The Distinctive Regulation of Cyanobacterial Glutamine Synthetase


1
Helmholtz Centre for Environmental Research, Department of Solar Materials, Permoserstrasse 15, D-04318 Leipzig, Germany
2
Instituto de Bioquímica Vegetal y Fotosíntesis, CSIC-Universidad de Sevilla, Américo Vespucio 49, E-41092 Seville, Spain
*
Author to whom correspondence should be addressed.
Life 2018, 8(4), 52; https://doi.org/10.3390/life8040052
Submission received: 25 September 2018
/
Revised: 23 October 2018
/
Accepted: 25 October 2018
/
Published: 27 October 2018
Abstract
:Glutamine synthetase (GS) features prominently in bacterial nitrogen assimilation as it catalyzes the entry of bioavailable nitrogen in form of ammonium into cellular metabolism. The classic example, the comprehensively characterized GS of enterobacteria, is subject to exquisite regulation at multiple levels, among them gene expression regulation to control GS abundance, as well as feedback inhibition and covalent modifications to control enzyme activity. Intriguingly, the GS of the ecologically important clade of cyanobacteria features fundamentally different regulatory systems to those of most prokaryotes. These include the interaction with small proteins, the so-called inactivating factors (IFs) that inhibit GS linearly with their abundance. In addition to this protein interaction-based regulation of GS activity, cyanobacteria use alternative elements to control the synthesis of GS and IFs at the transcriptional level. Moreover, cyanobacteria evolved unique RNA-based regulatory mechanisms such as glutamine riboswitches to tightly tune IF abundance. In this review, we aim to outline the current knowledge on the distinctive features of the cyanobacterial GS encompassing the overall control of its activity, sensing the nitrogen status, transcriptional and post-transcriptional regulation, as well as strain-specific differences.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Nitrogen (N) is an essential element to all life on earth as it is a significant fraction of crucial biomolecules such as amino acids, nucleic acids and a myriad of derivatives. Despite being the most common pure element in the atmosphere, molecular dinitrogen (N2) can only be utilized by few archaea and bacteria that possess N2-fixing nitrogenase (EC 1.18.6.1). This enzyme reduces the inert triple bond of N2 by providing enormous reducing power to yield bioavailable, dissolved inorganic N in the form of ammonium, which can be taken up and assimilated into organic forms by most microorganisms [1]. Thus bioavailable N species mostly remain scarce and are subject to strong fluctuations in natural habitats rendering N availability a key environmental factor for growth [2,3]. Therefore, it is not surprising that most microorganisms frugally husband their N pools and strongly regulate uptake and assimilation machineries in response to environmental changes.
1.1. GS Catalyzes a Core Reaction of N Assimilation in Bacteria
In E. coli and most other Gram-negatives, two primary pathways of ammonia assimilation have been described [4,5] (Figure 1). The initially discovered NAD(P)H/H+ glutamate dehydrogenase (GDH, EC 1.4.1.3) catalyzes the reductive, NAD(P)H/H+-dependent amination of 2-oxoglutarate (2OG) to yield glutamate and NAD(P)+, and was regarded as the main enzyme of ammonium assimilation in bacteria for a certain period [6]. Nevertheless, in 1970, Tempest and co-workers established another main N assimilatory system in bacteria, which comprises the subsequent action of the enzymes glutamine synthetase (GS; glutamate-ammonia ligase, EC 6.3.1.2) and glutamate synthase (formerly glutamine:2-oxoglutarate-aminotransferase (GOGAT), EC 1.4.1.14 (NAD(P)H+-dependent), and EC 1.4.7.1 (ferredoxin-dependent)). In the GS/GOGAT cycle GS facilitates the first step of N assimilation, the conversion of inorganic into organic N by incorporating ammonium into glutamate and yielding glutamine (Figure 1). Subsequently, with either NADPH/H+ or ferredoxin as reducing equivalents, GOGAT transfers the amine residue of glutamine to 2OG and forms two molecules of glutamate, which again serve as GS substrate [7,8]. Due to a greater affinity to ammonium and the dependence on ATP, the GS/GOGAT cycle is crucial for balancing glutamine pools and N-assimilation in enterobacteria and most Gram-negatives, i.e., during combined low ammonium availability and energy excess. In contrast, the GDH pathway is employed for glutamate synthesis upon energy limitation and ammonium profusion since the catalyzed reaction does not require ATP, while the enzyme features relatively high Km values for ammonium [5].
The family of GSs constitutes one of the oldest existing and operational gene families [9]. GSs are found in all living organisms and are split into at least three miscellaneous types (GSI, GSII, and GSIII), which differ in subunit stoichiometry and molecular weight [10,11,12,13]. Interestingly, studies which revealed the wide dispersion of GS aroused the suspicion that the GS types evolved ahead of the divergence of eukaryotes and prokaryotes by early gene duplication [9,11,14,15]. While GSII represents the generic eukaryotic enzyme which, however, also occurs in representatives of the Franciaceae, Rhizobiaceae (both plant symbionts), and Streptomycetaceae families [16,17,18], GSI is restricted to bacteria and archaea [11]. The third GS type, GSIII is generally found in a myriad of bacteria but several eukaryotic representatives (e.g., members of fungi, viridiplantae, amoebozoa, heterokonts, and haptophyta) as well as archaea appear to also bear GSIII-encoding genes (see Pfam entry PF12437).
1.2. GS Regulation Is Diverse among Distinct Organisms
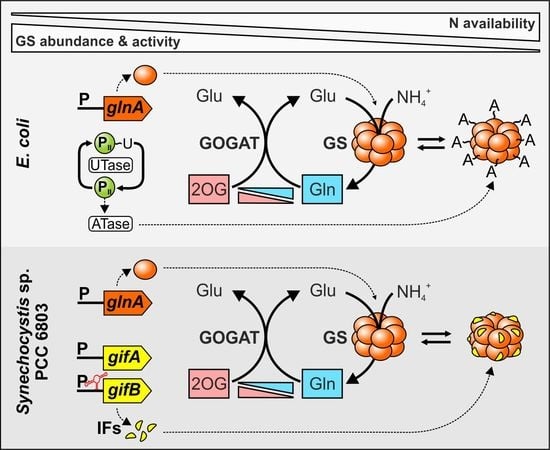
GSI (hereafter referred to as GS) consists of two superimposed hexameric rings, which are arranged in a centrosymmetric structure. The GS of enterobacteria is one of the best investigated and most comprehensively regulated enzymes known and represents the prevailing paradigm of bacterial GS regulation, which is addressed in most scientific textbooks. Enterobacterial GS is subject to feedback inhibition by several molecules of N and energy metabolism, which include among others tryptophan, histidine, carbamoyl phosphate, CTP, AMP, glucosamine-6-phosphate, NAD+, serine, and alanine, as well as glycine. These intermediates orchestrate a gradual inhibition of GS that mounts depending on the number of bound inhibitors, which is designated cumulative feedback inhibition [19,20,21]. The amino acids serine, alanine, and glycine were shown to interact with the glutamine binding site [22] and AMP appears to sequester the ATP binding cleft [23] while the remaining metabolites occupy distinct allosteric sites at the enzyme [20].
In addition, the enterobacterial GS is subject to posttranslational regulation by reversible adenylylation of specific tyrosine residues in each of the 12 enzyme subunits, which in turn raises the sensitivity of GS for feedback inhibition. The corresponding regulatory circuits, which govern the modification state of GS, are complex and involve two connected cycles of reversible protein nucleotidylation (Figure 2). In the first cycle, the ubiquitous signal transducing protein PII switches between uridylylated and unmodified states [24,25], while in the second cycle GS changes between adenylylated and unmodified forms [26,27]. Uridylylation of PII is catalyzed by the bifunctional uridylyltransferase (UTase) and depends on 2OG [24,25,28,29]. On the contrary, removal of the uridylyl moiety of PII by UTase requires glutamine [30]. In an analogical process, adenylylation of GS by the bifunctional adenylyltransferase (ATase) is a glutamine-dependent reaction that is opposed by the 2OG-dependent deadenylylation of GS [26,31,32,33,34].
In general, sensing of the cellular C/N status is of utmost importance for regulating N uptake and assimilation to ensure the maintenance of metabolic homeostasis. Since 2OG provides the carbon skeleton for the amidation reaction in the GS/GOGAT cycle but also participates in the tricarboxylic acid (TCA) cycle, it constitutes a branching point between carbon (C) and N metabolism, and is subject to fluctuations according to the cellular N status. Hence, 2OG is an indicator of C as well as N availability and its level accumulates in response to N limitation [35]. Moreover, because glutamine is the product of GS it is also suitable to reflect the external availability of N to the cell in addition to 2OG [35].
A tight connection of the PII and GS nucleotidylation cycles is ensured by the modification state of PII: adenylylation, and thus inactivation of GS by ATase strictly depends on the binding of unmodified PII, which accumulates during N-excess and high glutamine levels. On the other hand, deadenylylation and thus activation of GS by ATase only occurs upon binding of covalently modified PII(UMP), which accumulates during N limitation promoted by 2OG [25,29,36]. Hence, this bicyclic regulatory mechanism of enterobacteria constitutes a reciprocal control of the ATase- and UTase-catalyzed reactions by indicators of the cellular N status, 2OG and glutamine, to fine-tune GS activity accordingly (Figure 2) [28,37].
A third regulatory entity of enterobacterial GS is the widespread Ntr two-component system comprising the sensory histidine kinase NtrB and its response regulator NtrC. This system controls the N-dependent expression of the glnA gene, which encodes the GS monomers (Figure 2). The ATP-dependent autophosphorylation of NtrB yields active NtrC by transmission of the phosphoryl moiety. Subsequently, active NtrC promotes transcription of glnA, as well as other N-assimilatory genes, which also feature σ54-dependent promoters [40]. This process is antagonized by the emergence of unmodified PII protein, which triggers dephosphorylation and thus inactivation of NtrC (reviewed by [41]). In summary, UTase probes the cellular N status via both glutamine and 2OG concentrations thereby influencing the levels of unmodified and modified PII protein. This modification sensitizes or desensitizes GS for feedback inhibition by adenylylation/deadenylylation of the enzyme while the NtrB-NtrC system governs transcriptional control and therefore GS synthesis with respect to the cellular C/N balance (for a review see [39]).
GS regulation in enterobacteria appears relatively uniform, though investigation of an increasing number of bacteria beyond the enterobacterial family revealed striking differences in GS regulation among virtually every family investigated during the last two decades. Nevertheless, the assimilatory enzymes and several regulatory proteins involved appear to be mostly conserved in prokaryotes. For instance, the bifunctional GS of low G+C Gram-positive bacteria like Bacillus subtilis, which functions as both enzyme and regulator, is subject to feedback inhibition but devoid of posttranscriptional control [42]. During N-rich conditions, GS directly interacts with the transcriptional regulator of N assimilatory genes, TrnA and GlnK, an N-sensing PII-like protein. TrnA binding to GS triggers partial inhibition of enzymatic activity and ameliorates the impact of feedback inhibitors, especially of glutamine. In addition, GS binding obstructs DNA binding of TrnA. Under N-limiting conditions, GlnK forestalls interaction of TrnA with active GS, rendering the enzyme fully active [42,43]. In Clostridium saccharobutylicum NCP262, an endospore-forming obligate anaerobe, regulation of ammonium assimilation underlies a yet not fully understood mechanism encompassing post-transcriptional regulation of core enzymes via antisense RNAs as well as a specific RNA-binding regulatory protein of the ANTAR family, termed NitR [44], which is capable of transcription antitermination and thus induces expression of GS/GOGAT cycle genes upon N limitation [45]. Another mechanism of GS regulation was observed in several archaea where protein-protein interaction of GS with the PII-like protein GlnK was shown to increase GS activity in the presence of 2OG, while low 2OG levels thereof triggered GS inhibition by GlnK [46,47]. Taken together, these examples illustrate the complex and unique regulatory mechanisms of GS in diverse prokaryotes. This review aims to complement the body of knowledge on the diversified GS regulation throughout the prokaryotic domain by summarizing the insights gained into the exceptional mechanisms underlying GS regulation in cyanobacteria.
