Transmission of a Gammabaculovirus within Cohorts of Balsam Fir Sawfly (Neodiprion abietis) Larvae

{kind=link}
{kind=link}
Abstract
:1. Introduction
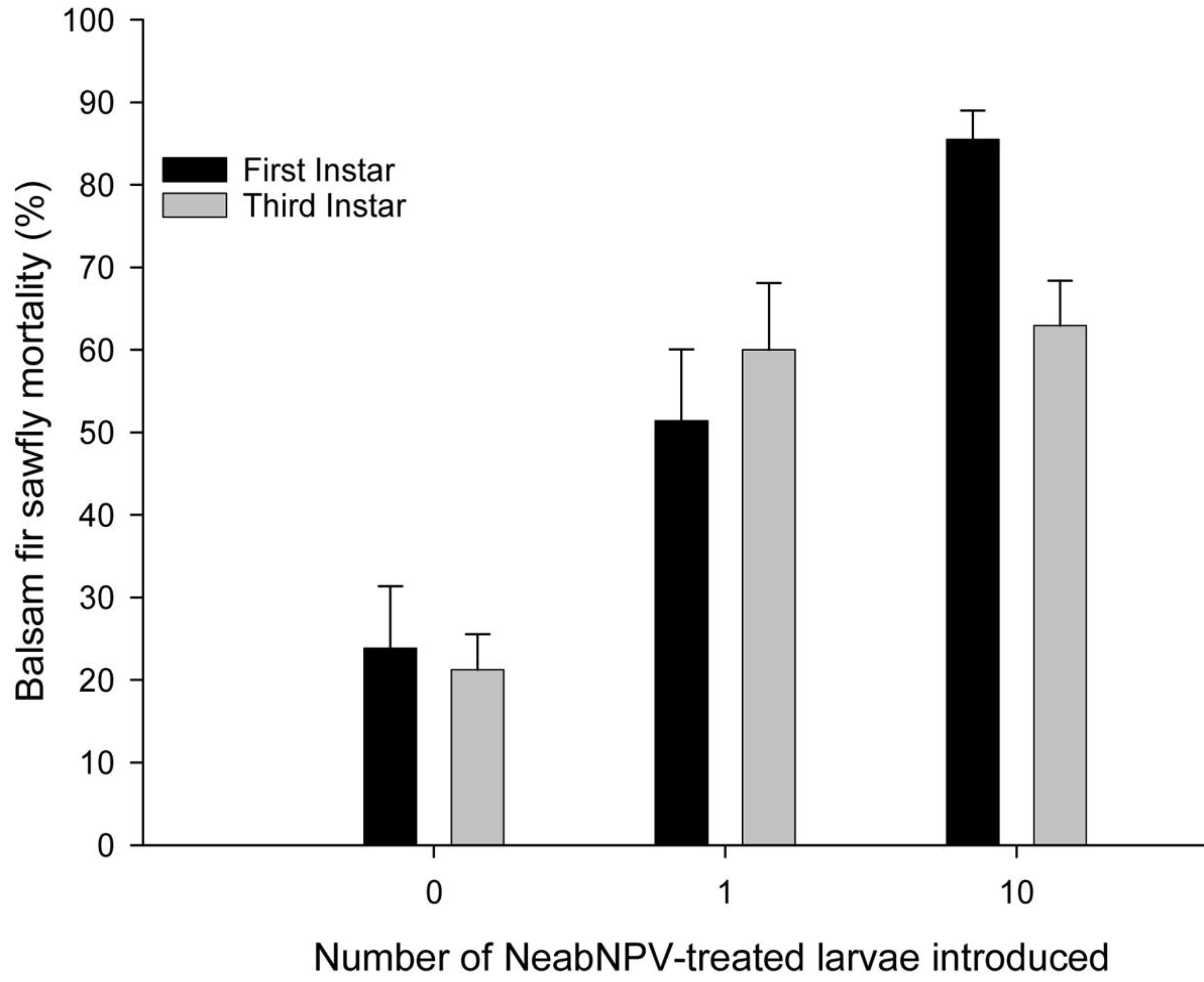
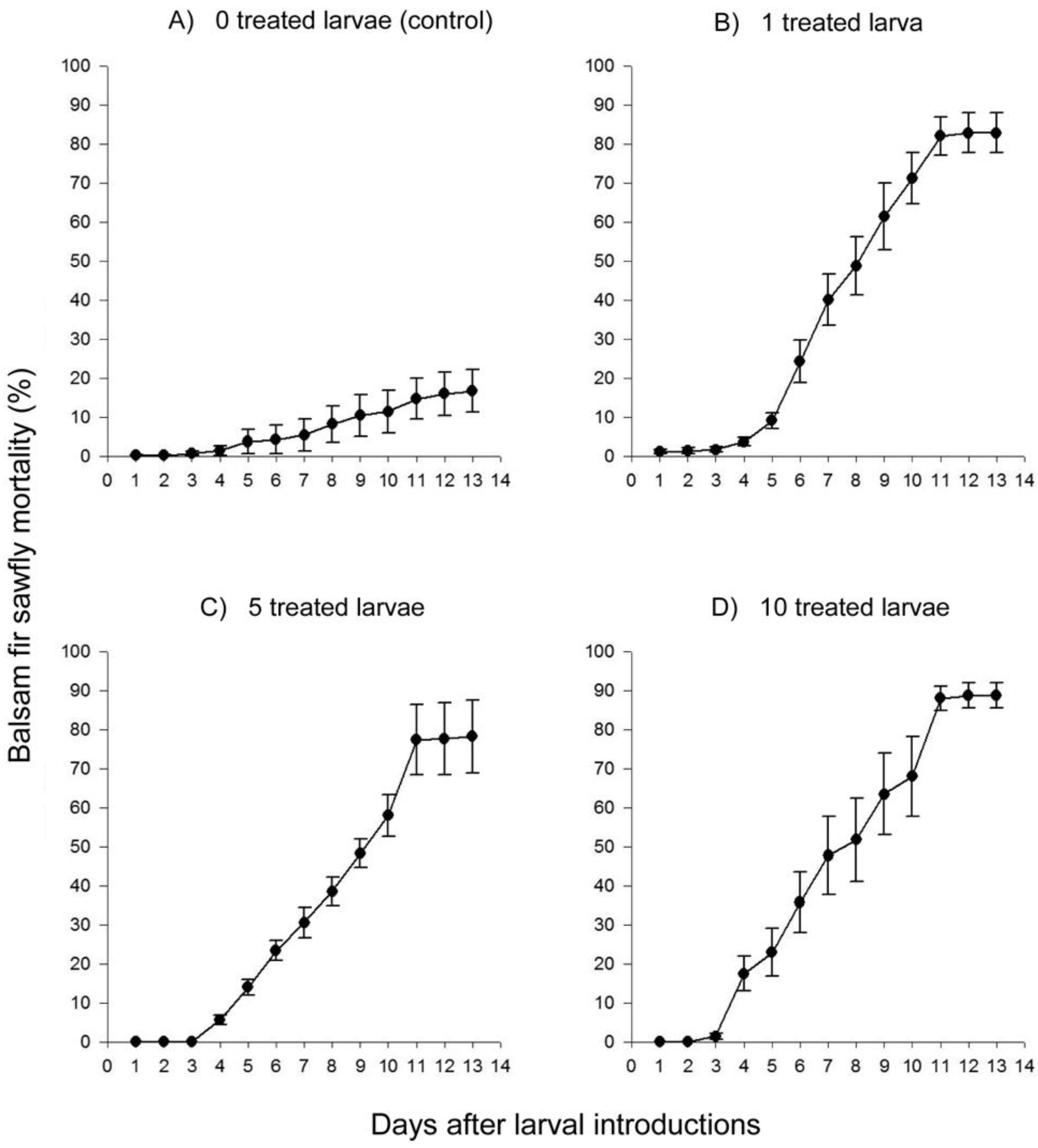
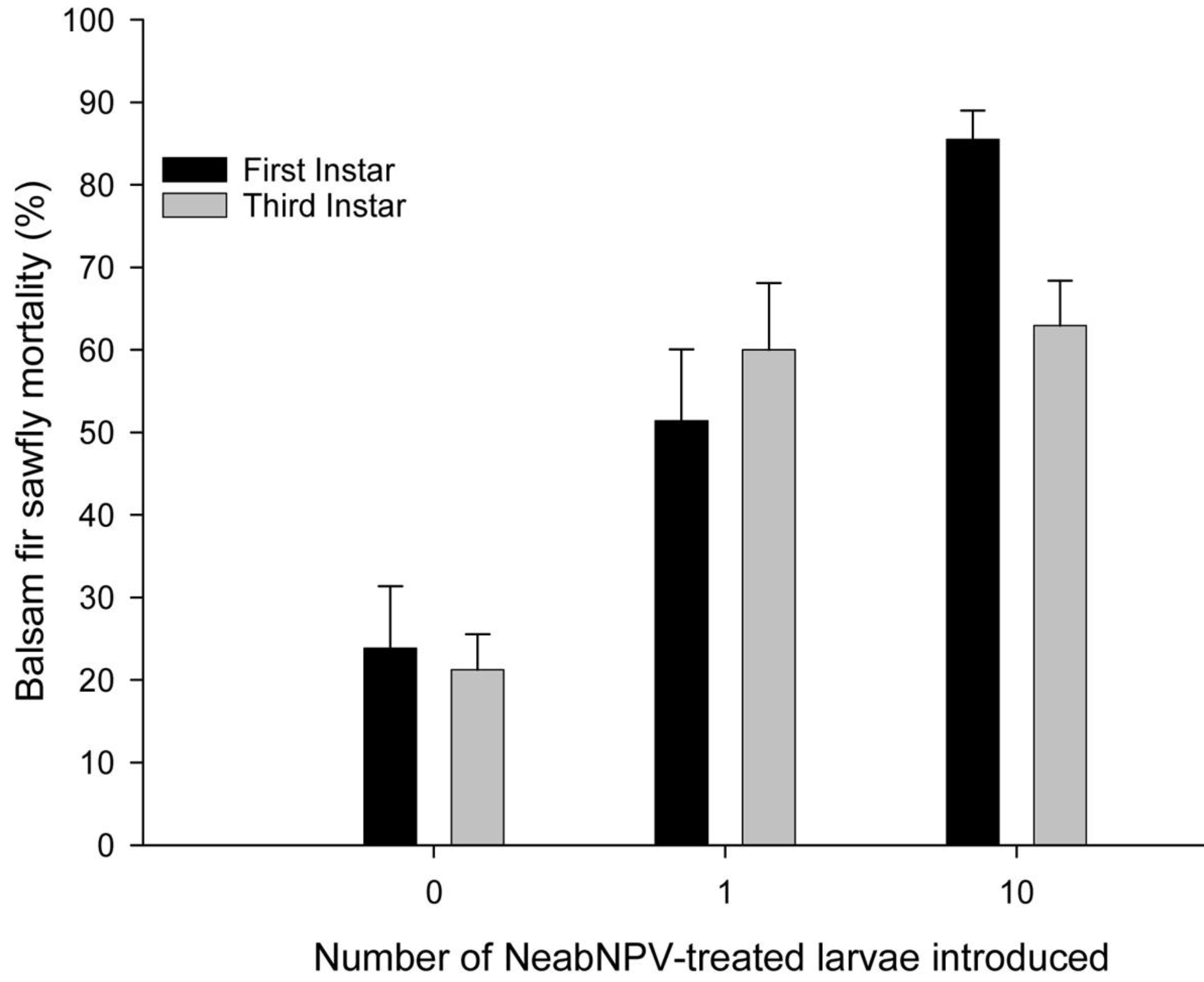
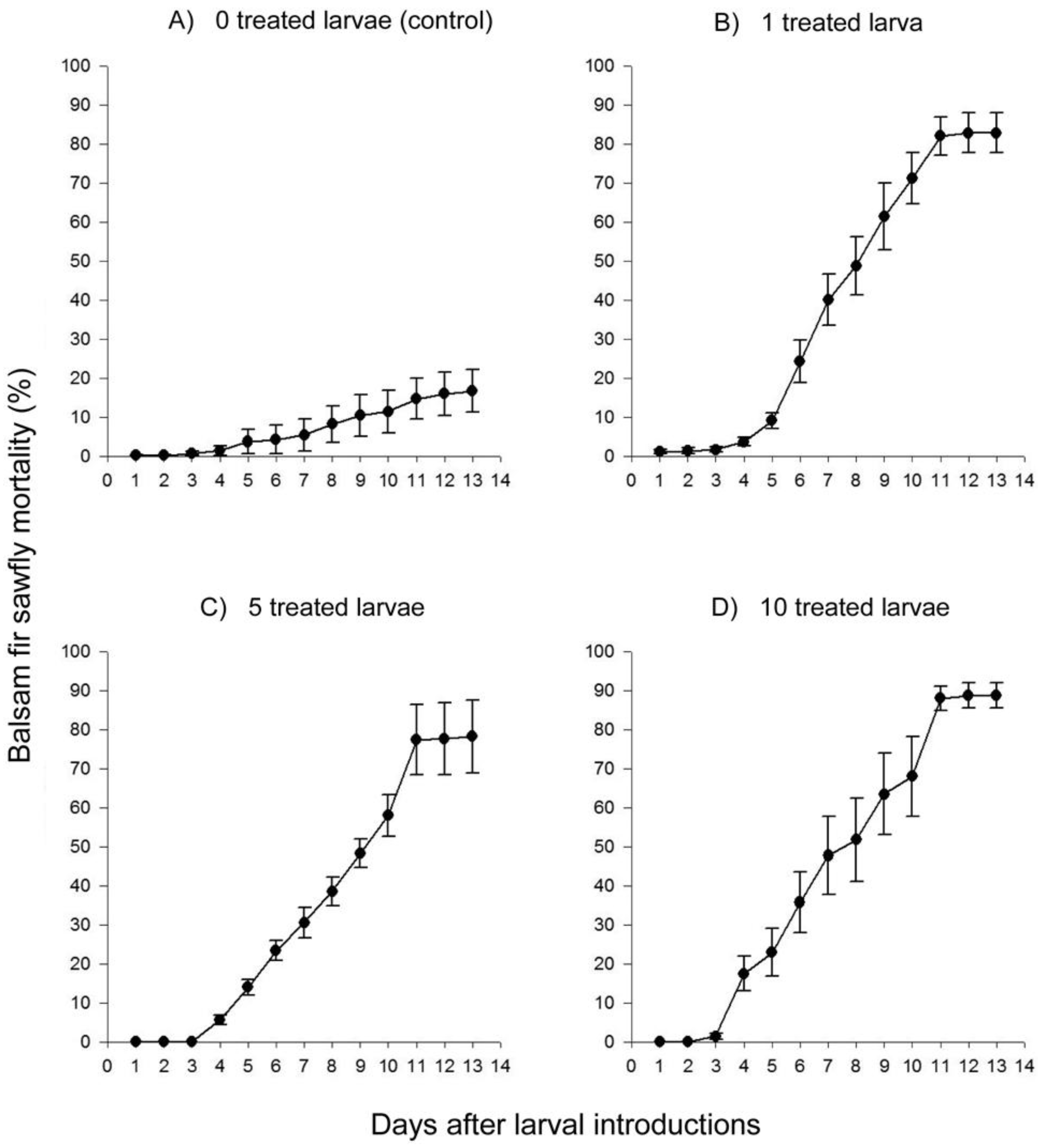
2. Results and Discussion
2.1. Field Study
2.2. Laboratory Study
3. Materials and Methods
