Resistance to Permethrin, β-cyfluthrin, and Diazinon in Florida Horn Fly Populations



Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Insecticide Susceptibility Assay
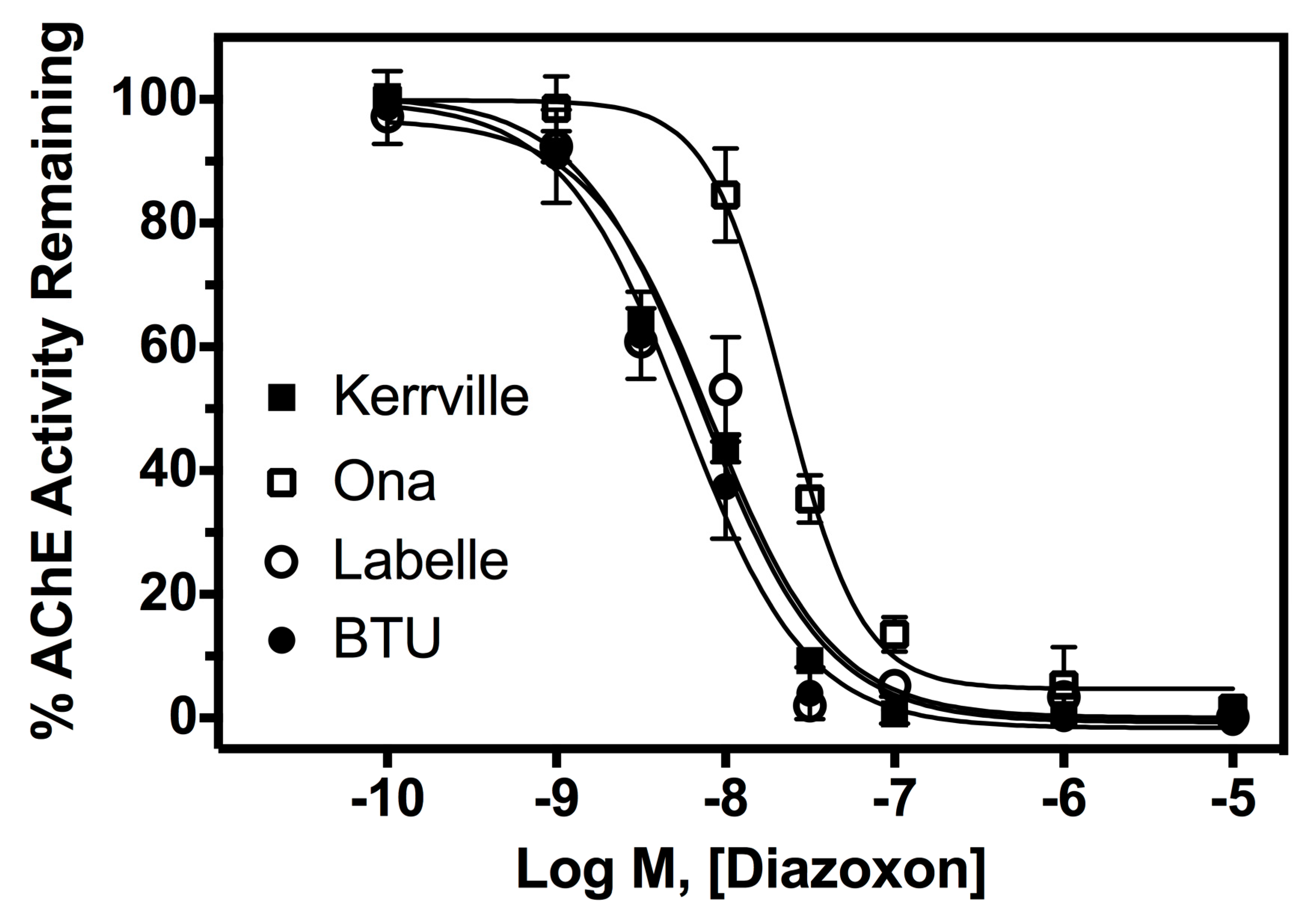
2.3. AChE Inhibition Assay
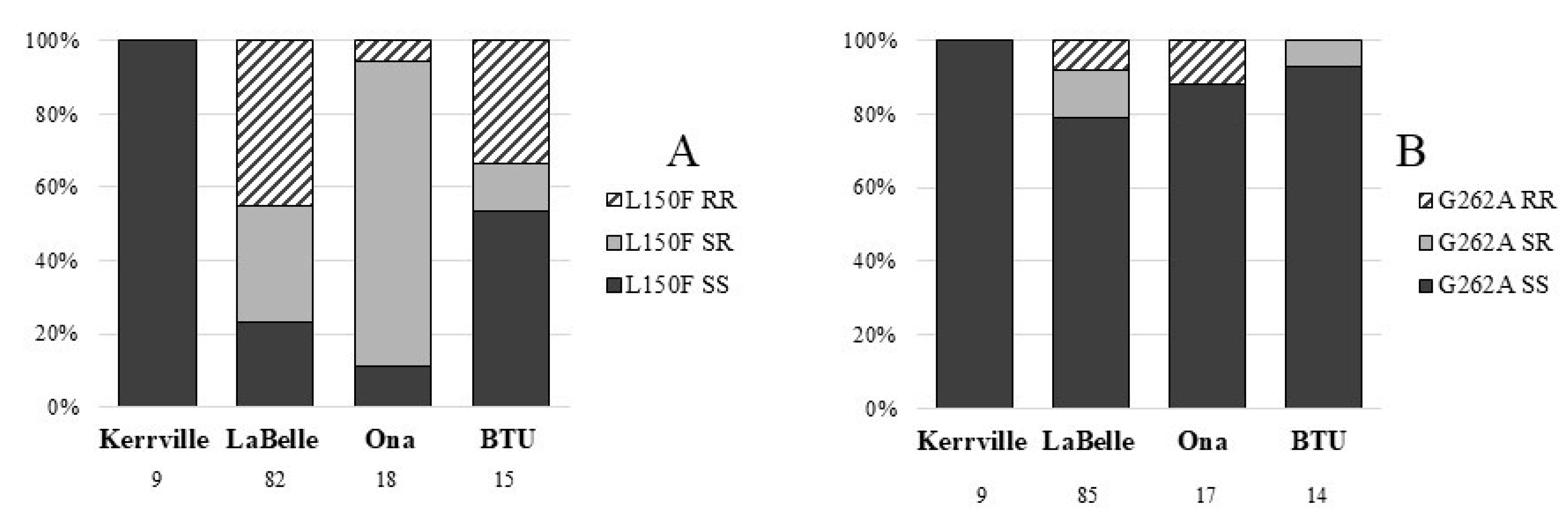
2.4. Genotyping
2.5. cP450 and CBE Assays
2.6. Statistics
3. Results
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kunz, S.E.; Murrel, K.D.; Lambert, G.; James, L.F.; Terrill, C.E. Estimated losses of livestock to pests. In CRC Handbook of Pest Management in Agriculture, 2nd ed.; Pimentel, D., Ed.; CRC Press: Boca Raton, FL, USA, 1991; Volume I, pp. 69–98. ISBN 978-0849338441. [Google Scholar]
- Li, A.Y.; Lohmeyer, K.H.; Miller, J.A. Dynamics and mechanisms of permethrin resistance in a field population of the horn fly, Haematobia irritans. Insect Sci. 2009, 16, 175–184. [Google Scholar] [CrossRef]
- Kunz, S.E.; Schmidt, C.D. The pyrethroid resistance problem in the horn fly (Diptera: Muscidae). J. Agric. Entomol. 1985, 2, 358–363. [Google Scholar]
- Byford, R.L.; Quisenberry, S.S.; Sparks, T.C.; Lockwood, J.A. Spectrum of insecticide cross-resistance in pyrethroid-resistant populations of Haematobia irritans (Diptera: Muscidae). J. Econ. Entomol. 1985, 78, 768–773. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Hinkle, N.C. Pyrethroid resistance in horn flies: The problem, causes, and possible solutions. J. Agric. Entomol. 1985, 2, 317–324. [Google Scholar]
- Guerrero, F.D.; Jamroz, R.C.; Kammlah, D.; Kunz, S.E. Toxicological and molecular characterization of pyrethroid-resistant horn flies, Haematobia irritans: Identification of kdr and super-kdr point mutations. Insect Biochem. Molec. Biol. 1997, 27, 745–755. [Google Scholar] [CrossRef]
- Bull, D.L.; Harris, R.L.; Pryor, N.W. The contribution of metabolism to pyrethroid and DDT resistance in the horn fly (Diptera: Muscidae). J. Econ. Entomol. 1988, 81, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Oyarzun, M.P.; Quiroz, A.; Birkett, M.A. Insecticide resistance in the horn fly: Alternative control strategies. Med. Vet. Entomol. 2008, 22, 188–202. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.S.; Martinez-Torres, D.; Hick, C.A.; Devonshire, A.L. Identification of mutations in the housefly para-type sodium channel gene associated with knockdown resistance (kdr) to pyrethroid insecticides. MGG 1996, 252, 51–60. [Google Scholar] [PubMed]
- Oyarzun, M.P.; Li, A.Y.; Figueroa, C.C. High levels of insecticide resistance in introduced horn fly (Diptera: Muscidae) populations and implications for management. J. Econ. Entomol. 2011, 104, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Barros, A.T.; Ottea, J.; Sanson, D.; Foil, L.D. Horn fly (Diptera: Muscidae) resistance to organophosphate insecticides. Vet. Parasitol. 2001, 96, 243–256. [Google Scholar] [CrossRef]
