
Biochemical Evaluation of Agaricus and Pleurotus Strains in Batch Cultures for Production Optimization of Valuable Metabolites


Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material, Media and Culture Conditions
2.2. Biomass Collection and Determination
2.3. Determination of Reducing Sugar Consumption
2.4. Determination of Intracellular Polysaccharides (IPS)
2.5. Determination of Proteins
2.6. Determination of RNA Levels
2.7. Determination of Total Glucans
2.8. Determination of Glucanase Activity
2.9. Determination of Growth Rates
2.10. Optimization Analysis
2.11. Statistical Analysis
3. Results and Discussion
3.1. Biomass Production–Glucose Consumption of the Substrate
3.2. IPS Determination
3.3. Protein Production
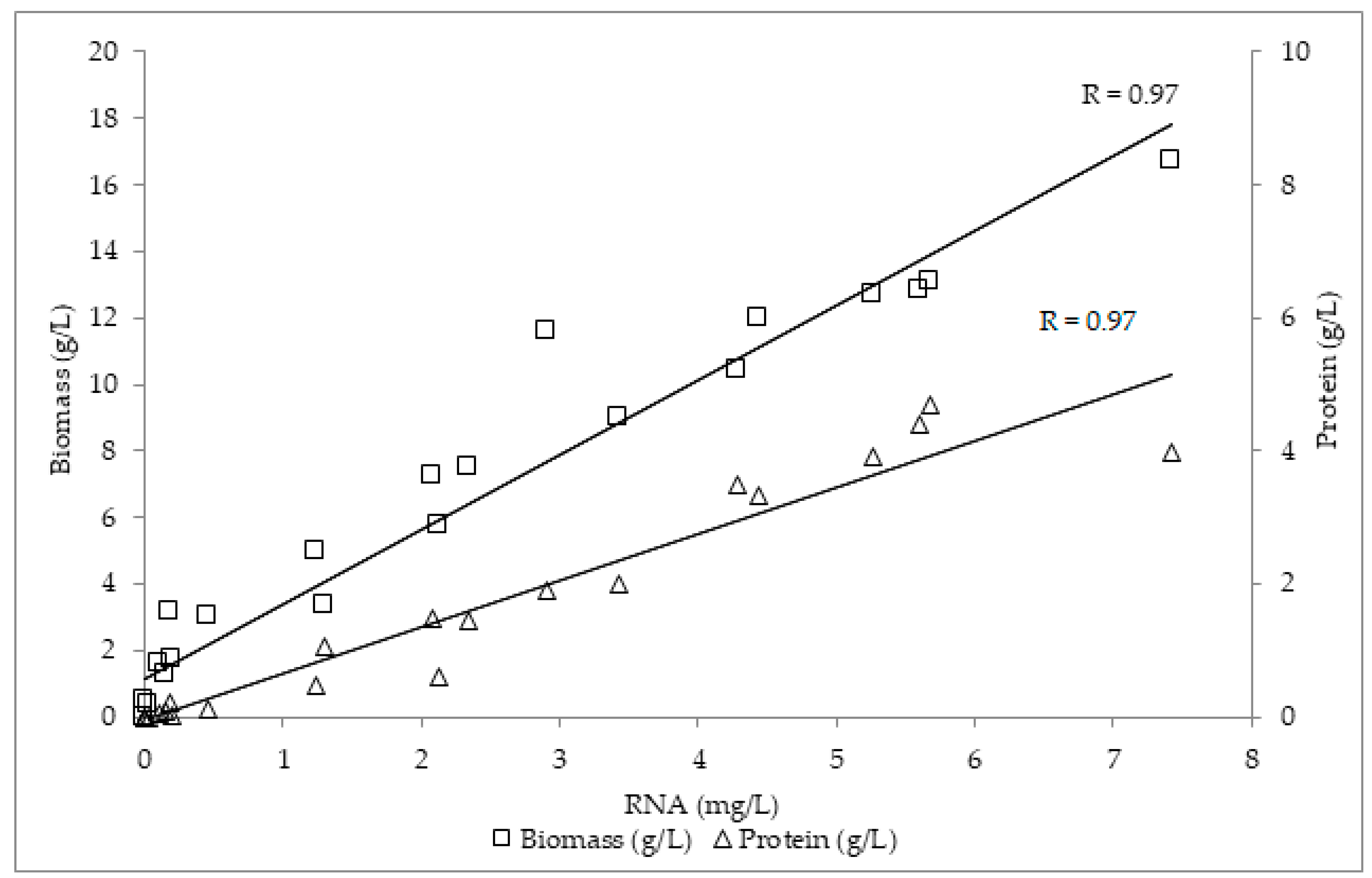
3.4. RNA Determination
3.5. Glucan Measurement
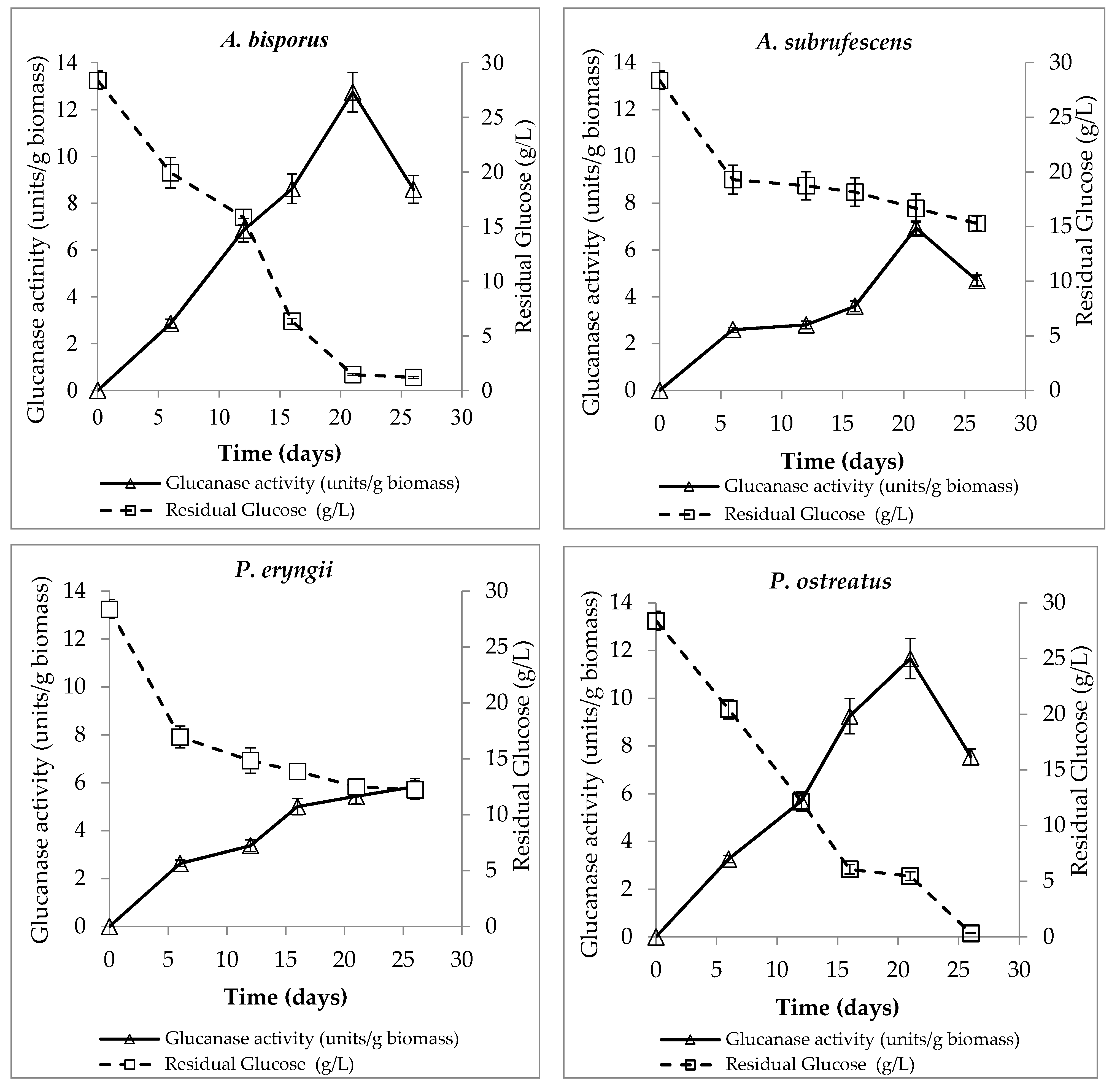
3.6. Glucanase Activity Measurement
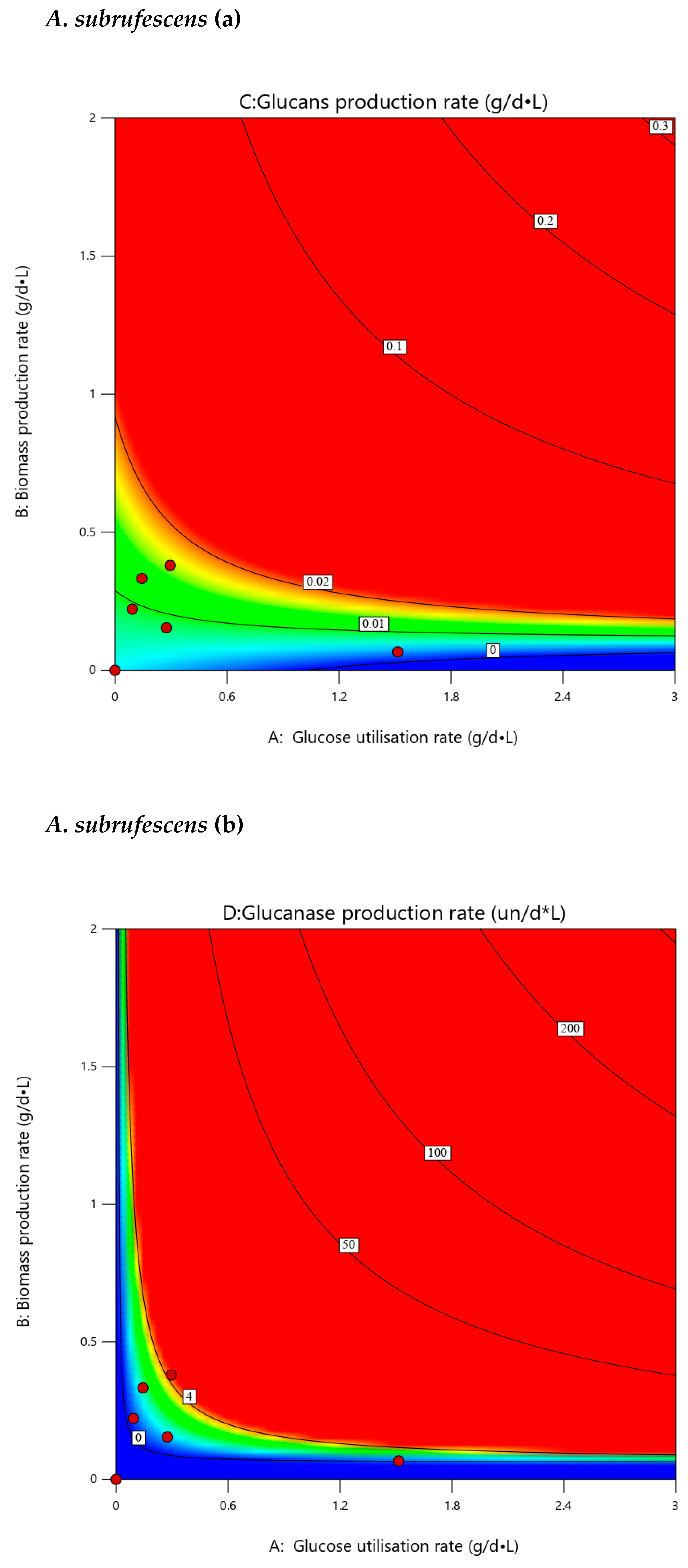
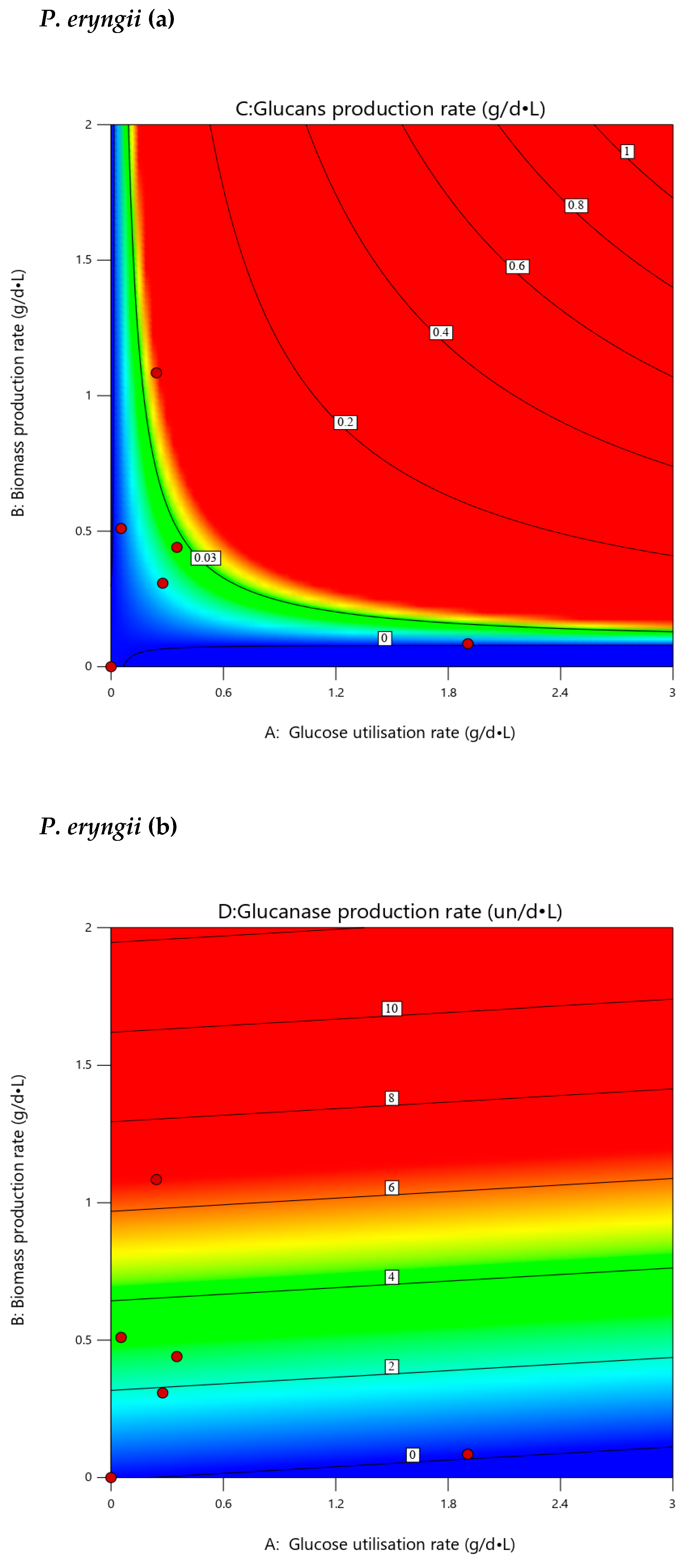
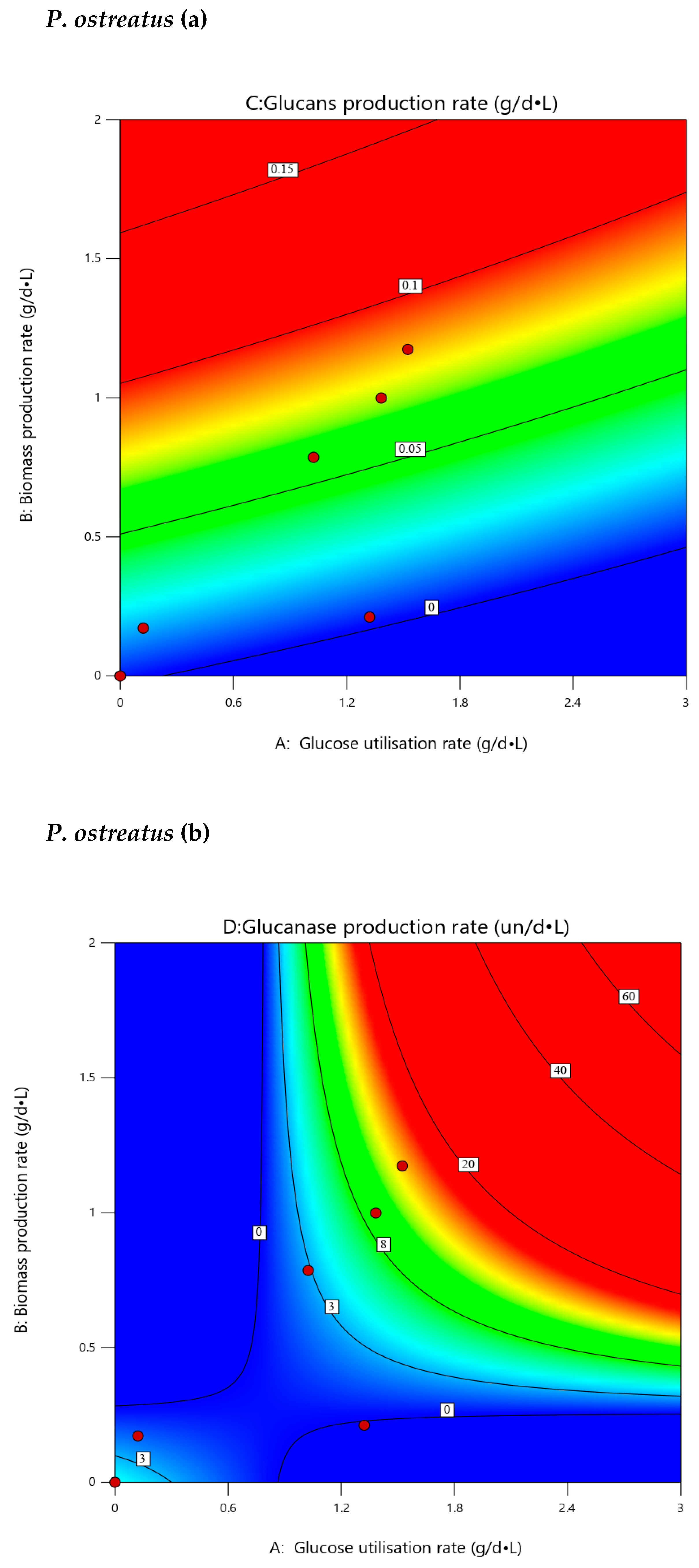
3.7. Extrapolated Optimization
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, K.; Mehra, R.; Guiné, R.P.F.; Lima, M.J.; Kumar, N.; Kaushik, R.; Ahmed, N.; Yadav, A.N.; Kumar, H. Edible Mushrooms: A comprehensive review on bioactive compounds with health benefits and processing aspects. Foods 2021, 10, 2996. [Google Scholar] [CrossRef]
- Karunarathna, S.C.; Chen, J.; Mortimer, P.E.; Xu, J.C.; Zhao, R.L.; Callac, P.; Hyde, K.D. A review of genus Agaricus in tropical and humid subtropical regions of Asia. Mycosphere 2016, 7, 417–439. [Google Scholar] [CrossRef]
- Chen, J.; Callac, P.; Parra, L.A.; Karunarathna, S.C.; He, M.Q.; Moinard, M.; De Kesel, A.; Raspé, O.; Wisitrassameewong, K.; Hyde, K.D.; et al. Study in Agaricus subgenus minores and allied clades reveals a new American subgenus and contrasting phylogenetic patterns in Europe and Greater Mekong Subregion. Persoonia 2017, 38, 170–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, J.; Weber, R. Introduction to Fungi, 3rd ed.; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Volman, J.J.; Helsper, J.P.; Wei, S.; Baars, J.J.; van Griensven, L.J.; Sonnenberg, A.S.; Mensink, R.P.; Plat, J. Effects of mushroom-derived beta-glucan-rich polysaccharide extracts on nitric oxide production by bone marrow-derived macrophages and nuclear factor-kappa B transactivation in Caco-2 reporter cells: Can effects be explained by structure? Mol. Nutr. Food Res. 2010, 54, 268–276. [Google Scholar]
- Beelman, R.B.; Royse, D.; Chikthimmah, N. Bioactive components in button mushroom Agaricus bisporus (J. Lge) Imbach (Agaricomycetideae) of nutritional, medicinal or biological importance. Int. J. Med. Mushr. 2003, 5, 321–337. [Google Scholar] [CrossRef]
- Chang, S.T.; Miles, P.G. Mushrooms: Cultivation, Nutritional Value, Medicinal Effect and Environmental Impact, 2nd ed.; CRC Press LLC: Boca Raton, FL, USA, 2004. [Google Scholar]
- Kalač, P. Chemical composition and nutritional value of European species of wild growing mushrooms: A review. Food Chem. 2009, 113, 9–16. [Google Scholar] [CrossRef]
- Kerrigan, R.W. Agaricus subrufescens, a cultivated edible and medicinal mushroom, and its synonyms. Mycologia 2005, 97, 12–24. [Google Scholar] [CrossRef]
