
Myo-D-inositol Trisphosphate Signalling in Oomycetes
by

Indu Muraleedharan Nair
1,2,
Emma Condon
1,2,
Barbara Doyle Prestwich
2 and
John James Mackrill
1,*
1
Department of Physiology, School of Medicine, University College Cork (UCC), T12 YT20 Cork, Ireland
2
Department of Plant Science, School of Biological, Earth and Environmental Sciences, University College Cork (UCC), T23 TK30 Cork, Ireland
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(11), 2157; https://doi.org/10.3390/microorganisms10112157
Submission received: 4 October 2022
/
Revised: 21 October 2022
/
Accepted: 27 October 2022
/
Published: 31 October 2022
(This article belongs to the Special Issue Feature Collection in Environmental Microbiology Section 2021-2022)
Abstract
:Oomycetes are pathogens of plants and animals, which cause billions of dollars of global losses to the agriculture, aquaculture and forestry sectors each year. These organisms superficially resemble fungi, with an archetype being Phytophthora infestans, the cause of late blight of tomatoes and potatoes. Comparison of the physiology of oomycetes with that of other organisms, such as plants and animals, may provide new routes to selectively combat these pathogens. In most eukaryotes, myo-inositol 1,4,5 trisphosphate is a key second messenger that links extracellular stimuli to increases in cytoplasmic Ca2+, to regulate cellular activities. In the work presented in this study, investigation of the molecular components of myo-inositol 1,4,5 trisphosphate signaling in oomycetes has unveiled similarities and differences with that in other eukaryotes. Most striking is that several oomycete species lack detectable phosphoinositide-selective phospholipase C homologues, the enzyme family that generates this second messenger, but still possess relatives of myo-inositol 1,4,5 trisphosphate-gated Ca2+-channels.
1. Introduction
Oomycetes are important pathogens worldwide, causing major losses globally [1]. However, they also play a role in nutrient cycling, and have the potential to produce useful chemicals [2,3]. They are generally found in damp terrestrial and freshwater environments, but have also been detected in arid regions such as deserts [4].
Ca2+ is a key second messenger utilized by all domains of life. Due to its high abundance in nature and potential cytotoxic effects, cells have evolved to maintain unstimulated cytoplasmic Ca2+ concentrations at sub-micromolar concentrations, compared to low millimolar extracellular levels [5]. Changes in cytoplasmic Ca2+ concentrations, due to the concerted actions of channels, transporters and buffers, control a wide array of cellular processes [6,7]. High cytoplasmic levels of Ca2+ can be lethal to cells [8]. Therefore, to avoid excessive Ca2+ accumulation, organisms have evolved mechanisms to expel excess Ca2+ and to store it in organelles such as the endoplasmic reticulum, vacuoles, mitochondria and the endolysosomal system [5,7,9,10]. In turn, this has facilitated the use of Ca2+ as a second messenger to regulate cell biology, by Ca2+ fluxes across the membranes of the cell-surface and intracellular stores.
Ca2+ release from intracellular stores can be rapidly triggered by a wide array of stimuli, allowing cells to produce fast responses [11]. Ca2+ channels located in the cell-surface membrane can be triggered to open by an array of stimuli, including mechanical forces, neurotransmitters, hormones, nutrients, membrane depolarization, or changes in temperature, allowing extracellular Ca2+ to flow into the cell [6,12]. However, for extracellular cues to activate Ca2+ channels in the membranes of intracellular Ca2+ stores, second messengers must be employed. There are several second messengers that elicit Ca2+ release from these intracellular stores, including myo-D-inositol 1,4,5-trisphosphate (IP3), cyclic ADP ribose, and nicotinic acid dinucleotide phosphate (NAADP) [7].
As the endoplasmic reticulum is a major Ca2+ store within most eukaryotic cells, its membrane is an important site for the action of Ca2+ release channels, including the IP3 receptors (IP3Rs) and ryanodine receptors (RyRs) [13]. In addition to the endoplasmic reticulum, plants and fungi also accumulate Ca2+ in vacuoles and other organelles. Paradoxically, structural homologues of mammalian IP3Rs have not been detected in the genomes of late-branching plant or late-branching fungal taxa [14]. However, they are encoded by the genomes of early-branching groups such as Volvox spp. [15] and Mucor spp [16]. In addition, whilst concentrated in the endoplasmic reticulum, IP3Rs and RyRs have also been reported to be present in the cell surface membrane, the sarcoplasmic reticulum, mitochondria, the endolysosomal system, the Golgi and the nuclear envelope [6,12].
In animal cells, depletion of intracellular Ca2+ stores, which can occur as a consequence of opening of IP3Rs, causes conformational changes in stromal interaction molecules (STIM): proteins that sense endoplasmic reticulum intraluminal Ca2+ concentrations. On sensing Ca2+-depleted stores, STIM can interact with Orai Ca2+ channels in the cell-surface membrane, thereby facilitating store-operated Ca2+ entry [17]. Another way by which Ca2+ can enter eukaryotic cells is via Na+/Ca2+-exchangers (NCX). These are surface membrane transporters that use the entry of three Na+ to expel one Ca2+. Under certain conditions, such as high intracellular Na+ or in depolarized cells, NCX can operate in reverse mode, driving Ca2+-entry [18]. Elevation of cytoplasmic Ca2+ via by any mechanism (entry or release) can be amplified by RyRs- or IP3R-dependent Ca2+-release, in a process termed Ca2+-induced Ca2+-release (CICR) [11].
Ca2+ plays a key role in controlling oomycete biology [1,9]. Many eukaryotic Ca2+ signalling components are conserved in oomycetes; however, several channels and receptor families are either apparently absent or possess distinctive structures. For example, homologues of Orai and STIM, essential components of store-operated Ca2+-entry in other eukaryotes, could not be detected in the oomycetes Phytophthora infestans and Saprolegnia declina. However, these oomycetes possess homologues of mammalian IP3Rs, whose opening would lead to Ca2+-store depletion [9,19].
G-protein-coupled receptors (GPCRs) and receptor tyrosine tyrosine kinases (RTK) are located in the surface membrane where they can initiate Ca2+ signalling in response to stimuli, such as nutrients, hormones and growth factors. Once activated, these receptors couple to enhanced phosphoinositide-selective phospholipase C (PI-PLC) activity. Such GPCRs are associated with heterotrimeric G-protein complexes, consisting of α, β and γ subunits. On ligand-receptor binding, the subunits dissociate and can then interact with transducer proteins. G-protein subunits of the αq or α11 subtypes can stimulate certain PI-PLC isozymes (PI-PLC β1–β3 in mammals) [20]. Ligand-activated RTKs have intrinsic kinase activity and initiate signalling by phosphorylating distinct PI-PLC isozymes (γ1 and γ2) at key tyrosine residues. Other PI-PLC families (δ, ε, η, ζ) are activated by various stimuli, including heterotrimeric G-protein βγ subunits (for η), monomeric G-proteins (such as Ras and Rho, for ε) and Ca2+ itself (for δ, ε, η, ζ) [21]. Active PI-PLCs hydrolyse a minor membrane phospholipid, phosphatidylinositol 4,5–biphosphate (PIP2), to generate the second messengers 1,2-diacylglycerol (DAG) and IP3, Figure 1. DAG is membrane associated lipid, which exerts its effects by stimulation of protein kinase C isozymes which phosphorylate target proteins, modifying their function, thereby regulating cellular activities [22].
In contrast to DAG, IP3 is a soluble second messenger which diffuses through the cytoplasm, to interact with IP3 receptors (IP3R) located in the membranes of intracellular Ca2+ stores, such as the endoplasmic reticulum. Binding of four molecules of IP3 per tetrameric IP3R promotes a channel-open conformational state of this complex, allowing Ca2+ release [23]. In addition, both IP3Rs and RyRs display biphasic relationships between channel gating and both cytoplasmic and intraluminal Ca2+ concentrations [11]. Preliminary analyses of oomycete conceptual proteomes indicates that IP3 signalling in oomycetes is distinct from that in other eukaryotes, such as plants and animals [4,9,19,24]. For example, the only candidate proteins in oomycetes sharing high identity with RyRs do so by homology with two “RyR-(R) domains”. These homologues are more closely related to the polycystic kidney disease (PKD) member of the transient receptor potential channel superfamily than they are to RyRs. The presence of these oomycete-specific PKDRR proteins in P. infestans has been verified using Western immunoblotting and immunofluorescent microscopy approaches [19]. Such differences between signalling molecules may provide a route for development of new strategies for combatting oomycete pathogens and consequently, are the focus of this review.
2. Oomycetes
Due to their similar morphologies, growth patterns and life cycles, oomycetes were long classified amongst fungi [4,25,26,27]. However, despite their superficial similarities, the ancestors of oomycetes and fungi diverged a long time ago and are found within different branches of the eukaryotic phylogenetic tree. From molecular clock models, oomycetes are estimated to have diverged from their last common ancestor with other eukaryotes about 430 to 400 million years ago. The two major orders, Peronosporales and Saprolegnialea, are estimated to have diverged from each other 225 to 190 million years ago [28]. Taxonomically placed into the Straminipilia kingdom, oomycetes are closely related to diatoms and brown algae [4]. As such, they are thought to have originated in marine environments, moving into freshwater and terrestrial environments as their hosts evolved [29].
Oomycete genomes (50–250 Mb) are much larger than those of fungi (10–40 Mb) [1]. Another main distinction between oomycetes and fungi are the components of their cell walls. In fungi, chitin is the main component, whereas in oomycetes there is very limited chitin, with the walls being made of mostly cellulose and other β-glucans [27,30].
