
Cross-Feeding and Enzymatic Catabolism for Mannan-Oligosaccharide Utilization by the Butyrate-Producing Gut Bacterium Roseburia hominis A2-183
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatic Analysis
2.1.1. Bioinformatic Prediction of Roseburia hominis A2-183 Gene Functions
2.1.2. Bioinformatic Analysis of RhMosBP, RhMOP130A, RhMan113A, and RhGal36A
2.2. Strains and Media
2.3. Monoculture and Coculture Fermentation Studies
2.4. Primer Design
2.5. Bacterial Growth Analysis
2.5.1. Genomic DNA Extraction
2.5.2. Quantitative PCR (qPCR)
2.6. Determination of Carbohydrate Utilization and Short-Chain Fatty Acids (SCFAs) Production
2.6.1. Utilization of Glucose and MOS/GMOS
2.6.2. Determination of Short-Chain Fatty Acids
2.7. Gene Expression Analyses
2.7.1. RNA Extraction and Purification
2.7.2. Monitoring Gene Expression
2.7.3. Relative Quantification of Gene Expression
2.8. Construction of Expression Plasmids for RhMOP130A, RhMan113A, and RhGal36A
2.9. Expression and Purification of RhMOP130A, RhMan113A, and RhGal36A
2.10. Protein Electrophoresis of RhMOP130A, RhMan113A, and RhGal36A
2.11. Saccharide Analysis by High-Performance Anion Exchange Chromatography–Pulsed Amperometric Detection (HPAEC–PAD)
2.12. Enzyme Activity Characterization of RhMOP130A
2.13. Enzyme Activity Characterization of RhMan113A
2.14. Enzyme Activity Characterization of RhGal36A
3. Results
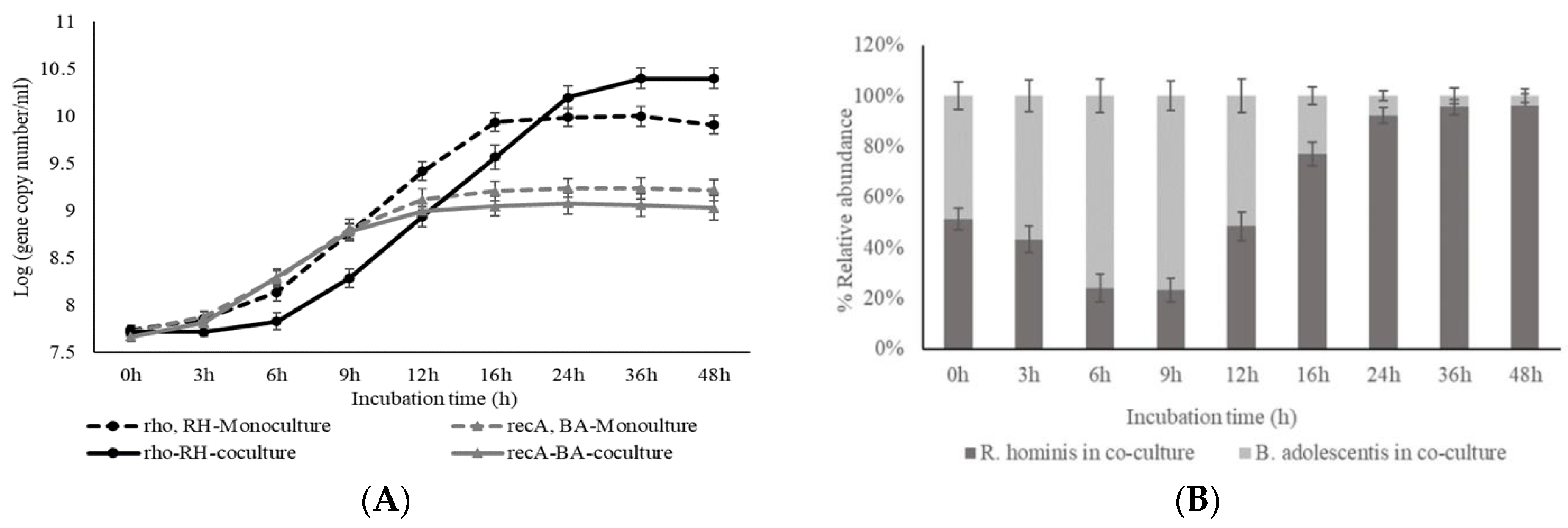
3.1. Mono- and Cocultivation of Bifidobacterium adolescentis EB1a (ATCC 15703) and Roseburia hominis A2-183 (DSMZ 16839)
3.1.1. Effect of Substrates on Bacterial Growth
3.1.2. Production of Short-Chain Fatty Acids (SCFAs) during Mono- and Cocultivation
3.1.3. Effect of Cocultivation on Glucose and MOS/GMOS Consumption
3.2. Role of the Locus RhMosUL and Other Genes in MOS/GMOS Utilization
3.2.1. Bioinformatic Analysis
3.2.2. Gene Expression Analysis, Using Reverse Transcription-Quantitative PCR (RT-qPCR)
3.2.3. Bioinformatic Analysis of RhMOP130A, RhMan113A, RhGal36A, and RhMosBP
3.2.4. Expression and Purification of RhMOP130A, RhMan113A, and RhGal36A
3.2.5. Catalytic Properties of RhMOP130A
3.2.6. Catalytic Properties of RhMan113A
3.2.7. Catalytic Properties of RhGal36A
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ndeh, D.; Gilbert, H.J. Biochemistry of Complex Glycan Depolymerisation by the Human Gut Microbiota. FEMS Microbiol. Rev. 2018, 42, 146–164. [Google Scholar] [CrossRef] [PubMed]
- Hou, K.; Wu, Z.X.; Chen, X.Y.; Wang, J.Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in Health and Diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353.e21. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- De Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut Microbiome and Health: Mechanistic Insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, J.; Wang, L. Role and Mechanism of Gut Microbiota in Human Disease. Front. Cell. Infect. Microbiol. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Xu, T.; Wu, X.; Liu, J.; Sun, J.; Wang, X.; Fan, G.; Meng, X.; Zhang, J.; Zhang, Y. The Regulatory Roles of Dietary Fibers on Host Health via Gut Microbiota-Derived Short Chain Fatty Acids. Curr. Opin. Pharmacol. 2022, 62, 36–42. [Google Scholar] [CrossRef]
