
Relationship between Organic Carbon and Opportunistic Pathogens in Simulated Glass Water Heaters


Abstract
:1. Introduction
2. Results
2.1. Correlations between Added Organic Carbon Concentration and 16S rRNA Gene Copies and HPC Counts


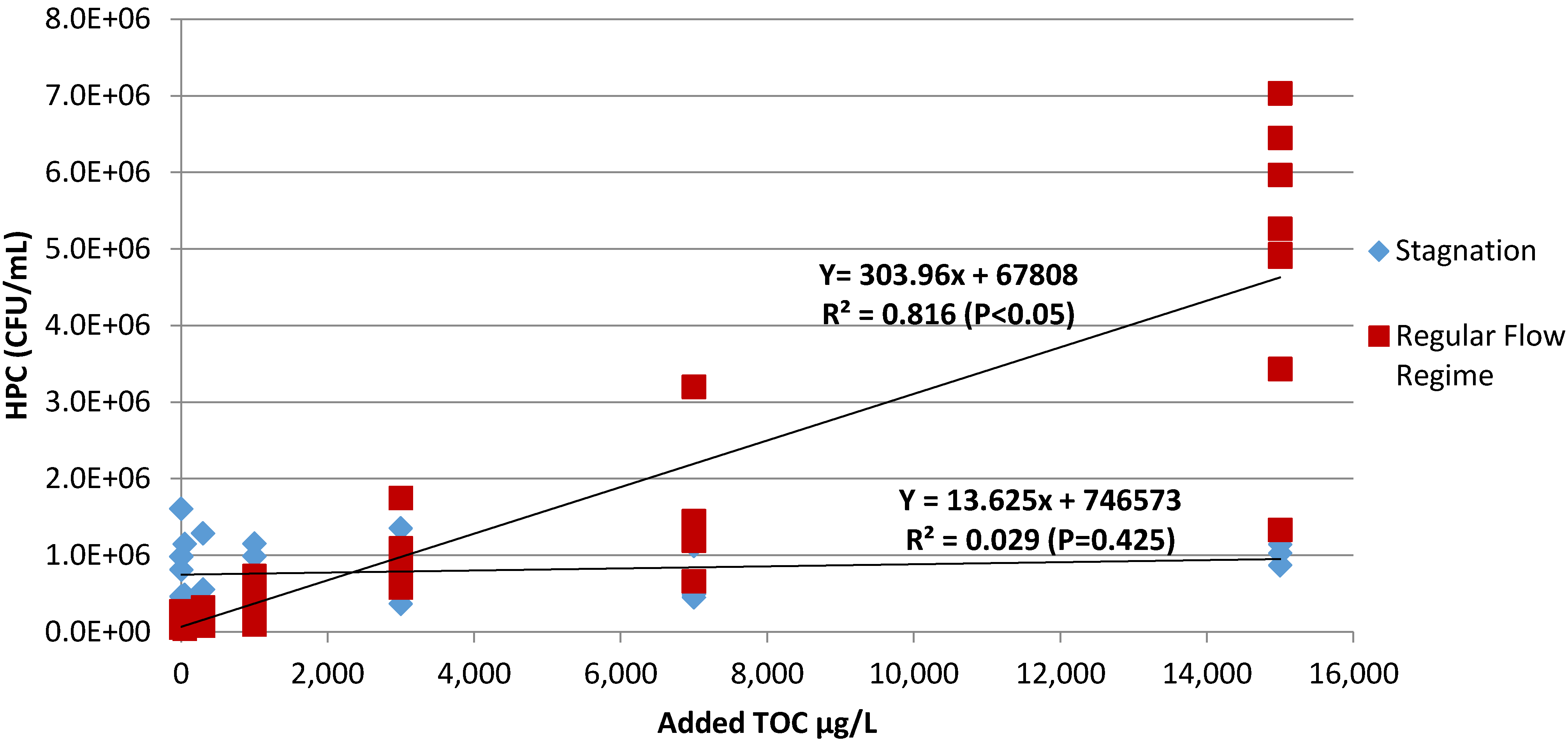{kind=link}
{kind=link}
{kind=link}
| Experiment Designator | Dates Performed | Average R2 Value 16S rRNA qPCR | Number of Time Points (n) 16S rRNA qPCR | Average R2 Value HPC | Number of Time Points (n) HPC |
|---|---|---|---|---|---|
| Preliminary Experiments | 12/1/2009–4/15/2010 | N/A | N/A | 0.952 | 11 |
| A1 | 4/28/2010–8/3/2010 | 0.971 | 3 | 0.889 | 9 |
| A2 | 10/13/2010–3/7/2011 | 0.626 | 3 | 0.645 | 3* |
| A3 | 3/31/2011–4/18/2011 | 0.168 | 3 | 0.295 | 3 |
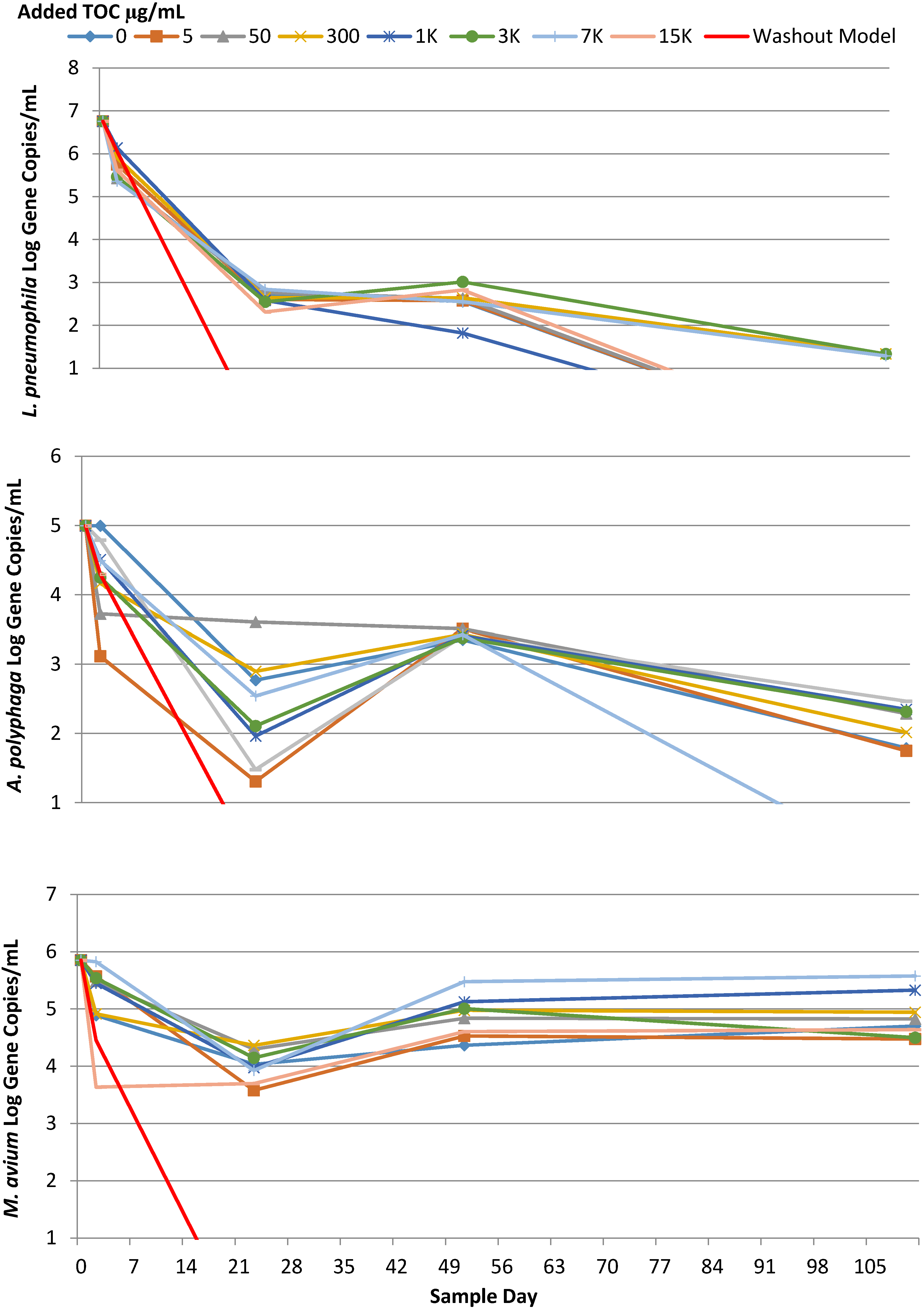
2.2. M. avium
| M. avium (R2 Value) qPCR | M. avium (R2 Value) Culture Enumeration | |
|---|---|---|
| Low Range TOC | Low Range TOC | |
| (0–1000 µg/L) | (0–1000 µg/L) | |
| Day 2 | 0.007 (p = 0.890) | 0.007 (p = 0.890) |
| Day 23 | 0.003 (p = 0.929) | 0.004 (p = 0.929) |
| Day 51 | 0.819 (p = 0.034) | 0.818 (p = 0.035) |
| Day 111 | 0.974 (p = 0.001) | N/A |
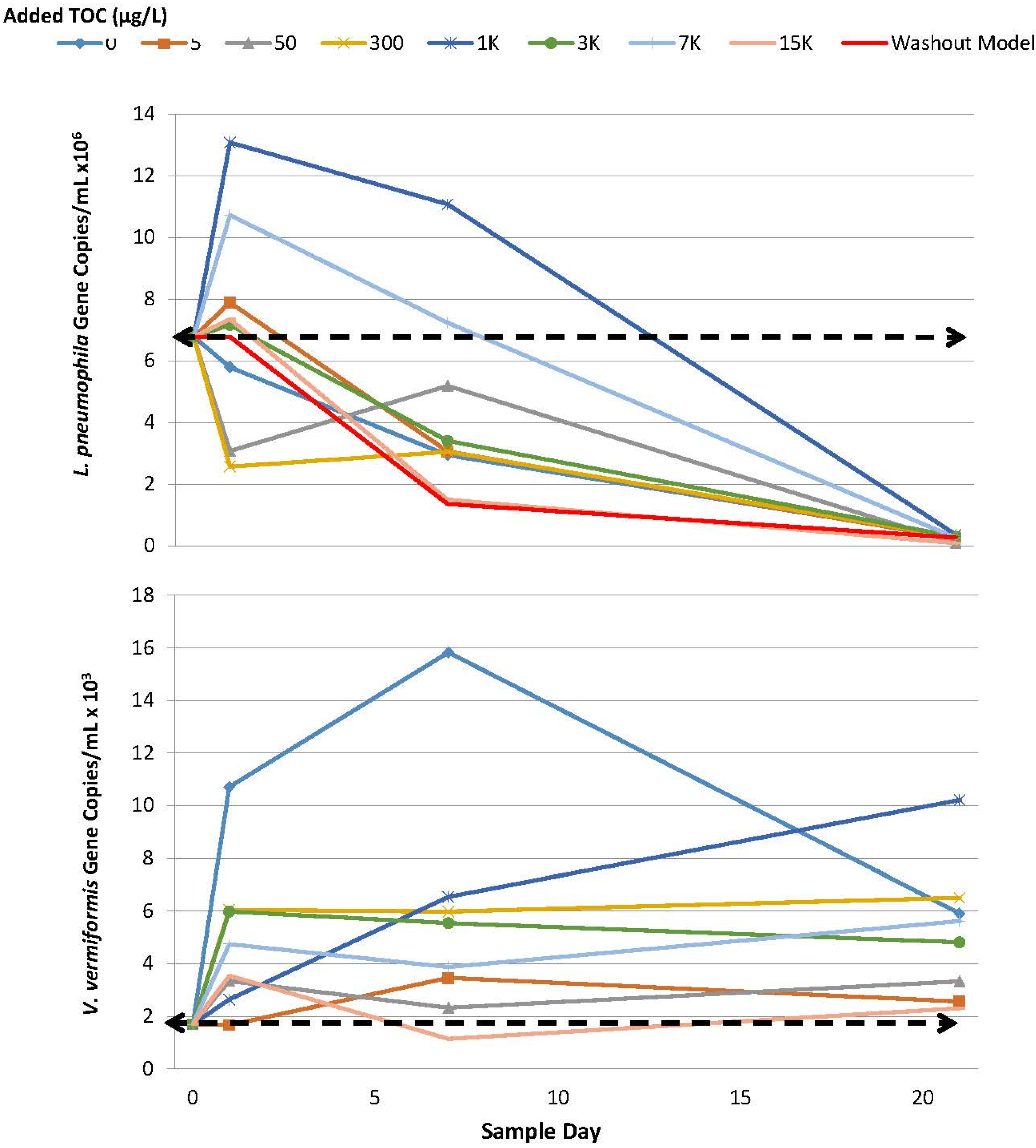
2.3. L. pneumophila
2.4. A. Polyphaga

| A. polyphaga (R2 Value) | |
|---|---|
| Low Range TOC | |
| (0–1000 µg/L) | |
| Day 2 | 0.991 (p = 0.004) |
| Day 23 | −0.125 (p = 0.559) |
| Day 51 | −0.118 (p = 0.569) |
| Day 111 | 0.683 (p = 0.084) |

3. Discussion
4. Experimental Section
- Water Source 1: Nanopure water with UV carbon destruction and Norton et al. (2004) recipe to create synthesized tap water [15] (Experiment: A1).
- Water Source 2: A mixture of 90% Water Source 1 mixed with 10% Blacksburg tap water that had been circulated through a granular activated carbon (GAC) filter. (Experiment A2)
