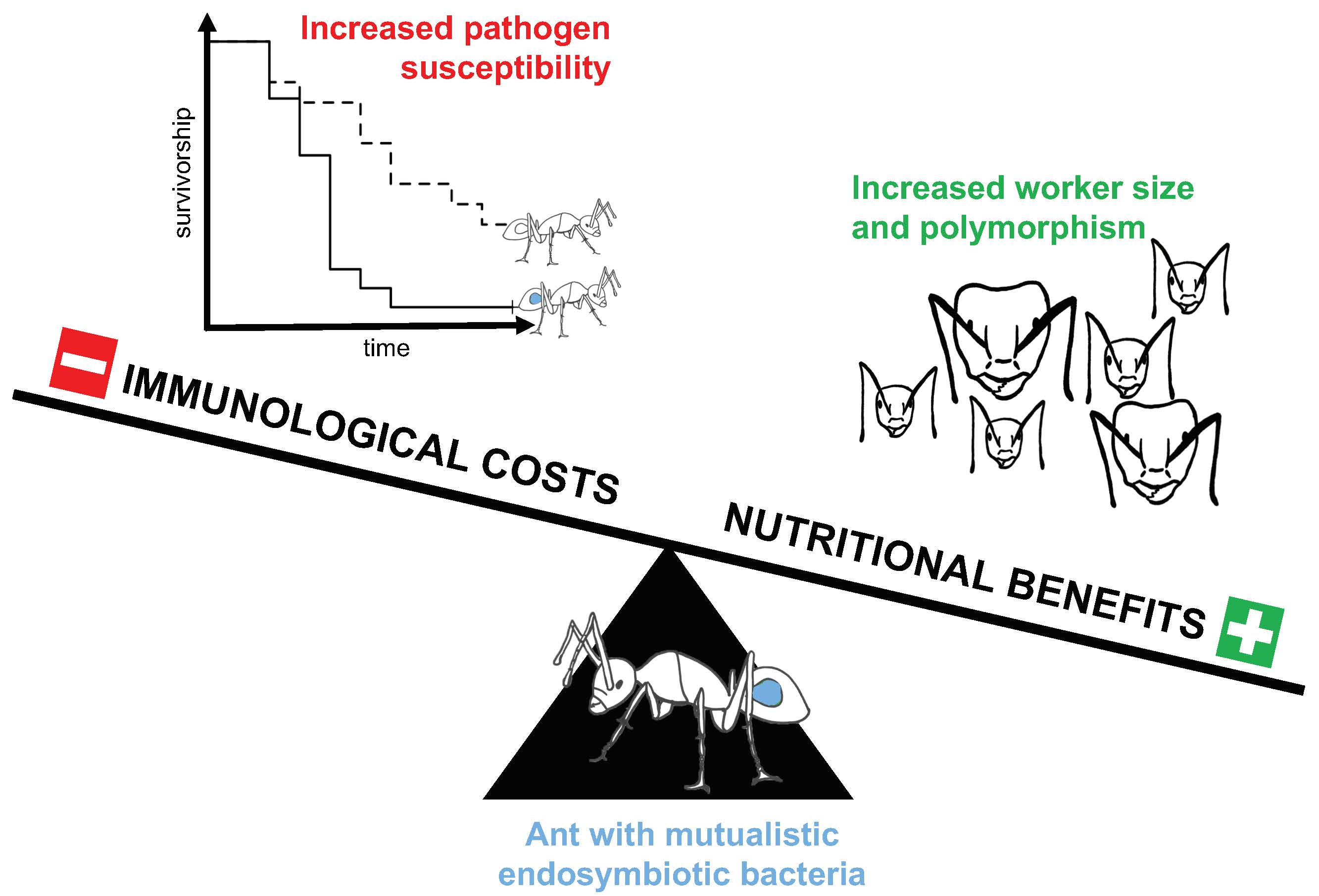
Camponotus floridanus Ants Incur a Trade-Off between Phenotypic Development and Pathogen Susceptibility from Their Mutualistic Endosymbiont Blochmannia


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
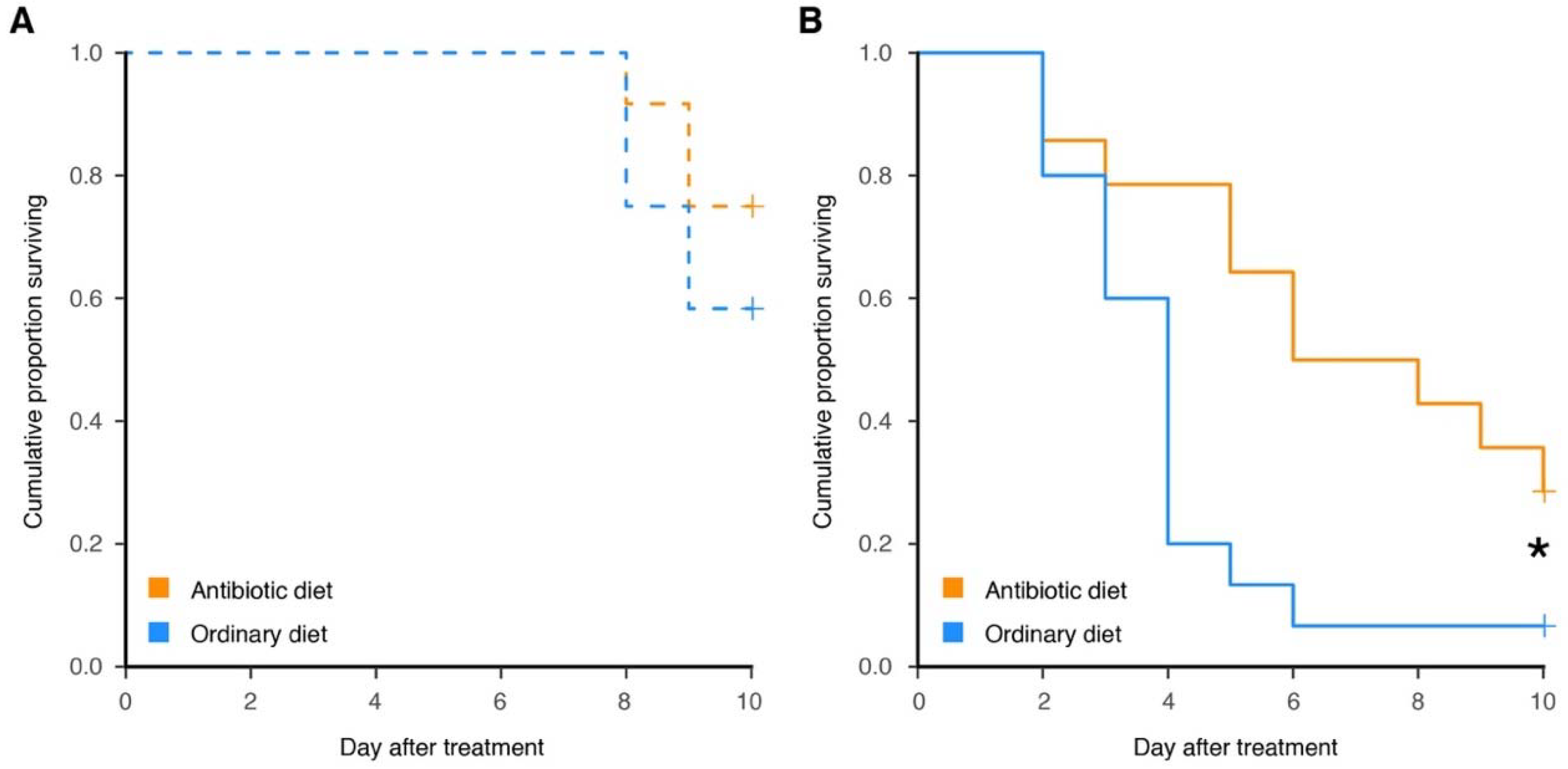{kind=link}
1. Introduction
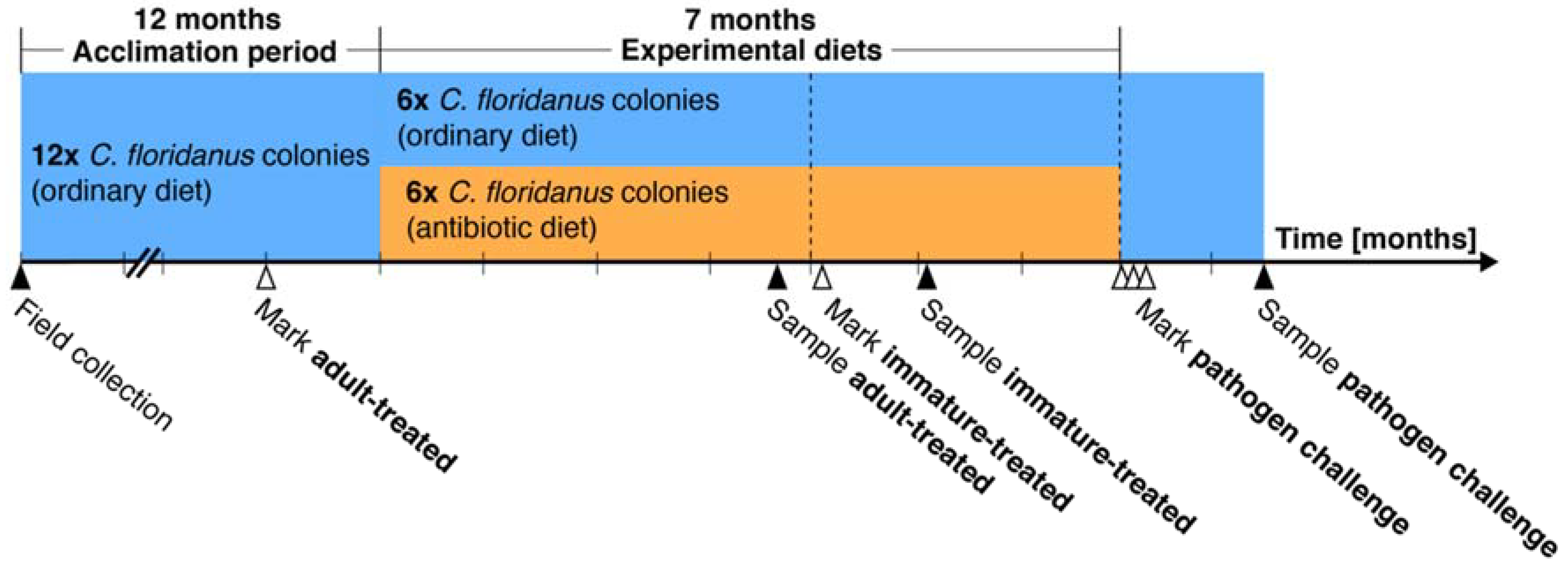
2. Materials and Methods
2.1. Ant Culture and Symbiont Reduction
2.2. Aging of Ants
2.3. Head Width, Melanisation, and Worker Polymorphism Assays
2.4. Fungal Entomopathogen
2.5. Statistical Analysis
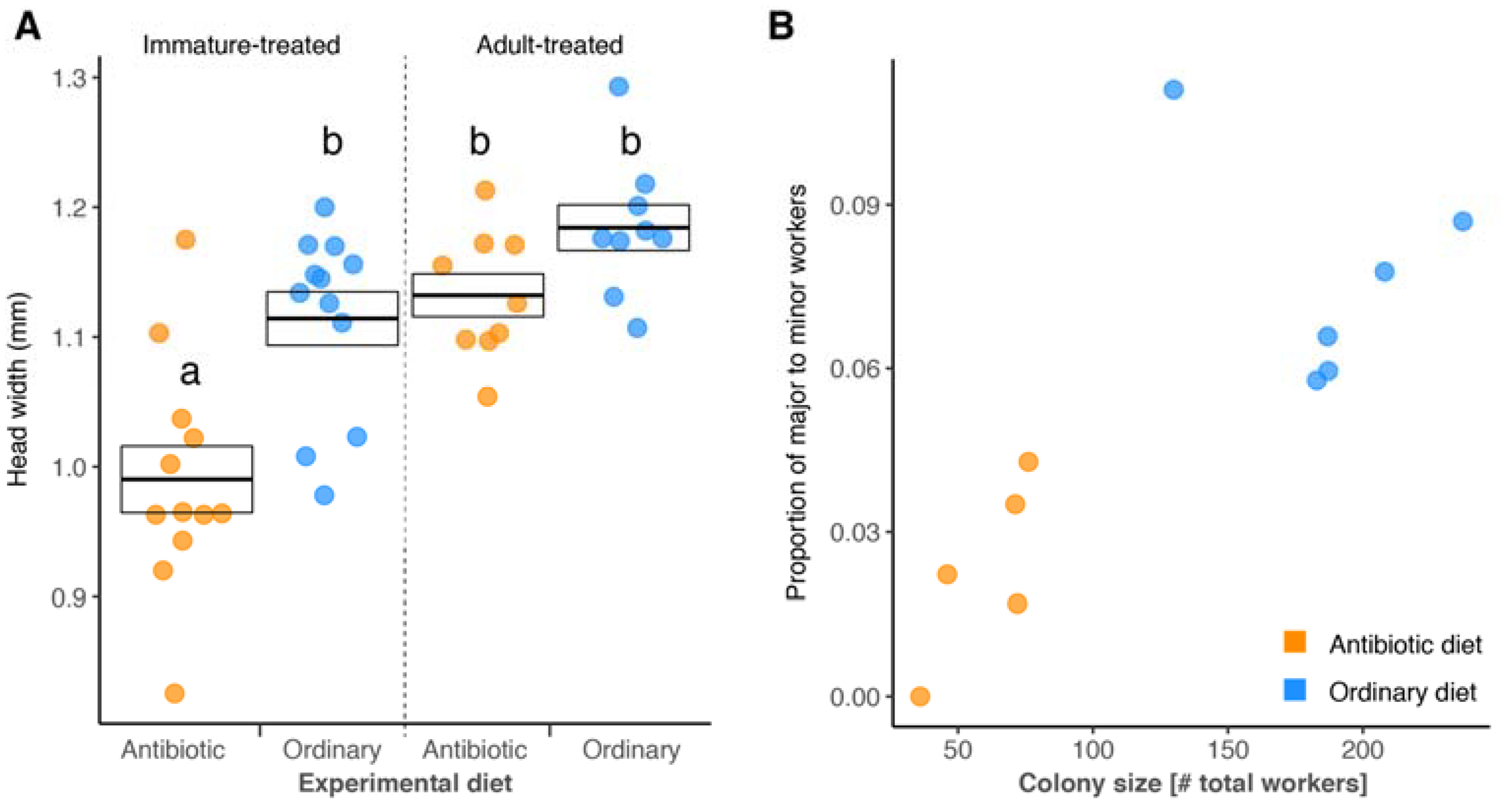
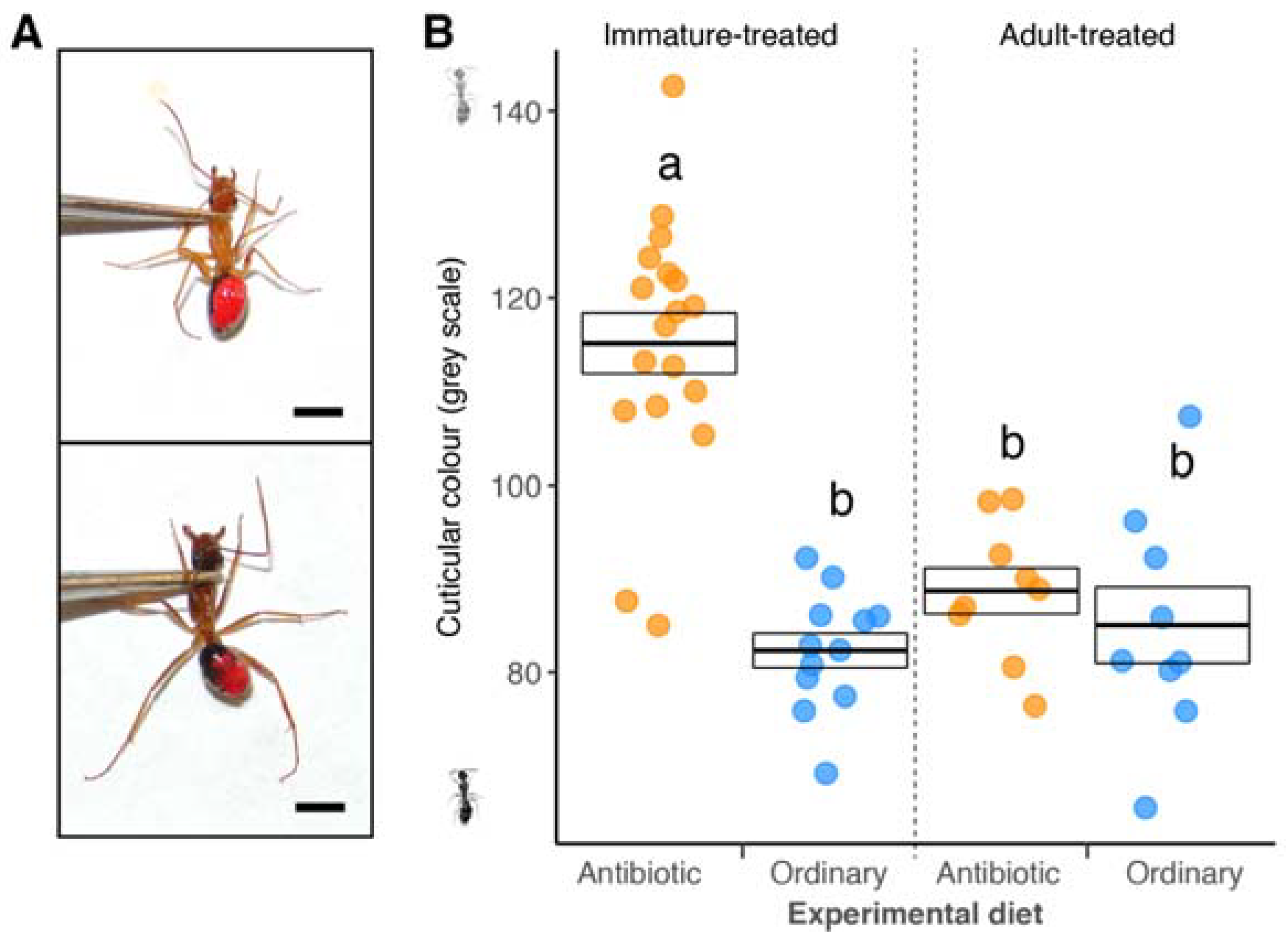
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Data Accessibility
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moran, N.A.; McCutcheon, J.P.; Nakabachi, A. Genomics and evolution of heritable bacterial symbionts. Annu. Rev. Genet. 2008, 42, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Baumann, P. Biology of bacteriocyte-associated endosymbionts of plant sap-sucking insects. Annu. Rev. Microbiol. 2005, 59, 155–189. [Google Scholar] [CrossRef] [PubMed]
- Wernegreen, J.J. Genome evolution in bacterial endosymbionts of insects. Nat. Rev. Genet. 2002, 3, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.M.; Moran, N.A. Heritable symbiosis: The advantages and perils of an evolutionary rabbit hole. Proc. Natl. Acad. Sci. USA 2015, 112, 10169–10176. [Google Scholar] [CrossRef] [PubMed]
- Nakabachi, A.; Shigenobu, S.; Sakazume, N.; Shiraki, T.; Hayashizaki, Y.; Carninci, P.; Ishikawa, H.; Kudo, T.; Fukatsu, T. Transcriptome analysis of the aphid bacteriocyte, the symbiotic host cell that harbors an endocellular mutualistic bacterium, Buchnera. Proc. Natl. Acad. Sci. USA 2005, 102, 5477–5482. [Google Scholar] [CrossRef] [PubMed]
- Zientz, E.; Dandekar, T.; Gross, R. Metabolic interdependence of obligate intracellular bacteria and their insect hosts. Microbiol. Mol. Biol. Rev. 2004, 68, 745–770. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.K.; Moran, N.A. Aphid genome expression reveals host-symbiont cooperation in the production of amino acids. Proc. Natl. Acad. Sci. USA 2011, 108, 2849–2854. [Google Scholar] [CrossRef] [PubMed]
