Density-Dependent Effects of Amphibian Prey on the Growth and Survival of an Endangered Giant Water Bug

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
: Amphibian predator-insect prey relationships are common in terrestrial habitats, but amphibian larvae are preyed upon by a variety of aquatic hemipterans in aquatic habitats. This paper suggests that the survival of the nymphs of the endangered aquatic hemipteran Kirkaldyia (=Lethocerus) deyrolli (Belostomatidae: Heteroptera) is directly and indirectly affected by the abundance of their amphibian larval prey (tadpoles). Young nymphs of K. deyrolli mainly feed on tadpoles, regardless of differences in prey availability. Nymphs provided with tadpoles grow faster than nymphs provided with invertebrate prey. Therefore, tadpole consumption seems to be required to allow the nymphs to complete their larval development. In addition, the survival of K. deyrolli nymphs was greater during the period of highest tadpole density (June) than during a period of low tadpole density (July). Higher tadpole density moderates predation pressure from the water scorpion Laccotrephes japonensis (Nepidae: Heteroptera) on K. deyrolli nymphs; i.e., it has a density-mediated indirect effect. These results suggest that an abundance of tadpoles in June provides food for K. deyrolli nymphs (a direct bottom-up effect) and moderates the predation pressure from L. japonensis (an indirect bottom-up effect). An abundance of amphibian prey is indispensable for the conservation of this endangered giant water bug species.1. Introduction
1.1. Trophic Interactions of Aquatic Insects and Amphibians
The trophic interactions of insects and amphibians have received a significant amount of attention by researchers. Amphibian adults eat a variety of terrestrial arthropods. Insects with a range of gape sizes [1,2] that co-exist in the same habitats as amphibians are exposed to high predation pressure from amphibians [3]. Generally, although amphibians (mainly those of the post-metamorphic stage) are sometimes preyed on by insects ([4], reviewed in Toledo [5]), amphibian predator–insect prey relationships are common in terrestrial habitats. In contrast, these predator–prey relationships are reversed for larval amphibians in aquatic habitats; i.e., amphibian larvae are preyed upon by a variety of aquatic insects such as Coleoptera [6], Heteroptera [7], and Odonata [8]. Amphibian larvae are keystone organisms for aquatic communities because they are important prey resources for aquatic insects. Insect predator-anuran larvae prey relationships in aquatic environments are appropriate model systems for examining predator–prey relationships.
Amphibian larvae show anti-predatory behavior (low activity) and morphological changes (larger heads) in response to predators in order to increase their chances of survival [9-16]. In a study of anurans, Vonesh et al. reported that reductions in larval density and size due to egg-stage predators facilitate larval survival in the presence of aquatic predators (predator-induced hatching plasticity) [17-19]. Thus, insect predator-anuran larvae prey relationships have been studied from the viewpoint of behavior and phenotypic plasticity in anuran larvae. However, very few studies have investigated the density-mediated indirect effects of insect predator–anuran larvae prey relationships. Here, I focused on insect predator–anuran larvae prey relationships from the viewpoint of density-mediated indirect effects.
1.2. Are There any Aquatic Heteropteran Predators of Amphibian Larvae?
Generally, dragonfly nymphs are mainly used as insect predators of amphibians in model predator–prey systems [9-12]. In addition, aquatic hemipterans play a significant role as the major component of the aquatic fauna of aquatic environments that are devoid of fish [20,21] and are often at the top of the food chain in such aquatic communities, preying upon a variety of aquatic animals [22,23]. As with Odonata predators, some researchers studied phenotypic changes in amphibians induced by aquatic hemipterans predators [24-26]. Certain environments also reduce predation by aquatic hemipteran predators. Kopp et al. [28] showed aquatic vegetation reduces predation rates on tadpoles by aquatic hemipteran predators. Swart and Taylor [27] revealed that tadpoles switched their preference away from black backgrounds in response to chemical signals from a predator (aquatic hemipterans) because the predator killed significantly more tadpoles on dark backgrounds than on light backgrounds. Therefore, aquatic hemipteran predators are regarded as important predators for anuran larvae.