- Water Source 3: A mixture of 90% Water Source 1 mixed with 10% source water from Blacksburg tap water that had been circulated through a GAC filter. Additional trace nutrients of manganese (6 µg/L), iron (42 µg/L) and zinc (375 µg/L) were added along with a stock solution of trace amino acids, at levels encountered in potable water systems [33] (Experiment A3).
4.1. First Phase of Testing: Experiment A1
4.2. First Set of Changes to Baseline: Experiment A2
4.3. Second Set of Changes to Baseline: Experiment A3
| Experiment | Inoculation/Stagnation Periods | Dates Performed | Water Formulation; Temperature; Water Change Rate |
|---|---|---|---|
| A1 | 23 April 2010 Inoculation with 10 mL from Blacksburg hot water tank Incubate 1 week at 37 °C | 5/18/10–8/3/10 | Norton et al., 2004; 37 °C; 3 water changes per week |
| 5 May 2010 Inoculation with A. polyphaga Incubate 1 week at 37 °C | |||
| 12 May 2010 Inoculation with L. pneumophila and M. avium Incubate 1 week at 37 °C | |||
| 18 May 2010 End stagnation period | |||
| A2 | 17 September 2010 Inoculation with A. polyphaga Incubate 1 week at 37 °C | 10/13/10–3/7/11 (5 months) | • 90% Norton et al., 2004/ 10% GAC filtered Blacksburg, VA tap water; 32 °C; 3 water changes per week |
| 27 Septermber 2010 Inoculation with L. pneumophila and M. avium Incubate 1 week at 37 °C | |||
| 13 October 2010 End stagnation period | |||
| A3 | 18 March 2010 Inoculation of V. vermiformis Incubate at 37 °C | 3/31/11–4/18/11 (3 weeks) | • 90% Norton et al., 2004/10% GAC filtered Blacksburg, VA tap water with addition of amino acid, Mg, Fe, Zn and pH decrease to 7.5; 32 °C; 1 water change per week |
| 21 March 2010 Inoculation of L. pneumophila Incubate at 37 °C | |||