- Gerardo, N.M.; Altincicek, B.; Anselme, C.; Atamian, H.; Barribeau, S.M.; de Vos, M.; Duncan, E.J.; Evans, J.D.; Gabaldón, T.; Ghanim, M.; et al. Immunity and other defenses in pea aphids, Acyrthosiphon pisum. Genome Biol. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.; Gibbs, R.A.; Gerardo, N.M.; Moran, N.; Nakabachi, A.; Stern, D.; Tagu, D.; Wilson, A.C.C.; Muzny, D.; Kovar, C.; et al. Genome sequence of the pea aphid Acyrthosiphon pisum. PLoS Biol. 2010, 8. [Google Scholar] [CrossRef] [Green Version]
- Nakabachi, A.; Ishikawa, H.; Kudo, T. Extraordinary proliferation of microorganisms in aposymbiotic pea aphids, Acyrthosiphon pisum. J. Invertebr. Pathol. 2003, 82, 152–161. [Google Scholar] [CrossRef]
- Stavrinides, J.; McCloskey, J.K.; Ochman, H. Pea aphid as both host and vector for the phytopathogenic bacterium Pseudomonas syringae. Appl. Environ. Microbiol. 2009, 75, 2230–2235. [Google Scholar] [CrossRef] [PubMed]
- Anselme, C.; Vallier, A.; Balmand, S.; Fauvarque, M.O.; Heddi, A. Host PGRP gene expression and bacterial release in endosymbiosis of the weevil Sitophilus zeamais. Appl. Environ. Microbiol. 2006, 72, 6766–6772. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, A.; Charif, D.; Vincent-Monegat, C.; Vallier, A.; Gavory, F.; Wincker, P.; Heddi, A. Host gene response to endosymbiont and pathogen in the cereal weevil Sitophilus oryzae. BMC Microbiol. 2012, 12, S14. [Google Scholar] [CrossRef] [PubMed]
- Shigenobu, S.; Stern, D.L. Aphids evolved novel secreted proteins for symbiosis with bacterial endosymbiont. Proc. R. Soc. B Biol. Sci. 2013, 280. [Google Scholar] [CrossRef] [PubMed]
- Anselme, C.; Perez-Brocal, V.; Vallier, A.; Vincent-Monegat, C.; Charif, D.; Latorre, A.; Moya, A.; Heddi, A. Identification of the Weevil immune genes and their expression in the bacteriome tissue. BMC Biol. 2008, 6, 43. [Google Scholar] [CrossRef] [PubMed]
- Login, F.H.; Heddi, A. Insect immune system maintains long-term resident bacteria through a local response. J. Insect Physiol. 2013, 59, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Simonet, P.; Gaget, K.; Balmand, S.; Ribeiro Lopes, M.; Parisot, N.; Buhler, K.; Duport, G.; Vulsteke, V.; Febvay, G.; Heddi, A.; et al. Bacteriocyte cell death in the pea aphid/Buchnera symbiotic system. Proc. Natl. Acad. Sci. USA 2018, 115, E1819–E1828. [Google Scholar] [CrossRef] [PubMed]