2. Cyanobacteria Evolved Exceptional GS Regulatory Mechanisms
As the evolutionary ancestors of chloroplasts, cyanobacteria represent a very ancient lineage and are the only bacterial group capable of performing water-splitting, oxygenic photosynthesis by combining the reductive power of two photosystems. This phylum shows substantial genomic and morphological divergence with unicellular and multicellular lifestyles, the latter showing a high degree of organization and differentiation. Cyanobacteria can be encountered in nearly any light-exposed habitat ranging from arctic regions, desert soils, rock surfaces, hot wells, and hypersaline lakes to fresh-water ecosystems, as well as ultraoligotrophic marine realms [48,49,50,51,52] and even non-light exposed habitats such as the deep terrestrial biosphere [53]. Cyanobacteria are utterly important in ecological terms and fundamentally affect global macronutrient cycles, as marine Prochlorococcus strains, for instance, account for 25% of the ocean’s net primary production [54]. Considering their metabolic performances and sheer abundance, cyanobacteria are prone to deplete nutrients in their vicinity, which renders nutrient deficiency a common factor for growth retardation. This is exemplified by the photosynthetic performance of early cyanobacteria, which caused the release of vast amounts of the photosynthetic byproduct O2 and simultaneous depletion of the majority of atmospheric carbon dioxide, which fundamentally affected living conditions on earth and paved the way for the evolution of aerobic metabolism and complex life [55].
For cyanobacteria, N represents a vital nutrient and accounts for up to 10% of their dry weight [56]. This is also illustrated by the fact that non-diazotrophic cyanobacteria, incapable of N2 fixation, tightly regulate their N assimilatory machinery and undergo substantial adaptations with respect to N availability [57]. For instance, prolonged N starvation eventually can result in degradation of the photosynthetic apparatus in a process designated as chlorosis [58]. Cyanobacteria are capable of assimilating dissolved N in form of ammonium, nitrate, and nitrite, and some can utilize urea, cyanate as well as several amino acids [56]. Notwithstanding, ammonium is the N species, which is directly incorporated by the GS/GOGAT cycle while other combined N sources require reduction prior to incorporation. Therefore, ammonium is the favored, least energy-demanding N source for most cyanobacteria [57]. Shortly after the discovery of GS in enterobacteria [7], the occurrence of GS/GOGAT and GDH was confirmed in the clade of cyanobacteria [59,60,61] and subsequent 13N-labeling experiments unequivocally established GS/GOGAT as the main N assimilatory pathway [62,63,64]. Apparently, GDH plays a rather auxiliary role in cyanobacterial N assimilation and imparts selective advantages in late stages of growth [65].
The cyanobacterial GS shares substantial similarities with the GSI of other bacteria, illustrated by the fact that it is able to complement a glnA-deficient E. coli mutant [66]. Nevertheless, the systems, which sense the N status and control GS in cyanobacteria, lack several known features characteristic for other, well-investigated bacterial clades. Purification and characterization of the GS from different cyanobacteria revealed a similar structure and subunit composition but, most strikingly, lacked covalent modifications characteristic of the enterobacterial GS [67,68,69]. Moreover, when compared to the enzyme of E. coli, GS of various cyanobacteria showed lower Km values and a higher sensitivity towards common GS inhibitors such as a variety of amino acids [68]. Furthermore, cyanobacterial GS features a marked specificity for ATP and is greatly inhibited by AMP. Thus, GS may respond more vigorously to differences in energy charge and adenine nucleotide availability, which are known to fluctuate in response to environmental alterations in cyanobacteria [69,70].
2.1. Sensing of the Cellular N Status and Transcriptional regulation of the glnA Gene in Cyanobacteria
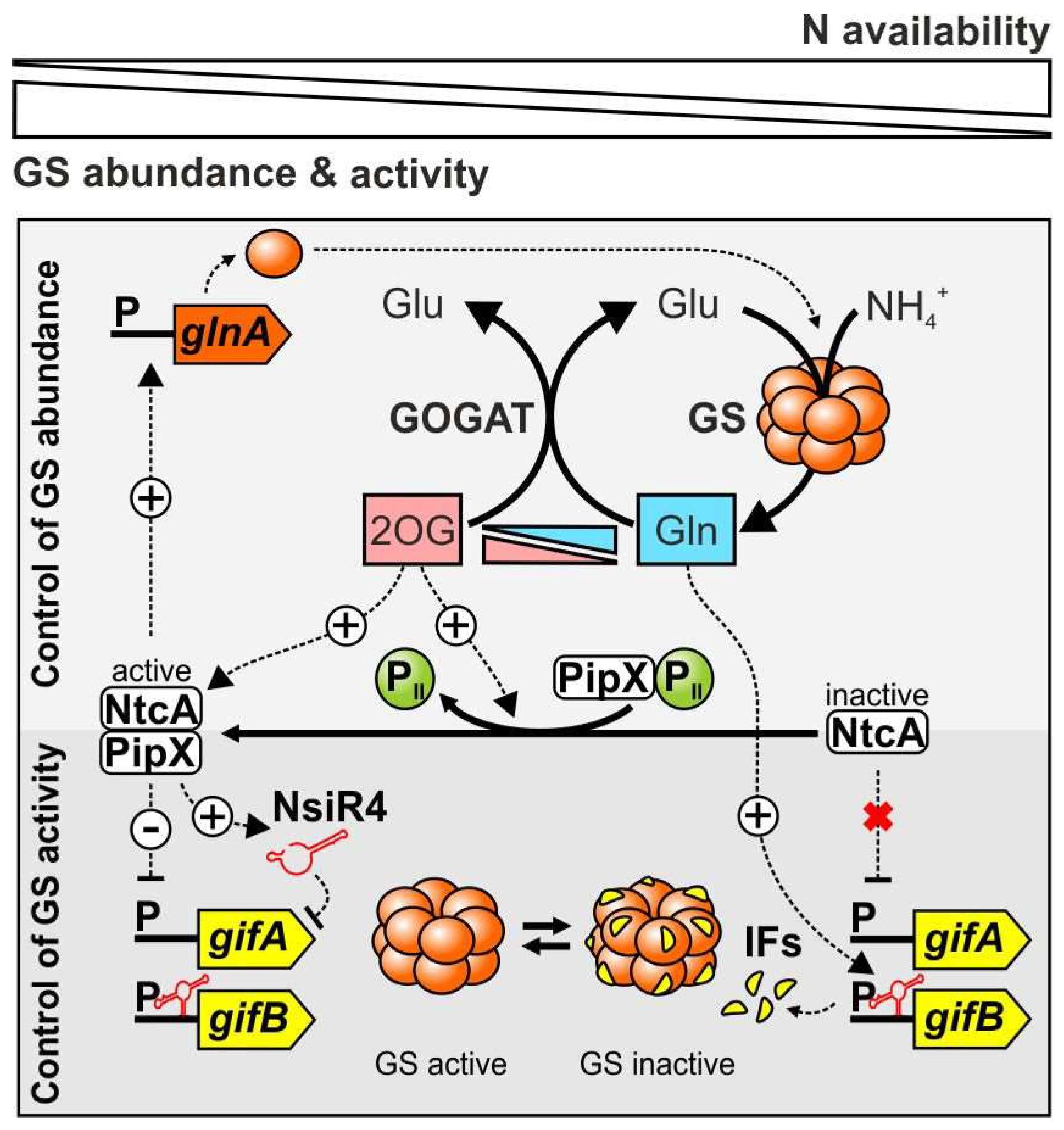
In cyanobacteria, the expression of the glnA gene is regulated with respect to the nature and abundance of the N source. N excess, e.g., induced by ammonium supplementation, results in low glnA expression, while cells grown on nitrate as sole N source feature elevated glnA levels only exceeded by N-starved cells [71,72,73,74,75]. Remarkably, the thoroughly investigated Ntr two-component system, which controls transcription of glnA in a wide range of bacteria, is absent in cyanobacteria [41,66]. Instead, the transcription of genes involved in N uptake and assimilation is mainly controlled by the highly conserved global transcriptional regulator NtcA, a protein of the CRP (the cyclic AMP receptor protein, also known as catabolite repressor protein) regulator family [76,77]. NtcA is restricted to cyanobacteria and can act both as an activator or repressor, depending on the location of its binding site with respect to the transcriptional start site (TSS) of the regulated genes [57]. Under N limitation, NtcA mediates the simultaneous up-regulation of multiple genes for N uptake systems (e.g., amt1 encoding an ammonium transporter) and other N-related genes including glnA as well as ntcA itself [57,78,79,80].
NtcA-dependent transcriptional regulation includes the interplay of the ubiquitous signal transduction protein PII with the co-activator protein PipX, which modulate NtcA activity with respect to the cellular N status [81,82,83]. The regulatory circuit focusing on the glnA gene is summarized in Figure 3. Despite being crucial for N sensing in other bacterial groups, preliminary findings suggested that cyanobacteria do not perceive the cellular N status by probing glutamine levels [2]. Instead, 2OG was shown to be the metabolite, which governs NtcA activity and thus sensing of the N status in cyanobacteria [84]. As an indicator of cellular C/N balance, 2OG directly modulates the binding affinity of the homodimeric NtcA to its target promoters [84,85,86], which contain the NtcA recognition sequence GTA-N8-TAC [87,88,89]. Moreover, the small co-activator PipX is capable of coalescing in alternate complexes with either NtcA or the PII protein [81]. Emergence of respective complexes is mutually exclusive, as high 2OG levels caused by N deprivation result in PipX-NtcA complex formation and up-regulation of N assimilatory genes including glnA (Figure 3) while PipX-PII complex formation is prevented. On the other hand, low 2OG levels during N excess initializes PipX-PII complex formation, which prevents PipX from boosting NtcA-mediated transcriptional regulation and significantly lowers NtcA binding affinities under these conditions [81,90]. Thus, this mechanism provides a functional connection of PII-mediated N signaling and NtcA-dependent gene regulation, which is unique to cyanobacteria and ensures proper expression of glnA and other N-related genes with respect to metabolic demands and N supply (Figure 3). Nevertheless, it should be noted that the interaction of PipX with PII is not only tuned by changes in 2OG levels, but also by changes in ADP. ADP strongly enhances the affinity of PII for PipX, and thus, increasing ADP levels allow PII-PipX complex formation even in the presence of 2OG [91].
2.2. Cyanobacterial GS Is Inactivated by Interaction with Small Proteins
In the early 1990s, GS in cyanobacteria was shown to be subject to a pronounced short-term inactivation upon addition of ammonium to nitrate grown cells [92]. Nevertheless, in contrast to the sophisticated adenylylation/deadenylylation system of enterobacteria, cyanobacterial GS was neither found adenylylated nor otherwise covalently modified [66]. Shortly after, it was shown that cyanobacterial GS, inactivated by ammonium supplementation in vivo, was reactivated in vitro by increasing pH, ionic strength or treatment with phosphatases, raising the possibility of non-covalent binding of any compound as the cause of GS inactivation [93]. Subsequent cross-linking experiments and mobility shift essays endorsed conclusions that rather polypeptides and not metabolites are involved in the observed inactivation [94]. Eventually, two homologous small proteins, namely the GS inactivating factors 7 and 17 (IF7 and IF17, respectively) were discovered due to the fact that two polypeptides of 7 and 17 kDa co-purified with ammonium-inactivated GS of the model strain Synechocystis sp. PCC 6803 (hereafter referred to as Synechocystis) [95]. These two proteins are encoded by the two separate genes gifA and gifB (glutamine synthetase inactivating factor A and B) with IF7 bearing remarkable resemblance to the C-terminal moiety of IF17 [95]. The regulatory interplay between GS and its IFs is illustrated in Figure 3.