Due to their evolutionary distance from ‘true fungi’, oomycetes utilize distinct genetic and biochemical mechanisms to infect their hosts. Genome sequences of important oomycete pathogens have also highlighted biochemical disparities between oomycetes and other eukaryotes [3,24]. Horizontal gene transfer between species and ‘host jumping’ are also thought to play a role in oomycete biochemical pathways, influencing pathogenicity, signal transduction and motility [2,4]. Within oomycetes, there are a wide variety of lifecycles, from biotrophy (obtaining nutrients from living hosts) to necrotrophy (obtaining them from dead tissues). Many pathogenic oomycetes, including Phytophthora spp., are hemi-biotrophic (switching from biotrophy to necrotrophy during disease progression) [2,4].
Oomycetes are the cause of many devastating plant and animal diseases globally, often leading to major losses of crops, trees and fish-stocks [26,31]. One example is Phytophthora infestans, which causes late blight in tomato and potato crops. This pathogen is well known as the agent that caused widespread devastation of the potato crop in Ireland and across Europe in the mid-19th century leading to the Great Irish Famine [29]. Oomycete pathogens, are notoriously difficult to control and can affect a wide range of hosts and ecosystems. Economic losses are not only incurred by crop failure, but also those involved in the application of pesticides against oomycetes. For P. infestans alone, global costs are estimated at about 10 billion US dollars per year [1]. Many anti-oomycete pesticides are becoming increasingly ineffective, due to development of resistance in oomycete populations, or the banning of these chemicals due to environmental and health concerns [32].
One of the most important factors in oomycete pathogenicity is the ability to rapidly produce zoospores, generated within structures called sporangia. Zoospores are released from sporangia during cold and wet conditions. These mobile spores then locate a new host, encyst, and germinate to infect the host via haustoria, instigating the infection cycle once again [27]. Ca2+ signalling plays an important role in regulating this aspect of oomycete infection. Therefore, understanding the underlying Ca2+ signalling mechanisms at play could allow for the infection to be targeted without harming the host [9].
However, despite the importance of these pathogens, knowledge pertaining to the mechanisms involved in controlling their cell biology is lacking. Understanding their unique signalling mechanisms may provide insight into new control methods [3,19]. As a result of their distinct biochemistry, classical fungicides have long proven ineffective against oomycetes. For example, oomycetes do not synthesise ergosterol making them resistant to the effects of azole fungicides [33]. Another example, Metalaxyl, interferes with RNA polymerases and has proven to be highly effective against oomycetes. This has a wide spectrum of activity, however, resistance in P. infestans was reported within a year of its initial use and is now widespread in certain subpopulations of this pathogen [34]. Thus, finding new ways to target these pathogens is imperative. A thorough understanding of oomycete physiology and infection mechanisms could open up a wide variety of possible control strategies using both chemical and biocontrol approaches.
3. Ca2+ and Phosphoinositide Signalling in Oomycetes
3.1. Oomycete Ca2+ Signalling
Like other eukaryotes, a large electrochemical gradient for Ca2+ exists across the cell membrane of oomycetes, with cytoplasmic levels of about 100 nM in Phytophthora cinnamomi [35], relative to the high micromolar to low millimolar concentrations in the extracellular environment. Changes in cytoplasmic Ca2+ concentration have been reported to regulate multiple aspects of oomycete biology [9]. For example, stretch-activated Ca2+ channels towards the hyphal tips in the oomycete Saprolegnia ferax act as a source of Ca2+ for sensing hyphal expansion [36]. Cold-shock is a stimulus that triggers zoosporogenesis and zoospore release in many Phytophthora spp. [1,37]. Cytokinesis and zoospore formation in P. cinnamomi sporangia is triggered by a cold stimulus is associated with a rise in cytoplasmic Ca2+ [36]. Oomycete zoospores migrate towards favourable, or away from unfavourable, chemical stimuli in a process termed chemotaxis. Silencing of the Gα subunit-encoding gene (potentially involved IP3 -mediated Ca2+ signalling) in P. sojae altered the chemotaxis of zoospores towards the soybean isoflavanone, daidzein [38]. The motility of Pythium porphyrae (cause of root-rot of the seaweed Porphyra yezoensis) zoospores is inhibited by extracellular Ca2+, whereas germination of its cysts is blocked by ethylene glycol-bis(2-aminoethylether)-N, N, N, N -tetraacetic acid (EGTA), a Ca2+ chelator [39].
3.2. Phosphoinositide and Myo-D-inositol 1,4,5-Trisphosphate (IP3) Signalling
IP3 is a widely-utilized second messenger found in most eukaryote systems [40]. IP3Rs and RyRs form a superfamily of tetrameric channels that release Ca2+ from intracellular stores [6,16]. The binding of signalling molecules (IP3, Ca2+, cADPR, possibly NAADP) or allosteric interactions with other proteins can activate these channels allowing release of Ca2+ [7].
In oomycetes, putative IP3-mediated Ca2+ signalling has been well studied in relation to zoosporogenesis [41]. The production and encystment of zoospores plays a key role in the pathogenicity of many oomycetes [25]. A short period of cold shock is required for zoospores to be released from sporangia. This cold shock reduces fluidity of the cell surface membrane, resulting in stimulation of membrane bound sensors such as mechano-sensitive cation channels and GPCRs [1,37]. In other eukaryotes, activation of GPCRs can initiate the PLC–IP3-Ca2+ release mechanism, whereas mechano-channel activation leads to Ca2+ -influx that can be amplified by CICR, with either mechanism potentially resulting in Ca2+ release from the endoplasmic reticulum or vacuoles via IP3Rs. Despite a clear role for Ca2+ in triggering zoospore encystment, PI-PLC homologues have yet to be detected in Phytophthora spp. [24]. However, pharmacological studies suggest that PLC–IP3-dependent signalling plays a role in P. infestans zoosporogenesis. Both the PLC-inhibitor U73122 and the IP3-antagonist 2-aminoethoxydiphenyl borate (2-APB) blocked zoosporogenesis in P. infestans [41]. Of a family of seven nuclear LIM interactor-interacting transcription factors, four members were upregulated in zoospores, with this expression being inhibited by either U73122 or 2-APB [42]. The properties of some of the chemicals used to study oomycete PI signalling are summarized in Table 1. A key point to note is that most of these small molecules exert effects on multiple targets in mammalian cells, bringing their specificity of effect in oomycetes into doubt.
In the current study, the presence of homologues of Homo sapiens PI signalling proteins was investigated in the oomycete orders Peronosporales, Saprolegniales and Albuginales. Animals, fungi, and plants were also examined, for comparative purposes. Query sequences were: Gαq, PI-PLCs (α, β, δ, ε, η and ζ), IP3R channels (ITPR), IP kinases (including ITPK1, IPPK, ITPKA and IPMK) and IP phosphatases (including IMPA1, INPP1, INPP4, INPP5 and MINPP1). Findings from this study are summarized in Figure 2 and are represented in full detail in Supplemental Table S1.
Homologues of Homo sapiens IP signalling proteins were investigated in Caenorhabditis elegans (Metazoa, Nematoda), Neurospora crassa (Fungi, Sordariales), Arabidopsis thaliana (Embryophyta, Bassicales), Phytophthora infestans (Oomycota, Peronosporales), Plasmopara halstedii (Oomycota, Peronosporales), Pythium oligandrum (Oomycota, Peronosporales), Saprolegnia parasitica (Oomycota, Saprolegniales), Pythium oligandrum (Oomycota, Saprolegniales) and Albugo laibaichii (Oomycota, Albuginales), using the Protein BLAST tool at NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins). The query genes were H. sapiens GNAQ, PI-PLCs (only δ1 shown), ITPR, IP-kinases (ITPK1, IPPK, ITPKA, IPMK), and IP-phosphatases (IMPA1, INPP1, INPP4, INPP5 and MINPP1). The look-up-table (LUT, upper left) indicate estimates of the probability that homology between sequences occurred by chance. The cut-off for this value used is 1 x10−4. ‘ND’ indicates no detectable homology. ‘C2’ indicates limited homology, due to a common C2 domain. Further details are available in Supplementary Table S1.
3.3. Oomycete GPCRs, G-Proteins and RTKs
The genomic sequences of P. sojae and P. ramorum indicate the presence of a single Gα and a single Gβ subunit in Phytophthora spp. [44]. However, there are 24 hypothetical GPCR proteins in these species, 12 of which are distinct from the GPCRs other eukaryotes in that they contain a C-terminal phosphophatidylinositol phosphate kinase (PIPK) domain. In P. infestans, silencing of one gene encoding a GPCR-PIPK (termed GK4) inhibited sporangial development and virulence [45]. In P. sojae, silencing of either GK4 or GK5 impaired pathogenicity, but only deletion of GK4 promoted encystment and reduced cyst germination [46]. Consequently, these GPCR-PIPKs have important roles in oomycete biology and might represent a means of directly coupling extracellular signals to increased phosphorylation of PI lipids, bypassingPLC activation [47].
In P. sojae, a putative GPCR called GPR11 is upregulated in zoospores. Silencing of this gene impaired zoospore release, encystment and germination, as well as pathogenicity. However, interaction between GPR11 and Gα could not be demonstrated using the yeast two-hybrid complementation system, nor did silencing of these genes have related effects on the expression of specific target genes [48].
