
The Marine Compound Tartrolon E Targets the Asexual and Early Sexual Stages of Cryptosporidium parvum
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Maintenance and Parasite Strains
2.2. Compounds
2.3. In Vitro Growth Inhibition Assay
2.4. Quantification of Parasite Growth by qPCR
2.5. Determination of Oocyst Excystation Rate
2.6. Sporozoite Viability Assay
2.7. Invasion Inhibition Assay
2.8. Quantification of Gene Expression by RT-qPCR
2.9. Statistics
3. Results
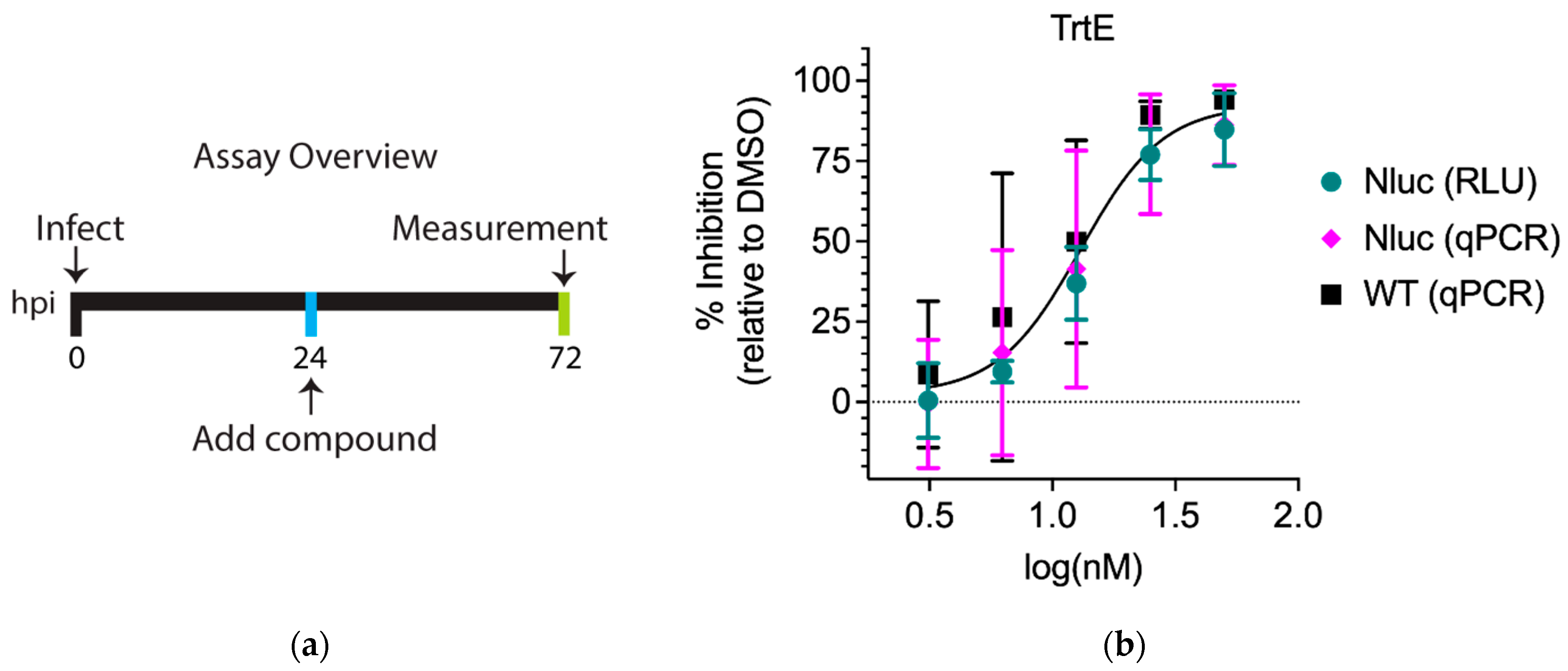
3.1. The Inhibition Curves of trtE against WT and Nluc C. parvum Strains Are Not Significantly Different
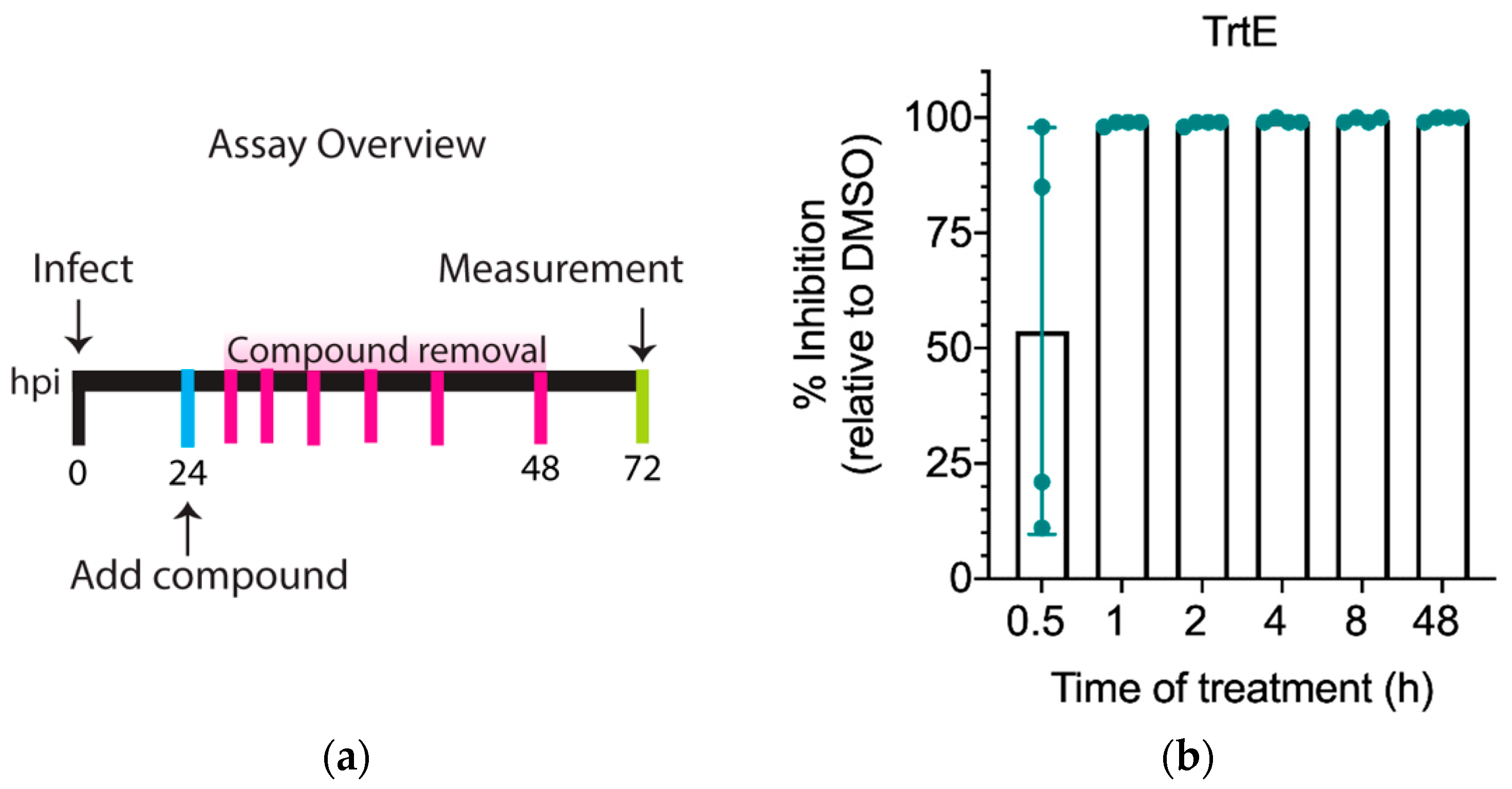
3.2. TrtE Kills C. parvum within the First Hour of Treatment
3.3. TrtE Has No Effect on Excystation