In vitro analysis of heterologically expressed IF7, IF17, and GS of Synechocystis unequivocally demonstrated that both factors act independently from each other and do not require additional modifications to exert a concentration-dependent inactivation of GS [95]. Remarkably, in contrast to the enterobacterial adenylylation system, GS inhibition by small proteins in cyanobacteria renders the enzyme entirely inactive [95]. Generation of ΔgifA and ΔgifB single as well as a ΔgifAgifB double knockout strains rendered GS partially or fully active when grown with ammonium, demonstrating that the proteins encoded by these genes are exclusively responsible for GS inactivation and thus constitute a major layer of GS regulation in cyanobacteria [95].
2.3. The Biochemical Mechanism of GS Inactivation by Protein-Protein Interaction
In light of the reactivation of ammonium-inactivated GS by increasing pH values [93] and the high amount of positively charged residues in both IFs supported the idea of an electrostatic interaction with GS, likely on the same, negatively charged site of the enzyme [95]. In fact, three conserved arginine residues were found to be essential and equally important for the interaction of the Synechocystis IF7 with GS both in vivo and in vitro [96]. Substitution of one of these residues by either neutral (alanine) or negatively charged residues (glutamic acid) eradicated any interaction of IF7 with GS and rendered the enzyme fully active, thus indicating that loss of one junction between both proteins impedes any interaction [96]. In contrast, IF17 also features three conserved R residues, however substitution of these had a less pronounced effect on GS inhibition. Intriguingly, the importance of these three residues for IF17 function appears to be disparate as, for instance, substitution of R110, which is situated in the C-terminal moiety of IF17, the homologous region to IF7, had the most significant effect on protein function, while only exchange of all three R residues featured a fully active GS [96].
The stability of both GS inactivating factors is promoted by the interaction with GS in vivo [97]. Nevertheless, the IFs from Synechocystis vary in several aspects. IF17 is subject to proteolytic degradation stimulated by the absence of ammonium [97]. In contrast, stability of IF7 is not affected by N availability. Nevertheless, the IF7 protein appears to be degraded by the constitutively expressed Prp1/Prp2 metalloprotease, while the contribution of further proteases was suggested [97]. Interestingly, rapid IF17 degradation, which occurs in vivo upon ammonium removal, could not be observed in vitro; thus, additional factors must govern the N-dependent degradation of IF17, independent of gifB transcription. The mechanisms involved in the different IF17 degradation pattern, however, were not identified but may include the 82-residue N-terminal moiety, which is not present in IF7 [95,97]. This N-terminal part clearly contributes to protein stability: IF17 devoid of its N-terminus was outright susceptible to degradation while addition of the IF17 N-terminus to IF7 conveyed enhanced stability of that protein [96]. Even though the interaction of both IFs with GS mainly involves the same amino acids, IF17 has a stronger inhibitory effect on GS compared to IF7 [95]. This is explained by two distinct IF17 properties: the greater binding affinity to GS, which is associated with a lysine residue (K102) that contributes to the function of IF17 as well as its N-terminal region, which seems to improve GS inactivation [96].
Recently, analysis of chimeric enzymes of Synechocystis and the filamentous, dinitrogen-fixing strain Anabaena sp. PCC 7120 (hereafter referred to as Anabaena) suggested a possible coevolution of cyanobacterial GS and their corresponding IFs [98]. Each enzyme features two residues in the C-terminal moiety, which are crucial for specific interaction with their respective inactivating factors. Intriguingly, no conservation of these critical residues in the C-termini of other cyanobacterial GSs was observed, irrespective of the number of IFs possessed by these cyanobacteria, thus strongly suggesting a coevolution of GSs and IFs [98]. Furthermore, investigation of the GS-IF complex highlighted several possible mechanisms for protein-mediated GS inactivation [98]. Complex formation could cause alteration of the GS quarternary structure, which would result in perturbation of the active site and thus elimination of enzymatic activity. Another possibility is the contribution of enzymatically relevant residues in the interaction with IFs. Last, peripheral binding of IFs alongside the GS surface could impede the passage of substrates or products to or from the active site [98].
2.4. GS Inactivating Factors Are Common in Cyanobacteria and Transcriptionally Regulated by NtcA
Transcriptional analyses in Synechocystis revealed that expression of gifA and gifB was strongly increased upon ammonium addition while diminished expression of both genes was observed under N deficiency [99]. Therefore, gifA and gifB transcript accumulation is in accordance with the observed inactivation of GS in the presence of ammonium. Consequently, sequence analysis of the upstream regions of the gifA and gifB genes revealed potential binding sites for the global N transcriptional regulator NtcA, which center at the core promoter elements at −30.5 and −7.5 bp, respectively, thereby potentially mediating repression of the downstream genes. This assumption was confirmed as a non-segregated NtcA mutant featured constitutive expression of gifA and gifB, independent from the N source [99]. Upon N-limiting conditions, gifA and gifB transcription is repressed by NtcA binding and therefore inversely regulated to the GS structural gene glnA, which is activated by NtcA simultaneously [99]. Thus, NtcA and the GS IFs operate on transcriptional and posttranslational levels to adjust GS activity according to N availability (Figure 3).
Derived from the IF7/IF17 system of Synechocystis in which GS inactivating factors were discovered first, cyanobacterial GS inactivating factors can be subsumed into IF7-like and IF17-like classes with the exception of genome-streamlined marine picocyanobacteria like Prochlorococcus, which appear to lack full-length homologs [96]. Most cyanobacterial strains bear IF7-like inactivating factors [100] but other configurations can occur. For instance, Thermosynechococcus elongatus features two proteins homologous to IF17 while Anabaena harbors only one IF7-like inactivating factor (IF7A). Functional characterization of the IF7/gifA homolog of Anabaena confirmed that GS is also subject to IF-mediated inhibition in this cyanobacterium [101]. Nevertheless, the regulatory characteristics differ in several aspects from the system described in Synechocystis. Expression of the IF7A-encoding gifA gene is also regulated by the global N transcriptional regulator NtcA in Anabaena but in contrast to gif promoters in Synechocystis, the upstream region harbors two NtcA binding sites. One site is centered at the −28.5 position, indicating a repressive role of NtcA for this promoter as the binding site features the very same distance to the −10 box in the NtcA repressed gifA promoter of Synechocystis [101]. Interestingly, the other site centers at −77.5 with respect to the TSS [101]. Despite the fact that binding motifs in canonical NtcA-activated promoters center at the −40.5 position, NtcA binding to the second site centering at position −77.5 is thought to activate gifA gene expression as several NtcA-activated promoters were shown to feature motifs upstream of −41.5 [99,100]. The simultaneous activation and repression of gifA expression upon NtcA binding in Anabaena is conflicting and the biological meaning is not understood. Nevertheless, gifA expression in Anabaena is strongly increased upon addition of NH4+, similar to the situation in Synechocystis [101]. In addition, another interesting finding of the latter study was the co-occurrence of a point mutation within the gifA gene in the ntcA mutant strain CSE2. This mutation leads to a premature stop codon, preventing synthesis of IF7A. Complementation of a WT gifA gene in the ntcA mutant strain resulted in poor growth, which points towards IF7A toxicity if gifA is not repressed by NtcA und thus de-regulated. Hence, the existence of two independent IFs in Synechocystis might explain why there is no fully segregated ntcA knockout mutant available for that strain thus far because single mutations, e.g., in gifA would not compensate the steady inactivation of GS.
In general, it can be stated that inactivation of GS is more efficient in strains, which possess two IF homologs. In Synechocystis for instance, the joint action of IF7 and IF17 during N-rich conditions rapidly inhibits GS activity and thereby, quickly restore the intracellular 2OG levels, which leads to repression of the gif genes once again. On the contrary, in strains harboring solitary IFs such as Anabaena this process is less effective, thus a rather continuous expression of gif genes is required to maintain appropriate levels of GS activity [95,101].
2.5. A Small Regulatory RNA Interacts with the gifA mRNA and Interferes with IF7 Production
In addition to transcriptional regulators, bacteria possess numerous and diverse means of RNA-mediated gene regulation. These regulatory RNA elements do not encode proteins and thus are non-coding. A major group of those non-coding RNAs (ncRNAs) can activate or repress gene expression at the post-transcriptional level by complementary base pairing with mRNAs. Those small regulatory RNAs (sRNAs) show short, imperfect base pairing interactions but frequently overlap with sequences required for translation initiation and hence, contribute to the specific and customized synthesis of the respective proteins [101,102,103,104].
In recent years, biocomputational predictions and studies mapping TSSs on a genome-wide scale gave rise to a plethora of sRNAs in cyanobacteria [89,105,106,107,108]. The expression of several sRNAs was vigorously enhanced upon N-limiting conditions indicating that they may act as regulatory elements in cyanobacterial N metabolism. For instance, three sRNAs upregulated during N depletion were identified in Anabaena and named N stress-induced RNAs (NsiR1, NsiR2, and NsiR3). Nevertheless, their detailed functions are yet to be elucidated [89,109,110]. In Synechocystis, the sRNA NsiR4 was verified as one of the most strongly induced transcripts during N-limiting conditions [106]. Albeit absent in α-cyanobacteria, NsiR4 is conserved among distantly related cyanobacteria as homologs were found in at least 38 cyanobacterial genomes covering all five morphological subsections [111]. These findings provided a strong hint that NsiR4 indeed possesses a biological function. Interestingly, examination of genomes and transcriptomes of different cyanobacteria including Anabaena [89], Synechoccoccus sp. PCC 7002 [112] and Synechocystis sp. PCC 6714 [113] revealed the existence of two distinct NsiR4 forms, which differ in length and appear to be mutually exclusive as no co-occurrence could be observed. Nevertheless, in all cases a perfect NtcA binding motif was found upstream of NsiR4 homologs centering around −41.5 bp with respect to the TSS. Accordingly, NsiR4 expression is activated upon N depletion, which was experimentally verified in Synechocystis and Anabaena [111].
Intriguingly, the gifA gene was computationally predicted as the most promising NsiR4 target and the direct interaction of both RNAs was experimentally verified [111]. Consistent with previously examined sRNAs, which diminish target stability upon binding [114] knockout of NsiR4 led to an increased gifA mRNA abundance while NsiR4 overexpression decreased gifA transcript abundance compared to the WT, i.e., after adding ammonium. Subsequent analysis clearly demonstrated that the interaction of NsiR4 with the gifA transcript negatively affects IF7 protein synthesis and in turn also impacts GS activity [111]. The transcriptional regulation of gifA by NtcA is obviously not sufficient to control GS activity and hence, an additional post-transcriptional mechanism has evolved that helps to determine whether or not IF7 is produced (Figure 3). With this, NsiR4 was the first sRNA shown to regulate the assimilation of a macronutrient in bacteria directly as it is involved in controlling the entry of inorganic N into cellular metabolism. Very recently, also a nitrogen stress repressed RNA (NsrR1) has been identified in heterocyst-forming cyanobacteria which regulates, in a coherent feed-forward loop together with NtcA, a protein involved in phycobilisome degradation under N limitation [115]. Nevertheless, RNA-based N regulation is not restricted to cyanobacteria as a diverse range of prokaryotes meanwhile was shown to possess sRNAs that control N metabolism (reviewed by [116]).
