The Feeding Rate of Predatory Mites on Life Stages of Bemisia tabaci Mediterranean Species

{kind=link}
{kind=link}
Abstract
:1. Introduction
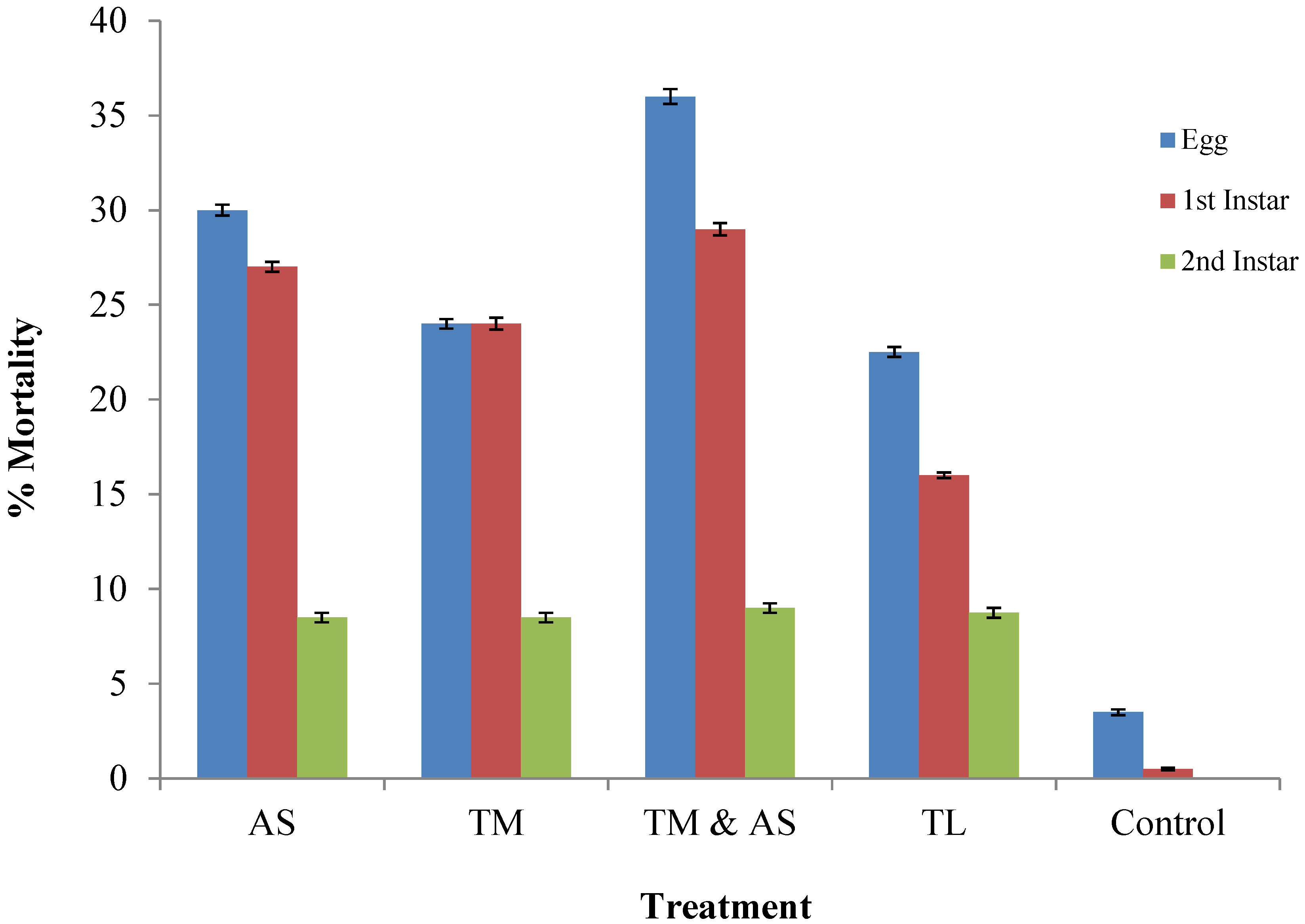
2. Experimental
2.1. Source of Insects
2.2. Feeding Trails
2.3. Data Analysis
3. Results and Discussion


4. Conclusions
Acknowledgments
Conflicts of Interest
References
- De Barro, P.J.; Hart, P.J. Mating interactions between two biotypes of the whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae) in Australia. Bull. Entomol. Res. 2000, 90, 103–112. [Google Scholar]
- Cock, M.J.N. Bemisia Tabaci, a Literature Survey: On the Cotton Whitefly with an Annotated Bibliography; FAO/CAB International Institute of Biologica-l Control: Ascot, UK, 1986; p. 21. [Google Scholar]
- Oliveira, M.R.V.; Henneberry, T.J.; Anderson, P. History, current status and collaborative research projects for Bemisia tabaci. Crop Prot. 2001, 20, 709–723. [Google Scholar] [CrossRef]
- Gelman, D.B.; Blackburn, M.B.; Hu, J.S. Identification of the molting hormone of the sweet potato (Bemisia tabaci) and greenhouse (Trialeurodes vaporariorum) whitefly. J. Insect Physiol. 2005, 51, 47–53. [Google Scholar] [CrossRef]
- Dalton, R. Whitefly infestations: The Christmas invasion. Nature 2006, 443, 898–900. [Google Scholar] [CrossRef]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Alegbejo, M.D. Whitefly transmitted plant viruses in Nigeria. J. Sustain. Agric. 2000, 17, 99–109. [Google Scholar]
- Simón, B.; Cenis, J.L.; Demichelis, S.; Rapisarda, C.; Cacigli, P.; Bosco, D. Survey of Bemisa tabaci (Hemiptera: Aleyrodidae) biotypes in Italy with the description of a new biotype (T) from Euphorbia characias. Bull. Entomol. Res. 2003, 93, 259–264. [Google Scholar]
- Dinsdale, A.; Cook, L.; Riginos, C.; Buckley, Y.M.; de Barro, P. Refined global analysis of bemisia tabaci (Hemiptera: Sternorrhyncha: Aleyrodoidea: Aleyrodidae) mitochondrial cytochrome oxidase 1 to identify species level genetic boundaries. Annu. Rev. Entomol. Soc. Am. 2010, 103, 196–208. [Google Scholar]
- Bethke, J.A.; Byrne, F.J.; Hodges, G.S.; McKenzie, C.L.; Shatters, R.G. First record of the Q biotype of the sweetpotato whitefly, Bemisia tabaci, in Guatemala. Phytoparasitica 2009, 37, 61–64. [Google Scholar] [CrossRef]
