
Acne, Microbiome, and Probiotics: The Gut–Skin Axis

 , ,
, ,
Abstract
:1. Introduction
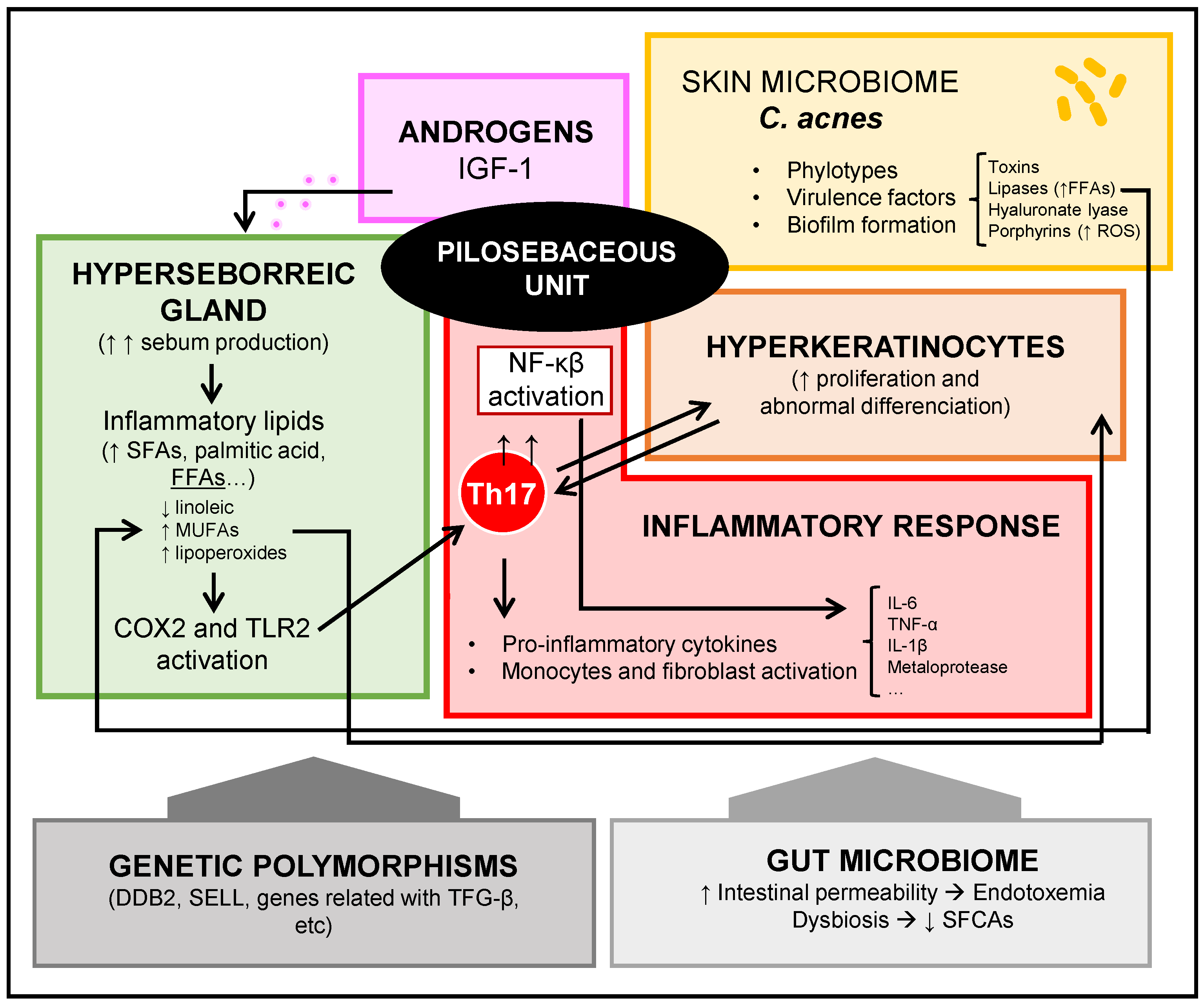
2. Pathogenesis of Acne Vulgaris
3. Acne Vulgaris and Skin Microbiota: Cutibacterium acnes
4. Acne Vulgaris and Gut Microbiota
5. Acne Vulgaris and Probiotics
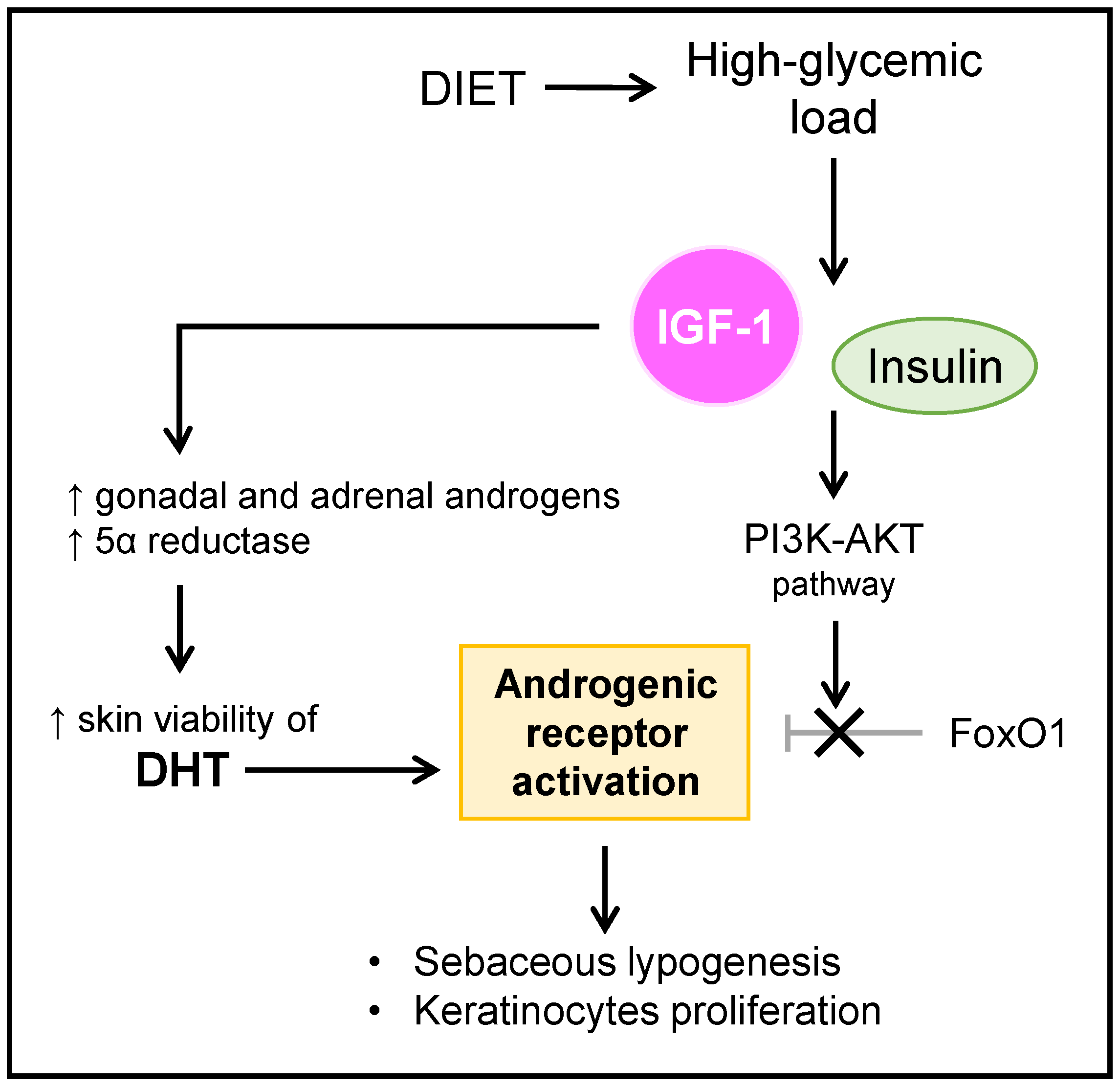
6. Acne Vulgaris and Diet
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Swaney, M.H.; Kalan, L.R. Living in Your Skin: Microbes, Molecules, and Mechanisms. Infect. Immun. 2021, 89, e00695-20. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Byrd, A.L.; Deming, C.; Conlan, S.; Kong, H.H.; Segre, J.A. Biogeography and individuality shape function in the human skin metagenome. Nature 2014, 514, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, E.A.; Nance, K.; Chen, S. The Gut-Brain Axis. Annu. Rev. Med. 2022, 73, 439–453. [Google Scholar] [CrossRef] [PubMed]
- Budden, K.F.; Gellatly, S.L.; Wood, D.L.; Cooper, M.A.; Morrison, M.; Hugenholtz, P.; Hansbro, P.M. Emerging pathogenic links between microbiota and the gut-lung axis. Nat. Rev. Microbiol. 2017, 15, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef] [Green Version]
- De Pessemier, B.; Grine, L.; Debaere, M.; Maes, A.; Paetzold, B.; Callewaert, C. Gut-Skin Axis: Current Knowledge of the Interrelationship between Microbial Dysbiosis and Skin Conditions. Microorganisms 2021, 9, 353. [Google Scholar] [CrossRef]
- Navarro-López, V.; Núñez-Delegido, E.; Ruzafa-Costas, B.; Sánchez-Pellicer, P.; Agüera-Santos, J.; Navarro-Moratalla, L. Probiotics in the Therapeutic Arsenal of Dermatologists. Microorganisms 2021, 9, 1513. [Google Scholar] [CrossRef]
- Sinha, S.; Lin, G.; Ferenczi, K. The skin microbiome and the gut-skin axis. Clin. Dermatol. 2021, 39, 829–839. [Google Scholar] [CrossRef]
- Fu, Y.; Lee, C.H.; Chi, C.C. Association of Psoriasis with Inflammatory Bowel Disease: A Systematic Review and Meta-analysis. JAMA Dermatol. 2018, 154, 1417–1423. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Boscá, A.; Navarro-López, V.; Martínez-Andrés, A.; Such, J.; Francés, R.; Horga de la Parte, J.; Asín-Llorca, M. Identification of Bacterial DNA in the Peripheral Blood of Patients with Active Psoriasis. JAMA Dermatol. 2015, 151, 670–671. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, C.A.; Monteleone, G.; McLaughlin, J.T.; Paus, R. The gut-skin axis in health and disease: A paradigm with therapeutic implications. Bioessays 2016, 38, 1167–1176. [Google Scholar] [CrossRef] [PubMed]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The Gut Microbiome as a Major Regulator of the Gut-Skin Axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heng, A.H.S.; Chew, F.T. Systematic review of the epidemiology of acne vulgaris. Sci. Rep. 2020, 10, 5754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksu, A.E.; Metintas, S.; Saracoglu, Z.N.; Gurel, G.; Sabuncu, I.; Arikan, I.; Kalyoncu, C. Acne: Prevalence and relationship with dietary habits in Eskisehir, Turkey. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Pereira Duquia, R.; da Silva Dos Santos, I.; de Almeida, H., Jr.; Martins Souza, P.R.; de Avelar Breunig, J.; Zouboulis, C.C. Epidemiology of Acne Vulgaris in 18-Year-Old Male Army Conscripts in a South Brazilian City. Dermatology 2017, 233, 145–154. [Google Scholar] [CrossRef]