| 31 March 2011 End stagnation period |
4.4. Nucleic Acid Extraction and Quantitative Polymerase Chain Reaction (qPCR) Deoxyribonucleic Acid (DNA) Extraction
4.5. Quantitative Polymerase Chain Reaction (qPCR)
4.6. Culture and Microscopic-Based Detection and Enumeration
4.6.1. L. pneumophila
4.6.2. V. Vermiformis/A. Polyphaga
4.6.3. M. avium
4.6.4. Heterotrophic Plate Counts (HPCs)
4.7. Total Organic Carbon (TOC) and Biodegradable Dissolved Organic Carbon (BDOC) Measurement
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References amd Notes
- Yoder, J.; Roberts, V.; Craun, G.F.; Hill, V.; Hicks, L.; Alexander, N.T.; Radke, V.; Calderon, R.L.; Hlavsa, M.C.; Beach, M.J.; et al. Surveillance for Waterborne Disease and Outbreaks Associated with Drinking Water and Water not Intended for Drinking—United States, 2005–2006. MMWR Surveill. Summ. 2008, 57, 39–62. [Google Scholar] [PubMed]
- Edwards, M.A.; Pruden, A.; Falkinham, J.O., III; Brazeau, R.; Williams, K.; Wang, H.; Martin, A.; Rhoads, W. Relationship Between Biodegradable Organic Matter and Pathogen Concentrations in Premise Plumbing; Water Research Foundation: Denver, CO, USA, 2013; p. 111. [Google Scholar]
- Bartrand, T.A.; Causey, J.; Clancy, J. Naegleria fowleri: An emerging drinking water pathogen. J. Am. Water Works Assoc. 2014, 106, E418–E432. [Google Scholar] [CrossRef]
- Falkinham, J.O., III; Hilborn, E.D.; Arduino, M.J.; Pruden, A.; Edwards, M.A. Epidemiology and Ecology of Opportunistic Premise Plumbing Pathogens: Legionella pneumophila, Mycobacterium avium, and Pseudomonas aeruginosa. Environ. Health Perspect. 2015, (in press). [Google Scholar] [CrossRef] [PubMed]