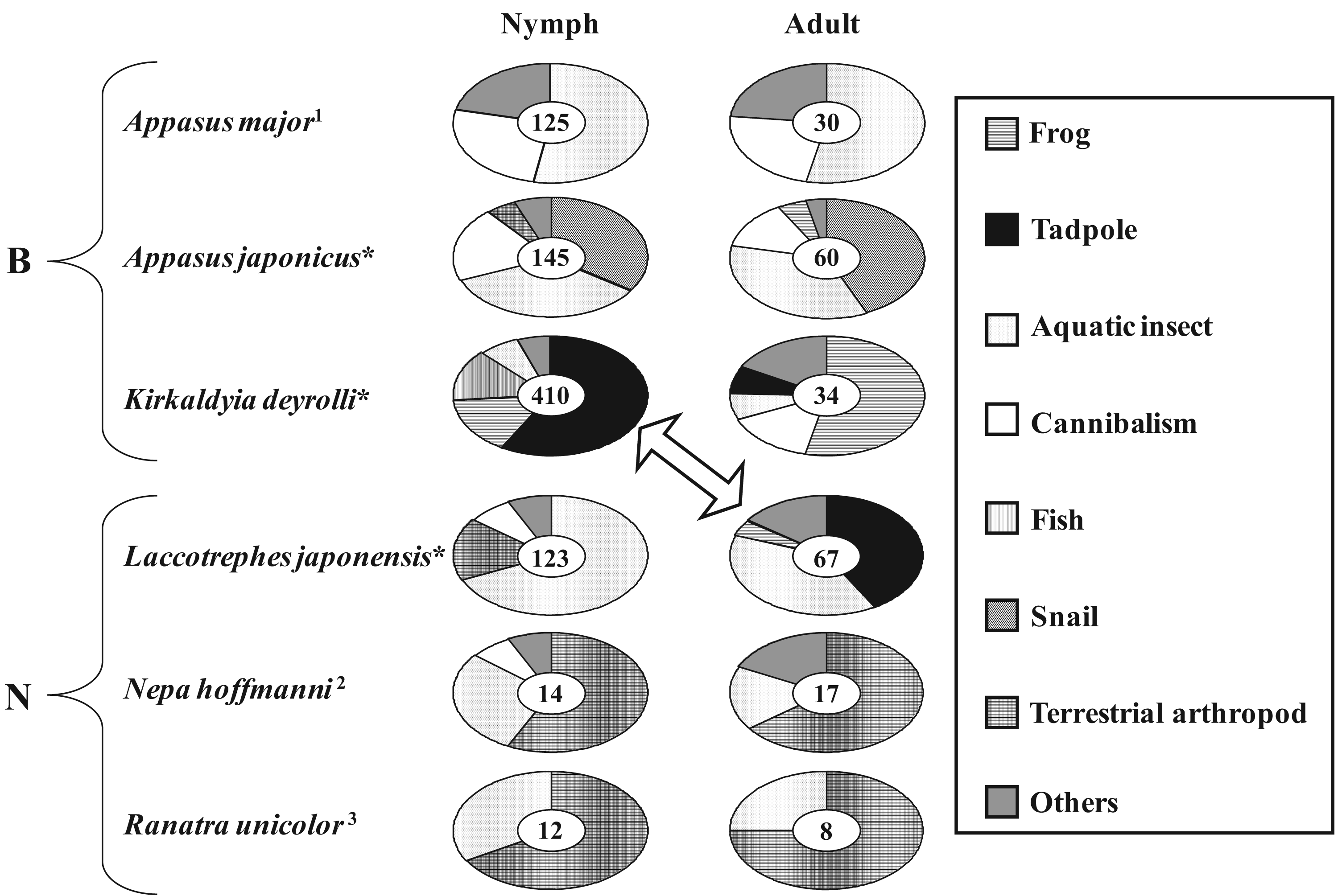
However, most aquatic hemipterans do not often eat tadpoles in Japan. Ohba and Nakasuji [7] investigated the feeding habits of aquatic bugs (Nepoidea, including Belostomatidae and Nepidae) by performing direct observations in wetland areas (see Figure 1) and obtaining data from the published literature [29-31]. As a result, it was found that sympatric species (Appasus japonicus, Kirkaldyia deyrolli, and Laccotrephes japonensis) displayed differences in their dietary components (Figure 2). Although tadpoles are preyed upon by a variety of aquatic insects, not all aquatic insect species eat tadpoles. Only K. deyrolli nymphs and L. japonensis adults greatly depend on tadpoles whereas A. japonicus does not eat tadpoles. Therefore, K. deyrolli nymphs and L. japonensis adults seem to be members of the same guild; i.e., they compete with each other, in Japanese wetlands [32].
In this paper, I first introduce that K. deyrolli nymphs feed on tadpoles. Second, I introduce that K. deyrolli nymph survival is indirectly affected by tadpoles; i.e., by a density-mediated indirect effect. Finally, I discuss aquatic heteropteran predator–amphibian larval prey relationships in relation to temporal dynamic interactions.
2. Tadpole-Feeding by K. deyrolli Nymphs
The quantity and quality of prey animals strongly affect a predator’s life history and abundance. Among predacious insects, predatory species that depend upon a particular prey animal, which are known as specialists, breed when their prey animal is abundant [33-36]. Specialist development coincides with the appearance of specific prey animals, especially during the nymphal period [37]. In rice fields, K. deyrolli nymphs prey upon tadpoles more than on other kinds of prey (Figure 2). The subfamily Lethocerinae, which has the largest body size among Belostomatidae, is known to be a vertebrate specialist [38-40]. Why K. deyrolli nymphs mostly feed on tadpoles is not well understood from the viewpoint of nymphal growth and seasonal occurrence.