- Mizuno, T.; Morimoto, M.; Minato, K.; Tsuchida, H. Polysaccharides from Agaricus blazei stimulate lymphocyte T-cell subsects in mice. Biosci. Biotechnol. Biochem. 1998, 62, 434–437. [Google Scholar] [CrossRef] [Green Version]
- Takaku, T.; Kimura, Y.; Okuda, H. Isolation of an Antitumor Compound from Agaricus blazei Murill and its mechanism of action. J. Nutr. 2001, 131, 1409–1413. [Google Scholar] [CrossRef]
- Morales, D.; Tejedor-Calvo, E.; Jurado-Chivato, N.; Polo, G.; Tabernero, M.; Ruiz-Rodríguez, A.; Largo, C.; Soler-Rivas, C. In vitro and in vivo testing of the hypocholesterolemic activity of ergosterol- and β-glucan-enriched extracts obtained from shiitake mushrooms (Lentinula edodes). Food Funct. 2019, 10, 7325–7332. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Bian, Z.; Xu, B. Beta-glucans from edible and medicinal mushrooms: Characteristics, physicochemical and biological activities. J. Food Compos. Anal. 2015, 41, 165–173. [Google Scholar] [CrossRef]
- Novak, M.; Vetvicka, V. β-glucans, history, and the present: Immunomodulatory aspects and mechanisms of action. J. Immunotoxicol. 2008, 5, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Lavelli, V.; Proserpio, C.; Gallotti, F.; Laureati, M.; Pagliarini, E. Circular reuse of bioresources: The role of Pleurotus spp. in the development of functional foods. Food Fuction. 2018, 9, 1353–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitra, K.; Venkatesh, R.; Dhanalakshmi, K.; Sharavanan, P.T.; Sasikumar, C.B.; Vijayakumari, K.K. Production and economic analysis of oyster mushroom (Pleurotus florida). Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 379–383. [Google Scholar] [CrossRef]
- Kues, U.; Liu, Y. Fruiting body production in basidiomycetes. Appl. Microbiol. Biotechnol. 2000, 54, 141–152. [Google Scholar] [CrossRef]
- Bergendiova, K.; Tibenska, E.; Majtan, J. Pleuran (β-glucan from Pleurotus ostreatus) supplementation, cellular immune response and respiratory tract infections in athletes. Eur. J. Appl. Physiol. 2011, 111, 2033–2040. [Google Scholar] [CrossRef]
- Synytsya, A.; Míčková, K.; Synytsya, A.; Jablonsky, I.; Spěváček, I.; Erban, V.; Kováříková, E.; Čopíková, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Li, S.; Shah, N.P. Characterization, antioxidative and bifidogenic effects of polysaccharides from Pleurotus eryngii after heat treatments. Food Chem. 2016, 197, 240–249. [Google Scholar] [CrossRef]
- Inácio, F.D.; Ferreira, R.O.; de Araujo, C.A.; Brugnari, T.; Castoldi, R.; Peralta, R.M.; de Souza, C.G. Proteases of wood rot fungi with emphasis on the genus Pleurotus. BioMed. Res. Int. 2015, 290161. [Google Scholar]
- Petre, M. Mushroom Biotechnology—Developments and Applications; Elsevier Inc.: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Eyal, J. Mushroom mycelium grown in submerged culture—Potential food applications. In Biotechnology and Food Ingredients; Goldberg, I.W., Ed.; Van Nostrand Reinhold: New York, NY, USA, 1991; pp. 31–64. [Google Scholar]
- Bakratsas, G.; Polydera, A.; Katapodis, P.; Stamatis, H. Recent trends in submerged cultivation of mushrooms and their application as a source of nutraceuticals and food additives. Future Foods 2021, 4, 100086. [Google Scholar] [CrossRef]
