.; Mehta, P.J. Solvolysis of adenine nucleosides. II. Effects of sugars and adenine substituents on alkaline solvolyses. J. Am. Chem. Soc. 1972, 94, 8542–8547. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, T.; Karlstrom, O. Heat-induced depyrimidination of deoxyribonucleic acid in neutral solution. Biochemistry 1973, 12, 5151–5154. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, H.S.; Chargaff, E. Studies on the nucleoside arrangement in deoxyribonucleic acids. I. The relationship between the production of pyrimidine nucleoside 3′,5′-diphosphates and specific features of nucleotide sequence. Biochim. Biophys. Acta 1957, 26, 596–608. [Google Scholar] [CrossRef]
- Venner, H. Research on nucleic acids. XII. Stability of the N-glycoside bond of nucleotides. Hoppe Seylers Z. Physiol. Chem. 1966, 344, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, R.; Danzig, M. Acidic hydrolysis of pyrimidine deoxyribonucleotides. Biochim. Biophys. Acta 1973, 319, 5–10. [Google Scholar] [CrossRef]
- Lindahl, T.; Nyberg, B. Rate of depurination of native deoxyribonucleic acid. Biochemistry 1972, 11, 3610–3618. [Google Scholar] [CrossRef] [PubMed]
- Kochetov, N.K.; Budowski, E.L. (Eds.) Organic Chemistry of Nucleic Acids; Plenum Press: London, UK; New York, NY, USA, 1982; pp. 477–532. [Google Scholar]
- Verlander, M.S.; Lohrmann, R.; Orgel, L.E. Catalysts for self-polymerization of adenosine cyclic 2′,3′-phosphate. J. Mol. Evol. 1973, 2, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Verlander, M.S.; Orgel, L.E. Analysis of high molecular weight material from the polymerization of adenosine cyclic 2′,3′-phosphate. J. Mol. Evol. 1974, 3, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Usher, D.A.; McHale, A.H. Nonenzymic joining of oligoadenylates on a polyuridylic acid template. Science 1976, 192, 53–54. [Google Scholar] [CrossRef] [PubMed]
- Usher, D.A.; McHale, A.H. Hydrolytic stability of helical RNA—Selective advantage for natural 3′,5′-bond. Proc. Natl. Acad. Sci. USA 1976, 73, 1149–1153. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Pino, S.; Ciciriello, F.; Di Mauro, E. Generation of long RNA chains in water. J. Biol. Chem. 2009, 284, 33206–33216. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Saladino, R.; Botta, G.; Giorgi, A.; Scipioni, A.; Pino, S.; Di Mauro, E. Generation of RNA molecules by a base-catalysed click-like reaction. ChemBioChem 2012, 13, 999–1008. [Google Scholar] [CrossRef] [PubMed]
- Šponer, J.E.; Šponer, J.; Giorgi, A.; Di Mauro, E.; Pino, S.; Costanzo, G. Untemplated nonenzymatic polymerization of 3′,5′ cGMP: A plausible route to 3′,5′-linked oligonucleotides in primordia. J. Phys. Chem. B 2015, 119, 2979–2989. [Google Scholar] [CrossRef] [PubMed]
- Morasch, M.; Mast, C.B.; Langer, J.K.; Schilcher, P.; Braun, D. Dry polymerization of 3′,5′-cyclic GMP to long strands of RNA. ChemBioChem 2014, 15, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Pino, S.; Timperio, A.M.; Šponer, J.E.; Šponer, J.; Nováková, O.; Šedo, O.; Zdráhal, Z.; Di Mauro, E. Non-enzymatic oligomerization of 3′,5′ cyclic AMP. PLoS ONE 2016, 11, e0165723. [Google Scholar] [CrossRef] [PubMed]