3.4. TrtE Inhibits Establishment of C. parvum Infection
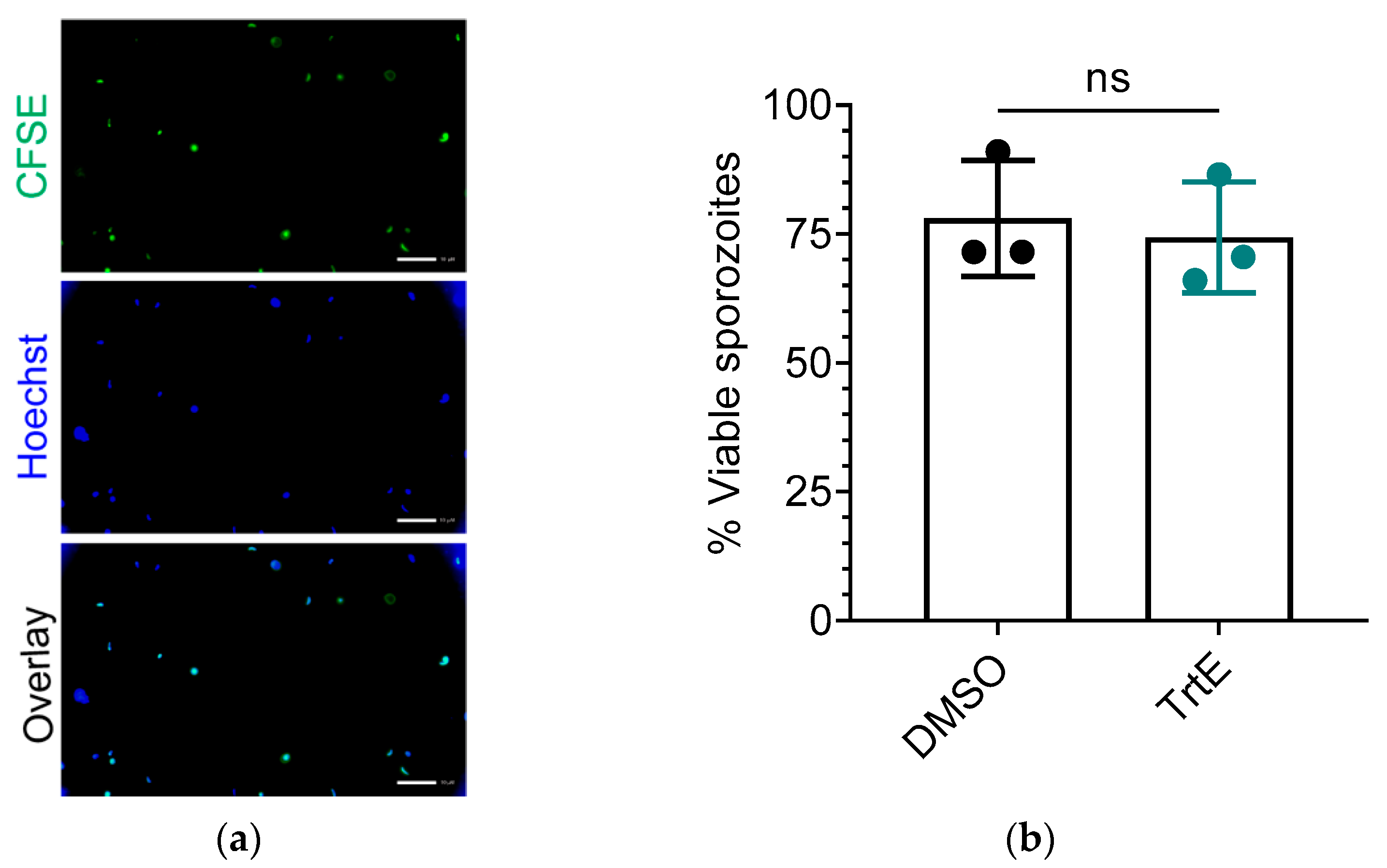
3.5. TrtE Does Not Reduce the Viability of C. parvum Sporozoites
3.6. TrtE Is Most Effective during the Asexual Stage of C. parvum
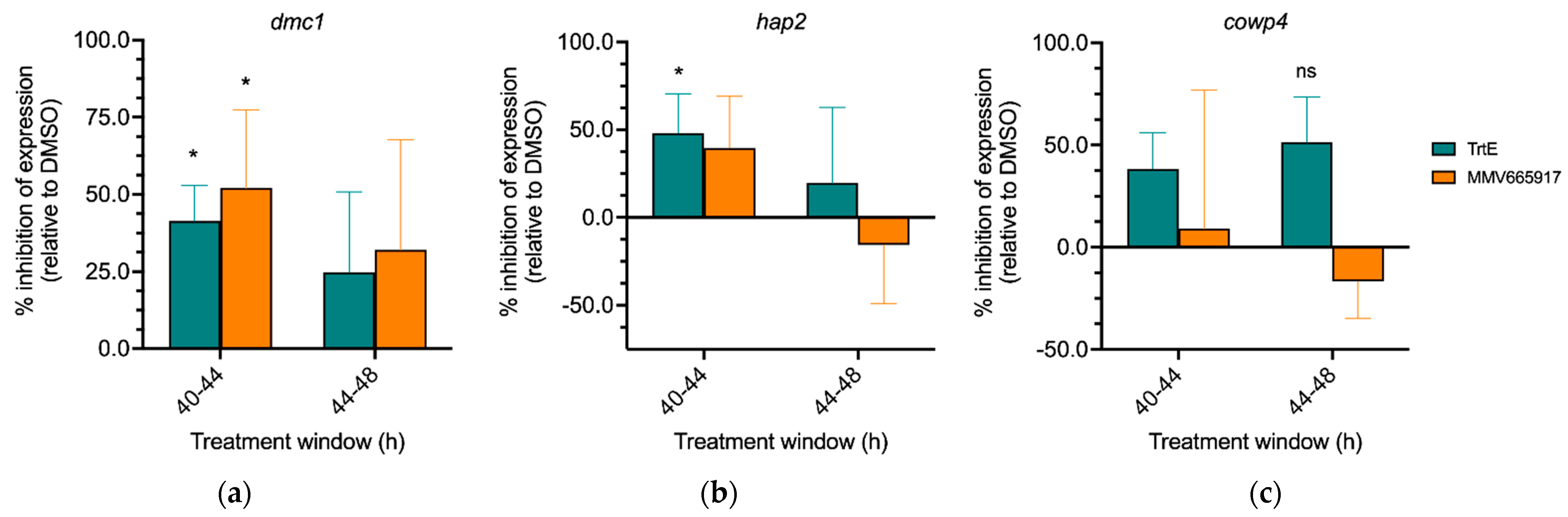
3.7. TrtE Targets Early Sexual Stages of Cryptosporidium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention (CDC) Cryptosporidiosis NNDSS Summary Report for 2019|Water-Related Topics|Healthy Water|CDC. Available online: https://www.cdc.gov/healthywater/surveillance/cryptosporidium/cryptosporidium-2019.html (accessed on 2 August 2022).
- Centers for Disease Control and Prevention (CDC) Pathogen & Environment|Cryptosporidium|Parasites|CDC. Available online: https://www.cdc.gov/parasites/crypto/pathogen.html (accessed on 2 August 2022).