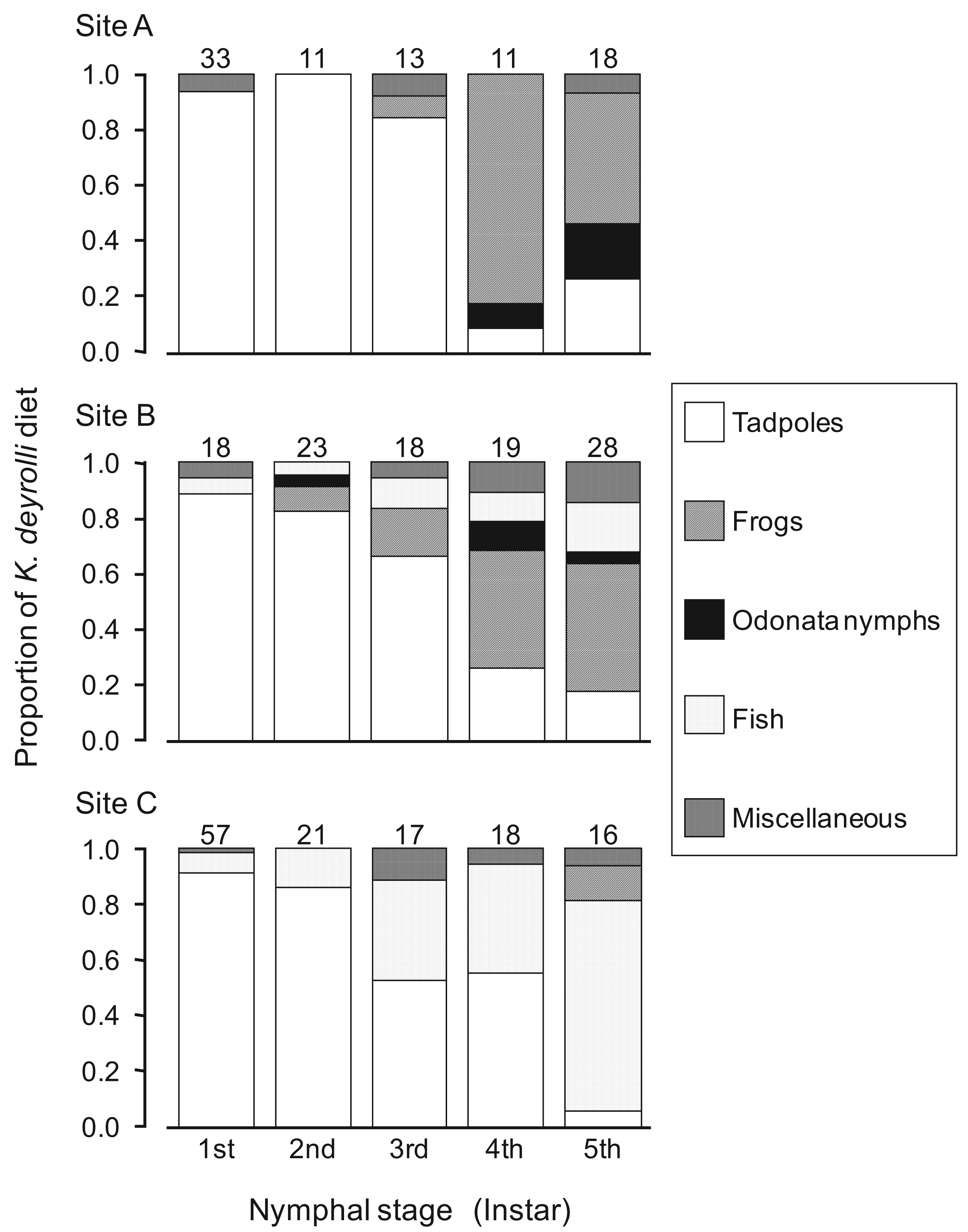
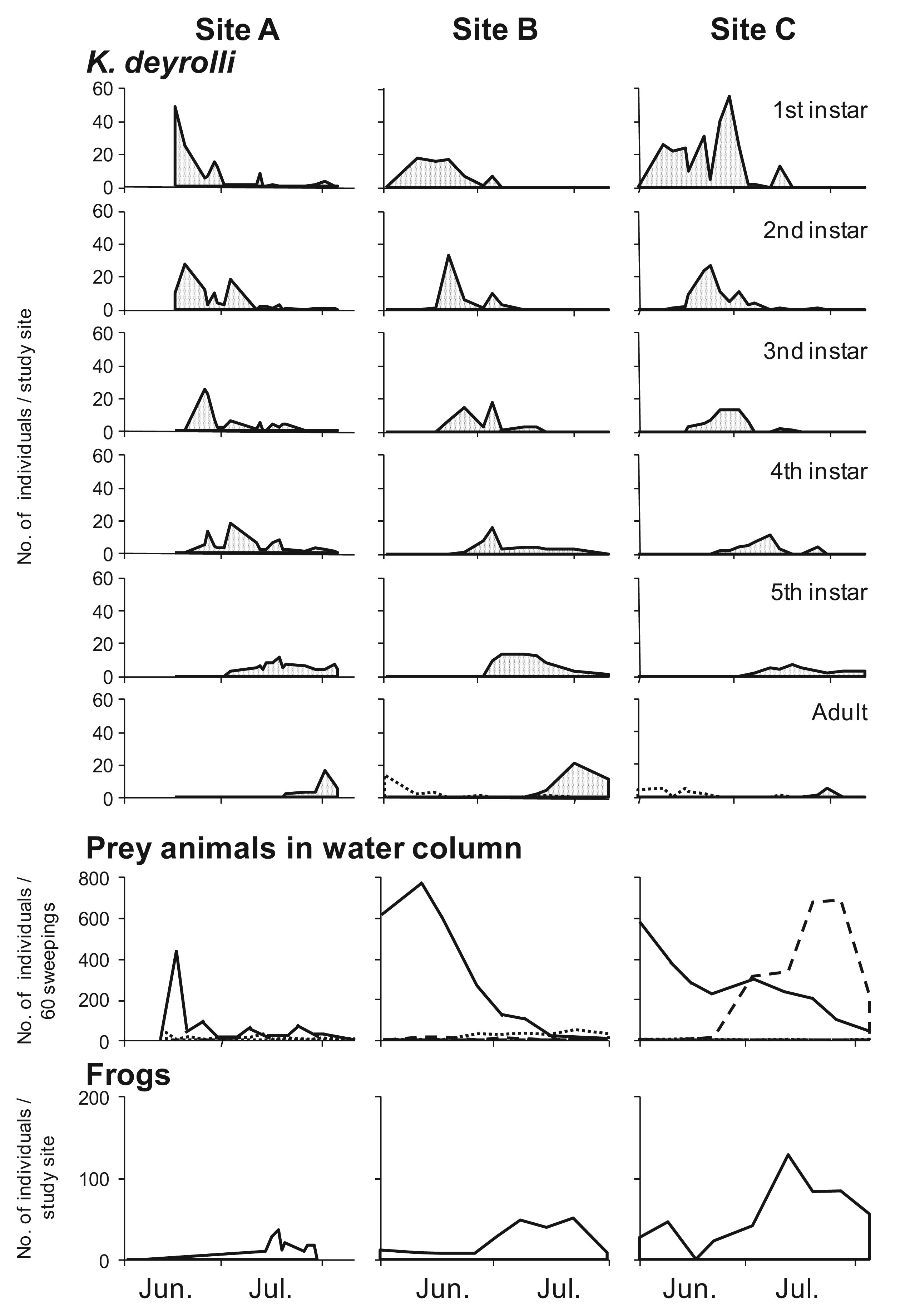
In general, the appearance of younger nymphs of predacious insects when prey animals are abundant is expected to moderate cannibalism due to food shortages [41-44]. Accordingly, Ohba et al. [45] studied the ontogenetic diet shift of K. deyrolli by quantifying instar abundance and analyzing captured prey and prey relative abundance in rice fields in three localities. The first to third-instar K. deyrolli nymphs mainly fed on tadpoles, regardless of differences in prey availability among the three localities (Figure 3). A rearing experiment demonstrated that K. deyrolli nymphs provided with tadpoles displayed greater growth rates at all nymphal stages, except for the final stage, than nymphs fed on dragonflynymphs. The emergence of young K. deyrolli nymphs seemed to coincide with the period when tadpoles became abundant in the rice fields (Figure 4). In addition, the appearance of younger K. deyrolli nymphs when tadpoles are abundant is expected to moderate cannibalism due to food shortages, as has been demonstrated for other predatory insects such as ladybirds [41-44]. Actually, the frequency of cannibalism in K. deyrolli nymphs is lower than that seen in A. japonicus nymphs in the field (Figure 2) [7].
3. Density-Mediated Indirect Effects of Tadpole Prey
Many specialists breed when certain prey are abundant in order to increase the chance of there being a sufficient amount of food to increase the growth and survival of their young, showing that a high prey density directly supports the predator population, e.g., a