- Vos, T.; Flaxman, A.D.; Naghavi, M.; Lozano, R.; Michaud, C.; Ezzati, M.; Shibuya, K.; Salomon, J.A.; Abdalla, S.; Aboyans, V.; et al. Years lived with disability (YLDs) for 1160 sequelae of 289 diseases and injuries 1990-2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2163–2196. [Google Scholar] [CrossRef]
- Bhate, K.; Williams, H.C. Epidemiology of acne vulgaris. Br. J. Dermatol. 2013, 168, 474–485. [Google Scholar] [CrossRef]
- Cordain, L.; Lindeberg, S.; Hurtado, M.; Hill, K.; Eaton, S.B.; Brand-Miller, J. Acne vulgaris: A disease of Western civilization. Arch. Dermatol. 2002, 138, 1584–1590. [Google Scholar] [CrossRef] [Green Version]
- Suh, D.H.; Kwon, H.H. What’s new in the physiopathology of acne? Br. J. Dermatol. 2015, 172, 13–19. [Google Scholar] [CrossRef]
- Afshar, M.; Gallo, R.L. Innate immune defense system of the skin. Vet. Dermatol. 2013, 24, e8–e9. [Google Scholar] [CrossRef] [PubMed]
- Moradi Tuchayi, S.; Makrantonaki, E.; Ganceviciene, R.; Dessinioti, C.; Feldman, S.R.; Zouboulis, C.C. Acne vulgaris. Nat. Rev. Dis. Primers 2015, 1, 15029. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Linking diet to acne metabolomics, inflammation, and comedogenesis: An update. Clin. Cosmet. Investig. Dermatol. 2015, 8, 371–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, B.S.; Seo, J.G.; Lee, G.S.; Kim, J.H.; Kim, S.Y.; Han, Y.W.; Kang, H.; Kim, H.O.; Rhee, J.H.; Chung, M.J.; et al. Antimicrobial activity of enterocins from Enterococcus faecalis SL-5 against Propionibacterium acnes, the causative agent in acne vulgaris, and its therapeutic effect. J. Microbiol. 2009, 47, 101–109. [Google Scholar] [CrossRef]
- Jugeau, S.; Tenaud, I.; Knol, A.C.; Jarrousse, V.; Quereux, G.; Khammari, A.; Dreno, B. Induction of toll-like receptors by Propionibacterium acnes. Br. J. Dermatol. 2005, 153, 1105–1113. [Google Scholar] [CrossRef]
- Lwin, S.M.; Kimber, I.; McFadden, J.P. Acne, quorum sensing and danger. Clin. Exp. Dermatol. 2014, 39, 162–167. [Google Scholar] [CrossRef]
- Alestas, T.; Ganceviciene, R.; Fimmel, S.; Müller-Decker, K.; Zouboulis, C.C. Enzymes involved in the biosynthesis of leukotriene B4 and prostaglandin E2 are active in sebaceous glands. J. Mol. Med. 2006, 84, 75–87. [Google Scholar] [CrossRef]
- Snodgrass, R.G.; Huang, S.; Choi, I.W.; Rutledge, J.C.; Hwang, D.H. Inflammasome-mediated secretion of IL-1β in human monocytes through TLR2 activation; modulation by dietary fatty acids. J. Immunol. 2013, 191, 4337–4347. [Google Scholar] [CrossRef] [Green Version]
- Nakatsuji, T.; Kao, M.C.; Zhang, L.; Zouboulis, C.C.; Gallo, R.L.; Huang, C.M. Sebum free fatty acids enhance the innate immune defense of human sebocytes by upregulating beta-defensin-2 expression. J. Investig. Dermatol. 2010, 130, 985–994. [Google Scholar] [CrossRef] [Green Version]
- Legrand-Poels, S.; Esser, N.; L’homme, L.; Scheen, A.; Paquot, N.; Piette, J. Free fatty acids as modulators of the NLRP3 inflammasome in obesity/type 2 diabetes. Biochem. Pharmacol. 2014, 92, 131–141. [Google Scholar] [CrossRef]