- Wei, J.; Xiang, L.; Li, X.; Song, Y.; Yang, C.; Ji, F.; Chung, A.C.K.; Li, K.; Lin, Z.; Cai, Z. Derivatization Strategy Combined with Parallel Reaction Monitoring for the Characterization of Short-Chain Fatty Acids and Their Hydroxylated Derivatives in Mouse. Anal. Chim. Acta 2020, 1100, 66–74. [Google Scholar] [CrossRef]
- Warne, R.W.; Kirschman, L.; Zeglin, L. Manipulation of Gut Microbiota during Critical Developmental Windows Affects Host Physiological Performance and Disease Susceptibility across Ontogeny. J. Anim. Ecol. 2019, 88, 845–856. [Google Scholar] [CrossRef]
- Jana, U.K.; Kango, N.; Pletschke, B. Hemicellulose-Derived Oligosaccharides: Emerging Prebiotics in Disease Alleviation. Front. Nutr. 2021, 8, 1–13. [Google Scholar] [CrossRef]
- Kolodziejczyk, A.A.; Zheng, D.; Elinav, E. Diet–Microbiota Interactions and Personalized Nutrition. Nat. Rev. Microbiol. 2019, 17, 742–753. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, G.; Arya, S.K. Mannans: An Overview of Properties and Application in Food Products. Int. J. Biol. Macromol. 2018, 119, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Price, J.; Mahurkar, A.; Rahnavard, G.; Crabtree, J.; Orvis, J.; Hall, A.B.; Brady, A.; Creasy, H.H.; McCracken, C.; Giglio, M.G.; et al. Strains, Functions and Dynamics in the Expanded Human Microbiome Project. Nature 2017, 550, 61–66. [Google Scholar] [CrossRef]
- Lindstad, L.J.; Lo, G.; Leivers, S.; Lu, Z.; Michalak, L.; Pereira, G.V.; Røhr, Å.K.; Martens, E.C.; McKee, L.S.; Louis, P.; et al. Human Gut Faecalibacterium prausnitzii Deploys a Highly Efficient Conserved System to Cross-Feed on b-Mannan derived Oligosaccharides. MBio 2021, 12, 1–18. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Wiemann, M.; Stålbrand, H. β-Mannanase BoMan26B from Bacteroides ovatus Produces Mannan-Oligosaccharides with Prebiotic Potential from Galactomannan and Softwood β-Mannans. LWT 2021, 151, 112215. [Google Scholar] [CrossRef]
- La Rosa, S.L.; Leth, M.L.; Michalak, L.; Hansen, M.E.; Pudlo, N.A.; Glowacki, R.; Pereira, G.; Workman, C.T.; Arntzen, M.; Pope, P.B.; et al. The Human Gut Firmicute Roseburia intestinalis Is a Primary Degrader of Dietary β-Mannans. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Bagenholm, V.; Reddy, S.K.; Bouraoui, H.; Morrill, J.; Kulcinskaja, E.; Bahr, C.M.; Aurelius, O.; Rogers, T.; Xiao, Y.; Logan, D.T.; et al. Galactomannan Catabolism Conferred by a Polysaccharide Utilization Locus of Bacteroides ovatus: Enzyme Synergy and Crystal Structure of a β-Mannanase. J. Biol. Chem. 2017, 292, 229–243. [Google Scholar] [CrossRef]