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and Aetiology of Diarrhoeal Disease in Infants and Young Children in Developing Countries (the Global Enteric Multicenter Study, GEMS): A Prospective, Case-Control Study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Nasrin, D.; Blackwelder, W.C.; Wu, Y.; Farag, T.; Panchalingham, S.; Sow, S.O.; Sur, D.; Zaidi, A.K.M.; Faruque, A.S.G.; et al. The Incidence, Aetiology, and Adverse Clinical Consequences of Less Severe Diarrhoeal Episodes among Infants and Children Residing in Low-Income and Middle-Income Countries: A 12-Month Case-Control Study as a Follow-on to the Global Enteric Multicenter Study (GEMS). Lancet Glob. Health 2019, 7, e568–e584. [Google Scholar] [CrossRef] [Green Version]
- Boudou, M.; Cleary, E.; ÓhAiseadha, C.; Garvey, P.; McKeown, P.; O’Dwyer, J.; Hynds, P. Spatiotemporal Epidemiology of Cryptosporidiosis in the Republic of Ireland, 2008–2017: Development of a Space-Time “Cluster Recurrence” Index. BMC Infect. Dis. 2021, 21, 880. [Google Scholar] [CrossRef] [PubMed]
- Omolabi, K.F.; Odeniran, P.O.; Soliman, M.E. A Meta-Analysis of Cryptosporidium Species in Humans from Southern Africa (2000–2020). J. Parasit. Dis. Off. Organ Indian Soc. Parasitol. 2021, 46, 304–316. [Google Scholar] [CrossRef]
- Yulfi, H.; Fakhrur Rozi, M.; Andriyani, Y.; Masyithah Darlan, D. Prevalence of Cryptosporidium Spp. and Blastocystis Hominis in Faecal Samples among Diarrheic HIV Patients in Medan, Indonesia. Med. Glas. Off. Publ. Med. Assoc. Zenica-Doboj Cant. Bosnia Herzeg. 2021, 18, 55–61. [Google Scholar] [CrossRef]
- Vanathy, K.; Parija, S.C.; Mandal, J.; Hamide, A.; Krishnamurthy, S. Cryptosporidiosis: A Mini Review. Trop. Parasitol. 2017, 7, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Sjöström, M.; Arvidsson, M.; Söderström, L.; Lilja, M.; Lindh, J.; Widerström, M. Outbreak of Cryptosporidium hominis in Northern Sweden: Persisting Symptoms in a 5-Year Follow-Up. Parasitol. Res. 2022, 121, 2043–2049. [Google Scholar] [CrossRef]
- Bouzid, M.; Hunter, P.R.; Chalmers, R.M.; Tyler, K.M. Cryptosporidium Pathogenicity and Virulence. Clin. Microbiol. Rev. 2013, 26, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Jumani, R.S.; Hasan, M.M.; Stebbins, E.E.; Donnelly, L.; Miller, P.; Klopfer, C.; Bessoff, K.; Teixeira, J.E.; Love, M.S.; McNamara, C.W.; et al. A Suite of Phenotypic Assays to Ensure Pipeline Diversity When Prioritizing Drug-like Cryptosporidium Growth Inhibitors. Nat. Commun. 2019, 10, 1862. [Google Scholar] [CrossRef] [Green Version]
- English, E.D.; Guérin, A.; Tandel, J.; Striepen, B. Live Imaging of the Cryptosporidium parvum Life Cycle Reveals Direct Development of Male and Female Gametes from Type I Meronts. PLoS Biol. 2022, 20, e3001604. [Google Scholar] [CrossRef] [PubMed]