3.1. Balsam Fir Sawfly Life History
3.2. NeabNPV
3.3. Field Study
3.4. Laboratory Study
3.5. Molecular Probing for the Presence of NeabNPV
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Smith, D.R. Systematics, life history and distribution of sawflies. In Sawfly Life HistoryAdaptations to Woody Plants; Wagner, M.R., Raffa, K.F., Eds.; Academic Press: New York, NY, USA, 1993; pp. 3–32. [Google Scholar]
- Wallace, D.R.; Cunningham, J.C. Diprionid sawflies. In Forest Insect Pests in Canada; Armstrong, J.A., Ives, W.G.H., Eds.; Natural Resources Canada: Ottawa, Canada, 1995; pp. 193–232. [Google Scholar]
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar] [CrossRef]
- Knerer, G.; Atwood, C. Diprionid sawflies: Polymorphism and speciation. Science 1973, 179, 1090–1099. [Google Scholar]
- Moreau, G. Past and present outbreaks of the balsam fir sawfly: An analytical review. Forest Ecol. Manag. 2006, 221, 215–219. [Google Scholar] [CrossRef]
- Canadian Council of Forest Ministers. Forest Insect National Tables for Areas of Moderate to Severe Defoliation. Available online: http://nfdp.ccfm.org/insects/national_e.php (Accessed on 27 July 2012).
