Association and Expression of Virulence from Plasmids of the Group B Strain in Pseudomonas syringae pv. eriobotryae



Abstract
1. Introduction
2. Results
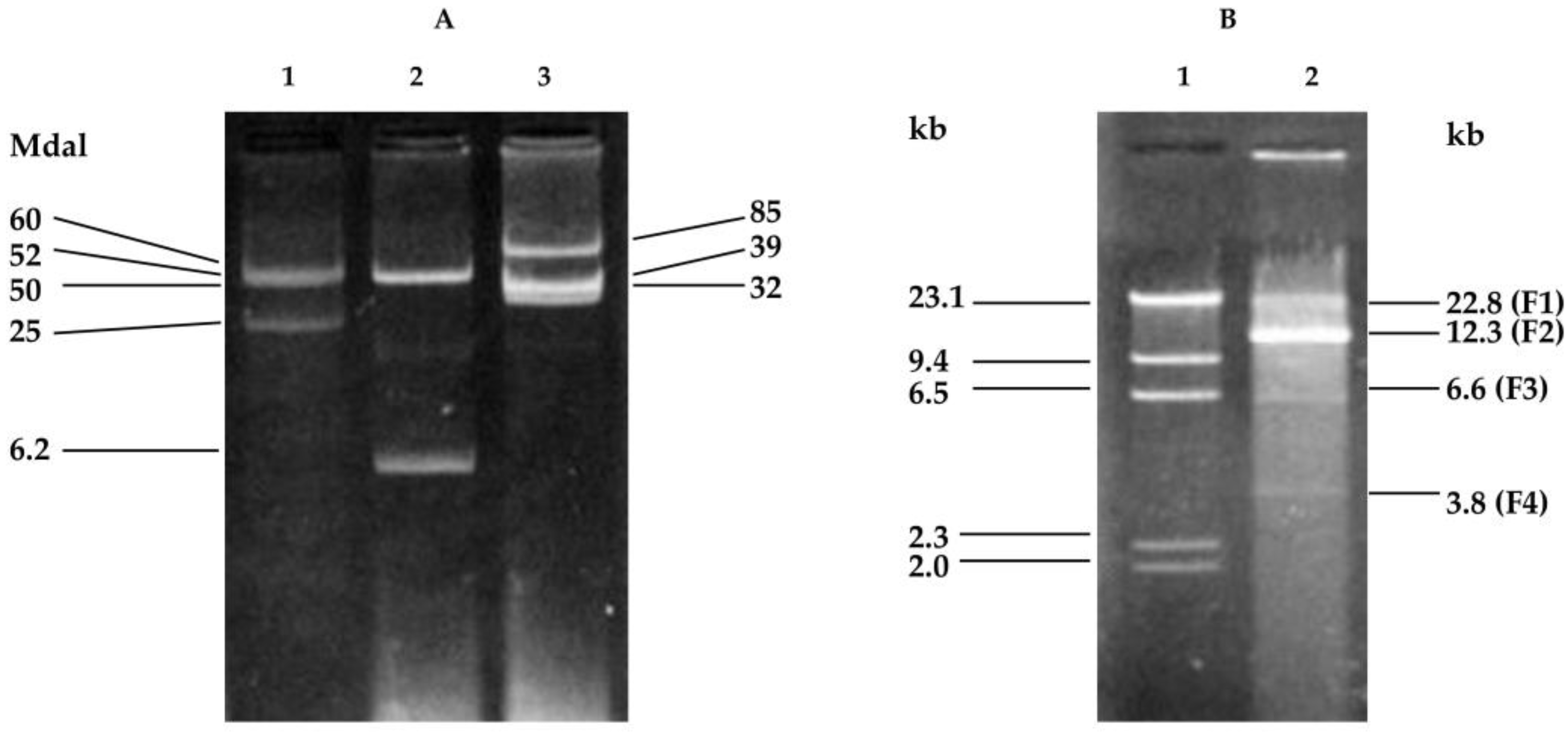
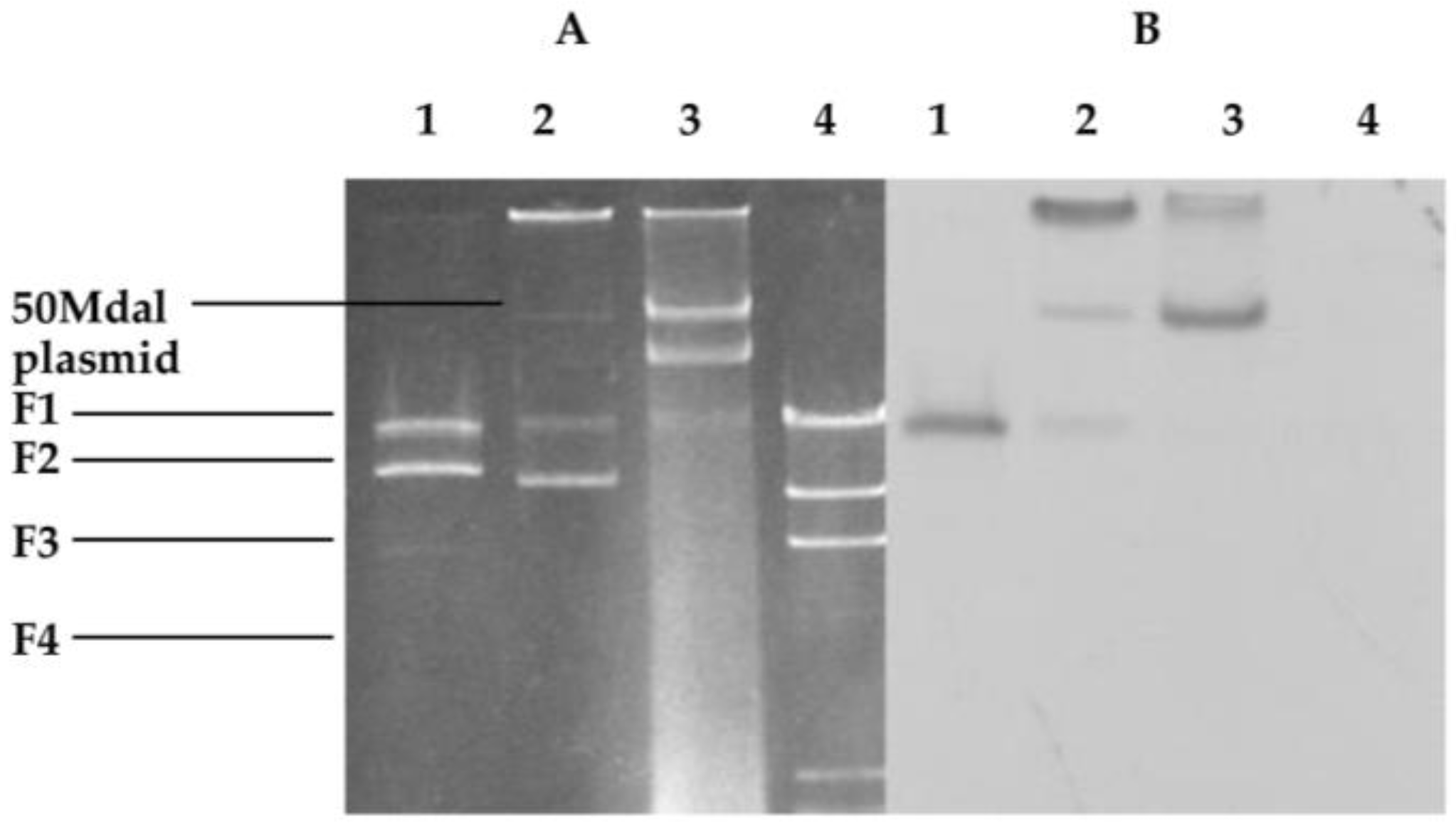
2.1. Cloning Fragments from Plasmid NAE89
2.2. Pathogenic Experiment
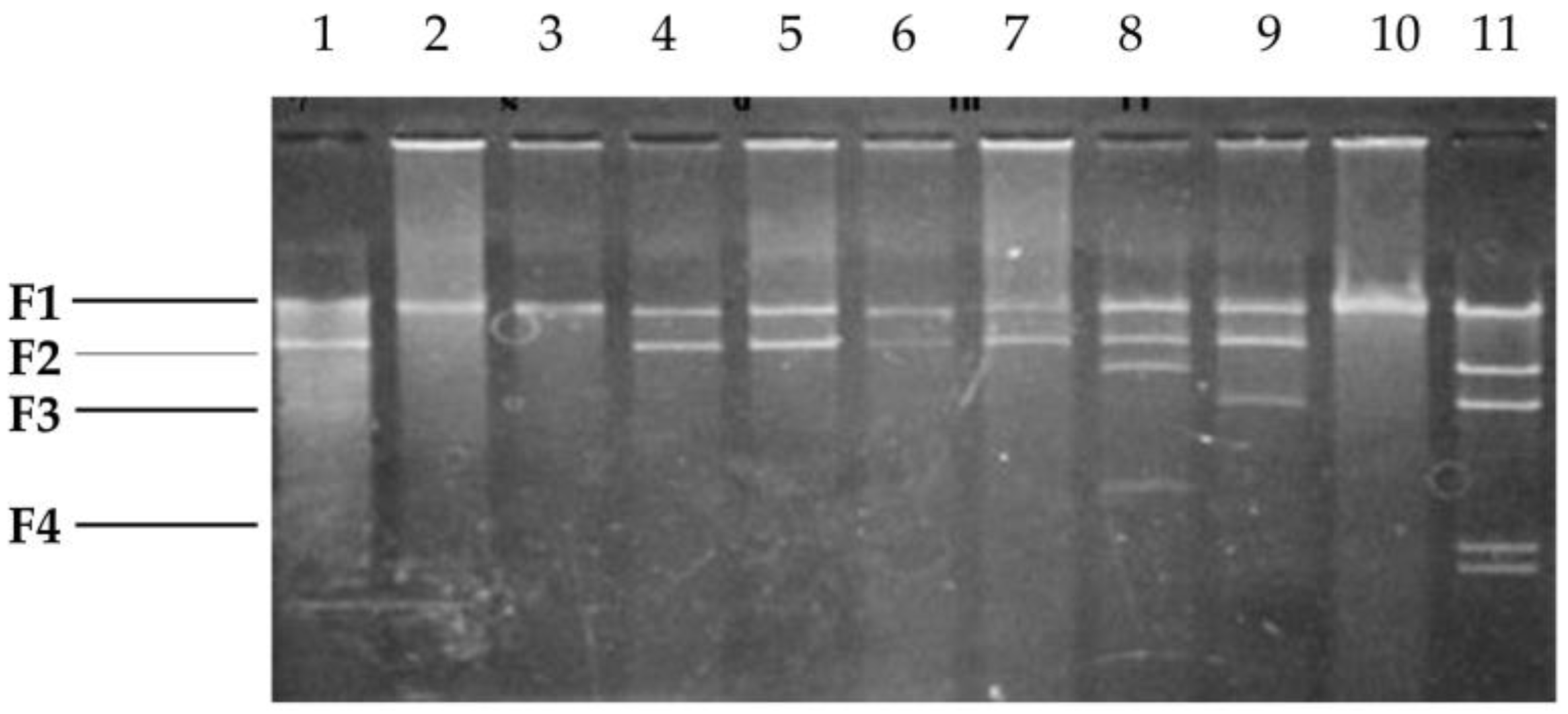
2.3. Southern Hybridization
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, Culture Conditions, and Chemicals
4.2. Plant Inoculation
4.3. General DNA Manipulation and Clone Plasmid
4.4. Southern Hybridization Experiments
Supplementary Materials
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Morita, A. Effect of the inoculation of Pseudomonas syringae pv. eriobotryae at seedling stage upon its growth and fruit-productivity. Ann. Phytopathol. Soc. Jpn. 1991, 57, 629–633. [Google Scholar]
- Fukuda, S.; Ishimoto, K.; Terakami, S.; Yamamoto, T.; Hiehata, N. Genetic mapping of the loquat canker resistance gene pse-c in loquat (Eriobotrya japonica). Sci. Hort. 2016, 200, 19–24. [Google Scholar] [CrossRef]
- Alippi, A.M.; Alippi, H.E. Stem canker of loquat: A new disease in Argentina. Rev. Argent. Microbiol. 1990, 22, 155–158. [Google Scholar] [PubMed]