- Tandel, J.; English, E.D.; Sateriale, A.; Gullicksrud, J.A.; Beiting, D.P.; Sullivan, M.C.; Pinkston, B.; Striepen, B. Life Cycle Progression and Sexual Development of the Apicomplexan Parasite Cryptosporidium parvum. Nat. Microbiol. 2019, 4, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Sparks, H.; Nair, G.; Castellanos-Gonzalez, A.; White, A.C. Treatment of Cryptosporidium: What We Know, Gaps, and the Way Forward. Curr. Trop. Med. Rep. 2015, 2, 181–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abubakar, I.; Aliyu, S.H.; Arumugam, C.; Usman, N.K.; Hunter, P.R. Treatment of Cryptosporidiosis in Immunocompromised Individuals: Systematic Review and Meta-Analysis. Br. J. Clin. Pharmacol. 2007, 63, 387–393. [Google Scholar] [CrossRef] [Green Version]
- Elshahawi, S.I.; Trindade-Silva, A.E.; Hanora, A.; Han, A.W.; Flores, M.S.; Vizzoni, V.; Schrago, C.G.; Soares, C.A.; Concepcion, G.P.; Distel, D.L.; et al. Boronated Tartrolon Antibiotic Produced by Symbiotic Cellulose-Degrading Bacteria in Shipworm Gills. Proc. Natl. Acad. Sci. USA 2013, 110, E295–E304. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, R.M.; Nepveux, V.F.J.; Abenoja, J.; Bowden, G.; Reis, P.; Beaushaw, J.; Bone Relat, R.M.; Driskell, I.; Gimenez, F.; Riggs, M.W.; et al. A Symbiotic Bacterium of Shipworms Produces a Compound with Broad Spectrum Anti-Apicomplexan Activity. PLoS Pathog. 2020, 16, e1008600. [Google Scholar] [CrossRef]
- Bowden, G.D.; Reis, P.M.; Rogers, M.B.; Bone Relat, R.M.; Brayton, K.A.; Wilson, S.K.; Di Genova, B.M.; Knoll, L.J.; Nepveux, V.F.J.; Tai, A.K.; et al. A Conserved Coccidian Gene Is Involved in Toxoplasma Sensitivity to the Anti-Apicomplexan Compound, Tartrolon E. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 1–7. [Google Scholar] [CrossRef]
- Vinayak, S.; Pawlowic, M.C.; Sateriale, A.; Brooks, C.F.; Studstill, C.J.; Bar-Peled, Y.; Cipriano, M.J.; Striepen, B. Genetic Modification of the Diarrheal Pathogen Cryptosporidium parvum. Nature 2015, 523, 477–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bone Relat, R.M.; Winder, P.L.; Bowden, G.D.; Guzmán, E.A.; Peterson, T.A.; Pomponi, S.A.; Roberts, J.C.; Wright, A.E.; O’Connor, R.M. High-Throughput Screening of a Marine Compound Library Identifies Anti-Cryptosporidium Activity of Leiodolide A. Mar. Drugs 2022, 20, 240. [Google Scholar] [CrossRef]
- Godiwala, N.T.; Vandewalle, A.; Ward, H.D.; Leav, B.A. Quantification of In Vitro and In Vivo Cryptosporidium parvum Infection by Using Real-Time PCR. Appl. Environ. Microbiol. 2006, 72, 4484–4488. [Google Scholar] [CrossRef] [Green Version]
- Behera, A.K.; Thorpe, C.M.; Kidder, J.M.; Smith, W.; Hildebrand, E.; Hu, L.T. Borrelia Burgdorferi-Induced Expression of Matrix Metalloproteinases from Human Chondrocytes Requires Mitogen-Activated Protein Kinase and Janus Kinase/Signal Transducer and Activator of Transcription Signaling Pathways. Infect. Immun. 2004, 72, 2864–2871. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Boxell, A.; Hijjawi, N.; Monis, P.; Ryan, U. Comparison of Various Staining Methods for the Detection of Cryptosporidium in Cell-Free Culture. Exp. Parasitol. 2008, 120, 67–72. [Google Scholar] [CrossRef]
- Kato, S.; Lindergard, G.; Mohammed, H.O. Utility of the Cryptosporidium oocyst Wall Protein (COWP) Gene in a Nested PCR Approach for Detection Infection in Cattle. Vet. Parasitol. 2003, 111, 153–159. [Google Scholar] [CrossRef]
- Funkhouser-Jones, L.J.; Ravindran, S.; Sibley, L.D. Defining Stage-Specific Activity of Potent New Inhibitors of Cryptosporidium parvum Growth In Vitro. mBio 2020, 11, e00052-20. [Google Scholar] [CrossRef] [Green Version]