G-protein mediated Ca2+ signalling potentially plays an important role in zoosporogenesis and cyst formation in P. infestans, since Gα (pigpa1) and Gβ (pigpb1) genes are upregulated during these developmental stages [49]. Silencing of the sole Gα subunit in P. infestans lead to aberrant swimming patterns in zoospores [43]. Similarly, in P. sojae, the Gα subunit is essential for chemotaxis towards soybean isoflavones [38]. It also plays a role in locating sites for penetration of host tissues and forms part of a signalling pathway with a regulator of G-protein signalling (RGS) proteins, a GTP-hydrolysing protein that can inactivate the Gα subunit [50]. In contrast, silencing of the Gβ subunit in P. infestans inhibited vegetative growth and formation of sporangia [51]. These experiments indicate that in oomycetes, different subunits of heterotrimeric G-proteins initiate separate signalling events, with distinct cellular outcomes. There is no evidence that homologues of mammalian heterotrimeric G-protein γ-subunits are encoded within the genomes of oomycetes [24,44].
Mastoparan is a peptide obtained from the venom of the wasp Vespula lewisii. In addition to membranolytic properties, mastoparan is a direct activator of heterotrimeric G-proteins, which mimics activated GPCRs [52]. In many eukaryotic cells, including those from animals and plants, application of mastoparan promotes PIP2 hydrolysis, accumulation of IP3 and DAG [53,54]. In contrast, mastoparan appears to stimulate phospholipase D (PLD) in P. infestans, leading to an increase in phosphatidic acid (PA) levels, with no evidence of increased PIP2 hydrolysis [21,43,49,55]. This suggests that the signalling mechanisms elicited by mastoparan in P. infestans are distinctive from those in other eukaryotes.
In metazoans (animals), the IP3 signalling pathway can also be activated through RTKs, which phosphorylate members of the PLCγ family to drive PIP2 hydrolysis. Although candidate tyrosine kinases have been identified in the genome of P. infestans [56], their relationships to metazoan RTKs and potential roles in Ca2+ signalling have not been established to date.
3.4. Oomycete Phosphoinositide-Selective Phospholipase C (PI-PLC) Isozymes
PI-PLCs hydrolyse the membrane phospholipid PIP2 to generate IP3 and DAG. IP3 can be inactivated (in terms of its ability to effectively gate IP3Rs) by progressive phosphorylation to generate higher order inositol polyphosphates. It can also be dephosphorylated to produce inositol, which can be used to regenerate PIP2 [57,58,59].
Substantial quantities of the PI-PLC substrate PIP2 could not be detected in P. infestans, preventing further analysis of its biological roles [55]. Furthermore, genomewide analysis of P. infestans failed to identify any PI-PLC homologues. However, PLD, DAG kinase, and phosphatidylinositol phosphate kinase (PIPK) encoding genes were detected in this oomycete [24]. In contrast, PIP2 has been unequivocally detected in the hyphae of P. cinnamomi, using liquid chromatography-mass spectroscopy approaches [60]. Chilling-induced zoosporogenesis of P. infestans (a Ca2+-dependent process) did not involve convincing IP3 accumulation [61]. In contrast, substantial IP3 accumulation was detected during pectin-stimulation of P. palmivora, with this plant-derived stimulus promoting differentiation of zoospores into cysts. However, the change in IP3 concentration was not rapid enough, nor was of large enough magnitude, to be the driver of zoospore differentiation. Furthermore, it has been estimated that the unstimulated cytosolic concentration of IP3 in P. palmivora zoospores was in the order of 5 μM [62], about two orders of magnitude greater than that in metazoan cells [63].
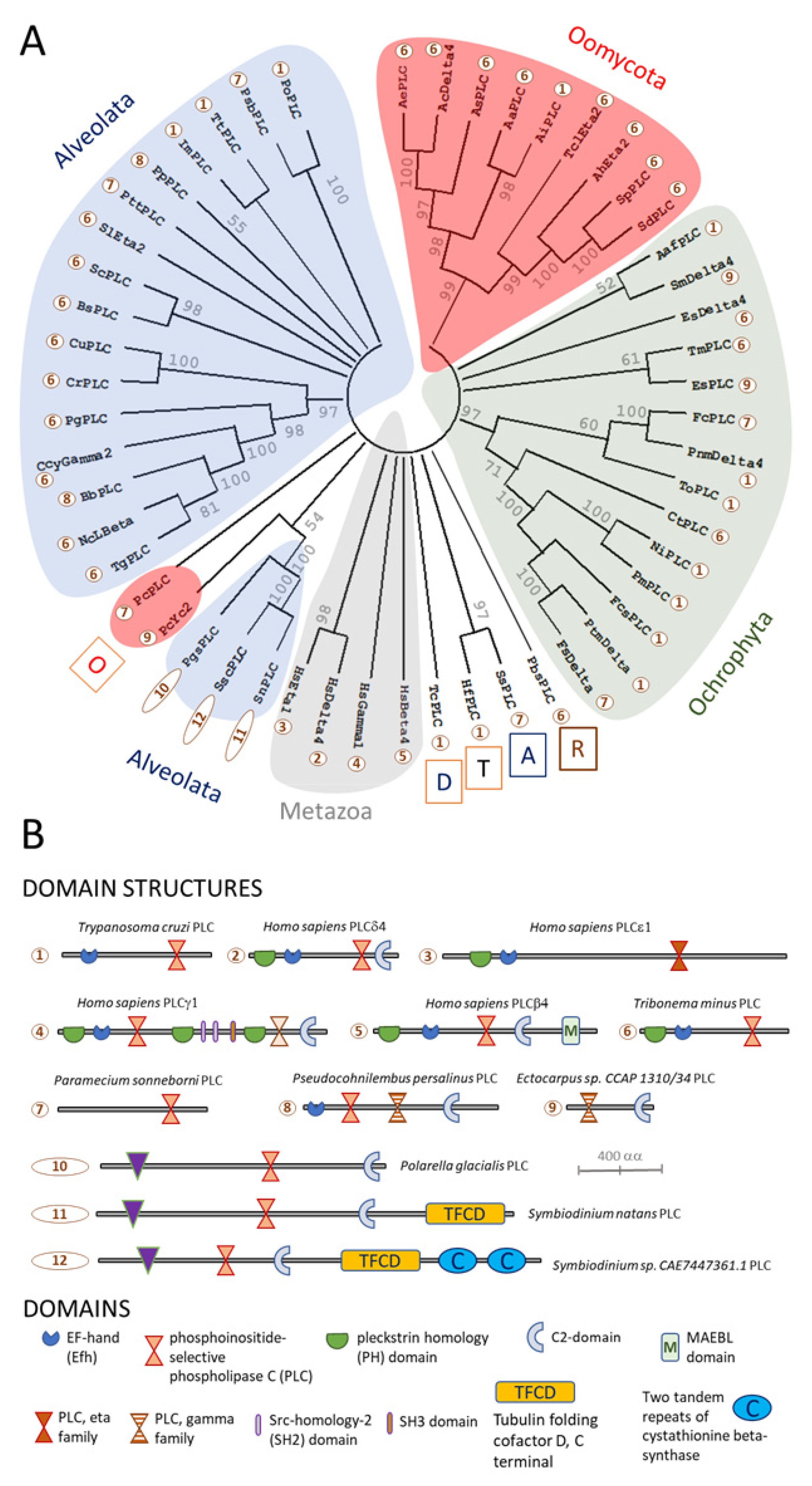
In the current study, searching for homologues of human PI-PLC proteins encoded by genomes from the Stramenopile-Alveolate-Rhizaria (SAR) group failed to detect these enzymes in P. infestans, Figure 2 and Figure 3. However, other oomycetes possess unambiguous homologues of PI-PLCs. These generally have a predicted protein domain structure consisting of a PLC-catalytic domain close to the C-terminus, in combination with an EF-hand Ca2+-binding site closer to the N-terminus, Figure 3 and Supplementary Table S2. Other oomycete PI-PLCs contain an additional candidate pleckstrin homology (PH) domain, N-terminal to the EF-hand domain. These predicted protein architectures are reminiscent of mammalian PI-PLCε isozymes, which are activated by increased cytoplasmic Ca2+ concentrations, suggesting that oomycete PI-PLCs of this type may serve to amplify transient Ca2+ signals [64]. Two distinctive types of oomycete PI-PLCs form phylogenetically distinct clusters. These types were only found in the genome of P. cactorum. One contains a PLC domain only; whereas the other bears an additional Ca2+-dependent lipid-binding, or C2-domain, Figure 3.
3.5. Phosphatidic Acid in Oomycetes
Phosphatidic acid (PA) is a minor structural component of eukaryotic membranes. It is an important second messenger in many organisms including plants, animals and oomycetes, through its abilities to interact with a range of proteins and to induce membrane curvature [65]. PA is involved in cyst production in both P. palmivora [62] and in P. infestans [55,62]. PA can be generated by hydrolysis of phosphatidylcholine by phospholipase D (PLD) isozymes [66]. The other product of PLD activity, choline, is reported to bind to sigma-1-receptors in the endoplasmic reticulum, sensitizing IP3R gating to IP3 [67]. An alternative pathway for formation of PA is through phosphorylation of DAG by DAG kinases [68]. PA can also be produced by a third pathway, requiring the action of lysophosphatidic acid acyltransferases on lysophosphatidic acid [65].
Both the PLD- and PLC-dependent PA production pathways are activated by mastoparan in plant and P. infestans cells [55]. In P. palmivora, pectin induces a large and rapid accumulation of PA, which are consistent with it playing a second messenger role in zoospore encystment [62]. It is hypothesized that in Phytophthora spp., the majority of PA production occurs via the PLD pathway. This could explain the large number of PLDs, the limited range of DAG kinases, and the apparent lack of PLC genes in P. infestans [24].