2.6. A Glutamine Riboswitch in the gifB mRNA Regulates IF17 Synthesis
Yet another type of ncRNAs are riboswitches, which fulfill integral regulatory tasks in bacteria [117,118,119]. Riboswitches reside in the 5’UTR of mRNAs and can modulate the expression of downstream genes by ligand-induced conformational changes. Riboswitches are composed of an aptamer, which specifically binds the ligand (e.g., metabolites, inorganic ions), and an expression platform, which determines the read out of genetic information by affecting transcription, translation or RNA processing [120,121,122,123]. In 2010, a myriad of highly structured ncRNAs with potential biological functions were determined computationally by examining genome and metagenome databases [124]. Two classes of RNA motifs, related in both structure and sequence, were shown to exclusively occur in cyanobacterial genomes and marine metagenomes of environmental samples. Several of these structured RNAs were found upstream of genes encoding for proteins pivotal for N regulation, such as the N regulatory protein PII, GS, and ammonium transporters, thus suggesting a potential regulatory role in N metabolism [124]. These conserved sequence elements were termed glnA and downstream-peptide (DP) motifs and annotated as RF01739 and RF01704 in the Rfam database [125], respectively. Intriguingly, both the glnA motif of Synechococcus elongatus as well as the DP motif of Synechococcus sp. CC9902 were shown to alter their secondary structure upon selective binding of l-glutamine in vitro, which classified them as glutamine-binding aptamers [126]. For the first time, this study acknowledged the possibility that glutamine might be also involved in perceiving the N status in cyanobacteria. Cyanobacteria were believed to lack glutamine signaling because homologs of the known, glutamine sensing enzymes were not found in cyanobacterial genomes [2].
Recently, detailed analysis revealed a widespread occurrence of the glnA aptamer in cyanobacterial strains. The DP aptamer, however, is apparently restricted to the monophyletic group of marine picocyanobacteria where it frequently exists in multiple copies [127]. Within the available cyanobacterial genome sequences the aptamers were mainly found to be associated with genes encoding for proteins, which harbor the domain of unknown function 4278 (DUF4278). Other associations such as with genes for GS, PII, and ammonium transporters as reported previously [124] appear to be restricted to sequences of environmental samples and hence yet uncultured strains. Interestingly, DUF4278 covers the N-terminal moiety of the GS inactivating factor IF17 encoded by gifB. This suggested that IF17 synthesis might be also controlled by a ncRNA, namely by a glutamine riboswitch. In Synechocystis it was experimentally proven that the glutamine binding aptamer present in the 5’UTR of gifB undergoes structural modulations, which in turn impact the production of IF17 [127]. These results unequivocally demonstrated that the described glnA aptamer represents the metabolite-sensing domain of a glutamine-binding riboswitch, thus renaming to type 1 glutamine riboswitch was proposed. Investigation of a DP aptamer of a Prochlorococcus strain revealed that these elements are also part of functional glutamine riboswitches and activate gene expression upon glutamine binding similar to the type 1 glutamine riboswitch. Hence, for those containing the DP aptamer the designation type 2 glutamine riboswitch was suggested [127].
The glutamine riboswitches are restricted but common to cyanobacteria and their discovery added an important piece in the puzzle of glutamine synthetase regulation in this bacterial group (Figure 3). Previously, 2OG-dependent transcriptional regulation of gifA and gifB by NtcA was considered to be exclusively responsible for the control of GS activity in cyanobacteria [84,99]. Nevertheless, the riboswitches were proven to be required for sufficient GS regulation as well and are thus additional key elements that act in a glutamine-dependent manner [127]. Hence, cyanobacteria also use glutamine as a signaling molecule for perceiving the N status, however via a unique RNA-based sensing mechanism.
2.7. The Regulation of GS in Marine Picocyanobacteria is Unclear
Marine picocyanobacteria constitute a monophyletic group within the cyanobacterial realm, which displays several remarkable traits. One representative that attained the reputation of a model organism for marine ecology is the genus Prochlorococcus [128]. Prochlorococcus diverge in genetically distinct ecotypes along the water column, mainly caused by the aptitude to cope with different light intensities resulting in a tailor-made outfit of photosynthetic pigmentation for the given light conditions [129]. While those strains possess the smallest genomes and cell sizes of any photosynthetic organism [130,131] it is the most abundant organism in the ocean and supposedly on earth thus being of fundamental ecological importance for global nutrient cycles [54]. The extremely nutrient-deficient niches occupied by these organisms prompted several outstanding adaptions: vast genome streamlining [132,133] accompanied by the substitution of protein regulators with ncRNAs [134] and a low GC content [132]. N limitation imposes rigorous constraints on the boundaries of biomass accumulation and primary productivity in marine ultraoligotrophic habitats [135]. Thus, N cost-minimizing measures such as the preference for amino acids reduced in N content [136] and the augmented usage of internal TSSs upon N limitation, resulting in shortened proteins with abated N content [137] allow Prochlorococcus strains to flourish in N-poor environments of the open ocean.
Despite the cost-minimizing loss of proteinaceous regulators, these strains retained NtcA and PII, the key proteins for N sensing and regulation of N assimilation in cyanobacteria [138]. Albeit featuring a similar structure, subunit composition, physicochemical properties, and enzymatic performance compared to the GS of other cyanobacteria, early studies in Prochlorococcus illustrated that N shortage causes a different response compared to other cyanobacteria [139,140]. For instance, GS activity in Prochlorococcus MED4 was shown to be substantially enhanced during long-term N starvation, while GS protein levels remained relatively constant under these conditions [140] thus contrasting common regulatory traits of other cyanobacterial GSs in which N deprivation causes a significant increase in GS protein abundance and activity [141].
The steady GS abundance in response to N availability probably resulting from an inexpensive, less sophisticated regulation of this pivotal enzyme may impart evolutionary advantages in habitats occupied by Prochlorococcus. These display rather stable N supplies rendering excessive GS regulation superfluous and costly [142]. Nevertheless, the glnA gene was shown to be one of the top responding transcripts during N stress in Prochlorococcus and appears to be regulated by NtcA, consistent with observations made in other cyanobacteria [143]. These contradicting results, substantial alterations on the transcriptional level concomitant with steady protein abundances may be due to differential GS regulatory mechanisms on the posttranscriptional level in observed strains. Since the apparent protein level results from the antagonistic processes of protein synthesis and degradation, constant GS protein levels during N-starvation may depend on a degradation process triggered by N deprivation. An explanation could be provided by the susceptibility of the Prochlorococcus GS for metal-catalyzed oxidation (MCO) prior to degradation upon N limitation [144,145], which also occurs in E. coli and B. subtilis [146,147]. This process promotes the degradation of unessential proteins to provide amino acids for the synthesis of proteins crucial to cope with nutrient limitation thus meeting the challenges imposed to Prochlorococcus strains.
Most compelling, the previously described type 2 glutamine riboswitch, which always occurs in multiple copy numbers and appears restricted to marine picocyanobacteria, is functionally associated to genes encoding for DUF4278 containing proteins. Intriguingly, these proteins share substantial similarities with the N-terminus of IF17 [127], which was shown to be disposable for IF17 function but substantially promotes protein stability in Synechocystis [96]. Previous findings ruled out IF-mediated GS regulation in the clade of genome-streamlined marine picocyanobacteria living in ultraoligotrophic habitats that feature a monotonic nutritional nourishment [96,142]. Nevertheless it cannot be excluded that the GS and corresponding IFs of picocyanobacteria underwent co-evolutionary adaptions, which enables the interaction with these putative inhibitory proteins. This is supported by the finding that GS/IF-interaction is somewhat specific since GS of Anabaena could not be inhibited by IF7 and IF17 of Synechocystis in vitro [101]. For instance, the gene PMM1846 of Prochlorococcus strain MED4 (also referred to as CCMP1986/NIES-2087) encodes a small protein of 7 kDa that exhibits striking resemblance to IFs of other cyanobacteria. It bears DUF4278, homologous to the N-terminus of IF17, and harbors a glutamine riboswitch in its 5’UTR that boosts gene expression during N excess [127]. Moreover, it appears to be transcriptionally regulated with respect to N availability, as a NtcA consensus motif [87] is located in its promoter region. Finally, the protein encoded by PMM1846 shares several physicochemical properties with IFs, which are characteristic for proteins that partake in electrostatic protein-protein interactions such as the prevalence of “electrostatic targeting residues” like arginine [96]. Nevertheless, extensive analysis is required to clarify whether these genes indeed encode IF-like inhibitors of GS in the inherently underinvestigated clade of picocyanobacteria.
2.8. GlnN Represents Another GS in Cyanobacteria
As mentioned, GS splits in three distinct classes, which significantly differ in subunit size, subunit arrangement and sequence. First assumptions that another glutamine synthesizing enzyme exists in cyanobacteria were furnished by a glnA-deficient mutant of Synechococcus sp. PCC 7002, which was able to grow without glutamine and retained most of the wild-type glutamine biosynthetic activity [75]. Shortly afterwards, the generation of a Synechocystis glnA mutant strain, which exhibited WT like growth in nitrate-containing media, devoid of glutamine supplementation strongly supported the idea of an additional GS enzyme, which facilitates N assimilation during glnA deficiency [12]. Subsequent complementation of a glnA deficient E. coli mutant strain with a Synechocystis library led to the discovery of the glnN gene encoding a GS enzyme, which only shares a minor resemblance with the amply investigated GSI and GSII mostly restricted to 17 strongly conserved residues in the predicted ATP and cation binding sites. However, the protein was found homologous to the GSIII, which was previously described in the family of Bacteroidaceae with a sequence-derived subunit size of 75 kDa and a putative hexameric arrangement [12,13,148,149]. Interestingly, glnN expression is substantially affected by the N source as ammonium supplementation suppresses glnN expression while N-free media strongly induced glnN transcript accumulation. Being dispensable for N assimilation and WT-like growth rates, GSIII accounts for 3% of total GS activity in nitrate-grown Synechocystis cells [12]. In contrast, total GS activity is raised upon N-limiting conditions during which GSIII provides about one fifth of total glutamine biosynthetic activity, as it was shown by a comparison of WT and glnN-deficient strains [12]. Moreover, the conjecture that a Synechocystis glnA/glnN double mutant was not viable illustrates the fact that there are most likely no other GS enzymes present in this strain [12]. Subsequent purification and characterization of the GSIII protein confirmed the previously predicted hexameric subunit stoichiometry and molecular mass of ~79 kDa [12,150]. Furthermore, it was shown that GSIII features similar kinetic properties and requires the same substrates and cofactors as described for the cyanobacterial GSI [67,150]. Considering that transition-state analogs of γ-glutamylphosphate, which irreversibly inhibit GSI, had the same effect on GlnN activity, while catalytically crucial residues are conserved in both enzymes, strongly suggests that the catalytic mechanism of GSIII matches the situation in GSI [150]. Furthermore, utilizing GSIII-specific antibodies, it was shown that several other cyanobacterial strains possess the glnN gene whose expression is induced upon N starvation, consistent with observations made in Synechocystis, while diazotrophic cyanobacteria apparently lack the glnN gene [150]. Even though the glnN promoter sequence features an imperfect NtcA consensus motif as well as N-dependent expression patterns, first attempts to show E. coli-expressed NtcA binding failed and it was assumed that interaction requires additional factors [76]. Subsequent scrutiny of NtcA-deficient strains, however, unequivocally demonstrated that glnN is another NtcA regulated gene in Synechocystis [58,99]. In light of these data it is tempting to speculate whether GSIII is particularly important for prolonged N starvation upon which non-diazotrophic cyanobacteria pull out all the stops to scavenge every available ammonium molecule. Notably, in Synechococcus sp. PCC 7942 GSIII was shown to promote the synthesis of phycobiliproteins when chlorotic cells were supplied with limiting amounts of combined N [58]. Hence, GSIII enhances the incorporation of combined N to recover after long-term N starvation as phycobiliproteins are utilized as N reservoirs for ensuing phases of N starvation. It remains enigmatic whether GSIII in Synechocystis and other GSI bearing strains is regulated by IFs. Given that GSIII in Synechocystis only occurs upon prolonged N starvation in physiologically relevant levels, a circumstance that causes fully active enzymes [12], IF-dependent inhibition of GSIII would be rather redundant. Recently, physiological examination of GSIII from Synechococcus sp. WH7803 revealed an unusual regulation of this enzyme as GSIII activity in this strain is highly responsive to light-dark changes while being unaffected by N availability [151]. Thus, the assumption that GSIII promotes recovery after prolonged N starvation [58] appears unlikely in Synechococcus sp. WH7803. Given that all sequenced Prochlorococcus strains lack the glnN gene, it appears that GSIII was lost in the course of streamlining in environments with stable nutrient availability [151]. Therefore, Synechococcus sp. WH7803 could be on the verge of losing its GSIII and the loss of its regulation by N availability might be the first step in this process.