- Smith, H.V.E. Annual Report of the Fire Patrol to the Annual Executive Meeting of The Newfoundland Forest Protection Association; Blackmore Printing Co. Ltd. Grand Falls: Newfoundland, Canada, 1947; pp. 46–63. [Google Scholar]
- Carroll, W.J. Some aspects of the Neodiprion abietis complex in Newfoundland. Ph.D. dissertation, State University College of Forestry, Syracuse University, Syracuse, NY, USA, 1962; p. 186. [Google Scholar]
- Duffy, S.P.; Young, A.M.; Morin, B.; Lucarotti, C.J.; Koop, B.F.; Levin, D.B. Sequence analysis and organization of the Neodiprion abietis nucleopolyhedrovirus genome. J. Virol. 2006, 80, 6952–6963. [Google Scholar] [CrossRef]
- Moreau, G.; Lucarotti, C.J.; Kettela, E.G.; Thurston, G.S.; Holmes, S.; Weaver, C.; Levin, D.B.; Morin, B. Aerial application of nucleopolyhedrovirus induces decline in increasing and peaking populations of Neodiprion abietis. Biol. Contr. 2005, 33, 65–73. [Google Scholar] [CrossRef]
- Lucarotti, C.J.; Moreau, G.; Kettela, E.G. Abietiv™—A viral biopesticide for control of the balsam fir sawfly. In Biological Control: A Global Pespective; Vincent, C., Goettel, M., Lazarovits, G., Eds.; CABI International: Wallingford, UK, 2007; pp. 353–361. [Google Scholar]
- Lucarotti, C.J.; Morin, B.; Graham, R.I.; Lapointe, R. Production, application, and field performance of Abietiv™, the balsam fir sawfly nucleopolyhedrovirus. Virol. Sinica. 2007, 22, 163–172. [Google Scholar] [CrossRef]
- Moreau, G. The Influence of Forest Management on Defoliator Populations: A Case Study with Neodiprion Abietis in Pre-commercially thinned and Natural Forest Stands. Ph.D. dissertation, Faculty of Forestry and Environmental Management, University of New Brunswick, New Brunswick, Canada, 2004; p. 166. [Google Scholar]
- Federici, B.A. Baculovirus pathogenesis. In The Baculoviruses; Miller, K.L., Ed.; Plenum Publishing Corporation: New York, NY, USA, 1997; pp. 33–59. [Google Scholar]
- Lucarotti, C.J.; Whittome-Waygood, B.H.; Lapointe, R.; Levin, D.B. Pathology of a Gammabaculovirus in its natural balsam fir sawfly (Neodiprion abietis) host. Psyche 2012. submitted. [Google Scholar]
- Campbell, C.S.; Quiring, D.T.; Kettela, E.G.; Lucarotti, C.J. Application of balsam fir sawfly nucleopolyhedrovirus against its natural host Neodiprion abietis (Hymenoptera Diprionidae). In Proceedings of the IUFRO Workshop on Forest Insect Population Dynamics and Host Influences, Kanazawa, Japan, 14-19 September 2003; pp. 86–89.