3.6. IP3Rs in Oomycetes
IP3Rs have been studied across a wide variety of eukaryotes due to their role in Ca2+ signalling. However, their functions within oomycetes are yet to be elucidated. Different oomycete species appear to possess different numbers of IP3R homologues. For example, P. infestans encodes one putative IP3R homologue, whereas the Saprolengnia diclina genome contains 4 candidate homologues [9,19].
Vertebrate genomes encode three closely related IP3R subunits (IP3R1–3, sharing 60–80% amino acid identity), which assemble into homo and heterotetrametric channel complexes. The size of the subunits, at approximately 2700 amino acid residues, make them among the largest ion channel complexes identified to date. The three IP3R subtypes differ in their subcellular distributions, affinities for IP3 (IP3R2 > IP3R1 > IP3R3) and modulation by associated proteins [72]. Even though in mammals, most cells express two or three different IP3R subtypes, they differ in their distribution between tissues and subcellular domains.
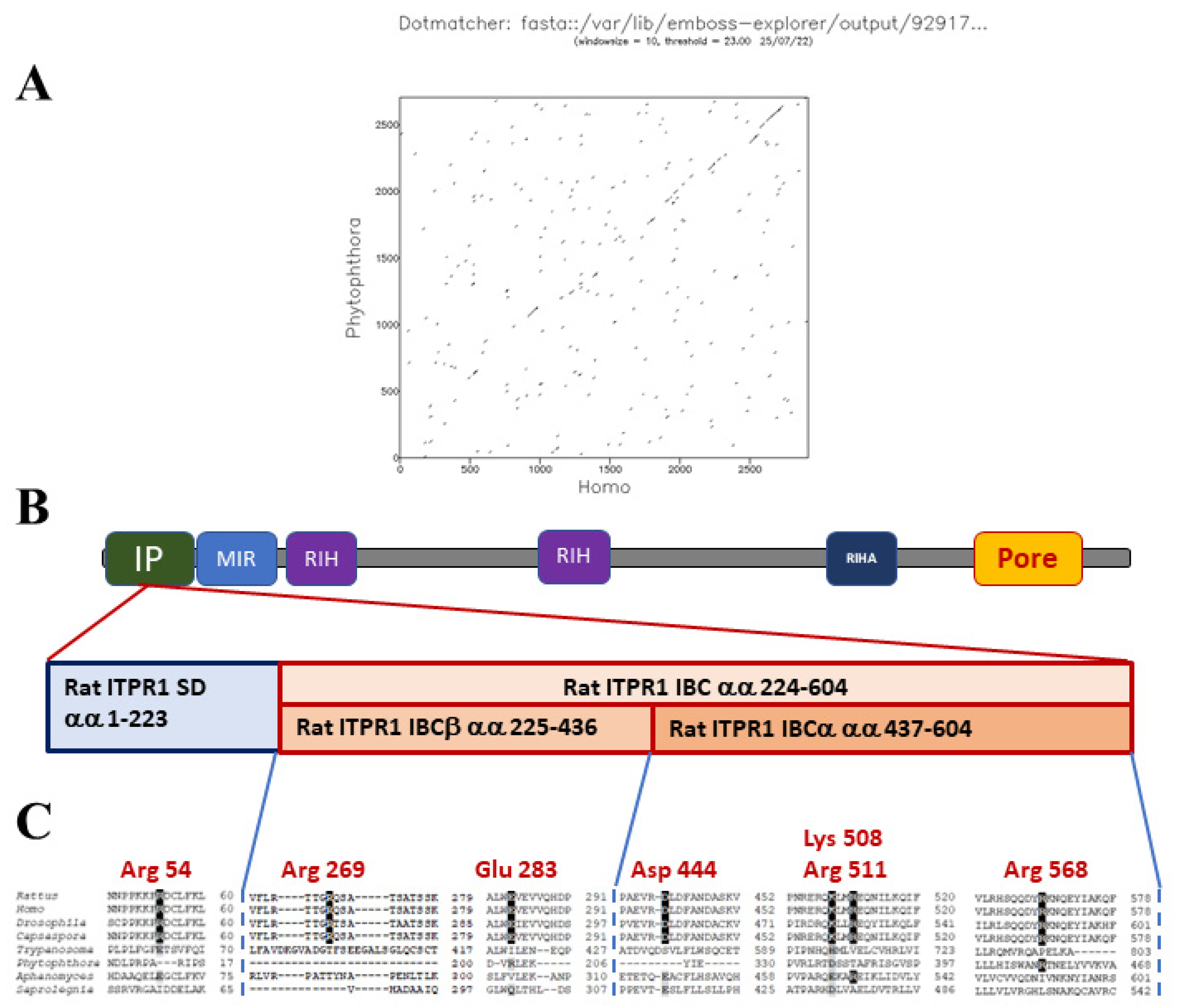
Mammalian IP3Rs are activated by the binding of IP3 to a clam-like structure, the IP3-binding core (IBC), located near the N-terminus of each IP3R subunit. Binding of IP3 alters the interaction between the IBC and the N-terminal suppressor domain (SD), Figure 4. These changes are proposed to disrupt interactions between the N-terminal regions of the four subunits of the IP3R, leading to gating of the intrinsic ion channel. The channel pore is formed by transmembrane domains towards the C-terminus of each IP3R subunit [73]. Binding of the IP3 to the receptor evokes conformational changes that expose of an activating Ca2+ binding site, leading to channel gating. There is also an inhibitory Ca2+ binding site, which promotes channel closure at higher Ca2+ concentrations [74].
Putative oomycete IP3R homologues were identified in conceptual translations of oomycete genomes, using the basic local alignment of sequences (BLAST) tool with human type 1 IP3R (ITPR1) protein as the query sequence [19]. Oomycete IP3Rs show poor conservation of amino acids within the IBC that are critical for IP3-binding to mammalian IP3Rs, Figure 4. This suggests that either IP3 is not an endogenous ligand of oomycete IP3Rs, or that these channels interact with the second messenger in a different way. We consider that the latter possibility is more likely, since at least some oomycetes display stimulus-dependent increases in IP3. This concept is not without precedent, since trypanosome IP3Rs display poor conservation of critical residues in the IBC, but have been experimentally demonstrated to act as IP3-activated Ca2+-release channels [75,76]. Furthermore, several oomycete Orders possess candidate PI-PLCs. Finally, all oomycete genomes analysed encode enzymes with the potential for the inactivation of IP3.
4. Enzymes That Inactivate IP3
IP3 is metabolized into forms less effective at IP3R gating, by removal of phosphate groups by inositol polyphosphate (IP) phosphatases and by phosphorylation by IPkinases. IP3 can be progressively dephosphorylated to inositol. DAG is phosphorylated by DAG-kinase-ɛ to form phosphatidic acid (PA). Additional enzymatic steps result in the formation of PIP2 from these products [57], Figure 1. Initially, IP3 is metabolized in two ways; either dephosphorylation by a 5-phosphatase to give inositol 1,4-bisphosphate (IP2) or through phosphorylation by IP3K to produce IP4 [58].
4.1. Oomycete IP Kinases
Metabolism of IP3 results in the production of diverse inositol polyphosphates (IPs), such as IP4, IP5, IP6, and PP-IP5 [59]. In mammals, inositol 1,4,5-trisphosphate 3-kinase (IP3 3-kinase/IP3K) plays a key role in maintaining Ca2+ homeostasis. It phosphorylates IP3 to inositol 1,3,4,5-tetrakisphosphate (IP4). IP4 is another second messenger, which inhibits store-operated Ca2+-entry; reduces translocation of Akt, Bkt and Itk protein kinases; remodels vesicular transport; regulates GTPase-activating proteins; and possibly mobilizes intracellular Ca2+ by acting synergistically with IP3 [77].
IP3 3-kinases (ITP3Ks) are a family of enzymes that phosphorylate IP3 to IP4 [59]. There are two major functional domains in mammalian ITP3Ks: a highly conserved C-terminal catalytic domain and a divergent N-terminal regulatory domain. The catalytic core of mammalian ITP3Ks consists of two domains: a large α/β-class structure and a small α-helical structure [78,79]. Three ITP3Ks are encoded by the human genome: ITPKA, ITPKB, and ITPKC. All share a conserved C-terminal catalytic domain but differ in mechanisms of regulation, as well as in their tissue distribution [80].
Mammalian ITP3Ks can be activated by calmodulin (CaM) in a Ca2+- dependent manner. CaM recognizes sequences which contain amphiphilic α-helices with clusters of positively charged and hydrophobic amino acids [81]. ITPKs are stimulated directly by Ca2+/calmodulin binding [82]. The mechanism by which this works is that CaM recognizes sequences that contain amphiphilic alpha-helices with clusters of positively charged and hydrophobic amino acids [83]. Certain sequences are required for CaM binding and enzyme activation and this level of stimulation by this interaction appears to be specific to cell, tissue, or isoform. ITP3Ks from nematodes and Arabidopsis thaliana lack the CaM-binding sites and therefore are insensitive to Ca2+ and CaM [80].
All ITPKs belong to a larger structural family, the inositol polyphosphate kinases or IPKs. Inositol phosphate multikinase (IPMK) is widely distributed in the kingdoms of animal, plant, and yeast [84]. Cellular IP3 serves as a substrate for both ITP3K and IPMK to form IP6. IPMK, but not ITP3K, is the major enzyme in IP6 synthesis. ITPK mainly functions in IP4 synthesis from IP3 [85]. Other kinases generate higher IPs. These higher IPs have multiple regulatory roles in cells, including control of DNA repair, endocytosis and chloride channel gating [58,59]. Inositol tetrakisphosphate 1-kinase (ITPK1) phosphorylates position 1 of Ins(3,4,5,6)P4 to generate Ins(1,3,4,5,6)P5, or the 5 position of Ins(1,3,4)P3 to produce Ins(1,3,4,5)P4 [86]. Inositol-pentakisphosphate 2-kinases (IPPK) phosphorylate Ins(1,3,4,5,6)P5 at the 2 position to generate Ins(1,2,3,4,5,6)P6 [87].