- Kistowska, M.; Gehrke, S.; Jankovic, D.; Kerl, K.; Fettelschoss, A.; Feldmeyer, L.; Fenini, G.; Kolios, A.; Navarini, A.; Ganceviciene, R.; et al. IL-1β drives inflammatory responses to propionibacterium acnes in vitro and in vivo. J. Investig. Dermatol. 2014, 134, 677–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zouboulis, C.C.; Schagen, S.; Alestas, T. The sebocyte culture: A model to study the pathophysiology of the sebaceous gland in sebostasis, seborrhoea and acne. Arch. Dermatol. Res. 2008, 300, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Shamloul, G.; Khachemoune, A. An updated review of the sebaceous gland and its role in health and diseases Part 1: Embryology, evolution, structure, and function of sebaceous glands. Dermatol. Ther. 2021, 34, e14695. [Google Scholar] [CrossRef] [PubMed]
- Ottaviani, M.; Camera, E.; Picardo, M. Lipid mediators in acne. Mediat. Inflamm. 2010, 2010, 858176. [Google Scholar] [CrossRef] [Green Version]
- Bienenfeld, A.; Azarchi, S.; Lo Sicco, K.; Marchbein, S.; Shapiro, J.; Nagler, A.R. Androgens in women: Androgen-mediated skin disease and patient evaluation. J. Am. Acad. Dermatol. 2019, 80, 1497–1506. [Google Scholar] [CrossRef]
- Hu, T.; Wei, Z.; Ju, Q.; Chen, W. Sex hormones and acne: State of the art. J. Dtsch. Dermatol. Ges. 2021, 19, 509–515. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Nikolakis, G.; Manna, P.R.; Skobowiat, C.; Zmijewski, M.; Li, W.; Janjetovic, Z.; Postlethwaite, A.; Zouboulis, C.C.; et al. Steroidogenesis in the skin: Implications for local immune functions. J. Steroid Biochem. Mol. Biol. 2013, 137, 107–123. [Google Scholar] [CrossRef] [Green Version]
- Dréno, B. What is new in the pathophysiology of acne, an overview. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 8–12. [Google Scholar] [CrossRef]
- Mirdamadi, Y.; Bommhardt, U.; Goihl, A.; Guttek, K.; Zouboulis, C.C.; Quist, S.; Gollnick, H. Insulin and Insulin-like growth factor-1 can activate the phosphoinositide-3-kinase /Akt/FoxO1 pathway in T cells in vitro. Dermatoendocrinology 2017, 9, e1356518. [Google Scholar] [CrossRef] [Green Version]
- Melnik, B.C. The role of transcription factor FoxO1 in the pathogenesis of acne vulgaris and the mode of isotretinoin action. G. Ital. Dermatol. Venereol. 2010, 145, 559–571. [Google Scholar]
- Li, J.; Al-Azzawi, F. Mechanism of androgen receptor action. Maturitas 2009, 63, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.A.; Bagatin, E. Adult-onset acne: Prevalence, impact, and management challenges. Clin. Cosmet. Investig. Dermatol. 2018, 11, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulden, V.; Clark, S.M.; Cunliffe, W.J. Post-adolescent acne: A review of clinical features. Br. J. Dermatol. 1997, 136, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Sardana, K.; Bansal, P.; Sharma, L.K.; Garga, U.C.; Vats, G. A study comparing the clinical and hormonal profile of late onset and persistent acne in adult females. Int. J. Dermatol. 2020, 59, 428–433. [Google Scholar] [CrossRef]
- Vexiau, P.; Chivot, M. Feminine acne: Dermatologic disease or endocrine disease? Gynecol. Obstet. Fertil. 2002, 30, 11–21. [Google Scholar] [CrossRef]
- Yoldemir, T. Postmenopausal hyperandrogenism. Climacteric 2022, 25, 109–117. [Google Scholar] [CrossRef]
- Imperato-McGinley, J.; Gautier, T.; Cai, L.Q.; Yee, B.; Epstein, J.; Pochi, P. The androgen control of sebum production. Studies of subjects with dihydrotestosterone deficiency and complete androgen insensitivity. J. Clin. Endocrinol. Metab. 1993, 76, 524–528. [Google Scholar] [CrossRef]