bottom-up effect. However, there is little information on the indirect role of higher prey density on the mediation of predation pressure on the predator from other predators within the same guild; i.e., density-mediated indirect effects, in aquatic environments including insect predator–anuran larva prey relationships. As mentioned above, K. deyrolli nymphs are considered to feed on tadpoles [45]. In addition, L. japonensis prey upon K. deyrolli nymphs [32,46] and tadpoles in rice fields [7]. Therefore, L. japonensis adults are intraguild predators of K. deyrolli nymphs. Consequently, it is predicted that the survivorship of K. deyrolli nymphs is affected by their common prey, tadpoles, via a bottom-up effect on survivorship as well as by their predator, L. japonensis adults, via a top-down effect. Thus, it is considered that density-mediated indirect relationships exist among the three animal species.
Ohba and Nakasuji [47] demonstrated that the survival of K. deyrolli nymphs, which are tadpole specialists, is affected by tadpoles (a direct bottom-up effect). We investigated the survival rates of the first instar nymphs of K. deyrolli in June (high tadpole density period) and July (low tadpole density period) using the Kiritani-Nakasuji-Manly method [48,49]. As circumstantial evidence, the survival rate of the first instar nymphs of K. deyrolli in a rice paddy field was higher in June (75.1%) than in July (54.2%) [47]. The differences in prey density between June and July probably affected the survival rates of the K. deyrolli nymphs, suggesting that bottom-up effects display seasonal variation.