Among the oomycetes investigated, ITP3K homologues were only detected in Pythium oligandrum and in Saprolegnia parasitica. These homologues only display weak homology with H. sapiens ITPKA and display greater identity to metazoan IMPK proteins, meaning that they are more likely to belong to the latter class of enzymes, Figure 2 and Supplementary Table S1. In contrast, well-supported homologues of IMPK were detected in all translated oomycete genomes investigated, suggesting that these multikinases are likely to operate in these organisms. Most oomycetes examined also possess ITPK1 and IPPK homologues, suggesting that they have the molecular apparatus for both inactivating the Ca2+-releasing second messenger IP3, and for generating higher IPs that have their own cellular roles.
4.2. Oomycete IP Phosphatases
The Ca2+-mobilizing activity of IP3 can also be ablated by IP phosphatases. Some IP phosphatases only dephosphorylate lipid-associated inositol groups (PI, PIP2, PIP3), some only soluble inositol phosphates, while others have broad substrate specificities [57]. Inositol monophosphatases (IMPAs) convert IP1 to inositol, a key step in the generation of PI lipids. IMPA can also dephosphorylate a wide range of substrates, including Ins(1,3)P2, Ins(1,4)P2, glucose-1-phosphate, fructose-1-phosphate and β-glycerol phosphate [88].
Inositol polyphosphate 1-phosphatases (INPP1) catalyses the removal of the 1-phosphate from Ins(1,4)P2 and Ins(1,3,4)P2. Both IMPA and INPP1 enzymes are inhibited by lithium, an ion that promotes IP3 accumulation and inhibits inositol formation within cells [89]. Inositol polyphosphate 4-phosphatase type I isozymes (INPP4A and INPP4B) remove the phosphate from the 4 position of the lipids PI(3,4)P2 and PI(1,4)P2, in addition to those of the soluble Ins(1,4)P2 and Ins(1,3,4)P3 [90]. Inositol polyphosphate-5-phosphatases are a multigene family of signalling enzymes. Of these, INPP5A, also called the SH2-containing inositol phosphatase (SHIP), dephosphorylates Ins(1,4,5)P3, Ins(1,3,4,5)P4 and PI(3,4,5)P3 at the 5 position, thereby inactivating these signalling molecules [91]. Multiple inositol polyphosphate phosphatase (MINPP1) is a member of the histidine phosphatase family of proteins, which removes phosphate groups from a wide variety of substrates, including IP3, IP4, IP5 and IP6. MINPP1 is highly conserved among eukaryotes [92]. It also serves as a signalling hub, in the interconversion of different forms of IP-containing molecules [93].
The current study provides strong evidence for homologues of IMPA1, INPP4 and MINPP1 in all oomycetes investigated, Figure 2. In contrast, no robust homologues of INPP5 were detected and matches obtained for INPP1 were dubious. For example, the hit retrieved from P. infestans with a H. sapiens INPP1 query sequence is annotated as an ecdysteroid kinase and probably weakly matches mammalian INPP1 through a shared nucleotidase domain. Overall, these findings indicate that oomycetes display both similarities and differences with other eukaryotes, in terms of their mechanisms for dephosphorylating IPs.
5. Perspectives
Oomycetes are economically important pathogens of plants and aquatic animals. The current work and previous studies have highlighted key differences in PI signalling between oomycetes and their hosts. while most oomycete genomes investigated appear to encode IP3R homologues, only those of saprolegnians contain robust PI-PLC homologues. Oomycete IP3R homologues display limited conservation of key residues required for IP3 binding by their mammalian counterparts. Furthermore, oomycete genomes encode an extensive suite of enzymes for IP3 metabolism: at the very least homologues of the IP kinases ITPK1, IPPK, and IPMK; and of the IP phosphatases IMPA1, INPP4 and MINPP1. Given these observations and conflicting findings on detection of PIP2 and IP3 in oomycetes, it is likely that IP3-signalling in these pathogens is very different from that in other eukaryotic groups. Potential mechanisms of coupling between extracellular stimuli and IP3Rs in oomycetes include amplification of Ca2+-influx by CICR or activation of Ca2+-dependent PI-PLCs (in some taxa); or PLD-dependent pathways, such as sensitization of IP3Rs via choline binding to sigma-1-receptors. Mechanisms of IP3R-mediated Ca2+-signalling that are distinct from those in mammals have been observed in other eukaryotes. For example, the IP3R of the euglenozoan Trypanosoma brucei is located in a specialized organelle called the acidocalciosome, where it is activated by products of intraluminal polyphosphate hydrolysis [76].
Key experimental steps in the further characterization of oomycete PI-signalling are: (1) lipidomic analyses of the turnover of signalling lipids in response to extracellular stimulation. For example, do mastoparan, cold-shock or cues from hosts stimulate the conversion of PIP2 to PIP? (2) Determination if oomycete IP3Rs can act as IP3-gated Ca2+-channels. This might be achieved in heterologous expression of these proteins. If these proteins form channels that are not gated by IP3, which second messenger(s) are they opened by? (3) Are oomycete IP-kinases and -phosphatases functional? If so, what are their biological roles and how do they differ from their counterparts in other eukaryotes?
Addressing these questions is likely to lead to new insights into oomycete biology and to novel strategies for combatting these pathogenic organisms.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms10112157/s1, Table S1: Homologues of different inositol phosphate signalling components in several taxonomic groups; Table S2: Homologues of phosphoinositide selective phospholipase C (PI-PLC) isozymes among the SAR group, and representatives of Metazoa and Discoba.
Author Contributions
This work was conceived and supervised by J.J.M. and B.D.P. Literature review and preparation of the original draft of this review were performed by I.M.N. and E.C. All authors contributed to the preparation and revision of the submitted version. J.J.M. curated and analysed protein data, generated figures and acquired funding that supported this work. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Science Foundation Ireland, Frontiers for the Future grant number 20/FFP-P/8686, awarded to JJM.
Data Availability Statement
All data used in this review are either presented in this work or in the associated Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Judelson, H.S.; Blanco, F.A. The Spores of Phytophthora: Weapons of the Plant Destroyer. Nat. Rev. Microbiol. 2005, 3, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.H.Y.; Tyler, B.M. Mechanisms and Evolution of Virulence in Oomycetes. Annu. Rev. Phytopathol. 2012, 50, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Judelson, H.S. Metabolic Diversity and Novelties in the Oomycetes. Annu. Rev. Microbiol. 2017, 71, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Thines, M. Phylogeny and Evolution of Plant Pathogenic Oomycetes—A Global Overview. Eur. J. Plant Pathol. 2014, 138, 431–447. [Google Scholar] [CrossRef]
- Plattner, H.; Verkhratsky, A. Inseparable Tandem: Evolution Chooses ATP and Ca2+ to Control Life, Death and Cellular Signalling. Philos. Trans. R Soc. Lond. B Biol. Sci. 2016, 371, 20150419. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E. Calcium Signaling. Cell 1995, 80, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The Versatility and Universality of Calcium Signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef]
- Zhivotovsky, B.; Orrenius, S. Calcium and Cell Death Mechanisms: A Perspective from the Cell Death Community. Cell Calcium 2011, 50, 211–221. [Google Scholar] [CrossRef]
- Zheng, L.; Mackrill, J.J. Calcium Signaling in Oomycetes: An Evolutionary Perspective. Front. Physiol. 2016, 7, 123. [Google Scholar] [CrossRef] [Green Version]
- Bose, J.; Pottosin, I.I.; Shabala, S.S.; Palmgren, M.G.; Shabala, S. Calcium Efflux Systems in Stress Signaling and Adaptation in Plants. Front. Plant Sci. 2011, 2, 85. [Google Scholar] [CrossRef]
- Ehrlich, B.E.; Kaftan, E.; Bezprozvannaya, S.; Bezprozvanny, I. The Pharmacology of Intracellular Ca(2+)-Release Channels. Trends Pharmacol. Sci. 1994, 15, 145–149. [Google Scholar] [CrossRef]
- Demidchik, V.; Shabala, S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium Transport across Plant Membranes: Mechanisms and Functions. New Phytol. 2018, 220, 49–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman-Gavrila, L.B.; Lew, R.R. Calcium Gradient Dependence of Neurospora Crassa Hyphal Growth. Microbiology 2003, 149, 2475–2485. [Google Scholar] [CrossRef] [PubMed]
- Krinke, O.; Novotná, Z.; Valentová, O.; Martinec, J. Inositol Trisphosphate Receptor in Higher Plants: Is It Real? J. Exp. Bot. 2007, 58, 361–376. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, G.L.; Brownlee, C. Ca2+ Signalling in Plants and Green Algae--Changing Channels. Trends Plant Sci. 2008, 13, 506–514. [Google Scholar] [CrossRef]