- Temeyer, K.B.; Brake, D.K.; Schlechte, K.G. Acetylcholinesterase of Haematobia irritans (Diptera: Muscidae): Baculovirus expression, biochemical properties, and organophosphate insensitivity of the G262A mutant. J. Med. Entom. 2012, 49, 589–594. [Google Scholar] [CrossRef]
- Temeyer, K.B.; Li, A.Y.; Lohmeyer, K.H.; Chen, A.C.; Olafson, P.U.; Sanson, D.W.; Foil, L.D. Acetylcholinesterase mutation in diazinon-resistant Haematobia irritans (L.) (Diptera: Muscidae). Vet. Parisitol. 2008, 154, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Foil, L.D.; Guerrero, F.D.; Bendele, K.G. Detection of target site resistance to pyrethroids and organophosphates in the horn fly using multiplex polymerase chain reaction. J. Med. Entomol. 2010, 47, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D.C.; Hinkle, N.C. A procedure for evaluation of horn fly, Haematobia irritans (L.), pyrethroid resistance by exposure to pyrethroid residues on glass. J. Agric. Entomol. 1986, 3, 100–102. [Google Scholar]
- Pitzer, J.B.; Kaufman, P.E.; TenBroeck, S.H. Assessing permethrin resistance in the stable fly (Diptera: Muscidae) in Florida by using laboratory selection and field evaluations. J. Econ. Entomol. 2010, 103, 2258–2263. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Anders, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Swale, D.R.; Tong, F.; Temeyer, K.B.; Li, A.; Lam, P.C.; Totrov, M.M.; Carlier, P.R.; Perez de Leon, A.A.; Bloomquist, J.R. Inhibitor profile of bis(n)-tacrines and N-methylcarbamates on acetylcholinesterase from Ripicephalus (Boophilus) microplus and Phlebotomus papatasi. Pestic. Biochem. Physiol. 2013, 106. [Google Scholar] [CrossRef] [PubMed]
- Jamroz, R.C.; Guerrero, F.D.; Kammlah, D.M.; Kunz, S.E. Role of the kdr and super-kdr sodium channel mutations in pyrethroid resistance: correlation of allelic frequency to resistance level in wild and laboratory populations of horn flies (Haematobia irritans). Insect Biochem. Molec. Biol. 1998, 28, 1031–1037. [Google Scholar] [CrossRef]
- Grant, D.F.; Bender, D.M.; Hammock, B.D. Quantative kinetic assays for glutathione s-transferase and general esterase in individual mosquitoes using an EIA reader. Insect Biochem. 1989, 19, 741–751. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: New York, NY, USA, 1971; pp. 1–350. ISBN 978-0521135900. [Google Scholar]