- Wernegreen, J.J.; Kauppinen, S.N.; Brady, S.G.; Ward, P.S. One nutritional symbiosis begat another: Phylogenetic evidence that the ant tribe Camponotini acquired Blochmannia by tending sap-feeding insects. BMC Evol. Biol. 2009, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Schroder, D.; Deppisch, H.; Obermayer, M.; Krohne, G.; Stackebrandt, E.; Holldobler, B.; Goebel, W.; Gross, R. Intracellular endosymbiotic: Bacteria of Camponotus species (carpenter ants): Systematics, evolution and ultrastructural characterization. Mol. Microbiol. 1996, 21, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Sauer, C.; Stackebrandt, E.; Gadau, J.; Holldobler, B.; Gross, R. Systematic relationships and cospeciation of bacterial endosymbionts and their carpenter ant host species: Proposal of the new taxon Candidatus Blochmannia gen. nov. Int. J. Syst. Evol. Micrbiol. 2000, 50, 1877–1886. [Google Scholar] [CrossRef] [PubMed]
- Sauer, C.; Dudaczek, D.; Holldobler, B.; Gross, R. Tissue localization of the endosymbiotic bacterium “Candidatus Blochmannia floridanus” in adults and larvae of the carpenter ant Camponotus floridanus. Appl. Environ. Microbiol. 2002, 68, 4187–4193. [Google Scholar] [CrossRef] [PubMed]
- Degnan, P.H.; Lazarus, A.B.; Brock, C.D.; Wernegreen, J.J. Host-symbiont stability and fast evolutionary rates in an ant-bacterium association: cospeciation of Camponotus species and their endosymbionts, Candidatus Blochmannia. Syst. Biol. 2004, 53, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Gil, R.; Silva, F.J.; Zientz, E.; Delmotte, F.; Gonzalez-Candelas, F.; Latorre, A.; Rausell, C.; Kamerbeek, J.; Gadau, J.; Holldobler, B.; et al. The genome sequence of Blochmannia floridanus: Comparative analysis of reduced genomes. Proc. Natl. Acad. Sci. USA 2003, 100, 9388–9393. [Google Scholar] [CrossRef] [PubMed]
- Degnan, P.H.; Lazarus, A.B.; Wernegreen, J.J. Genome sequence of Blochmannia pennsylvanicus indicates parallel evolutionary trends among bacterial mutualists of insects. Genome Res. 2005, 15, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Feldhaar, H.; Straka, J.; Krischke, M.; Berthold, K.; Stoll, S.; Mueller, M.J.; Gross, R. Nutritional upgrading for omnivorous carpenter ants by the endosymbiont Blochmannia. BMC Biol. 2007, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.L.; Thompson, J.W.; Dubois, L.G.; Moseley, M.A.; Wernegreen, J.J. Proteomic analysis of an unculturable bacterial endosymbiont (Blochmannia) reveals high abundance of chaperonins and biosynthetic enzymes. J. Proteome Res. 2013, 12, 704–718. [Google Scholar] [CrossRef] [PubMed]
- Wolschin, F.; Holldobler, B.; Gross, R.; Zientz, E. Replication of the endosymbiotic bacterium Blochmannia floridanus is correlated with the developmental and reproductive stages of its ant host. Appl. Environ. Microbiol. 2004, 70, 4096–4102. [Google Scholar] [CrossRef] [PubMed]