- Lai, M.; McCartney, W.O.; Morin, C.W. Canker of loquat caused by Pseudomonas sp. Phytopathology 1971, 61, 248–249. [Google Scholar] [CrossRef]
- Lai, M.; Morin, C.W.; Weigle, C.G. Two strains of Pseudomonas eriobotryae isolated for loquat cankers in California. Phytopathology 1972, 62, 310–313. [Google Scholar] [CrossRef]
- Lin, S.; Sharpe, R.H.; Janick, J. Loquat: Botany and Horticulture. Hort. Rev. 1999, 23, 233–276. [Google Scholar]
- McRae, E.M.; Hale, C.N. New plant disease record in New Zealand: Loquat canker. N. Z. J. Agric. Res. 1986, 29, 547–550. [Google Scholar] [CrossRef]
- Mukoo, H. Studies on the causal bacteria of the loquat canker (1). Bull. Natl. Inst. Agric. Sci. 1952, C-1, 1–190. [Google Scholar]
- Wimalajeewa, D.L.S.; Pascoe, I.G.; Jones, D.L. Bacterial stem canker of loquat. Australas. Plant Pathol. 1978, 7. [Google Scholar] [CrossRef]
- Morita, A. Studies on the loquat canker caused by Pseudomonas eriobotryae (Takimoto) Dowson. II: Grouping of the bacterial isolates on the basis of their pigment producibility and pathogenicity. Ann. Phytopathol. Soc. Jpn. 1978, 44, 6–13. [Google Scholar] [CrossRef]
- Morita, A. Studies on physiology and ecology of loquat canker. J. Gen. Plant Pathol. 2001, 67, 244–247. [Google Scholar] [CrossRef]
