A New Alien Invasive Longhorn Beetle, Xylotrechus chinensis (Cerambycidae), Is Infesting Mulberries in Catalonia (Spain)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects in the Wild
2.2. Insects in Semi-Field Conditions
2.3. Climatic Data and First Adult Emergences
2.4. Studies on Potential Alternative Hostplants (Common Grape Vine)
3. Results and Discussion
3.1. Arrival and Current Area Invaded in Catalonia
3.2. Description of Life Stages
3.3. Biology and Ecology
3.3.1. Mating Behaviour and Egg Production Capacity
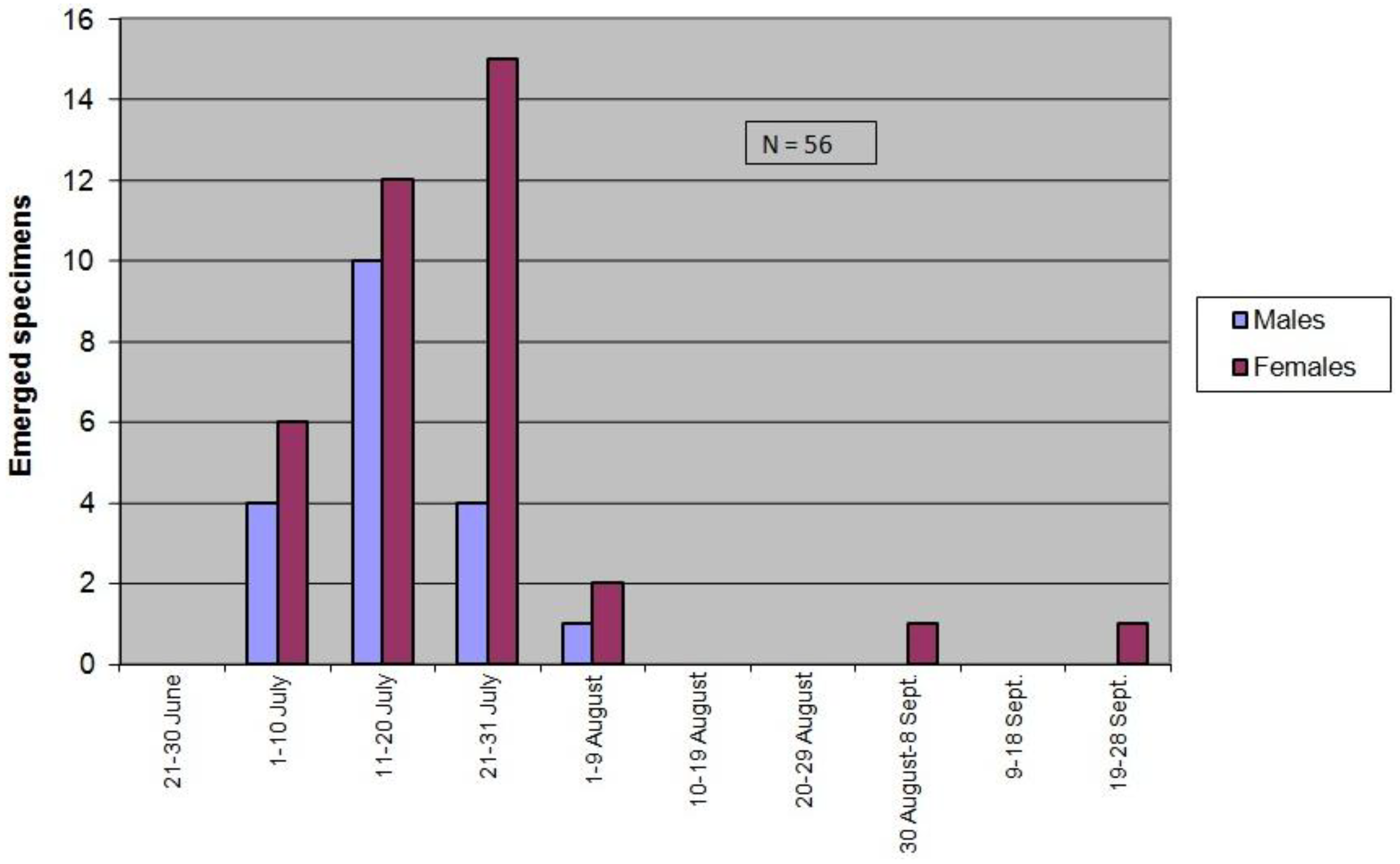
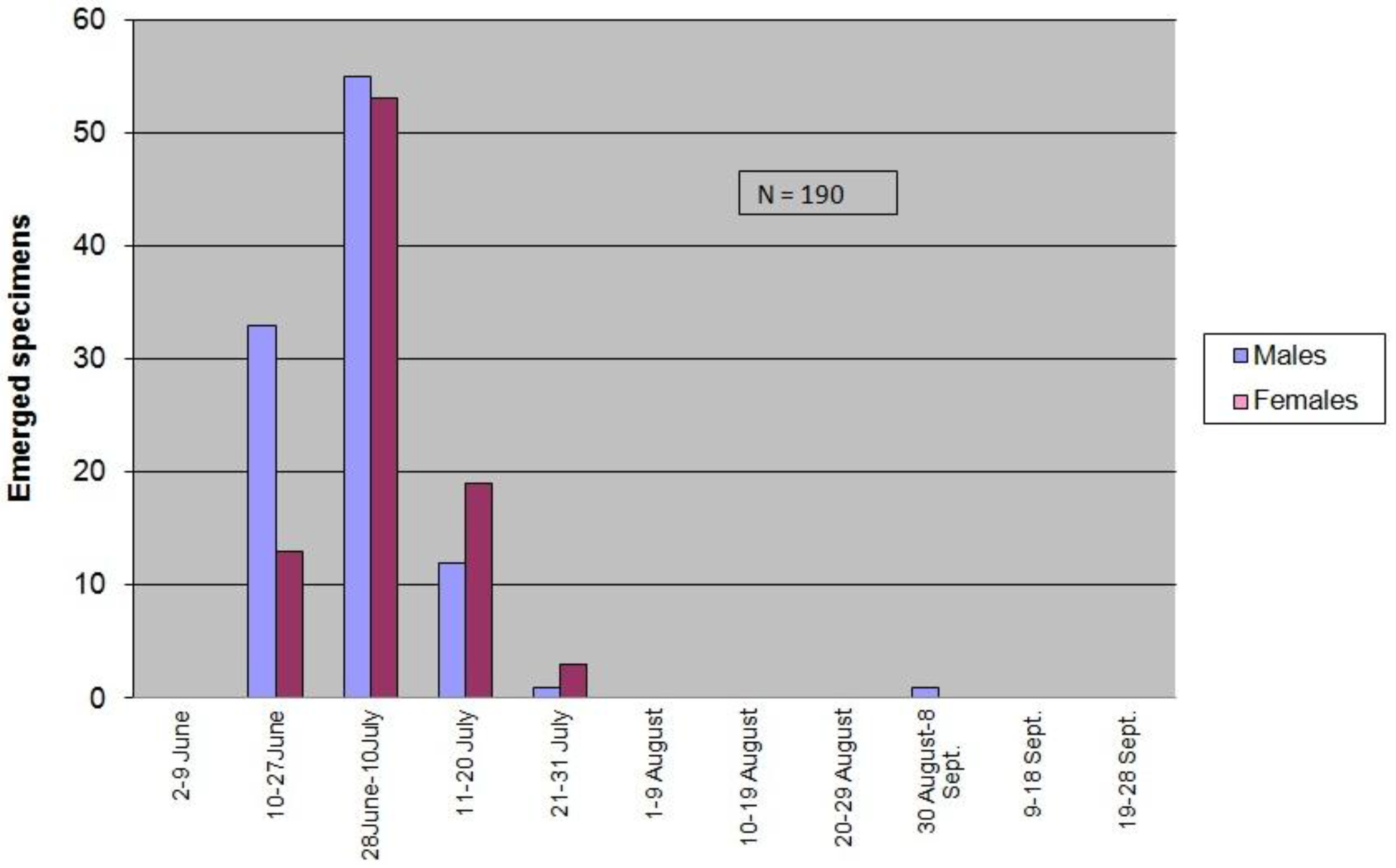
3.3.2. Seasonality
3.4. Larval Development and Damage to Mulberries in Catalonia
3.5. Host Plants Other Than Mulberries
3.6. Pathway and Vector Capacity of Pathogens
3.7. Possible Control Measures and Suggestions for Future Studies
- (a)
- Physical control. Removal and destruction (burning or grinding) of heavily infested mulberry trees. To date this has been the only measure used in the invaded towns.