- Stoll, S.; Feldhaar, H.; Fraunholz, M.J.; Gross, R. Bacteriocyte dynamics during development of a holometabolous insect, the carpenter ant Camponotus floridanus. BMC Microbiol. 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Zientz, E.; Beyaert, N.; Gross, R.; Feldhaar, H. Relevance of the endosymbiosis of Blochmannia floridanus and carpenter ants at different stages of the life cycle of the host. Appl. Environ. Microbiol. 2006, 72, 6027–6033. [Google Scholar] [CrossRef] [PubMed]
- Stoll, S.; Feldhaar, H.; Gross, R. Transcriptional profiling of the endosymbiont Blochmannia floridanus during different developmental stages of its holometabolous ant host. Environ. Microbiol. 2009, 11, 877–888. [Google Scholar] [CrossRef] [PubMed]
- De Souza, D.J.; Bezier, A.; Depoix, D.; Drezen, J.M.; Lenoir, A. Blochmannia endosymbionts improve colony growth and immune defence in the ant Camponotus fellah. BMC Microbiol. 2009, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratzka, C.; Gross, R.; Feldhaar, H. Gene expression analysis of the endosymbiont-bearing midgut tissue during ontogeny of the carpenter ant Camponotus floridanus. J. Insect Physiol. 2013, 59, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Kupper, M.; Stigloher, C.; Feldhaar, H.; Gross, R. Distribution of the obligate endosymbiont Blochmannia floridanus and expression analysis of putative immune genes in ovaries of the carpenter ant Camponotus floridanus. Arthropod Struct. Dev. 2016, 45, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Ratzka, C.; Liang, C.G.; Dandekar, T.; Gross, R.; Feldhaar, H. Immune response of the ant Camponotus floridanus against pathogens and its obligate mutualistic endosymbiont. Insect Biochem. Mol. Boil. 2011, 41, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Oster, G.F.; Wilson, E.O. Caste and Ecology in the Social Insects, 1978 ed.; Monographs in Population Biology; Princeton University Press: Princeton, NJ, USA, 1978; Volume 12, pp. 1–352. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Wilson, E.O. Which are the most prevalent ant genera. Stud. Entomol. 1976, 19, 187–200. [Google Scholar]
- Cremer, S.; Pull, C.D.; Furst, M.A. Social immunity: Emergence and evolution of colony-level disease protection. Annu. Rev. Entomol. 2018, 63, 105–123. [Google Scholar] [CrossRef] [PubMed]