- Robertson, J.L.; Preisler, H.K.; Russell, R.M. PoloPlus Probit and Logit Analysis User’s Guide; LeOra Software: Petaluma, CA, USA, 2003; pp. 1–36. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Am. Mosq. Control Assoc. 1987, 3, 302–303. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D.C.; Marchiondo, A.A. Toxicity of diazinon to pyrethroid resistant and susceptible horn flies, Haematobia irritans (L.): Laboratory studies and field trials. J. Agric. Entomol. 1987, 4, 262–270. [Google Scholar]
- Li, A.Y.; Guerrero, F.D.; Pruett, J.H. Involvement of esterases in diazinon resistance and biphasic effects of piperonyl butoxide on diazinon toxicity to Haematobia irritans (Diptera: Muscidae). Pest. Biochm. Physiol. 2007, 87, 147–155. [Google Scholar] [CrossRef]
- Rinkevich, F.D.; Leichter, C.A.; Lazo, T.A.; Hardstone, M.C.; Scott, J.G. Variable fitness costs for pyrethroid resistance alleles in the house fly, Musca domestica, in the absence of insecticide pressure. Pest. Biochem. Physiol. 2013, 105, 161–168. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Joyce, J.A. High levels of resistance in horn flies (Diptera: Muscidae) selected with cyhalothrin. J. Econ. Entomol. 1992, 85, 1587–1593. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, F.D.; Pruett, J.H.; Kunz, S.E.; Kammlah, D.M. Esterase profiles of diazinon-susceptible and -resistant horn flies (Diptera: Muscidae). J. Econ. Entomol. 1999, 92, 286–292. [Google Scholar] [CrossRef]
- Barros, A.; Saueressig, T.M.; Gomes, A.S.; Koller, W.W.; Furlong, J.; Girao, E.S.; da Cunha Pinheiro, A.; de Paula Jardim Alves-Branco, F.; de Fatima Munhos Sapper, M.; Braga, R.M.; et al. Susceptibility of the horn fly, Haematobia irritans (Diptera: Muscidae), to insecticides in Brazil. Rev. Bras. Parasitol. Vet. 2012, 21, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Domingues, L.N.; Guerrero, F.D.; Foil, L.D. Simultaneous detection of pyrethroid, organophosphate, and cyclodiene target site resistance in Haematobia irritans (Diptera: Muscidae) by multiplex polymerase chain reaction. J. Med. Entomol. 2014, 51, 964–970. [Google Scholar] [CrossRef] [PubMed]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Snodgrass, G.L.; Chen, M.S. Comparative study on glutathione S-transferase activity, cDNA, and gene expression between malathion susceptible and resistant strains of the tarnished plant bug, Lygus lineolaris. Pest. Biochm. Physiol. 2007, 87, 62–72. [Google Scholar] [CrossRef]
- Yang, M.L.; Zhang, J.Z.; Zhu, K.Y.; Xuan, T.; Liu, X.J.; Guo, Y.P.; Ma, E.B. Mechanisms of organophosphate resistance in a field population of oriental migratory locust, Locusta migratoria manilensis (Meyen). Arch. Insect Biochem. Physiol. 2009, 71, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Muthusamy, R.; Suganya, R.; Gowri, M.; Shivakumar, M.S. Biochemical mechanisms of organophosphate and pyrethroid resistance in red hairy caterpillar Amsacta albistriga (Lepidoptera: Arctiidae). J. Saudi Soc. Agric. Sci. 2013, 12, 47–52. [Google Scholar] [CrossRef]