- Márquez-rocha, F.J.; Guillén, G.K.; Sánchez, J.E.; Vazquez-Duhalt, R. Growth characteristics of Pleurotus ostreatus in bioreactor. Biotechnol. Technol. 1999, 13, 29–32. [Google Scholar] [CrossRef]
- Lee, B.C.; Bae, J.T.; Pyo, H.B.; Choe, T.B.; Kim, S.W.; Hwang, H.J.; Yun, J.W. Submerged culture conditions for the production of mycelial biomass and exopolysaccharides by the edible Basidiomycete Grifola frondosa. Enzym. Microb. Technol. 2004, 35, 369–376. [Google Scholar] [CrossRef]
- Kim, S.W.; Hwang, H.J.; Park, J.P.; Cho, Y.J.; Song, C.H.; Yun, J.W. Mycelial growth and exo-biopolymer production by submerged culture of various edible mushrooms under different media. Lett. Appl. Microbiol. 2002, 34, 56–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elisashvili, V.I.; Kachlishvili, E.T.; Wasser, S.P. Carbon and nitrogen source effects on basidiomycetes exopolysaccharide production. Priki. Biokhim. Mikrobiol. 2009, 45, 592–596. [Google Scholar] [CrossRef]
- Hamedi, A.; Ghanati, F.; Vahidi, H. Study on the effects of different culture conditions on the morphology of Agaricus blazei and the relationship between morphology and biomass or EPS production. Ann. Microbiol. 2012, 62, 699–707. [Google Scholar] [CrossRef]
- Zou, X. Fed-batch fermentation for hyperproduction of polysaccharide and ergosterol by medicinal mushroom Agaricus brasiliensis. Proc. Biochem. 2006, 41, 970–974. [Google Scholar] [CrossRef]
- Ma, Y.; Guan, C.Y.; Meng, X.J. Biological characteristics for mycelial growth of Agaricus bisporus. Appl. Mech. Mater. 2014, 508, 297–302. [Google Scholar] [CrossRef]
- Gaspar Júnior, P.J.; Tomizawa, M.M.; Schwan, R.F.; Rinker, D.L.; Dias, E.S. Nutritional requirements for growth of Agaricus brasiliensis. Acta Sci. Biol. Sci. 2011, 33, 93–97. [Google Scholar]
- Kim, H.H.; Na, J.G.; Chang, Y.K.; Chun, G.T.; Lee, S.; Jeong, Y.H. Optimization of submerged culture conditions for mycelial growth and exopolysaccharides production by Agaricus blazei. J. Microbiol. Biotechnol. 2004, 14, 944–951. [Google Scholar]
- Gern, R.M.M.; Wisbeck, E.; Rampinelli, R.J.; Ninow, J.L.; Furlan, S.A. Alternative medium for production of Pleurotus ostreatus biomass and potential antitumor polysaccharides. BioRes. Technol. 2008, 99, 76–82. [Google Scholar] [CrossRef]
- Singdevsachan, S.K.; Auroshree, P.; Mishra, J.; Baliyarsingh, B.; Tayung, K.; Thatoi, H. Mushroom polysaccharides as potential prebiotics with their antitumor and immunomodulating properties: A review. Bioact. Carbohydr. Diet. Fibre 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Diamantopoulou, P.; Papanikolaou, S.; Komaitis, M.; Aggelis, G.; Philippoussis, A. Patterns of major metabolites biosynthesis by different mushroom fungi grown on glucose-based submerged cultures. Bioprocess. Biosyst. Eng. 2014, 3, 1385–1400. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, H.; Chen, M.; Kan, S.; Shieh, C. Quantitative analysis of LED effects on exopolysaccharide and mycelial growth productions in Pleurotus eryngii cultures. J. Soc. Chem. Ind. 2012, 88, 1841–1846. [Google Scholar]
- Ahlborn, J.; Stephan, A.; Meckel, T.; Maheshwari, G.; Rühl, M.; Zorn, H. Upcycling of food industry side streams by basidiomycetes for production of a vegan protein source. Int. J. Recycl. Org. Waste Agric. 2019, 8, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Mumpuni, A.; Ekowati, N.; Purnomowati, P.; Purwati, E.S. Growth and protein content establishment of Pleurotus ostreatus on liquid and solid medium. Biosaintifika J. Biol. Biol. Educ. 2017, 9, 572–578. [Google Scholar] [CrossRef] [Green Version]