- Mackrill, J.J. Ryanodine Receptor Calcium Release Channels: An Evolutionary Perspective. Adv. Exp. Med. Biol. 2012, 740, 159–182. [Google Scholar] [CrossRef]
- Putney, J.W. Forms and Functions of Store-Operated Calcium Entry Mediators, STIM and Orai. Adv. Biol. Regul. 2018, 68, 88–96. [Google Scholar] [CrossRef]
- Matsuda, T.; Takuma, K.; Baba, A. Na(+)-Ca2+ Exchanger: Physiology and Pharmacology. Jpn. J. Pharmacol. 1997, 74, 1–20. [Google Scholar] [CrossRef]
- Zheng, L.; Prestwich, B.D.; Harrison, P.T.; Mackrill, J.J. Polycystic Kidney Disease Ryanodine Receptor Domain (PKDRR) Proteins in Oomycetes. Pathogens 2020, 9, 577. [Google Scholar] [CrossRef]
- Offermanns, S. G-Proteins as Transducers in Transmembrane Signalling. Prog. Biophys. Mol. Biol. 2003, 83, 101–130. [Google Scholar] [CrossRef]
- Gresset, A.; Sondek, J.; Harden, T.K. The Phospholipase C Isozymes and Their Regulation. In Phosphoinositides I: Enzymes of Synthesis and Degradation; Balla, T., Wymann, M., York, J.D., Eds.; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2012; Volume 58, pp. 61–94. ISBN 978-94-007-3011-3. [Google Scholar]
- Kikkawa, U. The Story of PKC: A Discovery Marked by Unexpected Twists and Turns. IUBMB Life 2019, 71, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Alzayady, K.J.; Wang, L.; Chandrasekhar, R.; Wagner, L.E.; Van Petegem, F.; Yule, D.I. Defining the Stoichiometry of Inositol 1,4,5-Trisphosphate Binding Required to Initiate Ca2+ Release. Sci. Signal. 2016, 9, ra35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijer, H.J.G.; Govers, F. Genomewide Analysis of Phospholipid Signaling Genes in Phytophthora Spp.: Novelties and a Missing Link. Mol. Plant Microbe Interact. 2006, 19, 1337–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latijnhouwers, M.; de Wit, P.J.G.M.; Govers, F. Oomycetes and Fungi: Similar Weaponry to Attack Plants. Trends Microbiol. 2003, 11, 462–469. [Google Scholar] [CrossRef]
- Kamoun, S.; Huitema, E.; Vleeshouwers, V.G. Resistance to Oomycetes: A General Role for the Hypersensitive Response? Trends Plant Sci. 1999, 4, 196–200. [Google Scholar] [CrossRef]
- Thines, M. Oomycetes. Curr. Biol. 2018, 28, R812–R813. [Google Scholar] [CrossRef] [Green Version]
- Matari, N.H.; Blair, J.E. A Multilocus Timescale for Oomycete Evolution Estimated under Three Distinct Molecular Clock Models. BMC Evol. Biol. 2014, 14, 101. [Google Scholar] [CrossRef] [Green Version]
- O’Hanlon, R.; McCracken, A.R.; Cooke, L.R. Diversity and Ecology of Phytophthora Species on the Island of Ireland. Biol. Environ. Proc. R. Ir. Acad. 2016, 116B, 27–51. [Google Scholar] [CrossRef]
- Lin, C.C.; Aronson, J.M. Chitin and Cellulose in the Cell Walls of the Oomycete, Apodachlya Sp. Arch. Mikrobiol. 1970, 72, 111–114. [Google Scholar] [CrossRef]
- Selosse, M.-A.; Strullu-Derrien, C.; Martin, F.M.; Kamoun, S.; Kenrick, P. Plants, Fungi and Oomycetes: A 400-Million Year Affair That Shapes the Biosphere. New Phytol. 2015, 206, 501–506. [Google Scholar] [CrossRef]
- Sukul, P.; Spiteller, M. Metalaxyl: Persistence, Degradation, Metabolism, and Analytical Methods. Rev. Environ. Contam. Toxicol. 2000, 164, 1–26. [Google Scholar] [PubMed]
- Lerksuthirat, T.; Sangcakul, A.; Lohnoo, T.; Yingyong, W.; Rujirawat, T.; Krajaejun, T. Evolution of the Sterol Biosynthetic Pathway of Pythium Insidiosum and Related Oomycetes Contributes to Antifungal Drug Resistance. Antimicrob. Agents Chemother. 2017, 61, e02352-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Zhou, Q.; Xi, J.; Li, D.-L.; Schnabel, G.; Zhan, J. Analysis of RPA190 Revealed Multiple Positively Selected Mutations Associated with Metalaxyl Resistance in Phytophthora Infestans. Pest Manag. Sci. 2018, 74, 1916–1924. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.L.; Hardham, A.R. A Transient Rise in Cytoplasmic Free Calcium Is Required to Induce Cytokinesis in Zoosporangia of Phytophthora Cinnamomi. Eur. J. Cell Biol. 1996, 69, 180–188. [Google Scholar] [PubMed]
- Garrill, A.; Jackson, S.L.; Lew, R.R.; Heath, I.B. Ion Channel Activity and Tip Growth: Tip-Localized Stretch-Activated Channels Generate an Essential Ca2+ Gradient in the Oomycete Saprolegnia Ferax. Eur. J. Cell Biol. 1993, 60, 358–365. [Google Scholar]
- Walker, C.A.; van West, P. Zoospore Development in the Oomycetes. Fungal Biol Rev. 2007, 21, 10–18. [Google Scholar] [CrossRef]
- Hua, C.; Wang, Y.; Zheng, X.; Dou, D.; Zhang, Z.; Govers, F.; Wang, Y. A Phytophthora Sojae G-Protein Alpha Subunit Is Involved in Chemotaxis to Soybean Isoflavones. Eukaryot. Cell 2008, 7, 2133–2140. [Google Scholar] [CrossRef] [Green Version]
- Addepalli, M.K.; Fujita, Y. Regulatory Role of External Calcium on Pythium Porphyrae (Oomycota) Zoospore Release, Development and Infection in Causing Red Rot Disease of Porphyra Yezoensis (Rhodophyta). FEMS Microbiol. Lett. 2002, 211, 253–257. [Google Scholar] [CrossRef]
- Plattner, H.; Verkhratsky, A. Ca2+ Signalling Early in Evolution--All but Primitive. J. Cell Sci. 2013, 126, 2141–2150. [Google Scholar] [CrossRef] [Green Version]
- Tani, S.; Yatzkan, E.; Judelson, H.S. Multiple Pathways Regulate the Induction of Genes during Zoosporogenesis in Phytophthora Infestans. Mol. Plant Microbe. Interact. 2004, 17, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Tani, S.; Kim, K.S.; Judelson, H.S. A Cluster of NIF Transcriptional Regulators with Divergent Patterns of Spore-Specific Expression in Phytophthora Infestans. Fungal Genet. Biol. 2005, 42, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Latijnhouwers, M.; Ligterink, W.; Vleeshouwers, V.G.A.A.; van West, P.; Govers, F. A Galpha Subunit Controls Zoospore Motility and Virulence in the Potato Late Blight Pathogen Phytophthora Infestans. Mol. Microbiol. 2004, 51, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Tyler, B.M.; Tripathy, S.; Zhang, X.; Dehal, P.; Jiang, R.H.Y.; Aerts, A.; Arredondo, F.D.; Baxter, L.; Bensasson, D.; Beynon, J.L.; et al. Phytophthora Genome Sequences Uncover Evolutionary Origins and Mechanisms of Pathogenesis. Science 2006, 313, 1261–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, C.; Meijer, H.J.G.; de Keijzer, J.; Zhao, W.; Wang, Y.; Govers, F. GK4, a G-Protein-Coupled Receptor with a Phosphatidylinositol Phosphate Kinase Domain in Phytophthora Infestans, Is Involved in Sporangia Development and Virulence. Mol. Microbiol 2013, 88, 352–370. [Google Scholar] [CrossRef]
- Yang, X.; Zhao, W.; Hua, C.; Zheng, X.; Jing, M.; Li, D.; Govers, F.; Meijer, H.J.G.; Wang, Y. Chemotaxis and Oospore Formation in Phytophthora Sojae Are Controlled by G-Protein-Coupled Receptors with a Phosphatidylinositol Phosphate Kinase Domain. Mol. Microbiol 2013, 88, 382–394. [Google Scholar] [CrossRef] [PubMed]
- van den Hoogen, J.; Govers, F. GPCR-Bigrams: Enigmatic Signaling Components in Oomycetes. PLoS. Pathog. 2018, 14, e1007064. [Google Scholar] [CrossRef]
- Wang, Y.; Li, A.; Wang, X.; Zhang, X.; Zhao, W.; Dou, D.; Zheng, X.; Wang, Y. GPR11, a Putative Seven-Transmembrane G Protein-Coupled Receptor, Controls Zoospore Development and Virulence of Phytophthora Sojae. Eukaryot. Cell 2010, 9, 242–250. [Google Scholar] [CrossRef] [Green Version]
- María Laxalt, A.; Latijnhouwers, M.; van Hulten, M.; Govers, F. Differential Expression of G Protein Alpha and Beta Subunit Genes during Development of Phytophthora Infestans. Fungal Genet. Biol. 2002, 36, 137–146. [Google Scholar] [CrossRef]
- Hua, C.; Zheng, X.; Wang, Y. G Protein Alpha Subunit May Help Zoospore to Find the Infection Site and Influence the Expression of RGS Protein. Commun. Integr. Biol. 2009, 2, 91–93. [Google Scholar] [CrossRef]
- Latijnhouwers, M.; Govers, F. A Phytophthora Infestans G-Protein Beta Subunit Is Involved in Sporangium Formation. Eukaryot. Cell 2003, 2, 971–977. [Google Scholar] [CrossRef] [Green Version]
- Higashijima, T.; Uzu, S.; Nakajima, T.; Ross, E.M. Mastoparan, a Peptide Toxin from Wasp Venom, Mimics Receptors by Activating GTP-Binding Regulatory Proteins (G Proteins). J. Biol. Chem. 1988, 263, 6491–6494. [Google Scholar] [CrossRef]
- Quarmby, L.M.; Yueh, Y.G.; Cheshire, J.L.; Keller, L.R.; Snell, W.J.; Crain, R.C. Inositol Phospholipid Metabolism May Trigger Flagellar Excision in Chlamydomonas Reinhardtii. J. Cell Biol. 1992, 116, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Mau, S.E.; Witt, M.R.; Vilhardt, H. Mastoparan, a Wasp Venom Peptide, Stimulates Release of Prolactin from Cultured Rat Anterior Pituitary Cells. J. Endocrinol. 1994, 142, 9–18. [Google Scholar] [CrossRef]
- Latijnhouwers, M.; Munnik, T.; Govers, F. Phospholipase D in Phytophthora Infestans and Its Role in Zoospore Encystment. Mol. Plant Microbe Interact. 2002, 15, 939–946. [Google Scholar] [CrossRef] [Green Version]