- Pruden, A.; Edwards, M.A.; Falkinham, J.O., III; Arduino, M.; Bird, J.; Birdnow, R.; Bédard, E.; Camper, A.; Clancy, J.; Hilborn, E.; et al. Research Needs for Opportunistic Pathogens in Premise Plumbing: Methodology, Microbial Ecology, and Epidemiology. Water Research Foundation Project 4379 Final Report; Water Research Foundation: Denver, CO, USA, 2013; p. 188. [Google Scholar]
- Snoeyink, V.S.; Hass, C.H.; Boulos, P.F.; Burlingame, G.A.; Camper, A.K.; Clark, R.N.; Edwards, M.A.; LeChevallier, M.W.; McMullen, L.D.; Moe, C.L.; et al. Drinking Water Distribution Systems: Assessing and Reducing Risks; National Research Council: Washington, D.C. USA, 2006; p. 392. [Google Scholar]
- Wang, H.; Edwards, M.; Falkinham, J.O., III; Pruden, A. Probiotic Approach to Pathogen Control in Potable Water Systems? Environ. Sci. Technol. 2013, 47, 10117–10128. [Google Scholar] [CrossRef] [PubMed]
- Pruden, A.; Edwards, M.A.; Falkinham, J.O., III; Arduino, M.; Bird, J.; Birdnow, R.; Bédard, E.; Camper, A.; Clancy, J.; Hilborn, E.; et al. Research Needs for Opportunistic Pathogens in Premise Plumbing: Methodology, Microbial Ecology, and Epidemiology. Water Research Foundation Project 4379 Final Report; Water Research Foundation: Denver, CO, USA, 2013; p. 188. [Google Scholar]
- Rhoads, W.; Pruden, A.; Edwards, M.A. Anticipating Challenges with in-building Disinfection for Control of Opportunistic Pathogens. Water Environ. Res. 2014, 86, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Van der Kooij, D.; van der Wielen, P.W.J.J. Microbial Growth in Drinking-Water Supplies Problems, Causes, Control and Research Needs; IWA Publishing: London, UK, 2013. [Google Scholar]