Intriguingly, one of the previously discovered glnN-bearing strains, Pseudanabaena 6903 was shown to lack a glnA gene and thus was the first known cyanobacterium in which only GSIII accounts for the biosynthesis of glutamine [152]. More recently, analysis of Synechococcus sp. RS9917 revealed that this marine cyanobacterium possesses two GSIII enzymes and lacks GSI [151]. In contrast to the situation in Synechocystis where nitrate-supplementation causes basal GSIII levels, glnN expression is strongly induced in nitrate-grown as well as N-starved cells of Pseudanabaena 6903, while ammonium addition triggers a rigorous inhibition of enzymatic activity accompanied by steady protein levels. This was not observed when N-starved cells were subjected to nitrate [152]. These findings clearly illustrate that GSIII in Pseudanabaena 6903 is subject to a yet unknown, rapid, ammonium-dependent inactivating mechanism, as minor GS activity would already be sufficient to ensure proper N supply under these conditions. Interestingly, glnN transcription in Pseudanabaena 6903 was also shown to be controlled by N, which is supported by the fact that the glnN promoter harbors an imperfect NtcA consensus sequence which is bound by NtcA of Synechocystis [152]. Thus, the regulatory pattern of glnN expression in Pseudanabaena 6903 is similar to the N-dependent control of glnA expression in Synechocystis as NtcA strongly induces the transcription of N-related genes during growth on nitrate or N-depleted media [76]. Thus, by employing NtcA-mediated N control of glnN transcription, Pseudanabaena 6903 deploys sufficient levels of its sole GS enzyme to meet metabolic demands during N-limiting conditions [152]. Unfortunately, there are no publicly available genomes of Pseudanabaena 6903, and thus the question whether GSIII is subject to protein-mediated regulation or an utterly different mechanism in this strain remains unanswered.
3. Concluding Remarks
Due to their photosynthetic lifestyle cyanobacteria are interesting hosts for the production of fuel components and valuable chemicals using light and CO2. However, their rising importance as microbial cell factories in biotechnology is contradicted by the current state of knowledge about metabolic regulation. As exemplified by the regulation of glutamine synthetase, cyanobacteria differ from other bacterial groups in many aspects. Hence, data that were obtained from genetic models like E. coli or B. subtilis can often not be applied. The intriguing example of cyanobacterial GS regulation suggests that further metabolic reactions might be controlled differently compared to other bacteria and that many regulatory factors still await discovery and/or characterization. Altogether, further basic research is required to make cyanobacteria more definable for metabolic engineering and thus also amenable for biotechnological applications.
Author Contributions
Conceptualization, S.K.; Original Draft Preparation, P.B. and S.K.; Review & Editing, P.B., M.I.M.-P., F.J.F. and S.K.
Funding
The work was financially supported by the Deutsche Forschungsgemeinschaft (DFG; Grant KL 3114/2-1 to S.K.) and by the Ministerio de Economía y Competitividad (MINECO; Grant BIO2016-75634-P to F.J.F., co-financed by European Regional Funds, FEDER).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2004, 2, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Forchhammer, K. Glutamine signalling in bacteria. Front. Biosci. J. Virtual Libr. 2007, 12, 358–370. [Google Scholar] [CrossRef]
- Zehr, J.P.; Ward, B.B. Nitrogen cycling in the ocean: New perspectives on processes and paradigms. Appl. Environ. Microbiol. 2002, 68, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Helling, R.B. Why does Escherichia coli have two primary pathways for synthesis of glutamate? J. Bacteriol. 1994, 176, 4664–4668. [Google Scholar] [CrossRef] [PubMed]
- Helling, R.B. Pathway choice in glutamate synthesis in Escherichia coli. J. Bacteriol. 1998, 180, 4571–4575. [Google Scholar] [PubMed]
- Halpern, Y.S.; Umbarger, H.E. Conversion of ammonia to amino groups in Escherichia coli. J. Bacteriol. 1960, 80, 285–288. [Google Scholar] [PubMed]
- Tempest, D.W.; Meers, J.L.; Brown, C.M. Synthesis of glutamate in Aerobacter aerogenes by a hitherto unknown route. Biochem. J. 1970, 117, 405–407. [Google Scholar] [CrossRef] [PubMed]
- Temple, S.J.; Vance, C.P.; Stephen Gantt, J. Glutamate synthase and nitrogen assimilation. Trends Plant Sci. 1998, 3, 51–56. [Google Scholar] [CrossRef]
- Kumada, Y.; Benson, D.R.; Hillemann, D.; Hosted, T.J.; Rochefort, D.A.; Thompson, C.J.; Wohlleben, W.; Tateno, Y. Evolution of the glutamine synthetase gene, one of the oldest existing and functioning genes. Proc. Natl. Acad. Sci. USA 1993, 90, 3009–3013. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.; Cullimore, J. Expression of three plant glutamine synthetase cDNA in Escherichia coli. Eur. J. Biochem. 1990, 193, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.R.; Masuchi, Y.; Robb, F.T.; Doolittlle, W.F. Evolutionary relationships of bacterial and archaeal glutamine synthetase genes. J. Mol. Evol. 1994, 38, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.C.; Florencio, F.J. A new type of glutamine synthetase in cyanobacteria: The protein encoded by the glnN gene supports nitrogen assimilation in Synechocystis sp. strain PCC 6803. J. Bacteriol. 1994, 176, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Southern, J.; Parker, J.R.; Woods, D.R. Novel structure, properties and inactivation of glutamine synthetase cloned from Bacteroides fragilis. Microbiology 1987, 133, 2437–2446. [Google Scholar] [CrossRef]
- Mathis, R.; Gamas, P.; Meyer, Y.; Cullimore, J.V. The presence of GSI-like genes in higher plants: Support for the paralogous evolution of GSI and GSII genes. J. Mol. Evol. 2000, 50, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.L.; Smith, G.J.; Alberte, R.S. Glutamine synthetase in marine algae: New surprises from an old enzyme. J. Phycol. 2002, 37, 793–795. [Google Scholar] [CrossRef]
- Darrow, R.A.; Knotts, R.R. Two forms of glutamine synthetase in free-living root-nodule bacteria. Biochem. Biophys. Res. Commun. 1977, 78, 554–559. [Google Scholar] [CrossRef]
- Edmands, J.; Noridge, N.A.; Benson, D.R. The actinorhizal root-nodule symbiont Frankia sp. strain CpI1 has two glutamine synthetases. Proc. Natl. Acad. Sci. USA 1987, 84, 6126–6130. [Google Scholar] [CrossRef] [PubMed]
- Kumada, Y.; Takano, E.; Nagaoka, K.; Thompson, C.J. Streptomyces hygroscopicus has two glutamine synthetase genes. J. Bacteriol. 1990, 172, 5343–5351. [Google Scholar] [CrossRef] [PubMed]
- Woolfolk, C.A.; Stadtman, E.R. Cumulative feedback inhibition in the multiple end product regulation of glutamine synthetase activity in Escherichia coli. Biochem. Biophys. Res. Commun. 1964, 17, 313–319. [Google Scholar] [CrossRef]
- Woolfolk, C.A.; Stadtman, E.R. Regulation of glutamine synthetase: III. Cumulative feedback inhibition of glutamine synthetase from Escherichia coli. Arch. Biochem. Biophys. 1967, 118, 736–755. [Google Scholar] [CrossRef]
- Woolfolk, C.A.; Shapiro, B.; Stadtman, E.R. Regulation of glutamine synthetase. I. Purification and properties of glutamine synthetase from Escherichia coli. Arch. Biochem. Biophys. 1966, 116, 177–192. [Google Scholar] [CrossRef]
- Liaw, S.H.; Pan, C.; Eisenberg, D. Feedback inhibition of fully unadenylylated glutamine synthetase from Salmonella typhimurium by glycine, alanine, and serine. Proc. Natl. Acad. Sci. USA 1993, 90, 4996–5000. [Google Scholar] [CrossRef] [PubMed]
- Liaw, S.H.; Jun, G.; Eisenberg, D. Interactions of nucleotides with fully unadenylylated glutamine synthetase from Salmonella typhimurium. Biochemistry 1994, 33, 11184–11188. [Google Scholar] [CrossRef] [PubMed]
- Adler, S.P.; Purich, D.; Stadtman, E.R. Cascade control of Escherichia coli glutamine synthetase. Properties of the PII regulatory protein and the uridylyltransferase-uridylyl-removing enzyme. J. Biol. Chem. 1975, 250, 6264–6272. [Google Scholar] [PubMed]
- Brown, M.S.; Segal, A.; Stadtman, E.R. Modulation of glutamine synthetase adenylylation and deadenylylation is mediated by metabolic transformation of the PII-regulatory protein. Proc. Natl. Acad. Sci. USA 1971, 68, 2949–2953. [Google Scholar] [CrossRef] [PubMed]
- Kingdon, H.S.; Shapiro, B.M.; Stadtman, E.R. Regulation of glutamine synthetase. VIII. ATP: Glutamine synthetase adenylyltransferase, an enzyme that catalyzes alterations in the regulatory properties of glutamine synthetase. Proc. Natl. Acad. Sci. USA 1967, 58, 1703–1710. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Oxidation of free amino acids and amino acid residues in proteins by radiolysis and by metal-catalyzed reactions. Annu. Rev. Biochem. 1993, 62, 797–821. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.; Rhee, S.G. Cascade control of Escherichia coli glutamine synthetase. Purification and properties of PII uridylyltransferase and uridylyl-removing enzyme. J. Biol. Chem. 1983, 258, 2246–2253. [Google Scholar] [PubMed]
- Mangum, J.H.; Magni, G.; Stadtman, E.R. Regulation of glutamine synthetase adenylylation and deadenylylation by the enzymatic uridylylation and deuridylylation of the PII regulatory protein. Arch. Biochem. Biophys. 1973, 158, 514–525. [Google Scholar] [CrossRef]
- Rhee, S.G.; Chock, P.B. Purification and characterization of uridylylated and unuridylylated forms of regulatory protein PII involved in the glutamine synthetase regulation in Escherichia coli. Isozymes 1983, 8, 141–153. [Google Scholar] [PubMed]
- Anderson, W.B.; Stadtman, E.R. Glutamine synthetase deadenylylation: A phosphorolytic reaction yielding ADP as nucleotide product. Biochem. Biophys. Res. Commun. 1970, 41, 704–709. [Google Scholar] [CrossRef]
- Shapiro, B.M. Glutamine synthetase deadenylylating enzyme system from Escherichia coli. Resolution into two components, specific nucleotide stimulation, and cofactor requirements. Biochemistry 1969, 8, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, B.M.; Stadtman, E.R. Glutamine synthetase deadenylylating enzyme. Biochem. Biophys. Res. Commun. 1968, 30, 32–37. [Google Scholar] [CrossRef]
- Shapiro, B.M.; Kingdon, H.S.; Stadtman, E.R. Regulation of glutamine synthetase. VII. Adenylyl glutamine synthetase: A new form of the enzyme with altered regulatory and kinetic properties. Proc. Natl. Acad. Sci. USA 1967, 58, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.A.; Dodsworth, J.A. Nitrogen regulation in bacteria and archaea. Annu. Rev. Microbiol. 2007, 61, 349–377. [Google Scholar] [CrossRef] [PubMed]