- Bågenholm, V.; Wiemann, M.; Reddy, S.K.; Bhattacharya, A.; Rosengren, A.; Logan, D.T.; Stålbrand, H. A Surface-Exposed GH26 -Mannanase from Bacteroides ovatus: Structure, Role, and Phylogenetic Analysis of BoMan26B. J. Biol. Chem. 2019, 294, 9100–9117. [Google Scholar] [CrossRef]
- Senoura, T.; Ito, S.; Taguchi, H.; Higa, M.; Hamada, S.; Matsui, H.; Ozawa, T.; Jin, S.; Watanabe, J.; Wasaki, J.; et al. New Microbial Mannan Catabolic Pathway That Involves a Novel Mannosylglucose Phosphorylase. Biochem. Biophys. Res. Commun. 2011, 408, 701–706. [Google Scholar] [CrossRef]
- Tamura, K.; Brumer, H. Glycan Utilization Systems in the Human Gut Microbiota: A Gold Mine for Structural Discoveries. Curr. Opin. Struct. Biol. 2021, 68, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Briggs, J.A.; Grondin, J.M.; Brumer, H. Communal Living: Glycan Utilization by the Human Gut Microbiota. Environ. Microbiol. 2021, 23, 15–35. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.A.; Kuipers, B.; Atashgahi, S.; Aalvink, S.; Smidt, H.; de Vos, W.M. Inter-Species Metabolic Interactions in an In-Vitro Minimal Human Gut Microbiome of Core Bacteria. NPJ Biofilms Microbiomes 2022, 8, 1–13. [Google Scholar] [CrossRef]
- Ejby, M.; Guskov, A.; Pichler, M.J.; Zanten, G.C.; Schoof, E.; Saburi, W.; Slotboom, D.J.; Abou Hachem, M. Two Binding Proteins of the ABC Transporter That Confers Growth of Bifidobacterium animalis subsp. lactis ATCC27673 on β-Mannan Possess Distinct Manno-Oligosaccharide-Binding Profiles. Mol. Microbiol. 2019, 112, 114–130. [Google Scholar] [CrossRef] [PubMed]
- Coyte, K.Z.; Rakoff-Nahoum, S. Understanding Competition and Cooperation within the Mammalian Gut Microbiome. Curr. Biol. 2019, 29, R538–R544. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.M.; Mulder, I.E.; Travis, A.J.; Lan, A.; Cerf-Bensussan, N.; Gaboriau-Routhiau, V.; Garden, K.; Logan, E.; Delday, M.I.; Coutts, A.G.P.; et al. Human Gut Symbiont Roseburia hominis Promotes and Regulates Innate Immunity. Front. Immunol. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Guo, Y.; Xie, J.P.; Deng, K.; Li, X.; Yuan, Y.; Xuan, Q.; Xie, J.; He, X.M.; Wang, Q.; Li, J.J.; et al. Prophylactic Effects of Bifidobacterium adolescentis on Anxiety and Depression-like Phenotypes after Chronic Stress: A Role of the Gut Microbiota-Inflammation Axis. Front. Behav. Neurosci. 2019, 13, 126. [Google Scholar] [CrossRef]
- Rivière, A.; Gagnon, M.; Weckx, S.; Roy, D.; Vuyst, L. De Mutual Cross-Feeding Interactions between Bifidobacterium longum subsp. longum NCC2705 and Eubacterium rectale ATCC 33656. Appl. Environ. Microbiol. 2015, 81, 7767–7781. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Gueimonde, M.; Duncan, S.H.; Flint, H.J.; De Los Reyes-Gavilan, C.G. Enhanced Butyrate Formation by Cross-Feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS Microbiol. Lett. 2015, 362, 1–7. [Google Scholar] [CrossRef]