- Thomas, J.M.; Ashbolt, N.J. Do free-living Amoeba in treated drinking water system present an emerging health risk? Environ. Sci. Technol. 2011, 45, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Van der Kooij, D. Assimilable organic carbon as indicator of bacterial regrowth. J. Am. Water Works Assoc. 1992, 84, 57. [Google Scholar]
- Camper, A.; Brastrup, K.; Sandvig, A.; Clement, J.; Spencer, C.; Capuzzi, A. Effect of distribution system materials on bacterial regrowth. J. Am. Water Works Assoc. 2003, 95, 107–121. [Google Scholar]
- LeChevallier, M.W.; Schulz, W.; Lee, R.G. Bacterial nutrients in drinking water. Appl. Environ. Microbiol. 1991, 57, 857–862. [Google Scholar] [PubMed]
- Norton, C.D.; LeChevallier, M.W.; Falkinham, J.O., III. Survival of Mycobacterium avium in a model distribution system. Water Res. 2004, 38, 1457–1466. [Google Scholar] [CrossRef] [PubMed]
- Wullings, B.A.; Bakker, G.; van der Kooij, D. Concentration and Diversity of Uncultured Legionella spp. in Two Unchlorinated Drinking Water Supplies with Different Concentrations of Natural Organic Matter. Appl. Environ. Microbiol. 2011, 77, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Van der Wielen, P.W.J.J.; van der Kooij, D. Nontuberculosis Mycobacteria, Fungi, and Opportunistic Pathogens in Unchlorinated Drinking Water in the Netherlands. Appl. Environ. Microbiol. 2013, 79, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.A.; Clarke, D.R. Microbiologically Influenced Corrosion in Hydronic Heating Systems. J. Fail. Anal. Prev. 2004, 4, 38–42. [Google Scholar] [CrossRef]
- Mathys, W.; Stanke, J.; Harmuth, M.; Junge-Mathys, E. Occurrence of Legionella in hot water systems of single-family residences in suburbs of two German cities with special reference to solar and district heating. Int. J. Hyg. Environ. Health 2008, 211, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Van der