- Anderson, W.B.; Hennig, S.B.; Ginsburg, A.; Stadtman, E.R. Association of ATP: Glutamine synthetase adenylyltransferase activity with the PI component of the glutamine synthetase deadenylylation system. Proc. Natl. Acad. Sci. USA 1970, 67, 1417–1424. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G.; Park, S.C.; Koo, J.H. The role of adenylyltransferase and uridylyltransferase in the regulation of glutamine synthetase in Escherichia coli. In Current Topics in Cellular Regulation; Shaltiel, S., Chock, P.B., Eds.; Modulation by Covalent Modification; Elsevier: Amsterdam, Netherlands, 1985; Volume 27, pp. 221–232. [Google Scholar]
- Stadtman, E.R. The story of glutamine synthetase regulation. J. Biol. Chem. 2001, 276, 44357–44364. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Regulation of glutamine synthetase activity. EcoSal Plus 2004, 1. [Google Scholar] [CrossRef] [PubMed]
- Weiss, V.; Kramer, G.; Dünnebier, T.; Flotho, A. Mechanism of regulation of the bifunctional histidine kinase NtrB in Escherichia coli. J. Mol. Microbiol. Biotechnol. 2002, 4, 229–233. [Google Scholar] [PubMed]
- Merrick, M.J.; Edwards, R.A. Nitrogen control in bacteria. Microbiol. Rev. 1995, 59, 604–622. [Google Scholar] [PubMed]
- Wray, L.V.; Zalieckas, J.M.; Fisher, S.H. Bacillus subtilis glutamine synthetase controls gene expression through a protein-protein interaction with transcription factor TnrA. Cell 2001, 107, 427–435. [Google Scholar] [CrossRef]
- Fedorova, K.; Kayumov, A.; Woyda, K.; Ilinskaja, O.; Forchhammer, K. Transcription factor TnrA inhibits the biosynthetic activity of glutamine synthetase in Bacillus subtilis. FEBS Lett. 2013, 587, 1293–1298. [Google Scholar] [CrossRef] [PubMed]
- Shu, C.J.; Zhulin, I.B. ANTAR: An RNA-binding domain in transcription antitermination regulatory proteins. Trends Biochem. Sci. 2002, 27, 3–5. [Google Scholar] [CrossRef]
- Stutz, H.E.; Quixley, K.W.M.; McMaster, L.D.; Reid, S.J. Co-regulation of the nitrogen-assimilatory gene cluster in Clostridium saccharobutylicum. Microbiol. Read. Engl. 2007, 153, 3081–3090. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, C.; Weidenbach, K.; Veit, K.; Forchhammer, K.; Schmitz, R.A. Unique mechanistic features of post-translational regulation of glutamine synthetase activity in Methanosarcina mazei strain Gö1 in response to nitrogen availability. Mol. Microbiol. 2005, 55, 1841–1854. [Google Scholar] [CrossRef] [PubMed]
- Pedro-Roig, L.; Camacho, M.; Bonete, M.-J. Regulation of ammonium assimilation in Haloferax mediterranei: Interaction between glutamine synthetase and two GlnK proteins. Biochim. Biophys. Acta BBA Proteins Proteom. 2013, 1834, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.R.; Castenholz, R.W. Evolution of thermotolerance in hot spring cyanobacteria of the genus Synechococcus. Appl. Environ. Microbiol. 2000, 66, 4222–4229. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.D.; Shukla, S.P.; Shukla, P.N.; Giri, D.D.; Singh, J.S.; Singh, P.; Kashyap, A.K. Cyanobacteria in Antarctica: Ecology, physiology and cold adaptation. Cell. Mol. Biol. 2004, 50, 575–584. [Google Scholar] [PubMed]
- Reed, R.H.; Chudek, J.A.; Foster, R.; Stewart, W.D.P. Osmotic adjustment in cyanobacteria from hypersaline environments. Arch. Microbiol. 1984, 138, 333–337. [Google Scholar] [CrossRef]
- Skulberg, O. Part 9. Terrestrial and Limnic Algae and Cyanobacteria. Available online: https://eurekamag.com/research/009/158/009158962.php (accessed on 6 July 2018).
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [PubMed]
- Puente-Sánchez, F.; Arce-Rodríguez, A.; Oggerin, M.; García-Villadangos, M.; Moreno-Paz, M.; Blanco, Y.; Rodríguez, N.; Bird, L.; Lincoln, S.A.; Tornos, F.; et al. Viable cyanobacteria in the deep continental subsurface. Proc. Natl. Acad. Sci. USA 2018, 201808176. [Google Scholar] [CrossRef] [PubMed]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincón, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.W.; Lomas, M.W.; Veneziano, D.; et al. Present and future global distributions of the marine cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [PubMed]
- Dismukes, G.C.; Klimov, V.V.; Baranov, S.V.; Kozlov, Y.N.; DasGupta, J.; Tyryshkin, A. The origin of atmospheric oxygen on Earth: The innovation of oxygenic photosynthesis. Proc. Natl. Acad. Sci. USA 2001, 98, 2170–2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, E.; Herrero, A. Assimilatory nitrogen metabolism and its regulation. In The Molecular Biology of Cyanobacteria; Advances in Photosynthesis; Springer: Dordrecht, The Netherlands, 1994; pp. 487–517. ISBN 978-0-7923-3273-2. [Google Scholar]
- Herrero, A.; Muro-Pastor, A.M.; Flores, E. Nitrogen control in cyanobacteria. J. Bacteriol. 2001, 183, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Sauer, J.; Dirmeier, U.; Forchhammer, K. The Synechococcus strain PCC 7942 glnN product (Glutamine Synthetase III) helps recovery from prolonged nitrogen chlorosis. J. Bacteriol. 2000, 182, 5615–5619. [Google Scholar] [CrossRef] [PubMed]
- Dharmawardene, M.W.; Haystead, A.; Stewart, W.D. Glutamine synthetase of the nitrogen-fixing alga Anabaena cylindrica. Arch. Mikrobiol. 1973, 90, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Lea, P.J.; Miflin, B.J. Glutamate synthase in blue-green algae. Biochem. Soc. Trans. 1975, 3, 381–384. [Google Scholar] [CrossRef] [PubMed]
- Neilson, A.H.; Doudoroff, M. Ammonia assimilation in blue-green algae. Arch. Mikrobiol. 1973, 89, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Meeks, J.C.; Wolk, C.P.; Thomas, J.; Lockau, W.; Shaffer, P.W.; Austin, S.M.; Chien, W.S.; Galonsky, A. The pathways of assimilation of 13NH4+ by the cyanobacterium, Anabaena cylindrica. J. Biol. Chem. 1977, 252, 7894–7900. [Google Scholar] [PubMed]
- Meeks, J.C.; Wolk, C.P.; Lockau, W.; Schilling, N.; Shaffer, P.W.; Chien, W.S. Pathways of assimilation of [13N]N2 and 13NH4+ by cyanobacteria with and without heterocysts. J. Bacteriol. 1978, 134, 125–130. [Google Scholar] [PubMed]
- Wolk, C.P.; Thomas, J.; Shaffer, P.W.; Austin, S.M.; Galonsky, A. Pathway of nitrogen metabolism after fixation of 13N-labeled nitrogen gas by the cyanobacterium Anabaena cylindrica. J. Biol. Chem. 1976, 251, 5027–5034. [Google Scholar] [PubMed]
- Chávez, S.; Lucena, J.M.; Reyes, J.C.; Florencio, F.J.; Candau, P. The presence of glutamate dehydrogenase is a selective advantage for the cyanobacterium Synechocystis sp. strain PCC 6803 under nonexponential growth conditions. J. Bacteriol. 1999, 181, 808–813. [Google Scholar] [PubMed]
- Fisher, R.; Tuli, R.; Haselkorn, R. A cloned cyanobacterial gene for glutamine synthetase functions in Escherichia coli, but the enzyme is not adenylylated. Proc. Natl. Acad. Sci. USA 1981, 78, 3393–3397. [Google Scholar] [CrossRef] [PubMed]
- Mérida, A.; Leurentop, L.; Candau, P.; Florencio, F.J. Purification and properties of glutamine synthetases from the cyanobacteria Synechocystis sp. strain PCC 6803 and Calothrix sp. strain PCC 7601. J. Bacteriol. 1990, 172, 4732–4735. [Google Scholar] [CrossRef] [PubMed]
- Orr, J.; Haselkorn, R. Kinetic and inhibition studies of glutamine synthetase from the cyanobacterium Anabaena 7120. J. Biol. Chem. 1981, 256, 13099–13104. [Google Scholar] [PubMed]
- Stacey, G.; Tabita, F.R.; Baalen, C.V. Nitrogen and ammonia assimilation in the cyanobacteria: Purification of glutamine synthetase from Anabaena sp. Strain CA. J. Bacteriol. 1977, 132, 596–603. [Google Scholar] [PubMed]
- Bottomley, P.J.; Stewart, W.D.P. ATP pools and transients in the blue-green alga, Anabaena cylindrica. Arch. Microbiol. 1976, 108, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Kupiec, R.; Gurevitz, M.; Zilberstein, A. Expression of glnA in the cyanobacterium Synechococcus sp. strain PCC 7942 is initiated from a single nif-like promoter under various nitrogen conditions. J. Bacteriol. 1993, 175, 7727–7731. [Google Scholar] [CrossRef] [PubMed]
- Elmorjani, K.; Liotenberg, S.; Houmard, J.; de Marsac, N.T. Molecular characterization of the gene encoding glutamine synthetase in the cyanobacterium Calothrix sp. PCC 7601. Biochem. Biophys. Res. Commun. 1992, 189, 1296–1302. [Google Scholar] [CrossRef]
- Orr, J.; Haselkorn, R. Regulation of glutamine synthetase activity and synthesis in free-living and symbiotic Anabaena spp. J. Bacteriol. 1982, 152, 626–635. [Google Scholar] [PubMed]
- Tumer, N.E.; Robinson, S.J.; Haselkorn, R. Different promoters for the Anabaena glutamine synthetase gene during growth using molecular or fixed nitrogen. Nature 1983, 306, 337–342. [Google Scholar] [CrossRef]
- Wagner, S.J.; Thomas, S.P.; Kaufman, R.I.; Nixon, B.T.; Stevens, S.E. The glnA gene of the cyanobacterium Agmenellum quadruplicatum PR-6 is nonessential for ammonium assimilation. J. Bacteriol. 1993, 175, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.C.; Muro-Pastor, M.I.; Florencio, F.J. Transcription of glutamine synthetase genes (glnA and glnN) from the cyanobacterium Synechocystis sp. strain PCC 6803 is differently regulated in response to nitrogen availability. J. Bacteriol. 1997, 179, 2678–2689. [Google Scholar] [CrossRef] [PubMed]
- Vega-Palas, M.A.; Flores, E.; Herrero, A. NtcA, a global nitrogen regulator from the cyanobacterium Synechococcus that belongs to the Crp family of bacterial regulators. Mol. Microbiol. 1992, 6, 1853–1859. [Google Scholar] [CrossRef] [PubMed]
- Herrero, A.; Paz-Yepes, J.; Vázquez-Bermúdez, M.F.; Flores, E. The NtcA-activated amt1 gene encodes a permease required for uptake of low concentrations of ammonium in the cyanobacterium Synechococcus sp. PCC 7942. Microbiology 2002, 148, 861–869. [Google Scholar] [CrossRef]
- Ohashi, Y.; Shi, W.; Takatani, N.; Aichi, M.; Maeda, S.; Watanabe, S.; Yoshikawa, H.; Omata, T. Regulation of nitrate assimilation in cyanobacteria. J. Exp. Bot. 2011, 62, 1411–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osanai, T.; Tanaka, K. Keeping in touch with PII: PII-interacting proteins in unicellular cyanobacteria. Plant Cell Physiol. 2007, 48, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, J.; Forchhammer, K.; Burillo, S.; Contreras, A. Interaction network in cyanobacterial nitrogen regulation: PipX, a protein that interacts in a 2-oxoglutarate dependent manner with PII and NtcA. Mol. Microbiol. 2006, 61, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, J.; Rodríguez-Mateos, F.; Salinas, P.; Lanza, V.F.; Dixon, R.; de la Cruz, F.; Contreras, A. PipX, the coactivator of NtcA, is a global regulator in cyanobacteria. Proc. Natl. Acad. Sci. USA 2014, 111, E2423–E2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forcada-Nadal, A.; Forchhammer, K.; Rubio, V. SPR analysis of promoter binding of Synechocystis PCC6803 transcription factors NtcA and CRP suggests cross-talk and sheds light on regulation by effector molecules. FEBS Lett. 2014, 588, 2270–2276. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, M.I.; Reyes, J.C.; Florencio, F.J. Cyanobacteria perceive nitrogen status by sensing intracellular 2-oxoglutarate levels. J. Biol. Chem. 2001, 276, 38320–38328. [Google Scholar] [CrossRef] [PubMed]