- (b)
- Chemical control. To date no insecticides have been tested for their efficacy against X. chinensis in Spain. However, to our understanding, two approaches are feasible:
- (1)
- Targeting ovipositing females, their eggs and first instar larvae. Since females lay eggs on mulberry barks (trunks and main branches) from mid-June to mid-August, spraying contact insecticide on such barks at the beginning of June should suffice to protect the trees. This assumes a two-month half-life under outdoor conditions of the insecticide used; if shorter, a second treatment might be carried out in mid-July. One such insecticide might be chlorpyrifos, sold under many brand names and registered and authorized in Spain for the protection of agricultural crops. It is an organophosphate acting as a contact insecticide and stomach poison, considered moderately hazardous to humans by the World Health Organization. It worked quite well against the Castniid Palm Borer, Paysandisia archon (Burmeister), an exotic invasive moth in Europe that attacks palm trees [29]. However, special regulations applying to insecticide use in public areas may hinder its implementation.
- (2)
- Targeting larvae (all instars) feeding in the tree phloem. This would require the use of a systemic insecticide, for instance abamectin, mostly a stomach poison, introduced into the xylem through endotherapy, i.e., the trunk injection of a sap-compatible solution of the insecticide. The xylem sap moves upwards through the vessels and the active ingredient is so transported to the phloem. This confines the applied insecticide only within the target tree, thereby making it particularly useful in urban situations. Abamectin has shown good results against bark beetles (see for instance [30]) and will be used experimentally in the town of Barberà del Vallès in the spring of 2018.
Injecting insecticide into the emergence holes seen on mulberry barks, apart from being very time-consuming, would not be efficient since they are not really well connected to the phloem cavities inhabited by larvae. Obviously, an emergence hole leads to an empty tunnel where the larva pupated and the hole is only made when the adult abandons the tree. - (c)
- (d)
- Biological control. During the course of this work we discovered the native stephanid wasp Stephanus serrator (Fabricius 1798) (Figure 10) as a likely parasitoid of X. chinensis larvae. Since it was not possible to find the corpse of the parasitized beetle larva within the several bolts infested by X. chinensis, we only can assume the latter served as the host for S. serrator. This assumption is reinforced by the fact that local woodborers do not generally appear as pests of mulberries in Spain [31] and especially because no bark beetles or woodboring beetles other than X. chinensis emerged from our bolts.
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Allison, J.D.; Mckenney, J.I.; Millar, J.G.; Mcelfresh, J.S.; Mitchell, R.F.; Hanks, L.M. Response of the woodborers Monochamus carolinensis and Monochamus titillator (Coleoptera: Cerambycidae) to known cerambycid pheromones in the presence and absence of the host plant volatile alfa-pinene. Environ. Entomol. 2012, 41, 1587–1596. [Google Scholar] [CrossRef] [PubMed]
- Linsley, E.G. Ecology of Cerambycidae. Annu. Rev. Entomol. 1959, 4, 99–138. [Google Scholar] [CrossRef]
- Ginzel, M.D.; Hanks, L.M. Contact pheromones as mate recognition cues of four species of longhorned beetles (Coleoptera: Cerambycidae). J. Insect Behav. 2003, 16, 181–187. [Google Scholar] [CrossRef]
- Hanks, L.M.; Millar, J.G. Sex and aggregation-sex pheromones of cerambycid beetles: Basic science and practical applications. J. Chem. Ecol. 2016, 42, 631–654. [Google Scholar] [CrossRef] [PubMed]
- Cocquempot, C.; Lindelöw, A. Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1. In Alien Terrestrial Arthropods of Europe. BioRisk 2010, 4, 193–218. [Google Scholar] [CrossRef]
- Cherepanov, A.I. Cerambycidae of Northern Asia. Volume 2. Cerambycinae. Part II; Otdelenie, S., Ed.; Nauka Publishers, Siberian Division: Novosibirsk, Russia, 1982; p. 354, (Translated from Russian in 1988. Amerind Publishing Co. Pvt. Ltd. New Delhi). [Google Scholar]
- Ohbayashi, N.; Niisato, T. Longicorn Beetles of Japan; Tokai University Press: Kanagawa, Japan, 2007; p. 818. [Google Scholar]
- Monné, M.A.; Hovore, F.T. Checklist of the Cerambycidae, or Longhorned Wood-Boring Beetles, of the Western Hemisphere; BioQuip Products: Rancho Dominguez, CA, USA, 2006; p. 394. [Google Scholar]
- Swift, I. A new species of Xylothrechus (Col: Cer: Cer: Clytini) from Southern California, U.S.A. Coleopt. Bull. 2007, 61, 287–291. [Google Scholar] [CrossRef]
- Alten, R.L. A new species of Xylotrechus (Coleoptera: Cerambycidae: Clytini) from Utah. Insecta Mundi 2014, 0355, 1–6. [Google Scholar]
- Sama, G.; Löbl, I. Cerambycinae. In Catalogue of Palaearctic Coleoptera. Vol. 6, Chrysomeloidea; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2010; Volume 6, pp. 143–207. [Google Scholar]
- Fauna Europaea. Available online: https://fauna-eu.org/ (accessed on 12 September 2017).