- Kurbanoglu, E.B.; Algur, O.F.; Zulkadir, A. Submerged production of edible mushroom Agaricus bisporus mycelium in ram horn hydrolysate. Ind. Crop. Prod. 2004, 19, 225–230. [Google Scholar] [CrossRef]
- Usman, M.; Murtaza, G.; Ditta, A. Nutritional, medicinal, and cosmetic value of bioactive compounds in button mushroom (Agaricus bisporus): A Review. Appl. Sci. 2021, 11, 5943. [Google Scholar] [CrossRef]
- Kim, K.; Choi, B.; Lee, I.; Lee, H.; Kwon, S.; Oh, K.; Kim, A.Y. Bioproduction of mushroom mycelium of Agaricus bisporus by commercial submerged fermentation for the production of meat analogue. J. Sci. Food Agric. 2011, 91, 1561–1568. [Google Scholar] [CrossRef]
- Elisashvili, V.; Penninckx, M.; Kachlishvili, E.; Tsiklauri, N.; Metreveli, E.; Kharziani, T.; Kvesitadze, G. Lentinus edodes and Pleurotus species lignocellulolytic enzymes activity in submerged and solid-state fermentation of lignocellulosic wastes of different composition. Bioresour. Technol. 2008, 99, 457–462. [Google Scholar] [CrossRef]
- Bettin, F.; Cousseau, F.; Martins, K.; Boff, N.A.; Zaccaria, S.; da Moura, S.M.; Dillon, P.; Dillon, A.J.P. Phenol removal by laccases and other phenol oxidases of Pleurotus sajor caju PS-2001 in submerged cultivations and aqueous mixtures. J. Env. Manag. 2019, 236, 581–590. [Google Scholar] [CrossRef]
- Bedade, D.K.; Singhal, R.S.; Turunen, O.; Deska, J.; Shamekh, S. Biochemical characterization of extracellular cellulase from Tuber maculatum mycelium produced under submerged fermentation. Appl. Biochem. Biotechnol 2017, 181, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Okereke, O.E.; Akanya, H.O.; Egwim, E.C. Purification and characterization of an acidophilic cellulase from Pleurotus ostreatus and its potential for agrowastes valorization. Biocatal. Agric. Biotechnol. 2017, 12, 253–259. [Google Scholar] [CrossRef]
- Bedade, D.; Berezina, O.; Singhal, R.; Deska, J.; Shamekh, S. Extracellular xylanase production from a new xylanase producer Tuber maculatum mycelium under submerged fermentation and its characterization. Biocatal. Agric. Biotechnol. 2017, 11, 288–293. [Google Scholar] [CrossRef]
- Bentil, J.A.; Thygesen, A.; Mensah, M.; Lange, L.; Meyer, A.S. Cellulase production by white-rot basidiomycetous fungi: Solid-state versus submerged cultivation. Appl. Microbiol. Biotechnol. 2018, 102, 5827–5839. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Bajar, S.; Devi, A.; Pant, D. An overview on the recent developments in fungal cellulase production and their industrial applications. Bioresour. Technol. Rep. 2021, 14, 100652. [Google Scholar] [CrossRef]
- Li, J.; Liu, X.; Yin, Z.; Hu, Z.; Zhang, K.-Q. An Overview on identification and regulatory mechanisms of long non-coding RNAs in Fungi. Front. Microbiol. 2021, 12, 638617. [Google Scholar] [CrossRef]
- García-Martínez, J.; Troulé, K.; Chávez, S.; Pérez-Ortín, J.E. Growth rate controls mRNA turnover in steady and non-steady states. RNA Biol. 2016, 13, 1175–1181. [Google Scholar] [CrossRef] [Green Version]