- Fukuta, S.; Ishimoto, K.; Sato, S.; Terakami, S.; Yamamoto, T.; Hiehata, N. Genetic mapping of the loquat canker resistance locus in bronze loquat (Eriobotrya deflexa). Tree Genet. Genomes 2014, 10, 875–883. [Google Scholar] [CrossRef]
- Kamiunten, H. Involvement of a plasmid in the expression of virulence in Pseudomonas syringae pv. eriobotryae. Ann. Phytopathol. Soc. Jpn. 1995, 61, 376–380. [Google Scholar] [CrossRef]
- Jackson, R.W.; Athanassopoulos, E.; Tsiamis, G.; Mansfield, W.J.; Arnold, L.D.; Gibbon, J.M.; Murillo, J.; Taylor, D.J.; Vivian, A. Identification of a pathogenicity island, which contains genes for virulence and a virulence, on a large native plasmid in the bean pathogen Pseudomonas syringae pathovar phaseolicola. Proc. Natl. Acad. Sci. USA 1999, 96, 10875–10880. [Google Scholar] [CrossRef] [PubMed]
- Hiehata, N.; Sato, Y.; Fukuda, S.; Terai, O. Inheritance of resistance to loquat canker (Pseudomonas syringae pv. eriobotryae, Group A) in loquat (Eriobotrya japonica). J. Jpn. Soc. Hort. Sci. 2002, 71, 255–261. [Google Scholar] [CrossRef]
- Kamiunten, H. Isolation and characterization of virulence gene psvA on a plasmid of Pseudomonas syringae pv. eriobotryae. Ann. Phytopathol. Soc. Jpn. 1999, 65, 501–509. [Google Scholar] [CrossRef][Green Version]