- Cremer, S.; Armitage, S.A.O.; Schmid-Hempel, P. Social immunity. Curr. Biol. 2007, 17, R693–R702. [Google Scholar] [CrossRef] [PubMed]
- Golkar, L.; LeBrun, R.A.; Ohayon, H.; Gounon, P.; Papierok, B.; Brey, P.T. Variation of larval susceptibility to Lagenidium giganteum in three mosquito species. J. Invertebr. Pathol. 1993, 62, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Feldhaar, H.; Gross, R. Immune reactions of insects on bacterial pathogens and mutualists. Microbes Infect. 2008, 10, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: Perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies. Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef] [PubMed]
- De Souza, D.J.; Devers, S.; Lenoir, A. Blochmannia endosymbionts and their host, the ant Camponotus fellah: Cuticular hydrocarbons and melanization. C. R. Biol. 2011, 334, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Armitage, S.A.O.; Boomsma, J.J. The effects of age and social interactions on innate immunity in a leaf-cutting ant. J. Insect Physiol. 2010, 56, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, S.; Rajakumar, R.; Abouheif, E.; Szyf, M. Epigenetic variation in the Egfr gene generates quantitative variation in a complex trait in ants. Nat. Commun. 2015, 6, 6513. [Google Scholar] [CrossRef] [PubMed]
- Avulova, S.; Rosengaus, R.B. Losing the battle against fungal infection: Suppression of termite immune defenses during mycosis. J. Insect Physiol. 2011, 57, 966–971. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Wills, B.D.; Powell, S.; Rivera, M.D.; Suarez, A.V. Correlates and consequences of worker polymorphism in ants. Annu. Rev. Entomol. 2018, 63, 575–598. [Google Scholar] [CrossRef] [PubMed]
- Dussutour, A.; Simpson, S.J. Carbohydrate regulation in relation to colony growth in ants. J. Exp. Biol. 2008, 211, 2224–2232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wills, B.D.; Chong, C.D.; Wilder, S.M.; Eubanks, M.D.; Holway, D.A.; Suarez, A.V. Effect of carbohydrate supplementation on investment into offspring number, size, and condition in a social insect. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.E. Developmental and physiological determinants of caste in social Hymenoptera—Evolutionary implications. Am. Nat. 1986, 128, 13–34. [Google Scholar] [CrossRef]
- Wheeler, D.E. The developmental basis of worker caste polymorphism in ants. Am. Nat. 1991, 138, 1218–1238. [Google Scholar] [CrossRef]