- Vives, E. Coleoptera, Cerambycidae. In Fauna Ibérica; Ramos, M.A., Ed.; Museo Nacional de Ciencias Naturales. CSIC: Madrid, Spain, 2000; Volume 12, pp. 1–716. ISBN 84-00-07887-X. [Google Scholar]
- Recalde, J.I.; San Martín, A.F. Descubrimiento de Xylotrechus stebbingi Gahan, 1906 (Coleoptera: Cerambycidae) en Navarra (norte de España), y otras especies saproxílicas del desfiladero de Gallipienzo. Arquivos Entomolóxicos 2015, 13, 347–350. [Google Scholar]
- Ocete, R.; López Martínez, M.A.; Prendes, C.; Lorenzo, C.D.; González Andújar, J.L.; Lara, M. Xylotrechus arvicola (Olivier) (Coleoptera, Cerambycidae), a new impacting pest on Spanish vineyards. Vitis 2002, 41, 211–212. [Google Scholar]
- Ocete, R.; Valle, J.M.; Artano, K.; Ocete, M.E.; López, M.A.; Pérez, M.A.; García, D.; Soria, F.J. Evolution of the spatio-temporal distribution of Xylotrechus arvicola (Olivier) (Coleoptera, Cerambycidae) in La Rioja vineyard (Spain). Vitis 2010, 9, 67–70. [Google Scholar]
- Orlinski, A.D. Exotic Forest Pest Information System for North America: Xylotrechus altaicus; North American Forest Commission: Newtown Square, PA, USA, 2000. [Google Scholar]
- Jackson, L.; Price, T.; Smith, G. Exotic Wood Borer/Bark Beetle. National Survey Guidelines; Animal and Plant Health Inspection Service: United States Department of Agriculture, USA. 2010; p. 246. Available online: www.aphis.usda.gov/about_aphis/programs_offices/lpa/index.shtml (accessed on 15 November 2017).
- Benker, U. Stowaways in Wood Packaging Material. Current Situation in Bavaria. Forstschutz Aktuell 2008, 44, 30–31. [Google Scholar]
- Han, Y.; Lyu, D. Taxonomic review of the genus Xylotrechus (Coleoptera: Cerambycidae: Cerambycinae) in Korea with a newly recorded species. Korean J. Appl. Entomol. 2010, 49, 69–82. [Google Scholar] [CrossRef]
- Secretaría General Técnica MAPAMA. Encuesta Sobre Superficies y Rendimientos de Cultivos de España 2016 (ESYRCE). Resultados Nacionales y Autonómicos; Ministerio de Agricultura y Pesca, Alimentación y Medio Ambiente: Madrid, Spain, 2016; p. 46.
- Iwabuchi, K.; Takahashi, J.; Sakai, T. Occurrence of 2, 3-octanediol and 2-hydroxy-3-octanone, possible male sex pheromone in Xylotrechus chinensis Chevrolat (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1987, 22, 110–111. [Google Scholar] [CrossRef]
- Iwabuchi, K. Mating behavior of Xylotrechus pyrrhoderus Bates (Coleoptera: Cerambycidae): 1. Behavioral sequences and existence of the male sex pheromone. Appl. Entomol. Zool. 1982, 17, 494–500. [Google Scholar] [CrossRef]
- Sarto i Monteys, V.; Quero, C.; Santa-Cruz, M.C.; Rosell, G.; Guerrero, A. Sexual communication in day-flying Lepidoptera with special reference to castniids or ‘butterfly-moths’. Bull. Entomol. Res. 2016, 106, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, Y.; Matsuyama, S.; Suzuki, T. Identification of 2,3-octanediol, 2-hydroxy-3-octanone, and 3-hydroxy-2-octanone from male Xylotrechus chinensis Chevrolat as possible sex pheromones (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 1987, 22, 25–28. [Google Scholar] [CrossRef]
- Flaherty, L.; Sweeney, J.D.; Pureswaran, D.; Quiring, D.T. Influence of host tree condition on the performance of Tetropium fuscum (Coleoptera: Cerambycidae). Environ. Entomol. 2011, 40, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Bolòs, O.; Vigo, J.; Masalles, R.M.; Ninot, J.M. Flora Manual dels Països Catalans, 3rd ed.; Editorial Pòrtic: Barcelona, Spain, 2005. [Google Scholar]