- Alan, V.; Jesús, M.; Robert, W.J. The roles of plasmids in phytopathogenic bacteria: mobile arsenal? Microbiology 2001, 147, 763–780. [Google Scholar]
- Gonzalez, C.F.; Pettit, E.A.; Valadez, V.A.; Provin, E.M. Mobilization, cloning and sequence determination of plasmid encoded polygalacturonase from a phytopathogenic Burkholderia Pseudomonas cepacia. Mol. Plant Microbe Interact. 1997, 10, 840–851. [Google Scholar] [CrossRef] [PubMed]
- Ezra, D.; Barash, I.; Valinsky, L.; Manulis, S. The dual function in virulence and host-range restriction of a gene isolated from the pPATHEhg plasmid of Erwinia herbicola pv. gypsophilae. Mol. Plant Microbe Interact. 2000, 13, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Hiehata, N.; Sato, Y.; Fukuda, S.; Tominaga, Y.; Terai, O.; Nesumi, H. Inheritance of resistance to loquat canker (Group C) in progenies derived from ‘Shiromogi’ loquat. J. Am. Soc. Hort. Sci. 2012, 137, 152–156. [Google Scholar]
- Hiehata, N.; Fukuda, S.; Terai, O. Evaluation of loquat canker resistance in new loquat cultivars released in Japan and introduced from foreign countries. Bull. Nagasaki. Fruit Tree Exp. Stat. 2007, 10, 14–21. [Google Scholar]