- Smith, C.R.; Anderson, K.E.; Tillberg, C.V.; Gadau, J.; Suarez, A.V. Caste determination in a polymorphic social insect: Nutritional, social, and genetic factors. Am. Nat. 2008, 172, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.M.; Hussain, M.; O’Neill, S.L.; Asgari, S. Wolbachia uses a host microRNA to regulate transcripts of a methyltransferase, contributing to dengue virus inhibition in Aedes aegypti. Proc. Natl. Acad. Sci. USA 2013, 110, 10276–10281. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.X.H.; Woolfit, M.; Huttley, G.A.; Rances, E.; Caragata, E.P.; Popovici, J.; O’Neill, S.L.; McGraw, E.A. Infection with a virulent strain of Wolbachia disrupts genome wide-patterns of cytosine methylation in the mosquito Aedes aegypti. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Thairu, M.W.; Hansen, A.K. Novel insights into insect-microbe interactions—Role of epigenomics and small RNAs. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.; Schrader, L.; Gil, R.; Manzano-Marin, A.; Florez, L.; Wheeler, D.; Werren, J.H.; Latorre, A.; Heinze, J.; Kaltenpoth, M.; et al. A novel intracellular mutualistic bacterium in the invasive ant Cardiocondyla obscurior. ISME J. 2016, 10, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.O. Cuticular Sclerotization and Tanning. In Insect Molecular Biology and Biochemistry; Gilbert, L.I., Ed.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2012; pp. 167–192. [Google Scholar]
- Qiao, L.; Du, M.H.; Liang, X.; Hao, Y.J.; He, X.; Si, F.L.; Mei, T.; Chen, B. Tyrosine Hydroxylase is crucial for maintaining pupal tanning and immunity in Anopheles sinensis. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Rolff, J. Effects of age and gender on immune function of dragonflies (Odonata, Lestidae) from a wild population. Can. J. Zool. 2001, 79, 2176–2180. [Google Scholar] [CrossRef]
- Doums, C.; Moret, Y.; Benelli, E.; Schmid-Hempel, P. Senescence of immune defence in Bombus workers. Ecol. Entomol. 2002, 27, 138–144. [Google Scholar] [CrossRef]
- Amdam, G.V.; Aase, A.L.T.O.; Seehuus, S.-C.; Fondrk, M.K.; Norberg, K.i.; Hartfelder, K. Social reversal of immunosenescence in honey bee workers. Exp. Gerontol. 2005, 40, 939–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakane, Y.; Lomakin, J.; Beeman, R.W.; Muthukrishnan, S.; Gehrke, S.H.; Kanost, M.R.; Kramer, K.J. Molecular and functional analyses of amino acid decarboxylases involved in cuticle tanning in Tribolium castaneum. J. Biol. Chem. 2009, 284, 16584–16594. [Google Scholar] [CrossRef] [PubMed]