- Giordano, M.; Palmucci, M.; Raven, J.A. Growth rate hypothesis and efficiency of protein synthesis under different sulphate concentrations in two green algae. Plant Cell. Env. 2015, 38, 2313–2317. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rio, D.C.; Ares, M., Jr.; Hannon, G.J.; Nilsen, T.W. Nondenaturing agarose gel electrophoresis of RNA. Cold Spring Harb. Protoc. 2010, 2010. [Google Scholar] [CrossRef] [PubMed]
- McCleary, B.V.; Draga, A. Measurement of β-glucan in mushrooms and mycelial products. J. AOAC Int. 2016, 99, 364–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levasseur, M.; Thompson, P.A.; Harrison, P.J. Physiological acclimation of marine phytoplankton to different nitrogen sources. J. Phycol. 1993, 29, 587–595. [Google Scholar] [CrossRef]
- Ahirwar, S.; Soni, H.; Rawat, H.K.; Prajapati, B.P.; Kango, N. Experimental design of response surface methodology used for utilisation of palm kernel cake as solid substrate for optimised production of fungal mannanase. Mycology 2016, 7, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Diamantopoulou, P.; Papanikolaou, S.; Kapoti, M.; Komaitis, M.; Aggelis, G.; Philippoussis, A. Mushroom polysaccharides and lipids synthesized in liquid agitated and static cultures. Part I: Screening various mushroom species. Appl. Biochem. Biotechnol. 2012, 167, 536–551. [Google Scholar] [CrossRef]
- Vamvakaki, A.N.; Kandarakis, I.; Kaminarides, S.; Komaitis, M.; Papanikolaou, S. Cheese whey as a renewable substrate for microbial lipid and biomass production by Zygomycetes. Eng. Life Sci. 2010, 10, 348–360. [Google Scholar] [CrossRef]
- Sarris, D.; Philippoussis, A.; Mallouchos, A.; Diamantopoulou, P. Valorization of low-cost, carbon-rich substrates by edible ascomycetes and basidiomycetes grown on liquid cultures. FEMS Microbiol. Lett. 2020, 367, fnaa168. [Google Scholar] [CrossRef]
- Lima, L.F.O.; Habu, S.; Gern, J.C.; Nascimento, B.M.; Parada, J.L.; Noseda, M.D. Production and characterization of the exopolysaccharides produced by Agaricus brasiliensis in submerged fermentation. Appl. Biochem. Biotechnol. 2008, 151, 283–294. [Google Scholar] [CrossRef]
- Liu, G.Q.; Wang, X.L. Selection of a culture medium for reducing costs and enhancing biomass and intracellular polysaccharide production by Agaricus blazei AB2003. Food Technol. Biotechnol. 2009, 47, 210–214. [Google Scholar]
- Özdal, M.; Gülmez, Ö.; Gür Özdal, Ö.; Algur, Ö.F. Antibacterial and antioxidant activity of mycelial extracts of different Pleurotus species. Food Health 2019, 5, 12–18. [Google Scholar] [CrossRef]
- El-Enshasy, H.; Daba, A.; El-Demellawy, M.; Ibrahim, A.; El Sayed, S.; El-Badry, I. Bioprocess development for large scale production of anticancer exo-polysaccharide by Pleurotus ostreatus in submerged culture. J. Appl. Sci. 2010, 10, 2523–2529. [Google Scholar] [CrossRef] [Green Version]
- Krüzselyi, D.; Kovács, D.; Vetter, J. Chemical analysis of king oyster mushroom (Pleurotus eryngii) fruit bodies. Acta Aliment. 2016, 45, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Diamantopoulou, P.; Papanikolaou, S.; Katsarou, E.; Komaitis, M.; Aggelis, G.; Philippoussis, A. Mushroom polysaccharides and lipids synthesized in liquid agitated and static cultures. Part II: Study of Volvariella volvacea. Appl. Biochem. Biotechnol. 2012, 167, 1890–1906. [Google Scholar] [CrossRef] [PubMed]