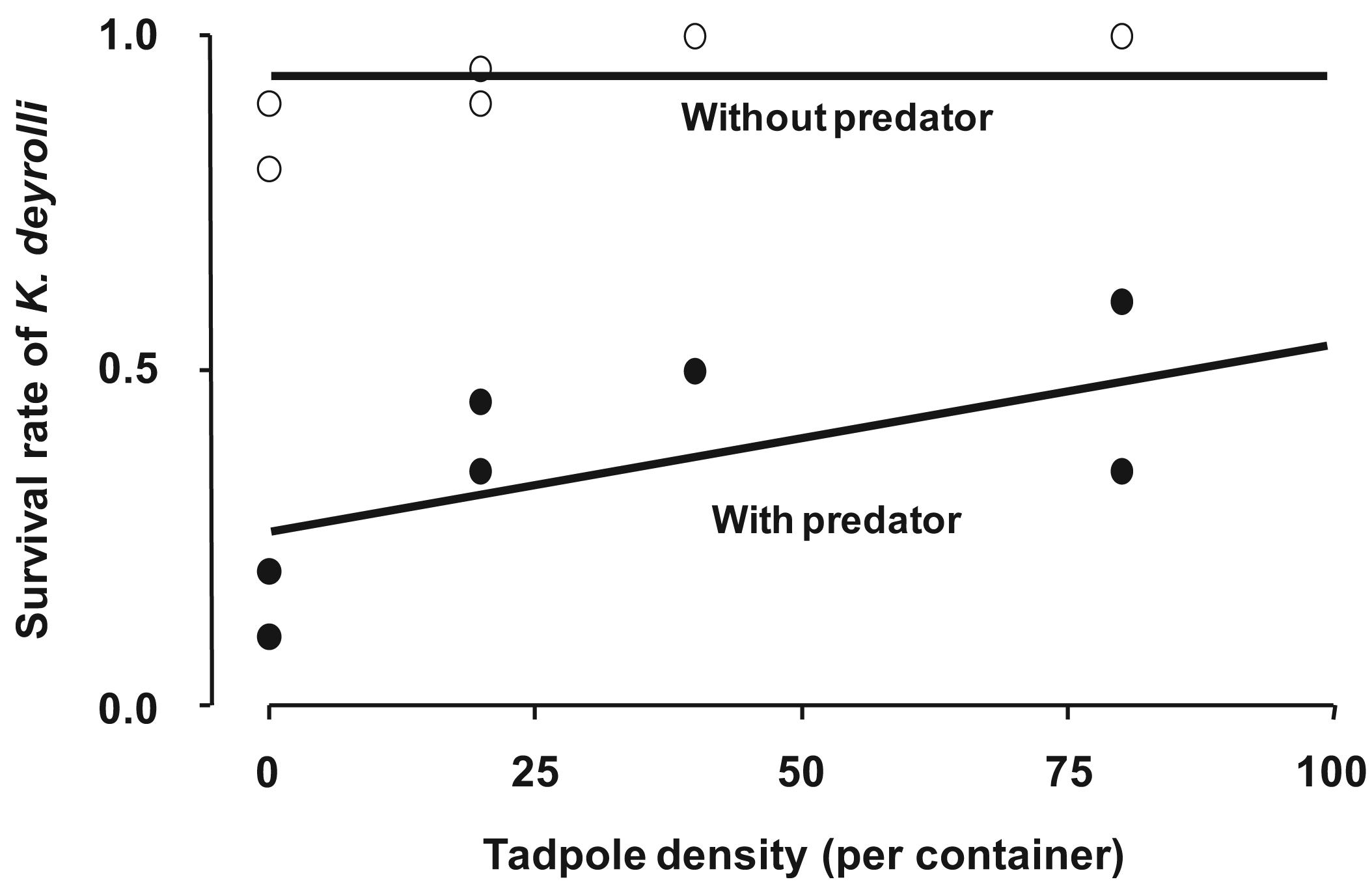
To examine whether a higher tadpole density moderates the predation pressure from L. japonensis adults on K. deyrolli nymphs, a field experiment was conducted in a rice field. As a result, it was found that a higher tadpole density moderated the predation pressure from L. japonensis adults on K. deyrolli nymphs (a density-mediated indirect effect; Figures 5 and 6) [50,51]. Prey abundance is an important factor for determining the frequency of intraguild predation [52-54], especially in terrestrial aphidophagous predator–aphid prey relationships. Thus, in rice fields the incidence of intraguild predation by L. japonensis adults might increase when the tadpole density decreases.
4. Conservation of an Endangered Giant Water Bug
The Japanese populations of K. deyrolli have decreased sharply during the last four decades, and this species is now included in the Red Data List of species in 45 of 47 Japanese prefectures [55,56]. Contributing factors such as decreases in suitable aquatic habitats, water pollution, and urbanization have been investigated and verified in previous studies [57-59]. In addition to these factors, it is important for the conservation of K. deyrolli to reveal the best food for them to eat. Regarding bottom-up effects, Hirai and Hidaka [60] and Hirai [61] emphasized that the frog population is very important for the conservation of the K. deyrolli population because frogs are major constituents of the diet of K. deyrolli adults.