- Roseland, C.R.; Kramer, K.J.; Hopkins, T.L. Cuticular strength and pigmentation of rust-red and black strains of Tribolium castaneum—Correlation with catecholamine and beta-alanine content. Insect Biochem. 1987, 17, 21–28. [Google Scholar] [CrossRef]
- Hopkins, T.L.; Kramer, K.J. Insect Cuticle Sclerotization. Ann. Rev. Entomol. 1992, 37, 273–302. [Google Scholar] [CrossRef]
- Klotz, J.H.; Mangold, J.R.; Vail, K.M.; Davis, L.R.; Patterson, R.S. A survey of the urban pest ants (Hymenoptera, Formicidae) of peninsular Florida. Fla. Entomol. 1995, 78, 109–118. [Google Scholar] [CrossRef]
- Klotz, J.H.; Greenberg, L.; Reid, B.L.; Davis, L. Spatial distribution of colonies of three carpenter ants, Camponotus pennsylvanicus, Camponotus floridanus, Camponotus laevigatus (Hymenoptera: Formicidae). Sociobiology 1998, 32, 51–62. [Google Scholar]
- Collingwood, C.A.; Agosti, D.; Sharaf, M.R.; van Harten, A. Order hymenoptera, family Formicidae. Arthropod Fauna UAE 2011, 4, 405–474. [Google Scholar]
- Wilson, K.; Cotter, S.C. Density-dependent prophylaxis in insects. In Phenotypic Plasticity of Insects: Mechanisms and Consequences; Ananthakrishnan, T.N., Whitman, T.W., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2009; pp. 381–420. [Google Scholar]
- Tang, H.P.; Kambris, Z.; Lemaitre, B.; Hashimoto, C. Two proteases defining a melanization cascade in the immune system of Drosophila. J. Biol. Chem. 2006, 281, 28097–28104. [Google Scholar] [CrossRef] [PubMed]
- Dean, P.; Gadsden, J.C.; Richards, E.H.; Edwards, J.P.; Keith Charnley, A.; Reynolds, S.E. Modulation by eicosanoid biosynthesis inhibitors of immune responses by the insect Manduca sexta to the pathogenic fungus Metarhizium anisopliae. J. Invertebr. Pathol. 2002, 79, 93–101. [Google Scholar] [CrossRef]
- Wei, G.; Lai, Y.L.; Wang, G.D.; Chen, H.; Li, F.; Wang, S.B. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, W.O.; Boomsma, J.J. Let your enemy do the work: within-host interactions between two fungal parasites of leaf-cutting ants. Proc. R. Soc. B Biol. Sci. 2004, 271 (Suppl. 3), S104–S106. [Google Scholar] [CrossRef] [PubMed]
- Konrad, M.; Pull, C.D.; Metzler, S.; Seif, K.; Naderlinger, E.; Grasse, A.V.; Cremer, S. Ants avoid superinfections by performing risk-adjusted sanitary care. Proc. Natl. Acad. Sci. USA 2018, 115, 2782–2787. [Google Scholar] [CrossRef] [PubMed]