- Tanigawa, R.; Shirokane, M.; Maeda Si, S.; Omata, T.; Tanaka, K.; Takahashi, H. Transcriptional activation of NtcA-dependent promoters of Synechococcus sp. PCC 7942 by 2-oxoglutarate in vitro. Proc. Natl. Acad. Sci. USA 2002, 99, 4251–4255. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-X.; Jiang, Y.-L.; He, Y.-X.; Chen, Y.-F.; Teng, Y.-B.; Chen, Y.; Zhang, C.-C.; Zhou, C.-Z. Structural basis for the allosteric control of the global transcription factor NtcA by the nitrogen starvation signal 2-oxoglutarate. Proc. Natl. Acad. Sci. USA 2010, 107, 12487–12492. [Google Scholar] [CrossRef] [PubMed]
- Giner-Lamia, J.; Robles-Rengel, R.; Hernández-Prieto, M.A.; Muro-Pastor, M.I.; Florencio, F.J.; Futschik, M.E. Identification of the direct regulon of NtcA during early acclimation to nitrogen starvation in the cyanobacterium Synechocystis sp. PCC 6803. Nucleic Acids Res. 2017, 45, 11800–11820. [Google Scholar] [CrossRef] [PubMed]
- Luque, I.; Flores, E.; Herrero, A. Molecular mechanism for the operation of nitrogen control in cyanobacteria. EMBO J. 1994, 13, 2862–2869. [Google Scholar] [CrossRef] [PubMed]
- Mitschke, J.; Vioque, A.; Haas, F.; Hess, W.R.; Muro-Pastor, A.M. Dynamics of transcriptional start site selection during nitrogen stress-induced cell differentiation in Anabaena sp. PCC7120. Proc. Natl. Acad. Sci. USA 2011, 108, 20130–20135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vázquez-Bermúdez, M.F.; Herrero, A.; Flores, E. 2-Oxoglutarate increases the binding affinity of the NtcA (nitrogen control) transcription factor for the Synechococcus glnA promoter. FEBS Lett. 2002, 512, 71–74. [Google Scholar] [CrossRef]
- Zeth, K.; Fokina, O.; Forchhammer, K. Structural basis and target-specific modulation of ADP sensing by the Synechococcus elongatus PII signaling protein. J. Biol. Chem. 2014, 289, 8960–8972. [Google Scholar] [CrossRef] [PubMed]
- Mérida, A.; Candau, P.; Florencio, F.J. Regulation of glutamine synthetase activity in the unicellular cyanobacterium Synechocystis sp. strain PCC 6803 by the nitrogen source: Effect of ammonium. J. Bacteriol. 1991, 173, 4095–4100. [Google Scholar] [CrossRef] [PubMed]
- Mérida, A.; Candau, P.; Florencio, F.J. In vitro reactivation of in vivo ammonium-inactivated glutamine synthetase from Synechocystis sp. PCC 6803. Biochem. Biophys. Res. Commun. 1991, 181, 780–786. [Google Scholar] [CrossRef]
- Reyes, J.C.; Florencio, F.J. A novel mechanism of glutamine synthetase inactivation by ammonium in the cyanobacterium Synechocystis sp. PCC 6803. Involvement of an inactivating protein. FEBS Lett. 1995, 367, 45–48. [Google Scholar] [CrossRef]
- García-Domínguez, M.; Reyes, J.C.; Florencio, F.J. Glutamine synthetase inactivation by protein–protein interaction. Proc. Natl. Acad. Sci. USA 1999, 96, 7161–7166. [Google Scholar] [CrossRef] [PubMed]
- Saelices, L.; Galmozzi, C.V.; Florencio, F.J.; Muro-Pastor, M.I. Mutational analysis of the inactivating factors, IF7 and IF17 from Synechocystis sp. PCC 6803: Critical role of arginine amino acid residues for glutamine synthetase inactivation. Mol. Microbiol. 2011, 82, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Galmozzi, C.V.; Fernández-Avila, M.J.; Reyes, J.C.; Florencio, F.J.; Muro-Pastor, M.I. The ammonium-inactivated cyanobacterial glutamine synthetase I is reactivated in vivo by a mechanism involving proteolytic removal of its inactivating factors. Mol. Microbiol. 2007, 65, 166–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saelices, L.; Robles-Rengel, R.; Florencio, F.J.; Muro-Pastor, M.I. A core of three amino acids at the carboxyl-terminal region of glutamine synthetase defines its regulation in cyanobacteria. Mol. Microbiol. 2015, 96, 483–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Domínguez, M.; Reyes, J.C.; Florencio, F.J. NtcA represses transcription of gifA and gifB, genes that encode inhibitors of glutamine synthetase type I from Synechocystis sp. PCC 6803. Mol. Microbiol. 2000, 35, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Luque, I.; Forchhammer, K. Nitrogen assimilation and C/N balance sensing. In The Cyanobacteria: Molecular Biology, Genetics and Evolution; Herrero, A., Flores, E., Eds.; Caister Academic Press: Norwich, UK, 2008; pp. 335–382. [Google Scholar]
- Galmozzi, C.V.; Saelices, L.; Florencio, F.J.; Muro-Pastor, M.I. Posttranscriptional regulation of glutamine synthetase in the filamentous cyanobacterium Anabaena sp. PCC 7120: Differential expression between vegetative cells and heterocysts. J. Bacteriol. 2010, 192, 4701–4711. [Google Scholar] [CrossRef] [PubMed]
- Waters, L.S.; Storz, G. Regulatory RNAs in bacteria. Cell 2009, 136, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Storz, G.; Vogel, J.; Wassarman, K.M. Regulation by small RNAs in bacteria: Expanding frontiers. Mol. Cell 2011, 43, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.G.H.; Romby, P. Small RNAs in bacteria and archaea: Who they are, what they do, and how they do it. Adv. Genet. 2015, 90, 133–208. [Google Scholar] [CrossRef] [PubMed]
- Kopf, M.; Hess, W.R. Regulatory RNAs in photosynthetic cyanobacteria. FEMS Microbiol. Rev. 2015, 39, 301–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopf, M.; Klähn, S.; Scholz, I.; Matthiessen, J.K.F.; Hess, W.R.; Voß, B. Comparative analysis of the primary transcriptome of Synechocystis sp. PCC 6803. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2014, 21, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Mitschke, J.; Georg, J.; Scholz, I.; Sharma, C.M.; Dienst, D.; Bantscheff, J.; Voß, B.; Steglich, C.; Wilde, A.; Vogel, J.; et al. An experimentally anchored map of transcriptional start sites in the model cyanobacterium Synechocystis sp. PCC6803. Proc. Natl. Acad. Sci. USA 2011, 108, 2124–2129. [Google Scholar] [CrossRef] [PubMed]
- Voss, B.; Georg, J.; Schön, V.; Ude, S.; Hess, W.R. Biocomputational prediction of non-coding RNAs in model cyanobacteria. BMC Genom. 2009, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, D.; Voss, B.; Oren, A.; Hess, W.R.; Muro-Pastor, A.M. Heterocyst-specific transcription of NsiR1, a non-coding RNA encoded in a tandem array of direct repeats in cyanobacteria. J. Mol. Biol. 2010, 398, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, A.M. The heterocyst-specific NsiR1 small RNA is an early marker of cell differentiation in cyanobacterial filaments. mBio 2014, 5, e01079-14. [Google Scholar] [CrossRef] [PubMed]
- Klähn, S.; Schaal, C.; Georg, J.; Baumgartner, D.; Knippen, G.; Hagemann, M.; Muro-Pastor, A.M.; Hess, W.R. The sRNA NsiR4 is involved in nitrogen assimilation control in cyanobacteria by targeting glutamine synthetase inactivating factor IF7. Proc. Natl. Acad. Sci. USA 2015, 112, E6243–E6252. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.; Bryant, D.A. Acclimation of the global transcriptome of the cyanobacterium Synechococcus sp. Strain PCC 7002 to nutrient limitations and different nitrogen sources. Front. Microbiol. 2012, 3, 145. [Google Scholar] [CrossRef] [PubMed]
- Kopf, M.; Klähn, S.; Scholz, I.; Hess, W.R.; Voß, B. Variations in the non-coding transcriptome as a driver of inter-strain divergence and physiological adaptation in bacteria. Sci. Rep. 2015, 5, 9560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.; Wagner, E.G.H. Target identification of small noncoding RNAs in bacteria. Curr. Opin. Microbiol. 2007, 10, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Escribano, I.; Vioque, A.; Muro-Pastor, A.M. NsrR1, a nitrogen stress-repressed sRNA, contributes to the regulation of nblA in Nostoc sp. PCC 7120. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Prasse, D.; Schmitz, R.A. Small RNAs involved in regulation of nitrogen metabolism. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Mironov, A.S.; Gusarov, I.; Rafikov, R.; Lopez, L.E.; Shatalin, K.; Kreneva, R.A.; Perumov, D.A.; Nudler, E. Sensing small molecules by nascent RNA: A mechanism to control transcription in bacteria. Cell 2002, 111, 747–756. [Google Scholar] [CrossRef]
- Nahvi, A.; Sudarsan, N.; Ebert, M.S.; Zou, X.; Brown, K.L.; Breaker, R.R. Genetic control by a metabolite binding mRNA. Chem. Biol. 2002, 9, 1043–1049. [Google Scholar] [CrossRef]
- Winkler, W.; Nahvi, A.; Breaker, R.R. Thiamine derivatives bind messenger RNAs directly to regulate bacterial gene expression. Nature 2002, 419, 952–956. [Google Scholar] [CrossRef] [PubMed]
- Barrick, J.E.; Breaker, R.R. The distributions, mechanisms, and structures of metabolite-binding riboswitches. Genome Biol. 2007, 8, R239. [Google Scholar] [CrossRef] [PubMed]
- Breaker, R.R. Riboswitches: From ancient gene-control systems to modern drug targets. Future Microbiol. 2009, 4, 771–773. [Google Scholar] [CrossRef] [PubMed]
- Dambach, M.D.; Winkler, W.C. Expanding roles for metabolite-sensing regulatory RNAs. Curr. Opin. Microbiol. 2009, 12, 161–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garst, A.D.; Edwards, A.L.; Batey, R.T. Riboswitches: Structures and mechanisms. Cold Spring Harb. Perspect. Biol. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, Z.; Wang, J.X.; Bogue, J.; Yang, J.; Corbino, K.; Moy, R.H.; Breaker, R.R. Comparative genomics reveals 104 candidate structured RNAs from bacteria, archaea, and their metagenomes. Genome Biol. 2010, 11, R31. [Google Scholar] [CrossRef] [PubMed]
- Kalvari, I.; Argasinska, J.; Quinones-Olvera, N.; Nawrocki, E.P.; Rivas, E.; Eddy, S.R.; Bateman, A.; Finn, R.D.; Petrov, A.I. Rfam 13.0: Shifting to a genome-centric resource for non-coding RNA families. Nucleic Acids Res. 2018, 46, D335–D342. [Google Scholar] [CrossRef] [PubMed]
- Ames, T.D.; Breaker, R.R. Bacterial aptamers that selectively bind glutamine. RNA Biol. 2011, 8, 82–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klähn, S.; Bolay, P.; Wright, P.R.; Atilho, R.M.; Brewer, K.I.; Hagemann, M.; Breaker, R.R.; Hess, W.R. A glutamine riboswitch is a key element for the regulation of glutamine synthetase in cyanobacteria. Nucleic Acids Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.W.; Olson, R.J.; Zettler, E.R.; Goericke, R.; Waterbury, J.B.; Welschmeyer, N.A. A novel free-living prochlorophyte abundant in the oceanic euphotic zone. Nature 1988, 334, 340–343. [Google Scholar] [CrossRef]