- Juul, A.; Holm, K.; Kastrup, K.W.; Pedersen, S.A.; Michaelsen, K.F.; Scheike, T.; Rasmussen, S.; Müller, J.; Skakkebaek, N.E. Free insulin-like growth factor I serum levels in 1430 healthy children and adults, and its diagnostic value in patients suspected of growth hormone deficiency. J. Clin. Endocrinol. Metab. 1997, 82, 2497–2502. [Google Scholar] [CrossRef]
- Ben-Amitai, D.; Laron, Z. Effect of insulin-like growth factor-1 deficiency or administration on the occurrence of acne. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 950–954. [Google Scholar] [CrossRef]
- Leyden, J.; Bergfeld, W.; Drake, L.; Dunlap, F.; Goldman, M.P.; Gottlieb, A.B.; Heffernan, M.P.; Hickman, J.G.; Hordinsky, M.; Jarrett, M.; et al. A systemic type I 5 alpha-reductase inhibitor is ineffective in the treatment of acne vulgaris. J. Am. Acad. Dermatol. 2004, 50, 443–447. [Google Scholar] [CrossRef]
- Seiffert, K.; Seltmann, H.; Fritsch, M.; Zouboulis, C.C. Inhibition of 5alpha-reductase activity in SZ95 sebocytes and Ha-CaT keratinocytes in vitro. Horm. Metab. Res. 2008, 39, 141–148. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wu, W.J.; Yang, J.K.; Cheng, H.; Zuo, X.B.; Lai, W.; Gao, T.W.; Ma, C.L.; Luo, N.; Huang, J.Q.; et al. Two new susceptibility loci 1q24.2 and 11p11.2 confer risk to severe acne. Nat. Commun. 2014, 5, 2870. [Google Scholar] [CrossRef] [PubMed]
- Navarini, A.A.; Simpson, M.A.; Weale, M.; Knight, J.; Carlavan, I.; Reiniche, P.; Burden, D.A.; Layton, A.; Bataille, V.; Allen, M.; et al. Genome-wide association study identifies three novel susceptibility loci for severe Acne vulgaris. Nat. Commun. 2014, 5, 4020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mina-Vargas, A.; Colodro-Conde, L.; Grasby, K.; Zhu, G.; Gordon, S.; Medland, S.E.; Martin, N.G. Heritability and GWAS Analyses of Acne in Australian Adolescent Twins. Twin. Res. Hum. Genet. 2017, 20, 541–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsaie, M.L.; Aly, D.G. The Immunogenetics of Acne. Adv. Exp. Med. Biol. 2022, 1367, 137–154. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, B.L.; Saklatvala, J.R.; Dand, N.; Hagenbeek, F.A.; Li, X.; Min, J.L.; Thomas, L.; Bartels, M.; Jan Hottenga, J.; Lupton, M.K.; et al. Genome-wide association meta-analysis identifies 29 new acne susceptibility loci. Nat. Commun. 2022, 13, 702. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, T.C. A bacteriological and microscopical study of over three hundred vesicular and pustular lesions of the skin, with a research upon the etiology of Acne vulgaris. Johns Hopkins Hosp. Rep. 1900, 9, 409–430. [Google Scholar]
- Christensen, G.J.; Brüggemann, H. Bacterial skin commensals and their role as host guardians. Benef. Microbes 2014, 5, 201–215. [Google Scholar] [CrossRef]
- McDowell, A.; Perry, A.L.; Lambert, P.A.; Patrick, S. A new phylogenetic group of Propionibacterium acnes. J. Med. Microbiol. 2008, 57, 218–224. [Google Scholar] [CrossRef] [Green Version]
- McDowell, A.; Barnard, E.; Nagy, I.; Gao, A.; Tomida, S.; Li, H.; Eady, A.; Cove, J.; Nord, C.E.; Patrick, S. An expanded multilocus sequence typing scheme for propionibacterium acnes: Investigation of ‘pathogenic’, ‘commensal’ and antibiotic resistant strains. PLoS ONE 2012, 7, e41480. [Google Scholar] [CrossRef] [Green Version]