- Murphy, R.C.; Ojo, K.K.; Larson, E.T.; Castellanos-Gonzalez, A.; Perera, B.G.K.; Keyloun, K.R.; Kim, J.E.; Bhandari, J.G.; Muller, N.R.; Verlinde, C.L.M.J.; et al. Discovery of Potent and Selective Inhibitors of Calcium-Dependent Protein Kinase 1 (CDPK1) from C. parvum and T. gondii. ACS Med. Chem. Lett. 2010, 1, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Shu, F.; Li, Y.; Chu, W.; Chen, X.; Zhang, Z.; Guo, Y.; Feng, Y.; Xiao, L.; Li, N. Characterization of Calcium-Dependent Protein Kinase 2A, a Potential Drug Target Against Cryptosporidiosis. Front. Microbiol. 2022, 13, 883674. [Google Scholar] [CrossRef]
- Baragaña, B.; Forte, B.; Choi, R.; Nakazawa Hewitt, S.; Bueren-Calabuig, J.A.; Pisco, J.P.; Peet, C.; Dranow, D.M.; Robinson, D.A.; Jansen, C.; et al. Lysyl-TRNA Synthetase as a Drug Target in Malaria and Cryptosporidiosis. Proc. Natl. Acad. Sci. USA 2019, 116, 7015–7020. [Google Scholar] [CrossRef] [Green Version]
- Buckner, F.S.; Ranade, R.M.; Gillespie, J.R.; Shibata, S.; Hulverson, M.A.; Zhang, Z.; Huang, W.; Choi, R.; Verlinde, C.L.M.J.; Hol, W.G.J.; et al. Optimization of Methionyl TRNA-Synthetase Inhibitors for Treatment of Cryptosporidium Infection. Antimicrob. Agents Chemother. 2019, 63, e02061-18. [Google Scholar] [CrossRef] [Green Version]
- Love, M.S.; McNamara, C.W. Phenotypic Screening Techniques for Cryptosporidium Drug Discovery. Expert Opin. Drug Discov. 2021, 16, 59–74. [Google Scholar] [CrossRef]
- Manjunatha, U.H.; Vinayak, S.; Zambriski, J.A.; Chao, A.T.; Sy, T.; Noble, C.G.; Bonamy, G.M.C.; Kondreddi, R.R.; Zou, B.; Gedeck, P.; et al. A Cryptosporidium PI(4)K Inhibitor Is a Drug Candidate for Cryptosporidiosis. Nature 2017, 546, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Jumani, R.S.; Bessoff, K.; Love, M.S.; Miller, P.; Stebbins, E.E.; Teixeira, J.E.; Campbell, M.A.; Meyers, M.J.; Zambriski, J.A.; Nunez, V.; et al. A Novel Piperazine-Based Drug Lead for Cryptosporidiosis from the Medicines for Malaria Venture Open-Access Malaria Box. Antimicrob. Agents Chemother. 2018, 62, e01505-17. [Google Scholar] [CrossRef] [Green Version]
- Choi, R.; Hulverson, M.A.; Huang, W.; Vidadala, R.S.R.; Whitman, G.R.; Barrett, L.K.; Schaefer, D.A.; Betzer, D.P.; Riggs, M.W.; Doggett, J.S.; et al. Bumped Kinase Inhibitors as Therapy for Apicomplexan Parasitic Diseases: Lessons Learned. Int. J. Parasitol. 2020, 50, 413–422. [Google Scholar] [CrossRef]
- Bessoff, K.; Sateriale, A.; Lee, K.K.; Huston, C.D. Drug Repurposing Screen Reveals FDA-Approved Inhibitors of Human HMG-CoA Reductase and Isoprenoid Synthesis That Block Cryptosporidium parvum Growth. Antimicrob. Agents Chemother. 2013, 57, 1804–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulverson, M.A.; Choi, R.; McCloskey, M.C.; Whitman, G.R.; Ojo, K.K.; Michaels, S.A.; Somepalli, M.; Love, M.S.; McNamara, C.W.; Rabago, L.M.; et al. Repurposing Infectious Disease Hits as Anti-Cryptosporidium Leads. ACS Infect. Dis. 2021, 7, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Nachipo, P.; Hermann, D.; Quinnan, G.; Gordon, M.A.; Van Voorhis, W.C.; Iroh Tam, P.-Y. Evaluating the Safety, Tolerability, Pharmacokinetics and Efficacy of Clofazimine in Cryptosporidiosis (CRYPTOFAZ): Study Protocol for a Randomized Controlled Trial. Trials 2018, 19, 456. [Google Scholar] [CrossRef] [Green Version]