- Moore, L.R.; Chisholm, S.W. Photophysiology of the marine cyanobacterium Prochlorococcus: Ecotypic differences among cultured isolates. Limnol. Oceanogr. 1999, 44, 628–638. [Google Scholar] [CrossRef]
- Chisholm, S.W.; Frankel, S.L.; Goericke, R.; Olson, R.J.; Palenik, B.; Waterbury, J.B.; West-Johnsrud, L.; Zettler, E.R. Prochlorococcus marinus nov. gen. nov. sp.: An oxyphototrophic marine prokaryote containing divinyl chlorophyll a and b. Arch. Microbiol. 1992, 157, 297–300. [Google Scholar] [CrossRef]
- Rocap, G.; Larimer, F.W.; Lamerdin, J.; Malfatti, S.; Chain, P.; Ahlgren, N.A.; Arellano, A.; Coleman, M.; Hauser, L.; Hess, W.R.; et al. Genome divergence in two Prochlorococcus ecotypes reflects oceanic niche differentiation. Nature 2003, 424, 1042–1047. [Google Scholar] [CrossRef] [PubMed]
- Dufresne, A.; Garczarek, L.; Partensky, F. Accelerated evolution associated with genome reduction in a free-living prokaryote. Genome Biol. 2005, 6, R14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovannoni, S.J.; Thrash, J.C.; Temperton, B. Implications of streamlining theory for microbial ecology. ISME J. 2014, 8, 1553–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steglich, C.; Futschik, M.E.; Lindell, D.; Voss, B.; Chisholm, S.W.; Hess, W.R. The challenge of regulation in a minimal photoautotroph: Non-coding RNAs in Prochlorococcus. PLoS Genet. 2008, 4, e1000173. [Google Scholar] [CrossRef]
- Moore, C.M.; Mills, M.M.; Arrigo, K.R.; Berman-Frank, I.; Bopp, L.; Boyd, P.W.; Galbraith, E.D.; Geider, R.J.; Guieu, C.; Jaccard, S.L.; et al. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 2013, 6, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Grzymski, J.J.; Dussaq, A.M. The significance of nitrogen cost minimization in proteomes of marine microorganisms. ISME J. 2012, 6, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Read, R.W.; Berube, P.M.; Biller, S.J.; Neveux, I.; Cubillos-Ruiz, A.; Chisholm, S.W.; Grzymski, J.J. Nitrogen cost minimization is promoted by structural changes in the transcriptome of N-deprived Prochlorococcus cells. ISME J. 2017, 11, 2267–2278. [Google Scholar] [CrossRef] [PubMed]
- Scanlan, D.J.; Ostrowski, M.; Mazard, S.; Dufresne, A.; Garczarek, L.; Hess, W.R.; Post, A.F.; Hagemann, M.; Paulsen, I.; Partensky, F. Ecological genomics of marine picocyanobacteria. Microbiol. Mol. Biol. Rev. MMBR 2009, 73, 249–299. [Google Scholar] [CrossRef] [PubMed]
- El Alaoui, S.; Diez, J.; Humanes, L.; Toribio, F.; Partensky, F.; García-Fernández, J.M. In vivo regulation of glutamine synthetase activity in the marine chlorophyll b-containing cyanobacterium Prochlorococcus sp. strain PCC 9511 (oxyphotobacteria). Appl. Environ. Microbiol. 2001, 67, 2202–2207. [Google Scholar] [CrossRef] [PubMed]
- El Alaoui, S.; Diez, J.; Toribio, F.; Gómez-Baena, G.; Dufresne, A.; García-Fernández, J.M. Glutamine synthetase from the marine cyanobacteria Prochlorococcus spp: Characterization, phylogeny and response to nutrient limitation. Environ. Microbiol. 2003, 5, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Florencio, F.J.; Reyes, J.C. Regulation of ammonium assimilation in cyanobacteria. In Photosynthetic Nitrogen Assimilation and Associated Carbon and Respiratory Metabolism; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2002; pp. 93–113. ISBN 978-0-7923-6336-1. [Google Scholar]
- García-Fernández, J.M.; de Marsac, N.T.; Diez, J. Streamlined regulation and gene loss as adaptive mechanisms in Prochlorococcus for optimized nitrogen utilization in oligotrophic environments. Microbiol. Mol. Biol. Rev. 2004, 68, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Tolonen, A.C.; Aach, J.; Lindell, D.; Johnson, Z.I.; Rector, T.; Steen, R.; Church, G.M.; Chisholm, S.W. Global gene expression of Prochlorococcus ecotypes in response to changes in nitrogen availability. Mol. Syst. Biol. 2006, 2, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Baena, G.; Diez, J.; García-Fernández, J.M.; El Alaoui, S.; Humanes, L. Regulation of glutamine synthetase by metal-catalyzed oxidative modification in the marine oxyphotobacterium Prochlorococcus. Biochim. Biophys. Acta 2001, 1568, 237–244. [Google Scholar] [CrossRef]
- Gómez-Baena, G.; Domínguez-Martín, M.A.; Donaldson, R.P.; García-Fernández, J.M.; Diez, J. Glutamine synthetase sensitivity to oxidative modification during nutrient starvation in Prochlorococcus marinus PCC 9511. PLoS ONE 2015, 10, e0135322. [Google Scholar] [CrossRef] [PubMed]
- Kimura, K.; Sugano, S. Inactivation of Bacillus subtilis glutamine synthetase by metal-catalyzed oxidation. J. Biochem. 1992, 112, 828–833. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Oliver, C.N.; Fulks, R.M.; Stadtman, E.R. Turnover of bacterial glutamine synthetase: Oxidative inactivation precedes proteolysis. Proc. Natl. Acad. Sci. USA 1981, 78, 2120–2124. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.T.; Parker, J.R.; Goodman, H.J.; Jones, D.T.; Woods, D.R. Molecular analysis of a novel glutamine synthetase of the anaerobe Bacteroides fragilis. J. Gen. Microbiol. 1989, 135, 3271–3279. [Google Scholar] [CrossRef] [PubMed]
- Pesole, G.; Bozzetti, M.P.; Lanave, C.; Preparata, G.; Saccone, C. Glutamine synthetase gene evolution: A good molecular clock. Proc. Natl. Acad. Sci. USA 1991, 88, 522–526. [Google Scholar] [CrossRef] [PubMed]
- García-Domínguez, M.; Reyes, J.C.; Florencio, F.J. Purification and characterization of a new type of glutamine synthetase from cyanobacteria. Eur. J. Biochem. 1997, 244, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Martín, M.A.; Díez, J.; García-Fernández, J.M. Physiological studies of glutamine synthetases I and III from Synechococcus sp. WH7803 reveal differential regulation. Front. Microbiol. 2016, 7, 969. [Google Scholar] [CrossRef] [PubMed]
- Crespo, J.L.; García-Domínguez, M.; Florencio, F.J. Nitrogen control of the glnN gene that codes for GS type III, the only glutamine synthetase in the cyanobacterium Pseudanabaena sp. PCC 6903. Mol. Microbiol. 1998, 30, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Ammonium assimilation reactions catalyzed by the glutamate dehydrogenase (GDH) and the glutamine synthetase/glutamate synthase (GS/GOGAT) cycle.
Figure 1.
Ammonium assimilation reactions catalyzed by the glutamate dehydrogenase (GDH) and the glutamine synthetase/glutamate synthase (GS/GOGAT) cycle.

Figure 2.
Simplified overview of N-dependent glutamine synthetase (GS) regulation in E. coli: GSI consists of two superimposed hexameric rings, which are arranged in a centrosymmetric structure. The bidirectional uridylyltransferase (UTase) senses both glutamine (Gln) and 2-oxoglutarate (2OG) and modifies the PII signal transducer accordingly. Unmodified PII promotes adenylylation of GS via the bidirectional adenylyltransferase (ATase), which enhances the sensitivity of the enzyme for feedback inhibition. Accordingly, modified PII facilitates deadenylylation of GS, rendering the enzyme fully active. Moreover, by impacting the bidirectional kinase/phosphatase activity of the sensor kinase NtrB, PII also regulates the NtrC-NtrB two-component system, which controls transcription of the GS encoding glnA gene. Under N excess, unmodified PII promotes NtrB-dependent dephosphorylation of the response regulator NtrC, which prevents activation of glnA expression [38,39].
Figure 2.
Simplified overview of N-dependent glutamine synthetase (GS) regulation in E. coli: GSI consists of two superimposed hexameric rings, which are arranged in a centrosymmetric structure. The bidirectional uridylyltransferase (UTase) senses both glutamine (Gln) and 2-oxoglutarate (2OG) and modifies the PII signal transducer accordingly. Unmodified PII promotes adenylylation of GS via the bidirectional adenylyltransferase (ATase), which enhances the sensitivity of the enzyme for feedback inhibition. Accordingly, modified PII facilitates deadenylylation of GS, rendering the enzyme fully active. Moreover, by impacting the bidirectional kinase/phosphatase activity of the sensor kinase NtrB, PII also regulates the NtrC-NtrB two-component system, which controls transcription of the GS encoding glnA gene. Under N excess, unmodified PII promotes NtrB-dependent dephosphorylation of the response regulator NtrC, which prevents activation of glnA expression [38,39].

Figure 3.
Regulatory network of GS in Synechocystis sp. PCC 6803. Increase in 2OG abundance during N deprivation promotes NtcA activation directly and via interaction with PipX. Active NtcA boosts transcription of the glnA gene encoding the GS monomers and of the sRNA NsiR4, which inhibits IF7 synthesis by interacting with the gifA mRNA. Simultaneously, transcription of the IF encoding gif genes is repressed by NtcA binding. N excess causes 2OG depletion, which results in PipX-PII complex formation and inactivation of NtcA. Thus, IFs accumulate and inactivate GS in a concentration-dependent manner due to absent NtcA-mediated repression. Furthermore, N availability is also sensed by a glutamine riboswitch within the 5’UTR of gifB, which controls the synthesis of IF17 (gifB). IF: inactivating factor, P: promoter.
Figure 3.
Regulatory network of GS in Synechocystis sp. PCC 6803. Increase in 2OG abundance during N deprivation promotes NtcA activation directly and via interaction with PipX. Active NtcA boosts transcription of the glnA gene encoding the GS monomers and of the sRNA NsiR4, which inhibits IF7 synthesis by interacting with the gifA mRNA. Simultaneously, transcription of the IF encoding gif genes is repressed by NtcA binding. N excess causes 2OG depletion, which results in PipX-PII complex formation and inactivation of NtcA. Thus, IFs accumulate and inactivate GS in a concentration-dependent manner due to absent NtcA-mediated repression. Furthermore, N availability is also sensed by a glutamine riboswitch within the 5’UTR of gifB, which controls the synthesis of IF17 (gifB). IF: inactivating factor, P: promoter.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bolay, P.; Muro-Pastor, M.I.; Florencio, F.J.; Klähn, S. The Distinctive Regulation of Cyanobacterial Glutamine Synthetase. Life 2018, 8, 52. https://doi.org/10.3390/life8040052
AMA Style
Bolay P, Muro-Pastor MI, Florencio FJ, Klähn S. The Distinctive Regulation of Cyanobacterial Glutamine Synthetase. Life. 2018; 8(4):52. https://doi.org/10.3390/life8040052
Chicago/Turabian StyleBolay, Paul, M. Isabel Muro-Pastor, Francisco J. Florencio, and Stephan Klähn. 2018. "The Distinctive Regulation of Cyanobacterial Glutamine Synthetase" Life 8, no. 4: 52. https://doi.org/10.3390/life8040052
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.