- Diamantopoulou, P.; Papanikolaou, S.; Aggelis, G.; Philippoussis, A. Adaptation of Volvariella volvacea metabolism in high carbon to nitrogen ratio media. Food Chem. 2016, 196, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-J.; Zhu, L.-L.; Li, D.-S.; Mi, Z.-Y.; Li, H.-M. Significance of inoculation density and carbon source on the mycelial growth and Tuber polysaccharides production by submerged fermentation of Chinese truffle Tuber sinense. Process. Biochem. 2008, 43, 576–586. [Google Scholar] [CrossRef]
- Zhong, J.J.; Tang, Y.-J. Submerged cultivation of medicinal mushrooms for production of valuable bioactive metabolites. Adv. Biochem. Eng. Biotechnol. 2004, 87, 25–59. [Google Scholar] [PubMed]
- Zhou, Y.; Hong-bo, S.; Chen, D. Effects of organic nitrogen and carbon sources on mycelial growth and polysaccharides production and their optimization in the submerged culture of Grifola umbellata, a Chinese medicinal herb. Afr. J. Biotechnol. 2009, 8, 5202–5214. [Google Scholar]
- Li, M.; Yu, L.; Zhao, J.; Zhang, H.; Chen, W.; Zhai, Q.; Tian, F. Role of dietary edible mushrooms in the modulation of gut microbiota. J. Funct. Foods 2021, 83, 104538. [Google Scholar] [CrossRef]
- Hwang, S.J.; Mau, J.L. Quality evaluation of Agaricus bisporus mycelia. Food Sci. 1997, 24, 44–55. [Google Scholar]
- Chang, H.L.; Chao, G.R.; Chen, C.C.; Mau, J.L. Non-volatile taste components of Agaricus blazei, Antrodia camphorata and Cordyceps militaris mycelia. Food Chem. 2001, 74, 203–207. [Google Scholar] [CrossRef]
- Smith, J.E.; Rowan, N.J.; Sullivan, R. Medicinal mushrooms: A rapidly developing area of biotechnology for cancer therapy and other bioactivities. Biotechnol. Lett. 2002, 24, 1839–1845. [Google Scholar] [CrossRef]
- Schumann, U.; Smith, N.A.; Wang, M.B. A fast and efficient method for preparation of high-quality RNA from fungal mycelia. BMC Res. Notes 2013, 6, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuero, R.; Ouellet, T.; Yu, J.; Mogongwa, N. Metal ion enhancement of fungal growth, gene expression and aflatoxin synthesis in Aspergillus flavus: RT-PCR characterization. J. Appl. Microbiol. 2003, 94, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Ohno, N.; Furukawa, M.; Miura, N.N.; Adachi, Y.; Motoi, M.; Yadomae, T. Antitumor β-glucan from the cultured fruit body of Agaricus blazei. Biol. Pharm. Bull. 2001, 24, 820–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitschke, J.; Modick, M.; Busch, E.; von Rekowski, R.W.; Altenbach, H.J.; Mölleken, H. A new colorimetric method to quantify b-1,3-1,6-glucans in comparison with total b-1,3-glucans in edible mushrooms. Food Chem. 2011, 127, 791–796. [Google Scholar] [CrossRef]
- Toledo, R.C.C.; Carvalho, M.A.; Lima, L.C.O.; Vilas-Boas, E.V.B.; Dias, E.S. Measurement of β-glucan and other nutritional characteristics in distinct strains of Agaricus subrufescens mushrooms. Afr. J. Biotechnol. 2013, 12, 6203–6209. [Google Scholar] [CrossRef]
- Mohamad, S.; Awang, M.; Rashid, R.; Ling, L.; Daud, F.; Hamid, A.; Ahmad, R.; Wan Yusoff, W. Optimization of mycelial biomass production in submerged culture fermentation of Pleurotus flabellatus using response surface methodology. Adv. Biosci. Biotechnol. 2015, 6, 419–426. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Species | Time (Days) | Biomass (g/L) | Residual Glucose (g/L) | Yx/Glc (g/g) | IPS (g/L) | Glucans (g/L) | IPS (%, w/w) | Glucans (%, w/w) |