Kooij, D.; Veenendaal, H.R.; Scheffer, W.J.H. Biofilm formation and multiplication of Legionella in a model warm water system with pipes of copper, stainless steel and cross-linked polyethylene. Water Res. 2005, 39, 2789–2798. [Google Scholar] [CrossRef] [PubMed]
- Filter, U.S. Application Note: Preventing Microbial Contamination in Analytical Grade Water with Regular Sanitation, 2001, Literature No.: LO-Sanit-AN-0201.
- Kayser, W.V.; Hickman, K.C.; Bond, W.W.; Favero, M.S.; Carson, L.A. Bacteriological Evaluation of an Ultra-Pure Water-Distilling System. Appl. Microbiol. 1975, 30, 704–706. [Google Scholar] [PubMed]
- Kulakov, L.A.; McAlister, M.B.; Ogden, K.L.; Larkin, M.J.; O’Hanlon, J.F. Analysis of bacteria contaminating ultrapure water in industrial systems. Appl. Environ. Microbiol. 2002, 68, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- McAlister, M.B.; Kulkaov, L.A.; Larkin, M.J.; Ogden, K.L. Analysis of bacterial contamination in different sections of a high-purity water system. Ultrapure Water 2001, 18, 18–26. [Google Scholar]
- Van der Kooij, D.; Oranje, J.P.; Hijnen, W.A.M. Growth of Pseudomonas aeruginosa in Tap Water in Relation to Utilization of Substrates at Concentrations of a Few Micrograms per Liter. Appl. Environ. Microbiol. 1982, 44, 1086–1095. [Google Scholar] [PubMed]
- Favero, M.S.; Carson, L.A.; Bond, W.W.; Petersen, N.J. Pseudomonas aeruginosa: Growth in distilled water from hospitals. Science 1971, 173, 836–838. [Google Scholar] [CrossRef] [PubMed]
- Van der Wielen, P.W.J.J. Opportunistic pathogens in unchlorinated drinking water in the Netherlands. In Proceedings of the American Water Works Association International Symposium on Waterborne Pathogens, Savannah, GA, USA, 13–14 April 2015.