- Dwyer, G. The roles of density, stage, and patchiness in the transmission of an insect virus. Ecology 1991, 72, 559–574. [Google Scholar] [CrossRef]
- Dwyer, G.; Elkinton, J.L.; Buonaccorsi, J.P. Host heterogeneity in susceptibility and disease dynamics: tests of a mathematical model. Amer. Natur. 1997, 150, 685–707. [Google Scholar]
- Moreau, G.; Eveleigh, E.S.; Lucarotti, C.J.; Quiring, D.T. Stage-specific responses to ecosystem alteration in an eruptive herbivorous insect. J. Appl. Ecol. 2006, 43, 28–34. [Google Scholar]
- Anstey, L.J.; Quiring, D.T.; Ostaff, D.P. Seasonal changes in intra-tree distribution of immature balsam fir sawfly (Hymenoptera: Diprionidae). Can. Entomol. 2002, 134, 529–538. [Google Scholar] [CrossRef]
- Cory, J.S.; Hails, R.S.; Sait, S.M. Baculovirus ecology. In The Baculoviruses; Miller, L.K., Ed.; Plenum Publishing Corporation: New York, NY, USA, 1997; pp. 301–339. [Google Scholar]
- Kaupp, W.J. Estimation of nuclear polyhedrosis virus produced in field populations of the European pine sawfly, Neodiprion sertifer (Geoff.) (Hymenoptera: Diprionidae). Can. J. Zool. 1982, 61, 1857–1861. [Google Scholar] [CrossRef]
- Kaupp, W.J. Persistence of Neodiprion sertifer (Hymenoptera: Diprionidae) nuclear polyhedrosis virus on Pinus contortata foliage. Can. Entomol. 1983, 115, 869–873. [Google Scholar] [CrossRef]
- Olofsson, E. Environmental persistence of the nuclear polyhedrovirus of the European spruce sawfly in relation to epizootics in Swedish scots pine forests. J. Invertebr. Pathol. 1987, 52, 119–129. [Google Scholar] [CrossRef]
- Olofsson, E. Persitence and dispersal of the nuclear polyhedrovirus of the European spruce sawfly (Geoffroy) [Hymenoptera:Diprionidae] in a virus-free lodgepole pine plantation in Sweden. Can. Entomol. 1988, 120, 887–892. [Google Scholar] [CrossRef]
- Olofsson, E. Dispersal of the nuclear polyhedrosis virus of Neodiprion sertifer from soil to pine foliage with dust. Entomol. Exper. Appl. 1988, 46, 181–186. [Google Scholar] [CrossRef]
- Olofsson, E. Transmission of the nuclear polyhedrosis virus of the european pine sawfly from adult to offspring. J. Invertebr. Pathol. 1989, 54, 322–330. [Google Scholar] [CrossRef]
- Young, S.Y.; Yearian, W.C. Intra- and inter-colony transmission of a nuclear polyhedrosis virus of loblolly pine sawfly Neodiprion taedae linearis. J. Entomol. Sci. 1987, 22, 29–34. [Google Scholar]
- Young, S.Y.; Yearian, W.C. Secondary transmission of a nuclear polyhedrosis virus of Neodiprion taedae [Hym.:Diprionidae] between larval colonies on loblolly pine. Entomophaga 1987, 34, 341–349. [Google Scholar]
- Young, S.Y.; Yearian, W.C. The influence of canopy location on intercolony transmission of nuclear polyhedrosis virus in larval populations of loblolly pine sawfly Neodiprion taedae linearis. J. Entomol. Sci. 1990, 25, 535–537. [Google Scholar]
- Bird, F.T. Transmission of some insect viruses with particular reference to ovarian transmission and its importance in the development of epizootics. J. Insect Pathol. 1961, 3, 352–380. [Google Scholar]
- Dwyer, G.; Elkinton, J.S. Host dispersal and the spatial spread of insect pathogens. Ecology 1995, 76, 1262–1275. [Google Scholar] [CrossRef]
- D’Amico, V.; Elkinton, J.S.; Dwyer, G.; Burand, J.P.; Buonaccorsi, J.P. Virus transmission in gypsy moths is not a simple mass action process. Ecology 1996, 77, 201–206. [Google Scholar] [CrossRef]