- Barnard, E.; Shi, B.; Kang, D.; Craft, N.; Li, H. The balance of metagenomic elements shapes the skin microbiome in acne and health. Sci. Rep. 2016, 6, 39491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitz-Gibbon, S.; Tomida, S.; Chiu, B.H.; Nguyen, L.; Du, C.; Liu, M.; Elashoff, D.; Erfe, M.C.; Loncaric, A.; Kim, J.; et al. Propionibacterium acnes strain populations in the human skin microbiome associated with acne. J. Investig. Dermatol. 2013, 133, 2152–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomholt, H.B.; Kilian, M. Population genetic analysis of Propionibacterium acnes identifies a subpopulation and epidemic clones associated with acne. PLoS ONE 2010, 5, e12277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.H.; Yoon, J.Y.; Park, S.Y.; Suh, D.H. Analysis of distribution patterns of Propionibacterium acnes phylotypes and Peptostreptococcus species from acne lesions. Br. J. Dermatol. 2013, 169, 1152–1155. [Google Scholar] [CrossRef]
- Lomholt, H.B.; Scholz, C.F.P.; Brüggemann, H.; Tettelin, H.; Kilian, M. A comparative study of Cutibacterium (Propionibacterium) acnes clones from acne patients and healthy controls. Anaerobe 2017, 47, 57–63. [Google Scholar] [CrossRef]
- McDowell, A.; Nagy, I.; Magyari, M.; Barnard, E.; Patrick, S. The opportunistic pathogen Propionibacterium acnes: Insights into typing, human disease, clonal diversification and CAMP factor evolution. PLoS ONE 2013, 8, e70897. [Google Scholar] [CrossRef] [Green Version]
- Dréno, B.; Pécastaings, S.; Corvec, S.; Veraldi, S.; Khammari, A.; Roques, C. Cutibacterium acnes (Propionibacterium acnes) and acne vulgaris: A brief look at the latest updates. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Paugam, C.; Corvec, S.; Saint-Jean, M.; Le Moigne, M.; Khammari, A.; Boisrobert, A.; Nguyen, J.M.; Gaultier, A.; Dréno, B. Propionibacterium acnes phylotypes and acne severity: An observational prospective study. J. Eur. Acad. Dermatol. Venereol. 2017, 31, e398–e399. [Google Scholar] [CrossRef]
- Tomida, S.; Nguyen, L.; Chiu, B.H.; Liu, J.; Sodergren, E.; Weinstock, G.M.; Li, H. Pan-genome and comparative genome analyses of propionibacterium acnes reveal its genomic diversity in the healthy and diseased human skin microbiome. mBio 2013, 4, e00003-13. [Google Scholar] [CrossRef] [Green Version]
- Mayslich, C.; Grange, P.A.; Dupin, N. Cutibacterium acnes as an Opportunistic Pathogen: An Update of Its Virulence-Associated Factors. Microorganisms 2021, 9, 303. [Google Scholar] [CrossRef]
- Valanne, S.; McDowell, A.; Ramage, G.; Tunney, M.M.; Einarsson, G.G.; O’Hagan, S.; Wisdom, G.B.; Fairley, D.; Bhatia, A.; Maisonneuve, J.F.; et al. CAMP factor homologues in Propionibacterium acnes: A new protein family differentially expressed by types I and II. Microbiology 2005, 151, 1369–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayslich, C.; Grange, P.A.; Castela, M.; Marcelin, A.G.; Calvez, V.; Dupin, N. Characterization of a Cutibacterium acnes Camp Factor 1-Related Peptide as a New TLR-2 Modulator in In Vitro and Ex Vivo Models of Inflammation. Int. J. Mol. Sci. 2022, 23, 5065. [Google Scholar] [CrossRef]