|---|---|---|---|---|---|---|---|---|
| A. bisporus | 0 | 0.00 ± 0.00 * | 28.39 ± 0.85 | - | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| 6 | 1.63 ± 0.11 | 19.93 ± 1.39 | 0.19 ± 0.04 | 0.29 ± 0.02 | 0.05 ± 0.00 | 17.78 ± 1.15 | 3.20 ± 0.21 | |
| 12 | 3.39 ± 0.25 | 15.85 ± 0.71 | 0.27 ± 0.02 | 0.74 ± 0.06 | 0.17 ± 0.01 | 21.91 ± 1.63 | 5.05 ± 0.38 | |
| 16 | 10.44 ± 0.77 | 6.34 ± 0.27 | 0.47 ± 0.03 | 3.43 ± 0.25 | 0.63 ± 0.05 | 32.82 ± 2.41 | 6.08 ± 0.45 | |
| 21 | 13.06 ± 0.87 b,d | 1.45 ± 0.09 | 0.48 ± 0.04 | 3.21 ± 0.21 | 1.05 ± 0.07 | 24.57 ± 1.64 | 8.05 ± 0.54 b,c,d | |
| 26 | 12.67 ± 0.86 | 1.20 ± 0.08 | 0.47 ± 0.02 | 2.57 ± 0.18 | 0.87 ± 0.06 | 20.32 ± 1.38 | 6.87 ± 0.47 | |
| A. subrufescens | 0 | 0.00 ± 0.00 | 28.39 ± 0.85 | - | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| 6 | 0.40 ± 0.02 | 19.30 ± 1.33 | 0.04 ± 0.03 | 0.08 ± 0.00 | 0.01 ± 0.00 | 19.50 ± 0.79 | 2.62 ± 0.11 | |
| 12 | 1.73 ± 0.10 | 18.74 ± 1.29 | 0.18 ± 0.02 | 0.61 ± 0.04 | 0.05 ± 0.00 | 35.29 ± 2.12 | 2.94 ± 0.18 | |
| 16 | 3.06 ± 0.20 | 18.16 ± 1.31 | 0.30 ± 0.03 | 0.70 ± 0.04 | 0.10 ± 0.01 | 22.92 ± 1.47 | 3.12 ± 0.20 | |
| 21 | 4.96 ± 0.21 | 16.68 ± 1.31 | 0.42 ± 0.04 | 1.39 ± 0.06 | 0.16 ± 0.01 | 28.07 ± 1.20 | 3.21 ± 0.14 | |
| 26 | 5.73 ± 0.28 a,c,d | 15.30 ± 0.67 | 0.44 ± 0.04 | 1.32 ± 0.07 | 0.27 ± 0.01 | 25.01 ± 1.24 | 4.70 ± 0.23 a,d | |
| P. eryngii | 0 | 0.00 ± 0.00 | 28.39 ± 0.85 | - | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| 6 | 0.51 ± 0.03 | 16.96 ± 0.97 | 0.04 ± 0.01 | 0.12 ± 0.01 | 0.01 ± 0.00 | 23.53 ± 1.22 | 2.63 ± 0.14 | |
| 12 | 3.15 ± 0.23 | 14.85 ± 1.14 | 0.23 ± 0.02 | 1.17 ± 0.09 | 0.10 ± 0.01 | 37.25 ± 2.73 | 3.31 ± 0.24 | |
| 16 | 7.49 ± 0.51 | 13.88 ± 0.60 | 0.52 ± 0.03 | 2.84 ± 0.19 | 0.30 ± 0.02 | 37.97 ± 2.58 | 4.00 ± 0.27 | |
| 21 | 9.03 ± 0.50 | 12.50 ± 0.53 | 0.57 ± 0.04 | 3.20 ± 0.18 | 0.43 ± 0.02 | 35.42 ± 1.97 | 4.74 ± 0.26 a,d | |
| 26 | 11.58 ± 0.67 b,d | 12.23 ± 0.84 | 0.72 ± 0.05 | 3.82 ± 0.22 | 0.35 ± 0.02 | 32.96 ± 1.91 | 3.03 ± 0.18 | |
| P. ostreatus | 0 | 0.00 ± 0.00 | 28.39 ± 0.85 | - | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| 6 | 1.27 ± 0.06 | 20.46 ± 0.88 | 0.16 ± 0.01 | 0.37 ± 0.02 | 0.04 ± 0.00 | 22.04 ± 1.01 | 3.06 ± 0.14 | |
| 12 | 7.26 ± 0.48 | 12.16 ± 0.91 | 0.45 ± 0.02 | 1.71 ± 0.11 | 0.29 ± 0.02 | 23.61 ± 1.56 | 4.00 ± 0.27 | |
| 16 | 11.96 ± 0.96 | 6.06 ± 0.42 | 0.54 ± 0.02 | 3.32 ± 0.27 | 0.69 ± 0.06 | 27.73 ± 2.22 | 5.81 ± 0.46 | |
| 21 | 12.82 ± 0.93 | 5.45 ± 0.39 | 0.56 ± 0.03 | 2.62 ± 0.19 | 0.79 ± 0.06 | 20.41 ± 1.48 | 6.19 ± 0.45 | |
| 26 | 16.75 ± 0.73 a,b,c | 0.33 ± 0.01 | 0.60 ± 0.04 | 3.56 ± 0.15 | 1.13 ± 0.05 | 21.23 ± 0.93 | 6.73 ± 0.29 a,b,c |
| Fungal Species | Time (Days) | RNA (mg/L) | Protein (g/L) | Glucanase Activity (Units/L) | RNA (mg/g Biomass) | Protein (%, w/w) | Glucanase Activity (Units/g Biomass) |
|---|---|---|---|---|---|---|---|
| A. bisporus | 0 | 0.000 ± 0.000 * | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| 6 | 0.109 ± 0.012 | 0.065 ± 0.004 | 4.668 ± 0.929 | 0.067 ± 0.007 | 4.000 ± 0.260 | 2.862 ± 0.186 | |