- Ghosh, S.; Pramanik, S. Structural Diversity, Functional Aspects and Future Therapeutic Applications of Human Gut Microbiome. Arch. Microbiol. 2021, 203, 5281–5308. [Google Scholar] [CrossRef]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef]
- Gasteiger, E. ExPASy: The Proteomics Server for in-Depth Protein Knowledge and Analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Almagro Armenteros, J.J.; Johansen, A.R.; Gíslason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 Predicts All Five Types of Signal Peptides Using Protein Language Models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B.; et al. MolProbity: More and Better Reference Data for Improved All-Atom Structure Validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A Program to Check the Stereochemical Quality of Protein Structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New Capabilities and Interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A Method for the Absolute Quantification of CDNA Using Real-Time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Byrne, E.; Björkmalm, J.; Bostick, J.P.; Sreenivas, K.; Willquist, K.; van Niel, E.W.J. Characterization and Adaptation of Caldicellulosiruptor Strains to Higher Sugar Concentrations, Targeting Enhanced Hydrogen Production from Lignocellulosic Hydrolysates. Biotechnol. Biofuels 2021, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Butler, S.; Al-Rudainy, B.; Wallberg, O.; Stålbrand, H. Enzymatic Conversion of Different Qualities of Refined Softwood Hemicellulose Recovered from Spent Sulfite Liquor. Molecules 2022, 27, 3207. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Ruthes, A.; Vilaplana, F.; Karlsson, E.N.; Adlecreutz, P.; Stålbrand, H. Enzyme Synergy for the Production of Arabinoxylo-Oligosaccharides from Highly Substituted Arabinoxylan and Evaluation of Their Prebiotic Potential. LWT 2020, 131, 109762. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of Stable Housekeeping Genes, Differentially Regulated Target Genes and Sample Integrity: BestKeeper–Excel-Based Tool Using Pair-Wise Correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of Housekeeping Genes for Gene Expression Studies in Human Reticulocytes Using Real-Time PCR. BMC Mol. Biol. 2006, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. MiRDeepFinder: A MiRNA Analysis Tool for Deep Sequencing of Plant Small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Pfaffl, M.W. Quantification Strategies in Real-Time Polymerase Chain Reaction. In Polymerase Chain Reaction: Theory and Technology; Caister Academic Press: Norfolk, UK, 2019. [Google Scholar]
- Morrill, J.; Kulcinskaja, E.; Sulewska, A.M.; Lahtinen, S.; Stålbrand, H. Bifidobacterium animalis subsp. lactis Bl-04 Possesses a Low-Affinity Mannan-Binding Module and Highlights the Diversity of Mannanolytic Enzymes. BMC Biochem. 2015, 16, 1–12. [Google Scholar] [CrossRef]