- Lheure, C.; Grange, P.A.; Ollagnier, G.; Morand, P.; Désiré, N.; Sayon, S.; Corvec, S.; Raingeaud, J.; Marcelin, A.G.; Calvez, V.; et al. TLR-2 Recognizes Propionibacterium acnes CAMP Factor 1 from Highly Inflammatory Strains. PLoS ONE 2016, 11, e0167237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.; Kang, D.; Barnard, E.; Li, H. Strain-Level Differences in Porphyrin Production and Regulation in Propionibacterium acnes Elucidate Disease Associations. mSphere 2016, 1, e00023-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnard, E.; Johnson, T.; Ngo, T.; Arora, U.; Leuterio, G.; McDowell, A.; Li, H. Porphyrin Production and Regulation in Cutaneous Propionibacteria. mSphere 2020, 5, e00793-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazipi, S.; Stødkilde-Jørgensen, K.; Scavenius, C.; Brüggemann, H. The Skin Bacterium Propionibacterium acnes Employs Two Variants of Hyaluronate Lyase with Distinct Properties. Microorganisms 2017, 5, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bek-Thomsen, M.; Lomholt, H.B.; Scavenius, C.; Enghild, J.J.; Brüggemann, H. Proteome analysis of human sebaceous follicle infundibula extracted from healthy and acne-affected skin. PLoS ONE 2014, 9, e107908. [Google Scholar] [CrossRef]
- Borrel, V.; Gannesen, A.V.; Barreau, M.; Gaviard, C.; Duclairoir-Poc, C.; Hardouin, J.; Konto-Ghiorghi, Y.; Lefeuvre, L.; Feuilloley, M.G.J. Adaptation of acneic and non acneic strains of Cutibacterium acnes to sebum-like environment. Microbiologyopen 2019, 8, e00841. [Google Scholar] [CrossRef] [Green Version]
- Dréno, B.; Gollnick, H.P.; Kang, S.; Thiboutot, D.; Bettoli, V.; Torres, V.; Leyden, J. Understanding innate immunity and inflammation in acne: Implications for management. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Farrar, M.D.; Ingham, E.; Holland, K.T. Heat shock proteins and inflammatory acne vulgaris: Molecular cloning, overexpression and purification of a propionibacterium acnes GroEL and DnaK homologue. FEMS Microbiol. Lett. 2000, 191, 183–186. [Google Scholar] [CrossRef]
- Cobian, N.; Garlet, A.; Hidalgo-Cantabrana, C.; Barrangou, R. Comparative Genomic Analyses and CRISPR-Cas Characterization of Cutibacterium acnes Provide Insights Into Genetic Diversity and Typing Applications. Front. Microbiol. 2021, 12, 758749. [Google Scholar] [CrossRef] [PubMed]
- Holmberg, A.; Lood, R.; Mörgelin, M.; Söderquist, B.; Holst, E.; Collin, M.; Christensson, B.; Rasmussen, M. Biofilm formation by Propionibacterium acnes is a characteristic of invasive isolates. Clin. Microbiol. Infect. 2009, 15, 787–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coenye, T.; Spittaels, K.J.; Achermann, Y. The role of biofilm formation in the pathogenesis and antimicrobial susceptibility of Cutibacterium acnes. Biofilm 2021, 4, 100063. [Google Scholar] [CrossRef]
- Jahns, A.C.; Lundskog, B.; Ganceviciene, R.; Palmer, R.H.; Golovleva, I.; Zouboulis, C.C.; McDowell, A.; Patrick, S.; Alexeyev, O.A. An increased incidence of Propionibacterium acnes biofilms in acne vulgaris: A case-control study. Br. J. Dermatol. 2012, 167, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Spittaels, K.J.; Coenye, T. Developing an in vitro artificial sebum model to study Propionibacterium acnes biofilms. Anaerobe 2018, 49, 21–29. [Google Scholar] [CrossRef]
- Ramage, G.; Tunney, M.M.; Patrick, S.; Gorman, S.P.; Nixon, J.R. Formation of Propionibacterium acnes biofilms on orthopaedic biomaterials and their susceptibility to antimicrobials. Biomaterials 2003, 24, 3221–3227. [Google Scholar] [CrossRef]
- Jahns, A.C.; Eilers, H.; Alexeyev, O.A. Transcriptomic analysis of Propionibacterium acnes biofilms in vitro. Anaerobe 2016, 42, 111–118. [Google Scholar] [CrossRef]