- Morton, S.C.; Zhang, Y.; Edwards, M. Implications of nutrient release from iron metal for microbial regrowth in water distribution systems. Water Res. 2005, 39, 2883–2892. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Edwards, M. Accelerated chloramine decay and microbial growth by nitrification in premise plumbing. J. Am. Water Works Ass. 2009, 101, 51–62. [Google Scholar]
- Proctor, C. Effect of Various Water Chemistry Factors on Legionella Proliferation and the Premise Plumbing Microbiome Composition. Master’s Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA. USA, 2013. [Google Scholar]
- Temmerman, R.; Vervaeren, H.; Noseda, B.; Boon, N.; Verstraete, W. Necrotrophic Growth of L. pneumophila. Appl. Environ. Microbiol. 2006, 72, 4323–4328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, M.; Benjamin, M.M. Effect of Preozonation on Coagulant-NOM Interactions. J. Am. Water Work Assoc. 1992, 84, 63–72. [Google Scholar]
- Strickhouser, A. Legionella pneumophila in domestic hot water systems: Evaluation of Detection Methods and Environmental Factors Affecting Survival. Master’s Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA. USA, 2007. [Google Scholar]
- Wadowsky, R.M.; Wolford, R.; McNamara, A.M.; Yee, R.B. Effect of temperature, pH, and oxygen level on the multiplication of naturally occurring Legionella pneumophila in potable water. Appl. Environ. Microbiol. 1985, 49, 1197–1205. [Google Scholar] [PubMed]
- Hoffman, P.; Friedman, H.; Bendinelli, M. Legionella Pneumophila: Pathogenesis and Immunity; Springer Science & Business Media, LLC: New York, NY, USA, 2008. [Google Scholar]
- Kuiper, M.W.; Valster, R.M.; Wullings, B.A.; Boonstra, H.; Smidt, H.; van der Kooij, D. Quantitative detection of the free-living Amoeba Hartmannella Vermiformis in surface water by using real-time PCR. Appl. Environ. Microbiol. 2006, 72, 5750–5756. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Edwards, M.A.; Falkinham, J.O., III; Pruden, A. Molecular Survey of Occurrence of Legionella spp., Mycobacterium spp., Pseudomonas aeruginosa and Amoeba Hosts in Two Chloraminated Drinking Water Distribution Systems. Appl. Environ. Microbiol. 2012, 78, 6285–6294. [Google Scholar] [CrossRef] [PubMed]
- Clescerl, L.S.; Greenberg, A.E.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Water Works Association and Water Environment Federation: Washington, DC, USA, 1998. [Google Scholar]
- Servais, P.; Billen, G.; Hascoet, M.C. Determination of the biodegradable fraction of dissolved organic matter in waters. Water Res. 1987, 21, 445–450. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, K.; Pruden, A.; Falkinham, J.O., III; Edwards, M. Relationship between Organic Carbon and Opportunistic Pathogens in Simulated Glass Water Heaters. Pathogens 2015, 4, 355-372. https://doi.org/10.3390/pathogens4020355
Williams K, Pruden A, Falkinham JO III, Edwards M. Relationship between Organic Carbon and Opportunistic Pathogens in Simulated Glass Water Heaters. Pathogens. 2015; 4(2):355-372. https://doi.org/10.3390/pathogens4020355
Chicago/Turabian StyleWilliams, Krista, Amy Pruden, Joseph O. Falkinham, III, and Marc Edwards. 2015. "Relationship between Organic Carbon and Opportunistic Pathogens in Simulated Glass Water Heaters" Pathogens 4, no. 2: 355-372. https://doi.org/10.3390/pathogens4020355