- Aruga, H. Principles of Sericulture; CRC Press: Boca Raton, FL, USA, 1994. (In Japanese) [Google Scholar]
- Sarto i Monteys, V.; Aguilar, L. The Castniid Palm Borer, Paysandisia archon (Burmeister, 1880) in Europe: Comparative biology, pest status and possible control methods (Lepidoptera: Castniidae). Nach. Entomol. Ver. Apollo NF 2005, 26, 61–94. [Google Scholar]
- Fettig, C.J.; Grosman, D.M.; Munson, A.S. Efficacy of abamectin and tebuconazole injections to protect Lodgepole Pine from mortality attributed to Mountain Pine Beetle (Coleoptera: Curculionidae) attack and progression of Blue Stain Fungi. J. Entomol. Sci. 2013, 48, 270–278. [Google Scholar] [CrossRef]
- Pérez Cañavate, J.G.; González Benavente, A.; Fernández Hernández, J.A.; Bañón Arias, S. La morera como árbol ornamental. Horticultura. 1992, 76, 13–23. [Google Scholar]
- Van Achterberg, C.; Yang, Z. New species of the genera Megischus Brullé and Stephanus Jurine from China (Hymenoptera: Stephanoidea: Stephanidae), with a key to world species of the genus Stephanus. Zool. Med. Leiden 2004, 78, 101–117. [Google Scholar]
- Selfa, J.; Ventura, D.; Carles-Tolrá, M.; Ortiz Sánchez, F.J.; Pérez-Fernández, T.; Pujade-Villar, J. Nuevos datos sobre la distribución de Stephanus serrator (Fabricius, 1798) en España (Hymenoptera, Stephanidae). Boln. Asoc. Esp. Entomol. 2014, 38, 191–195. [Google Scholar]
- Aguiar, A.P. World catalogue of the Stephanidae (Hymenoptera: Stephanoidea). Zootaxa 2004, 753, 1–120. [Google Scholar] [CrossRef]
- Van Achterberg, C. A revision of the Old World species of Megischus Brullé, Stephanus Jurine and Pseudomegischus gen. nov., with a key to the genera of the family Stephanidae (Hymenoptera: Stephanoidea). Zool. Verh. Leiden 2002, 339, 3–206. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virgin Females | #1 | #2 | #3 | #4 | #5 | #6 | #7 | #8 | #9 | #10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Eggs laid | 0 | 19 | 4 | 6 | 0 | 0 | 5 | 13 | 7 | 0 |
| Eggs remaining in the abdomen | 77 | 69 | 98 | 67 | 77 | 84 | 89 | 69 | 74 | 76 |
| Total eggs | 77 | 88 | 102 | 73 | 77 | 84 | 94 | 82 | 81 | 76 |
| Dead Mated Females | #1 | #2 | #3 | #4 | #5 | #6 | #7 | #8 | #9 | #10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Eggs remaining in the abdomen | 17 | 19 | 0 | 28 | 0 | 23 | 21 | 10 | 6 | 28 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarto i Monteys, V.; Torras i Tutusaus, G. A New Alien Invasive Longhorn Beetle, Xylotrechus chinensis (Cerambycidae), Is Infesting Mulberries in Catalonia (Spain). Insects 2018, 9, 52. https://doi.org/10.3390/insects9020052
Sarto i Monteys V, Torras i Tutusaus G. A New Alien Invasive Longhorn Beetle, Xylotrechus chinensis (Cerambycidae), Is Infesting Mulberries in Catalonia (Spain). Insects. 2018; 9(2):52. https://doi.org/10.3390/insects9020052
Chicago/Turabian StyleSarto i Monteys, Víctor, and Glòria Torras i Tutusaus. 2018. "A New Alien Invasive Longhorn Beetle, Xylotrechus chinensis (Cerambycidae), Is Infesting Mulberries in Catalonia (Spain)" Insects 9, no. 2: 52. https://doi.org/10.3390/insects9020052
APA StyleSarto i Monteys, V., & Torras i Tutusaus, G. (2018). A New Alien Invasive Longhorn Beetle, Xylotrechus chinensis (Cerambycidae), Is Infesting Mulberries in Catalonia (Spain). Insects, 9(2), 52. https://doi.org/10.3390/insects9020052