Previous studies have only focused on direct bottom-up effects of amphibian prey. However, the present study demonstrated an indirect bottom-up effect of amphibian prey. The emergence of young K. deyrolli nymphs appeared to coincide with the period when tadpoles became abundant in the rice fields, which allowed the young K. deyrolli nymphs to achieve greater growth (Figure 4 [45]). This indicates that a high prey density directly supports K. deyrolli nymphs. In addition to such direct effects, a high tadpole density was also demonstrated to have a beneficial indirect effect on the survival of K. deyrolli nymphs (Figure 6 [47]). Therefore, an abundance of amphibian prey is indispensable for the conservation of this endangered giant water bug species.
5. Conclusions
This paper introduced that young K. deyrolli nymphs mainly feed on tadpoles in rice fields and that nymphs provided with tadpoles grow faster than nymphs provided with invertebrate prey. A higher tadpole density moderates the predation pressure from the water scorpion L. japonensis on K. deyrolli nymphs; i.e., it has a density-mediated indirect effect. These results suggest that an abundance of tadpoles provides sufficient food for K. deyrolli nymph growth (a direct bottom-up effect) and moderates the predation pressure exerted on them by L. japonensis (an indirect bottom-up effect). An abundance of amphibian prey is indispensable for the conservation of this endangered giant water bug species.






References
- Toft, C.A. Feeding ecology of thirteen syntopic species of anurans in a seasonal tropical environment. Oecologia 1980, 45, 131–141. [Google Scholar]
- Hirai, T. Ontogenetic change in the diet of the pond frog, Rana nigromaculata. Ecol. Res. 2002, 17, 639–644. [Google Scholar]
- Honma, A.; Oku, S.; Nishida, T. Adaptive significance of death feigning posture as a specialized inducible defence against gape-limited predators. Proc. R. Soc. Biol. Sci. Ser. B 2006, 273, 1631–1636. [Google Scholar]
- Benard, M. Predators and mates: Conflicting selection on the size of male Pacific treefrogs (Pseudacris regilla). J. Herpetol. 2007, 41, 317–320. [Google Scholar]
- Toledo, L. Predation on seven South American anuran species by water bugs (Belostomatidae). Phyllomedusa 2003, 2, 105–108. [Google Scholar]
- Ohba, S. Ontogenetic dietary shift in the larvae of Cybister japonicus (Coleoptera: Dytiscidae) in Japanese rice fields. Environ. Entomol. 2009, 38, 856–860. [Google Scholar]
- Ohba, S.; Nakasuji, F. Dietary items of predacious aquatic bugs (Nepoidea: Heteroptera) in Japanese wetlands. Limnology 2006, 7, 41–43. [Google Scholar]
- Corbet, P.S. Dragonflies: Behaviour and Ecology of ODONATA; Cornell University Press: Ithaca, NY, USA, 1999. [Google Scholar]
- van Buskirk, J.; McCollum, S. Plasticity and selection explain variation in tadpole phenotype between ponds with different predator composition. Oikos 1999, 85, 31–39. [Google Scholar]
- Van Buskirk, J.; Relyea, R. Selection for phenotypic plasticity in Rana sylvatica tadpoles. Biol. J. Linn. Soc. 1998, 65, 301–328. [Google Scholar]
- van Buskirk, J. Phenotypic lability and the evolution of predator-induced plasticity in tadpoles. Evolution 2002, 56, 361–370. [Google Scholar]
- McIntyre, P.; Baldwin, S.; Flecker, A. Effects of behavioral and morphological plasticity on risk of predation in a Neotropical tadpole. Oecologia 2004, 141, 130–138. [Google Scholar]
- Kishida, O.; Nishimura, K. Bulgy tadpoles: Inducible defense morph. Oecologia 2004, 140, 414–421. [Google Scholar]
- Takahara, T.; Kohmatsu, Y.; Maruyama, A.; Yamaoka, R. Benefit of suites of defensive behavior induced by predator chemical cues on anuran tadpoles. Hyla Jpn. Behav. Ecol. Sociobiol. 2008, 63, 235–240. [Google Scholar]
- Hettyey, A.; Zsarnoczai, S.; Vincze, K.; Hoi, H.; Laurila, A. Interactions between the information content of different chemical cues affect induced defences in tadpoles. Oikos 2010, 119, 1814–1822. [Google Scholar]
- Jara, F.G.; Perotti, M.G. Toad tadpole responses to predator risk: Ontogenetic change between constitutive and inducible defenses. J. Herpetol. 2009, 43, 82–88. [Google Scholar]
- Vonesh, J.R. Egg predation and predator-induced hatching plasticity in the African reed frog, Hyperolius spinigularis. Oikos 2005, 110, 241–252. [Google Scholar]
- Vonesh, J.R. Sequential predator effects across three life stages of the African tree frog, Hyperolius spinigularis. Oecologia 2005, 143, 280–290. [Google Scholar]