- Judelson, H.S.; Ah-Fong, A.M.V. The Kinome of Phytophthora Infestans Reveals Oomycete-Specific Innovations and Links to Other Taxonomic Groups. BMC Genom. 2010, 11, 700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balla, T. Phosphoinositides: Tiny Lipids with Giant Impact on Cell Regulation. Physiol. Rev. 2013, 93, 1019–1137. [Google Scholar] [CrossRef] [PubMed]
- Michell, R.H. First Came the Link between Phosphoinositides and Ca2+ Signalling, and Then a Deluge of Other Phosphoinositide Functions. Cell Calcium 2009, 45, 521–526. [Google Scholar] [CrossRef]
- Hatch, A.J.; York, J.D. SnapShot: Inositol Phosphates. Cell 2010, 143, 1030–1030.e1. [Google Scholar] [CrossRef] [Green Version]
- Howard, J.M.; Beech, P.L.; Callahan, D.L. Detection of Endogenous Phosphatidylinositol 4,5-Bisphosphate in Phytophthora Cinnamomi. Lipids 2018, 53, 835–839. [Google Scholar] [CrossRef]
- Tani, S.; Judelson, H. Activation of Zoosporogenesis-Specific Genes in Phytophthora Infestans Involves a 7-Nucleotide Promoter Motif and Cold-Induced Membrane Rigidity. Eukaryot. Cell 2006, 5, 745–752. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Griffith, J.M.; Grant, B.R. Role of Phosphatide Acid during Differentiation of Phytophthora Palmivora Zoospores. J. Gen. Microbiol. 1992, 138, 451–459. [Google Scholar] [CrossRef]
- Luzzi, V.; Sims, C.E.; Soughayer, J.S.; Allbritton, N.L. The Physiologic Concentration of Inositol 1,4,5-Trisphosphate in the Oocytes of Xenopus Laevis. J. Biol. Chem. 1998, 273, 28657–28662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockcroft, S. The Latest Phospholipase C, PLCeta, Is Implicated in Neuronal Function. Trends Biochem. Sci. 2006, 31, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Tanguy, E.; Wang, Q.; Moine, H.; Vitale, N. Phosphatidic Acid: From Pleiotropic Functions to Neuronal Pathology. Front. Cell Neurosci. 2019, 13, 2. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Cambronero, J. Phospholipase D in Cell Signaling: From a Myriad of Cell Functions to Cancer Growth and Metastasis. J. Biol. Chem. 2014, 289, 22557–22566. [Google Scholar] [CrossRef] [Green Version]
- Brailoiu, E.; Chakraborty, S.; Brailoiu, G.C.; Zhao, P.; Barr, J.L.; Ilies, M.A.; Unterwald, E.M.; Abood, M.E.; Taylor, C.W. Choline Is an Intracellular Messenger Linking Extracellular Stimuli to IP3-Evoked Ca2+ Signals through Sigma-1 Receptors. Cell Rep. 2019, 26, 330–337.e4. [Google Scholar] [CrossRef] [Green Version]
- Mérida, I.; Avila-Flores, A.; Merino, E. Diacylglycerol Kinases: At the Hub of Cell Signalling. Biochem. J. 2008, 409, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The Rapid Generation of Mutation Data Matrices from Protein Sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Thillaiappan, N.B.; Chakraborty, P.; Hasan, G.; Taylor, C.W. IP3 Receptors and Ca2+ Entry. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.-D.; Velamakanni, S.; Ishiyama, N.; Stathopulos, P.B.; Rossi, A.M.; Khan, S.A.; Dale, P.; Li, C.; Ames, J.B.; Ikura, M.; et al. Structural and Functional Conservation of Key Domains in InsP3 and Ryanodine Receptors. Nature 2012, 483, 108–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foskett, J.K.; White, C.; Cheung, K.-H.; Mak, D.-O.D. Inositol Trisphosphate Receptor Ca2+ Release Channels. Physiol. Rev. 2007, 87, 593–658. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Bartlett, P.J.; Thomas, A.P.; Moreno, S.N.J.; Docampo, R. Acidocalcisomes of Trypanosoma Brucei Have an Inositol 1,4,5-Trisphosphate Receptor That Is Required for Growth and Infectivity. Proc. Natl. Acad. Sci. USA 2013, 110, 1887–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potapenko, E.; Negrão, N.W.; Huang, G.; Docampo, R. The Acidocalcisome Inositol-1,4,5-Trisphosphate Receptor of Trypanosoma Brucei Is Stimulated by Luminal Polyphosphate Hydrolysis Products. J. Biol. Chem. 2019, 294, 10628–10637. [Google Scholar] [CrossRef]
- Miller, A.T.; Chamberlain, P.P.; Cooke, M.P. Beyond IP3: Roles for Higher Order Inositol Phosphates in Immune Cell Signaling. Cell Cycle 2008, 7, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.J.; Hurley, J.H. Crystal Structure of the Catalytic Core of Inositol 1,4,5-Trisphosphate 3-Kinase. Mol. Cell 2004, 15, 703–711. [Google Scholar] [CrossRef]
- González, B.; Schell, M.J.; Letcher, A.J.; Veprintsev, D.B.; Irvine, R.F.; Williams, R.L. Structure of a Human Inositol 1,4,5-Trisphosphate 3-Kinase: Substrate Binding Reveals Why It Is Not a Phosphoinositide 3-Kinase. Mol. Cell 2004, 15, 689–701. [Google Scholar] [CrossRef]
- Xia, H.J.; Yang, G. Inositol 1,4,5-Trisphosphate 3-Kinases: Functions and Regulations. Cell Res. 2005, 15, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Takazawa, K.; Erneux, C. Identification of Residues Essential for Catalysis and Binding of Calmodulin in Rat Brain Inositol 1,4,5-Trisphosphate 3-Kinase. Biochem. J. 1991, 280, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Lloyd-Burton, S.M.; Yu, J.C.H.; Irvine, R.F.; Schell, M.J. Regulation of Inositol 1,4,5-Trisphosphate 3-Kinases by Calcium and Localization in Cells. J. Biol. Chem. 2007, 282, 9526–9535. [Google Scholar] [CrossRef] [PubMed]
- Franco-Echevarría, E.; Baños-Sanz, J.I.; Monterroso, B.; Round, A.; Sanz-Aparicio, J.; González, B. A New Calmodulin-Binding Motif for Inositol 1,4,5-Trisphosphate 3-Kinase Regulation. Biochem. J. 2014, 463, 319–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shears, S.B. How Versatile Are Inositol Phosphate Kinases? Biochem. J. 2004, 377, 265–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeds, A.M.; Sandquist, J.C.; Spana, E.P.; York, J.D. A Molecular Basis for Inositol Polyphosphate Synthesis in Drosophila Melanogaster. J. Biol. Chem. 2004, 279, 47222–47232. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Shears, S.B. Multitasking in Signal Transduction by a Promiscuous Human Ins(3,4,5,6)P(4) 1-Kinase/Ins(1,3,4)P(3) 5/6-Kinase. Biochem. J. 2000, 351, 551–555. [Google Scholar] [CrossRef]
- Fujii, M.; York, J.D. A Role for Rat Inositol Polyphosphate Kinases RIPK2 and RIPK1 in Inositol Pentakisphosphate and Inositol Hexakisphosphate Production in Rat-1 Cells. J. Biol. Chem. 2005, 280, 1156–1164. [Google Scholar] [CrossRef] [Green Version]
- McAllister, G.; Whiting, P.; Hammond, E.A.; Knowles, M.R.; Atack, J.R.; Bailey, F.J.; Maigetter, R.; Ragan, C.I. CDNA Cloning of Human and Rat Brain Myo-Inositol Monophosphatase. Expression and Characterization of the Human Recombinant Enzyme. Biochem. J. 1992, 284, 749–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiroz, J.A.; Gould, T.D.; Manji, H.K. Molecular Effects of Lithium. Mol. Interv. 2004, 4, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Ferron, M.; Vacher, J. Characterization of the Murine Inpp4b Gene and Identification of a Novel Isoform. Gene 2006, 376, 152–161. [Google Scholar] [CrossRef]
- Damen, J.E.; Liu, L.; Rosten, P.; Humphries, R.K.; Jefferson, A.B.; Majerus, P.W.; Krystal, G. The 145-KDa Protein Induced to Associate with Shc by Multiple Cytokines Is an Inositol Tetraphosphate and Phosphatidylinositol 3,4,5-Triphosphate 5-Phosphatase. Proc. Natl. Acad. Sci. USA 1996, 93, 1689–1693. [Google Scholar] [CrossRef] [Green Version]
- Chi, H.; Tiller, G.E.; Dasouki, M.J.; Romano, P.R.; Wang, J.; O’keefe, R.J.; Puzas, J.E.; Rosier, R.N.; Reynolds, P.R. Multiple Inositol Polyphosphate Phosphatase: Evolution as a Distinct Group within the Histidine Phosphatase Family and Chromosomal Localization of the Human and Mouse Genes to Chromosomes 10q23 and 19. Genomics 1999, 56, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Blero, D.; Payrastre, B.; Schurmans, S.; Erneux, C. Phosphoinositide Phosphatases in a Network of Signalling Reactions. Pflug. Arch. 2007, 455, 31–44. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Generalized scheme of phosphoinositide signalling in eukaryotic cells. Extracellular signals ca interact with cell-surface receptors, such as 7-transmembrane G-protein coupled receptors (GPCRs) or receptor tyrosine kinases (RTKs). GPCRs are associated with heterotrimeric GTP-binding proteins. The Gaq/11 subunits of these dissociate from activated receptors to stimulate b-subtypes of phosphosphingolipids-specific phospholipase C (PI-PLC). Activated RTKs stimulate g-subtypes of PI-PLC, by phosphorylating specific tyrosine residues. PI-PLCs cleave the minor membrane lipid phosphatidyl 4,5-bisphosphate (PIP2) to form soluble IP3 and membrane associated DAG. IP3 interacts with IP3Rs located in Ca2+-storage organelles such as the endoplasmic reticulum and vacuole, gating an intrinsic ion channel to release Ca2+ into the cytoplasm. IP3 can be inactivated by progressive dephosphorylation by inositol phosphate (IP) phosphatases, to generate inositol. This is recycled during the regeneration of PIP2. IP3 is also phosphorylated by IP kinases to form higher IPs, which have diverse biological functions distinct from the Ca2+-releasing role of IP3.