- Jones, C.M.; Gorman, K.; Denholm, I.; Williamson, M.S. High-throughput allelic discrimination of B and Q biotypes of the whitefly, Bemisia tabaci, using TaqMan allele-selective PCR. Pest Manag. Sci. 2008, 64, 12–15. [Google Scholar] [CrossRef]
- McKenzie, C.I.; Hodges, G.; Osborne, L.; Byrne, F.J.; Shatters, R.G. Distribution of Bemisia tabaci (Hemiptera: Aleyrodidae) Biotypes in Florida-Investigating the Q Invasion. Hortic. Entomol. 2009, 102, 670–676. [Google Scholar]
- Cuthbertson, A.G.S. Update on the status of Bemisia tabaci in the UK and the use of entomopathogenic fungi within eradication programmes. Insects 2013, 4, 198–205. [Google Scholar] [CrossRef]
- Cheek, S.; Macdonald, O. Statutory controls to prevent the establishment of Bemisia tabaci in the United Kingdom. Pestic. Sci. 1994, 42, 135–142. [Google Scholar] [CrossRef]
- Powell, M.E.; Cuthbertson, A.G.S.; Bell, H.A.; Boonham, N.; Morris, J.; Northing, P. First record of the Q Biotype of the sweetpotato whitefly, Bemisia tabaci, intercepted in the UK. Eur. J. Plant Pathol. 2012, 133, 797–801. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Head, J.; Walters, K.F.A.; Gregory, S.A. The efficacy of the entomopathogenic nematode, Steinernema feltiae, against the immature stages of Bemisia tabaci. J. Invertebr. Pathol. 2003, 83, 267–269. [Google Scholar] [CrossRef]
- Cuthbertson, A.G.S.; Mathers, J.J.; Croft, P.; Nattriss, N.; Blackburn, L.F.; Luo, W.; Northing, P.; Murai, T.; Jacobson, R.J.; Walters, K.F.A. Prey consumption rates and compatibility with pesticides of four predatory mites from the family Phytoseiidae attacking Thrips palmi Karny (Thysanoptera: Thripidae). Pest Manag. Sci. 2012, 68, 1289–1295. [Google Scholar] [CrossRef]
- Tashiro, H. Self-watering acrylic cages for confining insects and mites on detached leaves. J. Econ. Entomol. 1967, 60, 354–356. [Google Scholar]
- Cuthbertson, A.G.S.; Mathers, J.J.; Blackburn, L.F.; Korycinska, A.; Powell, M.E.; Luo, W.; Jacobson, R.J.; Northing, P. Population development of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) under simulated UK glasshouse conditions. Insects 2012, 4, 185–197. [Google Scholar]
- Messelink, G.J.; van Steenpaal, S.E.F.; Ramakers, P.M.J. Evaluation of phytoseiid predators for control of western flower thrips on greenhouse cucumber. Biocontrol 2006, 51, 753–768. [Google Scholar] [CrossRef]