- Iroh Tam, P.; Arnold, S.L.M.; Barrett, L.K.; Chen, C.R.; Conrad, T.M.; Douglas, E.; Gordon, M.A.; Hebert, D.; Henrion, M.; Hermann, D.; et al. Clofazimine for Treatment of Cryptosporidiosis in Human Immunodeficiency Virus Infected Adults: An Experimental Medicine, Randomized, Double-Blind, Placebo-Controlled Phase 2a Trial. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2021, 73, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.X.; Love, M.S.; McNamara, C.W.; Chi, V.; Woods, A.K.; Joseph, S.; Schaefer, D.A.; Betzer, D.P.; Riggs, M.W.; Iroh Tam, P.-Y.; et al. Pharmacokinetics and Pharmacodynamics of Clofazimine for Treatment of Cryptosporidiosis. Antimicrob. Agents Chemother. 2022, 66, e0156021. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.A.; Urban, S.; Roessner, U. A Historical Overview of Natural Products in Drug Discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nweze, J.A.; Mbaoji, F.N.; Li, Y.-M.; Yang, L.-Y.; Huang, S.-S.; Chigor, V.N.; Eze, E.A.; Pan, L.-X.; Zhang, T.; Yang, D.-F. Potentials of Marine Natural Products against Malaria, Leishmaniasis, and Trypanosomiasis Parasites: A Review of Recent Articles. Infect. Dis. Poverty 2021, 10, 9. [Google Scholar] [CrossRef]
- Martens, M.C.; Liu, Y.; Sanford, A.G.; Wallick, A.I.; Warner, R.C.; Li, R.; Davis, P.H. Analogs of Marinopyrrole A Show Enhancement to Observed In Vitro Potency against Acute Toxoplasma gondii Infection. Antimicrob. Agents Chemother. 2022, 66, e00794-21. [Google Scholar] [CrossRef]
- Tempone, A.G.; Pieper, P.; Borborema, S.E.T.; Thevenard, F.; Lago, J.H.G.; Croft, S.L.; Anderson, E.A. Marine Alkaloids as Bioactive Agents against Protozoal Neglected Tropical Diseases and Malaria. Nat. Prod. Rep. 2021, 38, 2214–2235. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. parvum Strain | Quantification Method | EC50 (nM) | 95% Confidence Interval (CI) |
|---|---|---|---|
| Nluc | Luminescence | 15.9 | 13.8 to 18.4 |
| Nluc | qPCR | 14.8 | 10.4 to 21.7 |
| WT | qPCR | 11.2 | 7.6 to 16.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cotto-Rosario, A.; Miller, E.Y.D.; Fumuso, F.G.; Clement, J.A.; Todd, M.J.; O’Connor, R.M. The Marine Compound Tartrolon E Targets the Asexual and Early Sexual Stages of Cryptosporidium parvum. Microorganisms 2022, 10, 2260. https://doi.org/10.3390/microorganisms10112260
Cotto-Rosario A, Miller EYD, Fumuso FG, Clement JA, Todd MJ, O’Connor RM. The Marine Compound Tartrolon E Targets the Asexual and Early Sexual Stages of Cryptosporidium parvum. Microorganisms. 2022; 10(11):2260. https://doi.org/10.3390/microorganisms10112260
Chicago/Turabian StyleCotto-Rosario, Alexis, Emma Y. D. Miller, Fernanda G. Fumuso, Jason A. Clement, Matthew J. Todd, and Roberta M. O’Connor. 2022. "The Marine Compound Tartrolon E Targets the Asexual and Early Sexual Stages of Cryptosporidium parvum" Microorganisms 10, no. 11: 2260. https://doi.org/10.3390/microorganisms10112260