- Vasconcelos, S.D.; Cory, J.S.; Wilson, K.R.; Sait, S.M.; Hails, R.S. Modified behaviour in baculovirus-infected lepidopteran larvae and its impact on the spatial distribution of inoculum. Biol. Control 1996, 7, 299–306. [Google Scholar] [CrossRef]
- Knell, R.J.; Begon, M.; Thompson, D.J. Transmission of Plodia interpunctella granulosis virus does not conform to the mass action model. J. Animal Ecol. 1998, 67, 592–599. [Google Scholar]
- Beisner, E.B.; Meiers, J.H. Population density and transmission of virus in experimental populations of the western tent caterpillar (Lepidoptera: Lasiocamidae). Environ. Entomol. 1999, 28, 1107–1113. [Google Scholar]
- Dwyer, G.; Dushoff, J.; Elkinton, J.; Levin, S. Pathogen-driven outbreaks in forest defoliators revisited: Building models from experimental data. Amer. Natur. 2000, 156, 105–120. [Google Scholar] [CrossRef]
- Hails, R.S.; Hernandez-Crespo, P.; Sait, S.M.; Donnelly, C.A.; Green, B.M.; Cory, J.S. Transmission patterns of natural and recombinant baculoviruses. Ecology 2002, 83, 906–916. [Google Scholar]
- D’Amico, V.; Elkinton, J.S.; Podgwaite, J.D.; Buonaccorsi, J.P.; Dwyer, G. Pathogen clumping: An explanation for non-linear transmission of an insect virus. Ecol. Entomol. 2005, 30, 383–390. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Population biology of infectious diseases: Part I. Nature 1979, 280, 361–367. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Infectious diseases and population cycles of forest insects. Science 1980, 210, 658–661. [Google Scholar]
- Anderson, R.M.; May, R.M. The population dynamics of microparasites and their invertebrate hosts. Phil. Trans. Roy. Soc. London B Biol. Sci. 1981, 291, 451–524. [Google Scholar] [CrossRef]
- Parsons, K.H.; Piene, H.; Farrell, J.; Quiring, D.T. Temporal patterns of balsam fir sawfly defoliation and growth loss in young balsam fir. Forest Ecol. Manag. 2003, 184, 33–46. [Google Scholar] [CrossRef]
- Parsons, K.; Quiring, D.; Piene, H.; Moreau, G. Relationship between balsam fir sawfly density and defoliation in balsam fir. Forest Ecol. Manag. 2005, 205, 325–331. [Google Scholar] [CrossRef]
- Health Canada. Registration Decision Neodiprion Abietis Newfoundland Strain; Health Canada Pest Management Regulatory Agency: Ottawa, Canada, 2009; pp. 1–9.
- Duffy, S.P.; Becker, E.M.; Whittome, B.; Lucarotti, C.J.; Levin, D.B. In vivo replication kinetics and transcription patterns of the balsam fir sawfly, Neodiprion abietis, nucleopolyhedrovirus. J. Gen. Virol. 2007, 88, 1945–1951. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Graves, R.; Quiring, D.T.; Lucarotti, C.J. Transmission of a Gammabaculovirus within Cohorts of Balsam Fir Sawfly (Neodiprion abietis) Larvae. Insects 2012, 3, 989-1000. https://doi.org/10.3390/insects3040989
Graves R, Quiring DT, Lucarotti CJ. Transmission of a Gammabaculovirus within Cohorts of Balsam Fir Sawfly (Neodiprion abietis) Larvae. Insects. 2012; 3(4):989-1000. https://doi.org/10.3390/insects3040989
Chicago/Turabian StyleGraves, Roger, Dan T. Quiring, and Christopher J. Lucarotti. 2012. "Transmission of a Gammabaculovirus within Cohorts of Balsam Fir Sawfly (Neodiprion abietis) Larvae" Insects 3, no. 4: 989-1000. https://doi.org/10.3390/insects3040989
APA StyleGraves, R., Quiring, D. T., & Lucarotti, C. J. (2012). Transmission of a Gammabaculovirus within Cohorts of Balsam Fir Sawfly (Neodiprion abietis) Larvae. Insects, 3(4), 989-1000. https://doi.org/10.3390/insects3040989