- Yu, Y.; Champer, J.; Agak, G.W.; Kao, S.; Modlin, R.L.; Kim, J. Different Propionibacterium acnes Phylotypes Induce Distinct Immune Responses and Express Unique Surface and Secreted Proteomes. J. Investig. Dermatol. 2016, 136, 2221–2228. [Google Scholar] [CrossRef] [Green Version]
- Ermertcan, A.T.; Öztürk, F.; Gündüz, K. Toll-like receptors and skin. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 997–1006. [Google Scholar] [CrossRef]
- Graham, G.M.; Farrar, M.D.; Cruse-Sawyer, J.E.; Holland, K.T.; Ingham, E. Proinflammatory cytokine production by human keratinocytes stimulated with Propionibacterium acnes and P. acnes GroEL. Br. J. Dermatol. 2004, 150, 421–428. [Google Scholar] [CrossRef]
- Jeremy, A.H.; Holland, D.B.; Roberts, S.G.; Thomson, K.F.; Cunliffe, W.J. Inflammatory events are involved in acne lesion initiation. J. Investig. Dermatol. 2003, 121, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Karagianni, F.; Pavlidis, A.; Malakou, L.S.; Piperi, C.; Papadavid, E. Predominant Role of mTOR Signaling in Skin Diseases with Therapeutic Potential. Int. J. Mol. Sci. 2022, 23, 1693. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Bloch, W.; Iden, S.; Rüegg, M.A.; Hall, M.N.; Leptin, M.; Partridge, L.; Eming, S.A. mTORC1 and mTORC2 regulate skin morphogenesis and epidermal barrier formation. Nat. Commun. 2016, 7, 13226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monfrecola, G.; Lembo, S.; Caiazzo, G.; De Vita, V.; Di Caprio, R.; Balato, A.; Fabbrocini, G. Mechanistic target of rapamycin (mTOR) expression is increased in acne patients’ skin. Exp. Dermatol. 2016, 25, 153–155. [Google Scholar] [CrossRef] [Green Version]
- Agamia, N.F.; Abdallah, D.M.; Sorour, O.; Mourad, B.; Younan, D.N. Skin expression of mammalian target of rapamycin and forkhead box transcription factor O1, and serum insulin-like growth factor-1 in patients with acne vulgaris and their relationship with diet. Br. J. Dermatol. 2016, 174, 1299–1307. [Google Scholar] [CrossRef]
- Jung, M.J.; Lee, J.; Shin, N.R.; Kim, M.S.; Hyun, D.W.; Yun, J.H.; Kim, P.S.; Whon, T.W.; Bae, J.W. Chronic Repression of mTOR Complex 2 Induces Changes in the Gut Microbiota of Diet-induced Obese Mice. Sci. Rep. 2016, 6, 30887. [Google Scholar] [CrossRef] [Green Version]
- Noureldein, M.H.; Eid, A.A. Gut microbiota and mTOR signaling: Insight on a new pathophysiological interaction. Microb. Pathog. 2018, 118, 98–104. [Google Scholar] [CrossRef]
- Strickle, A.; Kolmer, J.A.; Schamberg, J.F. Complement fixation in acne vulgaris. J. Cutan. Dis. 1916, 34, 166–178. [Google Scholar]
- Stokes, J.H.; Pillsbury, D.M. The effect on the skin of emotional and nervous states: III. Theoretical and practical consideration of a gastrointestinal mechanism. Arch. Derm. Syphilol. 1930, 22, 962–993. [Google Scholar] [CrossRef]
- Fukui, H.; Wiest, R. Changes of Intestinal Functions in Liver Cirrhosis. Inflamm. Intest. Dis. 2016, 1, 24–40. [Google Scholar] [CrossRef]
- Juhlin, L.; Michaëlsson, G. Fibrin microclot formation in patients with acne. Acta Derm. Venereol. 1983, 63, 538–540. [Google Scholar] [PubMed]
- Loveman, D.E.; Noojin, R.O.; Winkler, C.H., Jr. Comparative studies of enteric bacterial flora in acne vulgaris. J. Investig. Dermatol. 1955, 25, 135–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volkova, L.A.; Khalif, I.L.; Kabanova, I.N. Impact of the impaired intestinal microflora on the course of acne vulgaris. Klin. Med. (Mosk.) 2001, 79, 39–41. [Google Scholar] [PubMed]