- Kado, C.I.; Liu, S.T. Rapid procedure for detection and isolation of large and small plasmids. J. Bacteriol. 1981, 145, 1365–1373. [Google Scholar] [PubMed]
- Kamiunten, H. Loss of a plasmid in Pseudomonas syringae pv. eriobotryae is correlated with a change of symptoms. Ann. Phytopathol. Soc. Jpn. 1990, 56, 645–650. [Google Scholar] [CrossRef]
- Hasebe, A.; Tsushima, S.; Lida, S. Isolation and characterization of IS1416 from Pseudomonas glumae, a new member of the IS3 family. Plasmid 1998, 39, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Perbal, B. A Practical Guide to Molecular Cloning, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1988; pp. 396–399. [Google Scholar]






{kind=link}
{kind=link}
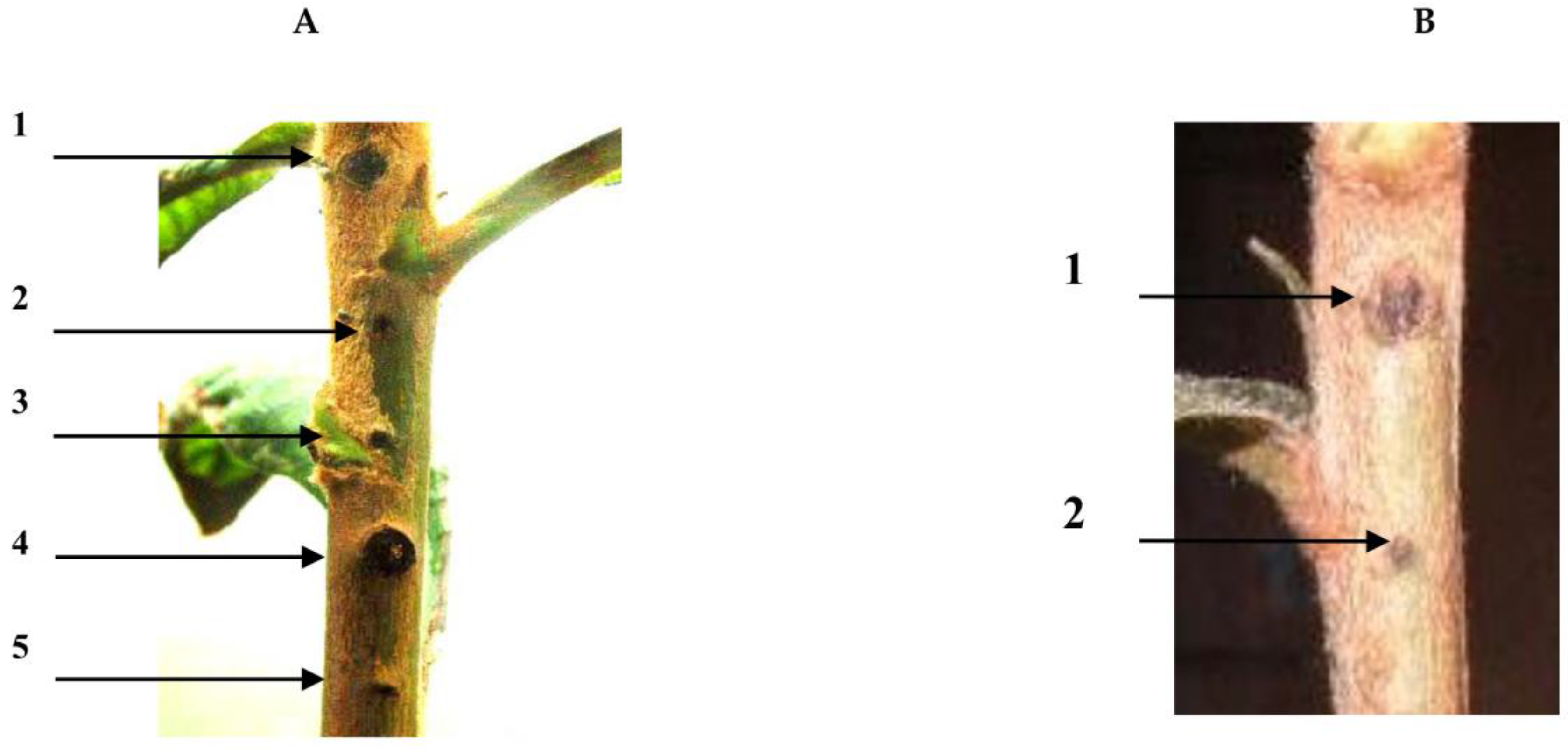{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids | Relevant Properties | Source or Reference |
|---|---|---|
| Pseudomonas syringae pv. eriobotryae | Nagasaki Fruit-Tree Experiment Station, Japan | |
| NAE6 | Wild-type virulence in group A | [13] |
| PE0 | Plasmid lost NAE 6 | [13] |
| NAE89 | Wild-type virulence in group B | Nippongene |
| NAE89-1 | The variant of lost halo gene formation | Nippongene |
| NAE57 | Wild-type in the C group | Nippongene |