- Nomikou, M.; Sabelis, M.W.; Janssen, A. Pollen subsidies promote whitefly control through the numerical response of predatory mites. Biocontrol 2010, 55, 253–260. [Google Scholar] [CrossRef]
- Calvo, F.J.; Bolckmans, K.; Belda, J.E. Control of Bemisia tabaci and Frankliniella occidentalis in cucumber by Amblyseius swirskii. Biocontrol 2011, 56, 185–192. [Google Scholar] [CrossRef]
- Xiao, Y.-F.; Avery, P.; Chen, J.-J.; McKenzie, C.; Osborne, L. Ornamental pepper as banker plants for establishment of Amblyseius swirskii (Acari: Phytoseiidae) for biological control of multiple pests in greenhouse vegetable production. Biol. Control 2012, 63, 279–286. [Google Scholar] [CrossRef]
- Onzo, A.; Zannou, I.D.; Adjibade, O.A.J.D.; Broutani, S.; Hanna, R. Potentialités de l’acarien prédateur Amblyseius swirskii (Athias-Henriot) (Acari: Phytoséiidae) dans la lutte biologique contre la mouche blanche Bemisia tabaci (Genn.), vecteur de la mosaïque du manioc en Afrique. Int. J. Biol. Chem. Sci. 2012, 6, 5085–5102. [Google Scholar]
- Calvo, F.J.; Bolckmans, K.; Belda, J.E. Biological control-based IPM in sweet pepper glasshouses using Amblyseius swirskii (Acri: Phytoseiidae). Biocontrol Sci. Technol. 2012, 22, 1398–1416. [Google Scholar] [CrossRef]
- Nomikou, M.; Janssen, A.; Schraag, R.; Sabelis, M.W. Vulnerability of Bemisia tabaci immatures to phytoseiid predators: Consequences for oviposition and influence of alternative food. Entomol. Exp. Appl. 2004, 110, 95–102. [Google Scholar] [CrossRef]
- Nomikou, M.; Janssen, A.; Schraag, R.; Sabelis, M.W. Phytoseiid predators suppress populations of Bemisia tabaci on cucumber plants with alternative food. Exp. App. Acarol. 2002, 27, 57–68. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cuthbertson, A.G.S. The Feeding Rate of Predatory Mites on Life Stages of Bemisia tabaci Mediterranean Species. Insects 2014, 5, 609-614. https://doi.org/10.3390/insects5030609
Cuthbertson AGS. The Feeding Rate of Predatory Mites on Life Stages of Bemisia tabaci Mediterranean Species. Insects. 2014; 5(3):609-614. https://doi.org/10.3390/insects5030609
Chicago/Turabian StyleCuthbertson, Andrew G. S. 2014. "The Feeding Rate of Predatory Mites on Life Stages of Bemisia tabaci Mediterranean Species" Insects 5, no. 3: 609-614. https://doi.org/10.3390/insects5030609