Figure 1.
Generalized scheme of phosphoinositide signalling in eukaryotic cells. Extracellular signals ca interact with cell-surface receptors, such as 7-transmembrane G-protein coupled receptors (GPCRs) or receptor tyrosine kinases (RTKs). GPCRs are associated with heterotrimeric GTP-binding proteins. The Gaq/11 subunits of these dissociate from activated receptors to stimulate b-subtypes of phosphosphingolipids-specific phospholipase C (PI-PLC). Activated RTKs stimulate g-subtypes of PI-PLC, by phosphorylating specific tyrosine residues. PI-PLCs cleave the minor membrane lipid phosphatidyl 4,5-bisphosphate (PIP2) to form soluble IP3 and membrane associated DAG. IP3 interacts with IP3Rs located in Ca2+-storage organelles such as the endoplasmic reticulum and vacuole, gating an intrinsic ion channel to release Ca2+ into the cytoplasm. IP3 can be inactivated by progressive dephosphorylation by inositol phosphate (IP) phosphatases, to generate inositol. This is recycled during the regeneration of PIP2. IP3 is also phosphorylated by IP kinases to form higher IPs, which have diverse biological functions distinct from the Ca2+-releasing role of IP3.

Figure 2.
Oomycete genomes encode putative Gαq/11 subunits, IP kinases and IP phosphatases, but only members of the Order Saprolegniales possess detectable PI-PLC homologues.
Figure 2.
Oomycete genomes encode putative Gαq/11 subunits, IP kinases and IP phosphatases, but only members of the Order Saprolegniales possess detectable PI-PLC homologues.

Figure 3.
Phylogeny and protein domain architectures of PI-PLC proteins from members of the Stramenopile-Alveolate-Rhizaria (SAR) group. Panel (A) shows a reconstruction of the evolutionary history of candidate PI-PLC homologues inferred from Maximum-Likelihood analyses, using the JTT model [69] with 500 bootstrap replicates [70]. Branches that were reproduced in less than 50% of the bootstrap replicates were collapsed (node information indicates the % of replicates). This analysis used 53 amino acid sequence, with 3274 positions in the final dataset, and was conducted using MEGA11 software [71]. Letters in boxes represent taxonomic groups: A, alveolates; D, discobans; O, oomycetes; R, rhizarians; and T, thraustochytrids. For species names, see Table S2. Numbers within circles represent protein domain architectures. Panel (B) shows these different types of architectures, which are shown approximately to scale. Additionally, shown is a key to protein domains.
Figure 3.
Phylogeny and protein domain architectures of PI-PLC proteins from members of the Stramenopile-Alveolate-Rhizaria (SAR) group. Panel (A) shows a reconstruction of the evolutionary history of candidate PI-PLC homologues inferred from Maximum-Likelihood analyses, using the JTT model [69] with 500 bootstrap replicates [70]. Branches that were reproduced in less than 50% of the bootstrap replicates were collapsed (node information indicates the % of replicates). This analysis used 53 amino acid sequence, with 3274 positions in the final dataset, and was conducted using MEGA11 software [71]. Letters in boxes represent taxonomic groups: A, alveolates; D, discobans; O, oomycetes; R, rhizarians; and T, thraustochytrids. For species names, see Table S2. Numbers within circles represent protein domain architectures. Panel (B) shows these different types of architectures, which are shown approximately to scale. Additionally, shown is a key to protein domains.

Figure 4.
Comparison of IP3R proteins from mammals with that from P. infestans. Panel (A) shows an amino acid identity plot between IP3Rs from H. sapiens (ITPR1) with that from P. infestans. Note the limited identity between the proteins, apart from at the C-terminal channel domain [19]. Panel (B) displays the key protein domains (IP: IP3-binding domain; MIR: mannosyltransferase, IP3R and RyR domain; RIH: RyR and IP3R homology domain; RIHA: RIH-associated domain; Pore) within a mammalian IP3R (Rattus norvegicus ITPR1) and expanded view of the IP domain. This includes the suppressor domain (SD) and the β- and α-segments of the IP3 binding core (IBC). Panel (C) represents a multiple sequence alignment of candidate IP domain from the IP3Rs of several eukaryotic species: Rattus norvegicus ITPR1, Homo sapiens ITPR1, Drosophila melanogaster ITPR, Capsaspora owczarzaki ITPR, Phytophthora infestans ITPR, Aphanomyces astaci ITPR and Saprolegnia parasitica ITPR. Note the lack of conservation of residues that are critical for interactions between IP3 and mammalian IP domains, indicated in red text.
Figure 4.
Comparison of IP3R proteins from mammals with that from P. infestans. Panel (A) shows an amino acid identity plot between IP3Rs from H. sapiens (ITPR1) with that from P. infestans. Note the limited identity between the proteins, apart from at the C-terminal channel domain [19]. Panel (B) displays the key protein domains (IP: IP3-binding domain; MIR: mannosyltransferase, IP3R and RyR domain; RIH: RyR and IP3R homology domain; RIHA: RIH-associated domain; Pore) within a mammalian IP3R (Rattus norvegicus ITPR1) and expanded view of the IP domain. This includes the suppressor domain (SD) and the β- and α-segments of the IP3 binding core (IBC). Panel (C) represents a multiple sequence alignment of candidate IP domain from the IP3Rs of several eukaryotic species: Rattus norvegicus ITPR1, Homo sapiens ITPR1, Drosophila melanogaster ITPR, Capsaspora owczarzaki ITPR, Phytophthora infestans ITPR, Aphanomyces astaci ITPR and Saprolegnia parasitica ITPR. Note the lack of conservation of residues that are critical for interactions between IP3 and mammalian IP domains, indicated in red text.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Small molecules used to investigate PI signalling in oomycetes. This indicates the structural class, targets in mammalian cells and examples of effects on oomycetes, with accompanying reference citations.
Table 1.
Small molecules used to investigate PI signalling in oomycetes. This indicates the structural class, targets in mammalian cells and examples of effects on oomycetes, with accompanying reference citations.
| Small Molecule | Structural Class | Targets in Mammals | Example of Effects on Oomycetes |
|---|---|---|---|
| U73122 | Aminosteroid | PI-PLC inhibitor. Activator of H.sapiens PLCβ3. Ca2+-pump inhibitor. 5-lipoxygenase inhibitor. | Block of zoosporogenesis in P.infestans [41] Inhibition of LIM-interactor transcription factor expression in P. infestans [42] |
| Mastoparan | Peptide | PI-PLC activator. PI-PLC inhibitor. Membrane permeabilization. | Accumulation of phosphatidic acid in P.infestans (activation of phospholipase D?) [43] |
| 2-amino- ethoxydiphenyl borate (2-APB) | Diphenylborane | IP3R inhibitor SOCE modulator TRP channel activator Ca2+-pump inhibitor | Block of zoosporogenesis in P.infestans [41]. Inhibition of LIM-interactor transcription factor expression in P.infestans [42] |
| Ethylene glycol-bis(2-amino-ethylether)-N, N, N, N -tetraacetic acid (EGTA) | Aminopolycarboxylic acid | Ca2+-chelator Mg2+-chelator | Block of cyst germination in Pythium porphyrae [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nair, I.M.; Condon, E.; Prestwich, B.D.; Mackrill, J.J. Myo-D-inositol Trisphosphate Signalling in Oomycetes. Microorganisms 2022, 10, 2157. https://doi.org/10.3390/microorganisms10112157
AMA Style
Nair IM, Condon E, Prestwich BD, Mackrill JJ. Myo-D-inositol Trisphosphate Signalling in Oomycetes. Microorganisms. 2022; 10(11):2157. https://doi.org/10.3390/microorganisms10112157
Chicago/Turabian StyleNair, Indu Muraleedharan, Emma Condon, Barbara Doyle Prestwich, and John James Mackrill. 2022. "Myo-D-inositol Trisphosphate Signalling in Oomycetes" Microorganisms 10, no. 11: 2157. https://doi.org/10.3390/microorganisms10112157
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.