| E. coli DH5 | E. coli bacteria and bacterium strain | Nippongene |
| pET-3a (psvA) | Vector inserted with BamHI to psvA used as a probe in this study, isolated from a bacteria lesion of the strain group A | Stratagene |
| pRK2013 | Helper plasmid | This study |
| pLAFR3 | Cosmid vector | This study |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khanh, T.D.; Xuan, T.D. Association and Expression of Virulence from Plasmids of the Group B Strain in Pseudomonas syringae pv. eriobotryae. Pathogens 2018, 7, 41. https://doi.org/10.3390/pathogens7020041
Khanh TD, Xuan TD. Association and Expression of Virulence from Plasmids of the Group B Strain in Pseudomonas syringae pv. eriobotryae. Pathogens. 2018; 7(2):41. https://doi.org/10.3390/pathogens7020041
Chicago/Turabian StyleKhanh, Tran Dang, and Tran Dang Xuan. 2018. "Association and Expression of Virulence from Plasmids of the Group B Strain in Pseudomonas syringae pv. eriobotryae" Pathogens 7, no. 2: 41. https://doi.org/10.3390/pathogens7020041
APA StyleKhanh, T. D., & Xuan, T. D. (2018). Association and Expression of Virulence from Plasmids of the Group B Strain in Pseudomonas syringae pv. eriobotryae. Pathogens, 7(2), 41. https://doi.org/10.3390/pathogens7020041