- Vonesh, J.R.; Bolker, B.M. Compensatory larval responses shift trade-offs associated with predator-induced hatching plasticity. Ecology 2005, 86, 1580–1591. [Google Scholar]
- Runck, C.; Blinn, D.W. Role of Belostoma bakeri (Heteroptera) in the trophic ecology of a fishless desert spring. Limnol. Oceanogr. 1994, 39, 1800–1812. [Google Scholar]
- Blaustein, L. Influence of the predatory backswimmer, Notonecta maculata, on invertebrate community structure. Ecol. Entomol. 1998, 23, 246–252. [Google Scholar]
- Waters, T.F. Secondary production in inland waters 1. Adv. Ecol. Res. 1977, 10, 91–164. [Google Scholar]
- Runck, C.; Blinn, W. Population dynamics and secondary production by Ranatra montezuma (Heteroptera: Nepidae). J. North. Am. Benthol. Soc. 1990, 9, 262–270. [Google Scholar]
- Gomez, V.I.; Kehr, A.I. Morphological and developmental responses of anuran larvae (Physalaemus albonotatus) to chemical cues from the predators Moenkhausia dichoroura (Characiformes: Characidae) and Belostoma elongatum (Hemiptera: Belostomatidae). Zool. Stud. 2011, 50, 203–210. [Google Scholar]
- Kehr, A.I.; Gómez, V.I. Intestinal, body and tail plasticity in Rhinella schneideri (Bufonidae) tadpoles induced by a predator insect (Belostoma elegans). Adv. Stud. Biol. 2009, 1, 85–94. [Google Scholar]
- Hoverman, J.T.; Relyea, R.A. Temporal environmental variation and phenotypic plasticity: A mechanism underlying priority effects. Oikos 2008, 117, 23–32. [Google Scholar]
- Swart, C.C.; Taylor, R.C. Behavioral interactions between the giant water bug (Belostoma lutarium) and tadpoles of Bufo woodhousii. Southeast. Nat. 2004, 3, 13–24. [Google Scholar]
- Kopp, K.; Wachlevski, M.; Eterovick, P.C. Environmental complexity reduces tadpole predation by water bugs. Can. J. Zool. 2006, 84, 136–140. [Google Scholar]
- Okada, H.; Nakasuji, F. Comparative studies on the seasonal occurrence, nymphal development and food menu in two giant water bugs, Diplonychus japonicus Vuillefroy and Diplonychus major Esaki (Hemiptera: Belostomatidae). Res. Popul. Ecol. 1993, 35, 15–22. [Google Scholar]
- Ban, Y. Some observation on the life cycle of the water scorpion, Ranatra unicolor Scott (Hemiptera: Nepidae). Yamanoshita Bay, Lake Biwa. Verh. Int. Verein. Limnol. 1981, 21, 1621–1625. [Google Scholar]
- Ban, Y. Water Scorpion NEPA Hoffmani; Bun-ichi Sogo Shuppan: Tokyo, Japan, 1988. [Google Scholar]
- Ohba, S. Notes on predators and their effect on the survivorship of the endangered giant water bug, Kirkaldyia (=Lethocerus) deyrolli (Heteroptera, Belostomatidae), in Japanese rice fields. Hydrobiologia 2007, 583, 377–381. [Google Scholar]
- Evans, E. Feeding specialization in predatory insects: Hunting and attack behavior of two stinkbug species (Hemiptera: Pentatomidae). Am. Midl. Nat. 1982, 108, 96–104. [Google Scholar]
- Sota, T. Activity patterns, diets and interspecific interactions of coexisting spring and autumn breeding carabids: Carabus yaconinus and Leptocarabus kumagaii (Coleoptera, Carabidae). Ecol. Entomol. 1985, 10, 315–324. [Google Scholar]
- Hagen, K.S. Biology and ecology of predaceous Coccinellidae. Annu. Rev. Entomol. 1962, 7, 289–326. [Google Scholar]
- Albuquerque, G.; Tauber, M.; Tauber, C. Life-history adaptations and reproductive costs associated with specialization in predacious insects. J. Anim. Ecol. 1997, 66, 307–317. [Google Scholar]
- Elkinton, J.S. Population Ecology. In Encyclopedia of Insects; Resh, V.H., Carde, R.T., Eds.; Academic press: San Diego, CA, USA, 2003; pp. 933–947. [Google Scholar]
- Smith, R.L. Evolution of Paternal Care in the Giant Water Bugs (Heteroptera: Belostomatidae). In The evolution of social behavior in insects and arachnids; Choe, J., Crespi, B., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 116–149. [Google Scholar]
- Swart, C.C.; Deaton, L.E.; Felgenhauer, B.E. The salivary gland and salivary enzymes of the giant waterbugs (Heteroptera; Belostomatidae). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 145, 114–122. [Google Scholar]
- Ohba, S. Field observation of predation on a turtle by a giant water bug. Entomol. Sci. 2011, 14, 364–365. [Google Scholar]
- Takahashi, K. Intra- and inter-specific predations of lady beetles in spring alfalfa fields. Jpn. J. Entomol. 1989, 57, 199–203. [Google Scholar]