| 12 | 1.298 ± 0.119 | 1.055 ± 0.078 | 23.198 ± 2.542 | 0.383 ± 0.035 | 31.119 ± 2.312 | 6.843 ± 0.508 | |
| 16 | 4.280 ± 0.403 | 3.487 ± 0.256 | 89.959 ± 4.158 | 0.410 ± 0.039 | 33.400 ± 2.448 | 8.617 ± 0.632 | |
| 21 | 5.681 ± 0.473 | 4.702 ± 0.314 | 166.439 ± 4.250 | 0.435 ± 0.036 | 36.006 ± 2.402 b,c | 12.744 ± 0.850 b,c | |
| 26 | 5.258 ± 0.446 | 3.921 ± 0.267 | 108.826 ± 4.295 | 0.415 ± 0.035 | 30.947 ± 2.107 | 8.589 ± 0.585 | |
| A. subrufescens | 0 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| 6 | 0.035 ± 0.002 | 0.002 ± 0.000 | 1.037 ± 0.105 | 0.087 ± 0.006 | 0.522 ± 0.021 | 2.593 ± 0.105 | |
| 12 | 0.197 ± 0.019 | 0.041 ± 0.002 | 4.847 ± 0.824 | 0.114 ± 0.011 | 2.342 ± 0.141 | 2.800 ± 0.168 | |
| 16 | 0.459 ± 0.047 | 0.116 ± 0.007 | 11.023 ± 1.092 | 0.150 ± 0.015 | 3.800 ± 0.244 | 3.600 ± 0.231 | |
| 21 | 1.245 ± 0.085 | 0.488 ± 0.021 | 34.425 ± 1.481 | 0.251 ± 0.017 | 9.828 ± 0.420 | 6.938 ± 0.296 a,c,d | |
| 26 | 2.133 ± 0.169 | 0.612 ± 0.030 | 26.944 ± 1.166 | 0.372 ± 0.030 | 10.675 ± 0.529 a,c,d | 4.700 ± 0.233 | |
| P. eryngii | 0 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| 6 | 0.002 ± 0.000 | 0.006 ± 0.000 | 1.343 ± 0.108 | 0.003 ± 0.000 | 1.117 ± 0.058 | 2.633 ± 0.136 | |
| 12 | 0.189 ± 0.023 | 0.218 ± 0.016 | 10.629 ± 1.236 | 0.060 ± 0.007 | 6.909 ± 0.506 | 3.372 ± 0.247 | |
| 16 | 2.344 ± 0.253 | 1.463 ± 0.099 | 37.489 ± 1.699 | 0.313 ± 0.034 | 19.535 ± 1.326 | 5.005 ± 0.340 | |
| 21 | 3.422 ± 0.301 | 2.000 ± 0.111 | 49.111 ± 1.509 | 0.379 ± 0.033 | 22.145 ± 1.229 a,b,d | 5.439 ± 0.302 | |
| 26 | 2.907 ± 0.270 | 1.911 ± 0.111 | 67.646 ± 1.694 | 0.251 ± 0.023 | 16.501 ± 0.957 | 5.842 ± 0.339 a,b,d | |
| P. ostreatus | 0 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 | 0.000 ± 0.000 |
| 6 | 0.145 ± 0.012 | 0.104 ± 0.005 | 4.141 ± 0.748 | 0.114 ± 0.009 | 8.168 ± 0.375 | 3.259 ± 0.150 | |
| 12 | 2.085 ± 0.211 | 1.487 ± 0.098 | 41.352 ± 1.876 | 0.287 ± 0.029 | 20.462 ± 1.348 | 5.692 ± 0.375 | |
| 16 | 4.437 ± 0.444 | 3.318 ± 0.265 | 110.651 ± 3.696 | 0.371 ± 0.037 | 27.742 ± 2.217 | 9.252 ± 0.739 | |
| 21 | 5.602 ± 0.506 | 4.409 ± 0.319 | 149.565 ± 4.217 | 0.437 ± 0.039 | 34.392 ± 2.487 b,d | 11.667 ± 0.843 b,c | |
| 26 | 7.420 ± 0.518 | 3.991 ± 0.174 | 126.433 ± 1.646 | 0.443 ± 0.031 | 23.827 ± 1.039 | 7.548 ± 0.329 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Argyropoulos, D.; Psallida, C.; Sitareniou, P.; Flemetakis, E.; Diamantopoulou, P. Biochemical Evaluation of Agaricus and Pleurotus Strains in Batch Cultures for Production Optimization of Valuable Metabolites. Microorganisms 2022, 10, 964. https://doi.org/10.3390/microorganisms10050964
Argyropoulos D, Psallida C, Sitareniou P, Flemetakis E, Diamantopoulou P. Biochemical Evaluation of Agaricus and Pleurotus Strains in Batch Cultures for Production Optimization of Valuable Metabolites. Microorganisms. 2022; 10(5):964. https://doi.org/10.3390/microorganisms10050964
Chicago/Turabian StyleArgyropoulos, Dimitrios, Charoula Psallida, Paraskevi Sitareniou, Emmanouil Flemetakis, and Panagiota Diamantopoulou. 2022. "Biochemical Evaluation of Agaricus and Pleurotus Strains in Batch Cultures for Production Optimization of Valuable Metabolites" Microorganisms 10, no. 5: 964. https://doi.org/10.3390/microorganisms10050964