- Ladevéze, S.; Tarquis, L.; Cecchini, D.A.; Bercovici, J.; André, I.; Topham, C.M.; Morel, S.; Laville, E.; Monsan, P.; Lombard, V.; et al. Role of Glycoside Phosphorylases in Mannose Foraging by Human Gut Bacteria. J. Biol. Chem. 2013, 288, 32370–32383. [Google Scholar] [CrossRef]
- Li, A.; Benkoulouche, M.; Ladeveze, S.; Durand, J.; Cioci, G.; Laville, E.; Potocki-Veronese, G. Discovery and Biotechnological Exploitation of Glycoside-Phosphorylases. Int. J. Mol. Sci. 2022, 23, 3043. [Google Scholar] [CrossRef]
- Reddy, S.K.; Bågenholm, V.; Pudlo, N.A.; Bouraoui, H.; Koropatkin, N.M.; Martens, E.C.; Stålbrand, H. A β-Mannan Utilization Locus in Bacteroides ovatus Involves a GH36 α-Galactosidase Active on Galactomannans. FEBS Lett. 2016, 590, 2106–2118. [Google Scholar] [CrossRef]
- Grimaud, F.; Pizzut-Serin, S.; Tarquis, L.; Ladevèze, S.; Morel, S.; Putaux, J.L.; Potocki-Veronese, G. In Vitro Synthesis and Crystallization of β-1,4-Mannan. Biomacromolecules 2019, 20, 846–853. [Google Scholar] [CrossRef] [PubMed]
- You, X.; Qin, Z.; Yan, Q.; Yang, S.; Li, Y.; Jiang, Z. Structural Insights into the Catalytic Mechanism of a Novel Glycoside Hydrolase Family 113 -1,4-Mannanase from Amphibacillus xylanus. J. Biol. Chem. 2018, 293, 11746–11757. [Google Scholar] [CrossRef] [PubMed]
- Merceron, R.; Foucault, M.; Haser, R.; Mattes, R.; Watzlawick, H.; Gouet, P. The Molecular Mechanism of Thermostable α-Galactosidases AgaA and AgaB Explained by X-Ray Crystallography and Mutational Studies. J. Biol. Chem. 2012, 287, 39642–39652. [Google Scholar] [CrossRef] [PubMed]
- Nakae, S.; Ito, S.; Higa, M.; Senoura, T.; Wasaki, J.; Hijikata, A.; Shionyu, M.; Ito, S.; Shirai, T. Structure of Novel Enzyme in Mannan Biodegradation Process 4-O-β-d-Mannosyl-d-Glucose Phosphorylase MGP. J. Mol. Biol. 2013, 425, 4468–4478. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Saburi, W.; Odaka, R.; Kato, K.; Sakurai, N.; Komoda, K.; Nishimoto, M.; Kitaoka, M.; Mori, H.; Yao, M. Structural Insights into the Difference in Substrate Recognition of Two Mannoside Phosphorylases from Two GH130 Subfamilies. FEBS Lett. 2016, 590, 828–837. [Google Scholar] [CrossRef]
- Zhang, Y.; Ju, J.; Peng, H.; Gao, F.; Zhou, C.; Zeng, Y.; Xue, Y.; Li, Y.; Henrissat, B.; Gao, G.F.; et al. Biochemical and Structural Characterization of the Intracellular Mannanase AaManA of Alicyclobacillus acidocaldarius Reveals a Novel Glycoside Hydrolase Family Belonging to Clan GH-A. J. Biol. Chem. 2008, 283, 31551–31558. [Google Scholar] [CrossRef]
- Couturier, M.; Touvrey-Loiodice, M.; Terrapon, N.; Drula, E.; Buon, L.; Chirat, C.; Henrissat, B.; Helbert, W. Functional Exploration of the Glycoside Hydrolase Family GH113. PLoS ONE 2022, 17, e0267509. [Google Scholar] [CrossRef]
- Linares, C.N.; Dilokpimol, A.; Stålbrand, H.; Mäkelä, M.R.; de Vries, R.P. Recombinant Production and Characterization of Six Novel GH27 and GH36 α-Galactosidases from Penicillium subrubescens and Their Synergism with a Commercial Mannanase during the Hydrolysis of Lignocellulosic Biomass. Bioresour. Technol. 2020, 295, 122258. [Google Scholar] [CrossRef]