- Costanzo, G.; Giorgi, A.; Scipioni, A.; Timperio, A.M.; Mancone, C.; Tripodi, M.; Kapralov, M.; Krasavin, E.; Kruse, H.; Šponer, J.; et al. Nonenzymatic oligomerization of 3′,5′-cyclic CMP induced by proton and UV irradiation hints at a nonfastidious origin of RNA. ChemBioChem 2017, 18, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Cassone, G.; Šponer, J.; Saija, F.; Di Mauro, E.; Saitta, A.M.; Šponer, J.E. Stability of 2′,3′ and 3′,5′ cyclic nucleotides in formamide and in water: A theoretical insight into the factors controlling the accumulation of nucleic acid building blocks in the prebiotic pool. Phys. Chem. Chem. Phys. 2017, 19, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Chwang, A.K.; Sundaralingam, M. The crystal and molecular structure of guanosine 3′,5′-cyclic monophosphate (cyclic GMP) sodium tetrahydrate. Acta Crystallogr. B 1974, 30, 1233–1240. [Google Scholar] [CrossRef]
- Kervio, E.; Sosson, M.; Richert, C. The effect of leaving groups on binding and reactivity in enzyme-free copying of DNA and RNA. Nucleic Acids Res. 2016, 44, 5504–5514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lohrmann, R. Formation of nucleoside 5′-phosphoramidates under potentially prebiological conditions. J. Mol. Evol. 1977, 10, 137–154. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Ferris, J.P. Kinetic and mechanistic analysis of dinucleotide and oligonucleotide formation from the 5′-phosphorimidazolide of adenosine on Na+-montmorillonite. J. Am. Chem. Soc. 1994, 116, 7564–7572. [Google Scholar] [CrossRef] [PubMed]
- Mansy, S.S.; Schrum, J.P.; Krishnamurthy, M.; Tobe, S.; Treco, D.A.; Szostak, J.W. Template-directed synthesis of a genetic polymer in a model protocell. Nature 2008, 454, 122–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orgel, L.E. Prebiotic chemistry and the origin of the RNA world. Crit. Rev. Biochem. Mol. Biol. 2004, 39, 99–123. [Google Scholar] [PubMed]
- Jauker, M.; Griesser, H.; Richert, C. Copying of RNA sequences without pre-activation. Angew. Chem. Int. Ed. 2015, 54, 14559–14563. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, L.; Maurel, M.C.; Deamer, D. Salt-promoted synthesis of RNA-like molecules in simulated hydrothermal conditions. J. Mol. Evol. 2015, 80, 86–97. [Google Scholar] [CrossRef] [PubMed]
- DeGuzman, V.; Vercoutere, W.; Shenasa, H.; Deamer, D. Generation of oligonucleotides under hydrothermal conditions by non-enzymatic polymerization. J. Mol. Evol. 2014, 78, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Šponer, J.E.; Šponer, J.; Di Mauro, E. Four ways to oligonucleotides without phosphoimidazolides. J. Mol. Evol. 2015, 82, 5–10. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saladino, R.; Šponer, J.E.; Šponer, J.; Costanzo, G.; Pino, S.; Di Mauro, E. Chemomimesis and Molecular Darwinism in Action: From Abiotic Generation of Nucleobases to Nucleosides and RNA. Life 2018, 8, 24. https://doi.org/10.3390/life8020024
Saladino R, Šponer JE, Šponer J, Costanzo G, Pino S, Di Mauro E. Chemomimesis and Molecular Darwinism in Action: From Abiotic Generation of Nucleobases to Nucleosides and RNA. Life. 2018; 8(2):24. https://doi.org/10.3390/life8020024
Chicago/Turabian StyleSaladino, Raffaele, Judit E. Šponer, Jiří Šponer, Giovanna Costanzo, Samanta Pino, and Ernesto Di Mauro. 2018. "Chemomimesis and Molecular Darwinism in Action: From Abiotic Generation of Nucleobases to Nucleosides and RNA" Life 8, no. 2: 24. https://doi.org/10.3390/life8020024