- Oremus, G.; Guerrero, F.D.; Alison, M.W., Jr.; Kimball, M.M.; Kim, J.H.; Foil, L.D. Effects of mid-season avermectin treatments on pyrethroid resistance in horn fly (Diptera: Muscidae) populations at three locations in Louisiana. Vet. Parisitol. 2006, 141, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Barros, A.T.; Gomes, A.; Ismael, A.P.; Koller, W.W. Susceptibility to diazinon in populations of the horn fly, Haematobia irritans (Diptera: Muscidae), in central Brazil. Mem. Inst. Oswaldo. Cruz. 2002, 97, 905–908. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Site | Permethrin | β-cyfluthrin | Diazinon | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LC50 value A | 95% CI | Slope | RR B | LC50 value | 95% CI | Slope | RR | LC50 value | 95% CI | Slope | RR | |
| Kerrville | 0.023 | 0.016–0.032 | 2.47 | 0.0067 | 0.0044–0.0096 | 1.55 | 0.02 | 0.14–0.029 | 2.45 | |||
| Labelle | 1.53 * | 1.01–2.6 | 1.29 | 66.6 | ND C | ND | ND | ND | >0.295 D | ND | 0.158 | >14.7 |
| BTU | 0.13 * | 0.055–0.19 | 1.36 | 5.7 | 0.01 | 0.0013–0.13 | 0.79 | 1.4 | 0.06 * | 0.046–0.27 | 1.33 | 3 |
| Ona | 0.068 | 0.013–0.36 | 0.85 | 3.0 | <0.001 | ND | ND | <0.15 | ND E | ND | ND | ND |
| Collection Site | Cytochrome P450 | General Esterase |
|---|---|---|
| Kerrville | 29.1 ± 8.9 | 13.5 ± 5.4 |
| Labelle | 50.5 ± 8.22 | 40.6 ± 10.3 |
| Ona | 23.3 ± 10.1 | 23.0 ± 4.5 |
| BTU | 32.1 ± 18.4 | 32.3 ± 6.8 |
| ANOVA | ||
| F-Value | 0.93 | 2.69 |
| p-Value | 0.44 | 0.08 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holderman, C.J.; Swale, D.R.; Bloomquist, J.R.; Kaufman, P.E. Resistance to Permethrin, β-cyfluthrin, and Diazinon in Florida Horn Fly Populations. Insects 2018, 9, 63. https://doi.org/10.3390/insects9020063
Holderman CJ, Swale DR, Bloomquist JR, Kaufman PE. Resistance to Permethrin, β-cyfluthrin, and Diazinon in Florida Horn Fly Populations. Insects. 2018; 9(2):63. https://doi.org/10.3390/insects9020063
Chicago/Turabian StyleHolderman, Chris J., Daniel R. Swale, Jeffery R. Bloomquist, and Phillip E. Kaufman. 2018. "Resistance to Permethrin, β-cyfluthrin, and Diazinon in Florida Horn Fly Populations" Insects 9, no. 2: 63. https://doi.org/10.3390/insects9020063
APA StyleHolderman, C. J., Swale, D. R., Bloomquist, J. R., & Kaufman, P. E. (2018). Resistance to Permethrin, β-cyfluthrin, and Diazinon in Florida Horn Fly Populations. Insects, 9(2), 63. https://doi.org/10.3390/insects9020063