- Vojvodic, S.; Boomsma, J.J.; Eilenberg, J.; Jensen, A.B. Virulence of mixed fungal infections in honey bee brood. Front. Zool. 2012, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.N.; Hughes, W.O.H. Arboreality and the evolution of disease resistance in ants. Ecol. Entomol. 2011, 36, 588–595. [Google Scholar] [CrossRef]
- Shykoff, J.A.; Schmid-Hempel, P. Parasites and the advantage of genetic variability within social insect colonies. Proc. R. Soc. B Biol. Sci. 1991, 243, 55–58. [Google Scholar] [CrossRef]
- Tragust, S.; Mitteregger, B.; Barone, V.; Konrad, M.; Ugelvig, L.V.; Cremer, S. Ants disinfect fungus-exposed brood by oral uptake and spread of their poison. Curr. Biol. 2013, 23, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Hughes, W.O.H.; Eilenberg, J.; Boomsma, J.J. Trade-offs in group living: transmission and disease resistance in leaf-cutting ants. Proc. R. Soc. B Biol. Sci. 2002, 269, 1811–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugelvig, L.V.; Cremer, S. Social prophylaxis: Group interaction promotes collective immunity in ant colonies. Curr. Biol. 2007, 17, 1967–1971. [Google Scholar] [CrossRef] [PubMed]
- Konrad, M.; Vyleta, M.L.; Theis, F.J.; Stock, M.; Tragust, S.; Klatt, M.; Drescher, V.; Marr, C.; Ugelvig, L.V.; Cremer, S. Social transfer of pathogenic fungus promotes active immunisation in ant colonies. PLoS Biol. 2012, 10. [Google Scholar] [CrossRef] [PubMed]
- Bos, N.; Lefèvre, T.; Jensen, A.B.; d’Ettorre, P. Sick ants become unsociable. J. Evol. Biol. 2011, 25, 342–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinotte, V.M.; Freedman, S.N.; Ugelvig, L.V.; Seid, M.A. Camponotus floridanus Ants Incur a Trade-Off between Phenotypic Development and Pathogen Susceptibility from Their Mutualistic Endosymbiont Blochmannia. Insects 2018, 9, 58. https://doi.org/10.3390/insects9020058
Sinotte VM, Freedman SN, Ugelvig LV, Seid MA. Camponotus floridanus Ants Incur a Trade-Off between Phenotypic Development and Pathogen Susceptibility from Their Mutualistic Endosymbiont Blochmannia. Insects. 2018; 9(2):58. https://doi.org/10.3390/insects9020058
Chicago/Turabian StyleSinotte, Veronica M., Samantha N. Freedman, Line V. Ugelvig, and Marc A. Seid. 2018. "Camponotus floridanus Ants Incur a Trade-Off between Phenotypic Development and Pathogen Susceptibility from Their Mutualistic Endosymbiont Blochmannia" Insects 9, no. 2: 58. https://doi.org/10.3390/insects9020058