- Osawa, N. A life table of the ladybird beetle Harmonia axyridis Pallas (Coleoptera, Coccinellidae) in relation to the aphid abundance. Jpn. J. Entomol. 1992, 60, 575–579. [Google Scholar]
- Agarwala, B.; Dixon, A. Laboratory study of cannibalism and interspecific predation in ladybirds. Ecol. Entomol. 1992, 17, 303–309. [Google Scholar]
- Hironori, Y.; Katsuhiro, S. Cannibalism and interspecific predation in two predatory ladybirds in relation to prey abundance in the field. Entomophaga 1997, 42, 153–163. [Google Scholar]
- Ohba, S.; Miyasaka, H.; Nakasuji, F. The role of amphibian prey in the diet and growth of giant water bug nymphs in Japanese rice fields. Popul. Ecol. 2008, 50, 9–16. [Google Scholar]
- Ohba, S.; Swart, C.C. Intraguild predation of water scorpion Laccotrephes japonensis (Nepidae: Heteroptera). Ecol. Res. 2009, 24, 1207–1211. [Google Scholar]
- Ohba, S.; Nakasuji, F. Density-mediated indirect effects of a common prey tadpole on interaction between two predatory bugs: Kirkaldyia deyrolli and Laccotrephes japonensis. Popul. Ecol. 2007, 49, 331–336. [Google Scholar]
- Kiritani, K.; Nakasuji, F. Estimation of the stage-specific survival rate in the insect population with overlapping stages. Res. Popul. Ecol. 1967, 9, 143–152. [Google Scholar]
- Manly, B.F.J. Extensions to Kiritani and Nakasuji's method for analysing insect stage-frequency data. Res. Popul. Ecol. 1976, 17, 191–199. [Google Scholar]
- Holt, R.; Lawton, J. The ecological consequences of shared natural enemies. Annu. Rev. Ecol. Syst. 1994, 25, 495–520. [Google Scholar]
- Holt, R.D. Predation, apparent competition, and the structure of prey communities. Theor. Popul. Biol. 1977, 12, 197–229. [Google Scholar]
- Polis, G.; Myers, C.; Holt, R. The ecology and evolution of intraguild predation: Potential competitors that eat each other. Annu. Rev. Ecol. Syst. 1989, 20, 297–330. [Google Scholar]
- Lucas, E.; Coderre, D.; Brodeur, J. Intraguild predation among aphid predators: Characterization and influence of extraguild prey density. Ecology 1998, 79, 1084–1092. [Google Scholar]
- Hodge, M.A. The implications of intraguild predation for the role of spiders in biological control. J. Arachnol. 1999, 27, 351–362. [Google Scholar]
- Search system of Japanese red data. Available online: http://www.jpnrdb.com/index.html (accessed on 16 May 2011).
- Japan Environment Agency. Threatened Wildlife of Japan. Red Data Book, 2nd ed.; Environment Agency of Japan: Tokyo, Japan, 2000; in Japanese. [Google Scholar]
- Ho, C.; Kim, H.; Kim, J.G. Landscape analysis of the effects of artificial lighting around wetland habitats on the giant water bug Lethocerus deyrollei in Jeju Island. J. Ecol. Field. Biol. 2009, 32, 83–86. [Google Scholar]
- Yoon, T.; Kim, D.; Kim, S.; Jo, S.; Bae, Y. Light-attraction flight of the giant water bug, Lethocerus deyrolli (Hemiptera: Belostomatidae), an endangered wetland insect in east Asia. Aquat. Insects 2010, 32, 195–203. [Google Scholar]
- Nagaba, Y.; Tufail, M.; Inui, H.; Takeda, M. Hormonal regulation and effects of four environmental pollutants on vitellogenin gene transcription in the giant water bug, Lethocerus deyrollei (Hemiptera: Belostomatidae). J. Insect Conserv. 2011, 15, 421–431. [Google Scholar]
- Hirai, T.; Hidaka, K. Anuran-dependent predation by the giant water bug, Lethocerus deyrollei (Hemiptera: Belostomatidae), in rice fields of Japan. Ecol. Res. 2002, 17, 655–661. [Google Scholar]
- Hirai, T. Diet composition of the endangered giant water bug Lethocerus deyrolli (Hemiptera: Belostomatidae) in the rice fields of Japan: Which is the most important prey item among frogs, fish, and aquatic insects? Entomol. Sci. 2007, 10, 333–336. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ohba, S.-y. Density-Dependent Effects of Amphibian Prey on the Growth and Survival of an Endangered Giant Water Bug. Insects 2011, 2, 435-446. https://doi.org/10.3390/insects2040435
Ohba S-y. Density-Dependent Effects of Amphibian Prey on the Growth and Survival of an Endangered Giant Water Bug. Insects. 2011; 2(4):435-446. https://doi.org/10.3390/insects2040435
Chicago/Turabian StyleOhba, Shin-ya. 2011. "Density-Dependent Effects of Amphibian Prey on the Growth and Survival of an Endangered Giant Water Bug" Insects 2, no. 4: 435-446. https://doi.org/10.3390/insects2040435
APA StyleOhba, S.-y. (2011). Density-Dependent Effects of Amphibian Prey on the Growth and Survival of an Endangered Giant Water Bug. Insects, 2(4), 435-446. https://doi.org/10.3390/insects2040435