- Hirmas, B.; Gasaly, N.; Orellana, G.; Vega-Sagardía, M.; Saa, P.; Gotteland, M.; Garrido, D. Metabolic Modeling and Bidirectional Culturing of Two Gut Microbes Reveal Cross-Feeding Interactions and Protective Effects on Intestinal Cells. mSystems 2022, 7, e00646-22. [Google Scholar] [CrossRef]
- Moens, F.; Verce, M.; De Vuyst, L. Lactate- and Acetate-Based Cross-Feeding Interactions between Selected Strains of Lactobacilli, Bifidobacteria and Colon Bacteria in the Presence of Inulin-Type Fructans. Int. J. Food Microbiol. 2017, 241, 225–236. [Google Scholar] [CrossRef]
- D’Souza, G.; Shitut, S.; Preussger, D.; Yousif, G.; Waschina, S.; Kost, C. Ecology and Evolution of Metabolic Cross-Feeding Interactions in Bacteria. Nat. Prod. Rep. 2018, 35, 455–488. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Specific Activity of RhMOP130A (kat/mol) | ||
|---|---|---|
| Substrate | Synthesis Direction | Phosphorolysis Direction |
| M1 | - | NA |
| M2 | 2.98 ± 0.10 | - |
| M3 | 1.13 ± 0.08 | 1.02 ± 0.13 |
| M4 | 1.16 ± 0.11 | 0.65 ± 0.04 |
| M5 | 1.45 ± 0.08 | 0.58 ± 0.02 |
| GM3 | NA | - |
| G2M5 | NA | - |
| RhMan113A | ||||
|---|---|---|---|---|
| Substrate | Specific Activity (kat/mol) | kcat (s−1) | KM (mM) | kcat/KM (s−1 mM−1) |
| M2 | - | NA | NA | NA |
| M3 | 4.9 ± 0.02 | NA | NA | NA |
| M4 | 10.2 ± 0.2 | 10.2 ± 1.1 | 3.3 ± 1.1 | 3.1 ± 0.4 |
| M5 | 7.0 ± 0.1 | 3.2 ± 0.7 | 2.1 ± 1.8 | 1.5 ± 0.9 |
| M6 | 5.8 ± 0.4 | NA | NA | NA |
| GM3 | - | NA | NA | NA |
| Substrate | Specific Activity of RhGal36A in (kat/mol) |
|---|---|
| pNP-gal | 234.7 ± 2.9 |
| Raffinose | + |
| GM3 | 17.1 ± 0.7 |
| G2M5 | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattacharya, A.; Majtorp, L.; Birgersson, S.; Wiemann, M.; Sreenivas, K.; Verbrugghe, P.; Van Aken, O.; Van Niel, E.W.J.; Stålbrand, H. Cross-Feeding and Enzymatic Catabolism for Mannan-Oligosaccharide Utilization by the Butyrate-Producing Gut Bacterium Roseburia hominis A2-183. Microorganisms 2022, 10, 2496. https://doi.org/10.3390/microorganisms10122496
Bhattacharya A, Majtorp L, Birgersson S, Wiemann M, Sreenivas K, Verbrugghe P, Van Aken O, Van Niel EWJ, Stålbrand H. Cross-Feeding and Enzymatic Catabolism for Mannan-Oligosaccharide Utilization by the Butyrate-Producing Gut Bacterium Roseburia hominis A2-183. Microorganisms. 2022; 10(12):2496. https://doi.org/10.3390/microorganisms10122496
Chicago/Turabian StyleBhattacharya, Abhishek, Lovisa Majtorp, Simon Birgersson, Mathias Wiemann, Krishnan Sreenivas, Phebe Verbrugghe, Olivier Van Aken, Ed W. J. Van Niel, and Henrik Stålbrand. 2022. "Cross-Feeding and Enzymatic Catabolism for Mannan-Oligosaccharide Utilization by the Butyrate-Producing Gut Bacterium Roseburia hominis A2-183" Microorganisms 10, no. 12: 2496. https://doi.org/10.3390/microorganisms10122496