- Deng, Y.; Wang, H.; Zhou, J.; Mou, Y.; Wang, G.; Xiong, X. Patients with Acne Vulgaris Have a Distinct Gut Microbiota in Comparison with Healthy Controls. Acta Derm. Venereol. 2018, 98, 783–790. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Liu, L.; Chen, L.; Zhou, L.; Xiong, X.; Deng, Y. Gender-Specific Differences in Gut Microbiota Composition Associated with Microbial Metabolites for Patients with Acne Vulgaris. Ann. Dermatol. 2021, 33, 531–540. [Google Scholar] [CrossRef]
- Balić, A.; Vlašić, D.; Žužul, K.; Marinović, B.; Bukvić Mokos, Z. Omega-3 Versus Omega-6 Polyunsaturated Fatty Acids in the Prevention and Treatment of Inflammatory Skin Diseases. Int. J. Mol. Sci. 2020, 21, 741. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, A.; Bruhs, A.; Schwarz, T. The short-chain fatty acid sodium butyrate functions as a regulator of the skin immune system. J. Investig. Dermatol. 2017, 137, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Yan, H.M.; Zhao, H.J.; Guo, D.Y.; Zhu, P.Q.; Zhang, C.L.; Jiang, W. Gut microbiota alterations in moderate to severe acne vulgaris patients. J. Dermatol. 2018, 45, 1166–1171. [Google Scholar] [CrossRef]
- Lee, D.K.; Kim, M.J.; Ham, J.W.; An, H.M.; Cha, M.K.; Lee, S.W.; Park, C.I.; Shin, S.H.; Lee, K.O.; Kim, K.J.; et al. In vitro evaluation of antibacterial activities and anti-inflammatory effects of Bifidobacterium spp. addressing acne vulgaris. Arch. Pharm. Res. 2012, 35, 1065–1071. [Google Scholar] [CrossRef]
- Thompson, K.G.; Rainer, B.M.; Antonescu, C.; Florea, L.; Mongodin, E.F.; Kang, S.; Chien, A.L. Minocycline and Its Impact on Microbial Dysbiosis in the Skin and Gastrointestinal Tract of Acne Patients. Ann. Dermatol. 2020, 32, 21–30. [Google Scholar] [CrossRef]
- Rainer, B.M.; Thompson, K.G.; Antonescu, C.; Florea, L.; Mongodin, E.F.; Kang, S.; Chien, A.L. Impact of lifestyle and demographics on the gut microbiota of acne patients and the response to minocycline. J. Dermatol. Treat. 2021, 32, 934–935. [Google Scholar] [CrossRef] [PubMed]
- Bowe, W.P.; Filip, J.C.; DiRienzo, J.M.; Volgina, A.; Margolis, D.J. Inhibition of propionibacterium acnes by bacteriocin-like inhibitory substances (BLIS) produced by Streptococcus salivarius. J. Drugs Dermatol. 2006, 5, 868–870. [Google Scholar] [PubMed]
- Oh, S.; Kim, S.H.; Ko, Y.; Sim, J.H.; Kim, K.S.; Lee, S.H.; Park, S.; Kim, Y.J. Effect of bacteriocin produced by Lactococcus sp. HY 449 on skin-inflammatory bacteria. Food Chem. Toxicol. 2006, 44, 1184–1190. [Google Scholar] [CrossRef] [PubMed]
- Deidda, F.; Amoruso, A.; Nicola, S.; Graziano, T.; Pane, M.; Mogna, L. New Approach in Acne Therapy: A Specific Bacteriocin Activity and a Targeted Anti IL-8 Property in Just 1 Probiotic Strain, the L. salivarius LS03. J. Clin. Gastroenterol. 2018, 52, S78–S81. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kuo, S.; Shu, M.; Yu, J.; Huang, S.; Dai, A.; Two, A.; Gallo, R.L.; Huang, C.M. Staphylococcus epidermidis in the human skin microbiome mediates fermentation to inhibit the growth of Propionibacterium acnes: Implications of probiotics in acne vulgaris. Appl. Microbiol. Biotechnol. 2014, 98, 411–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosseau, C.; Devine, D.A.; Dullaghan, E.; Gardy, J.L.; Chikatamarla, A.; Gellatly, S.; Yu, L.L.; Pistolic, J.; Falsafi, R.; Tagg, J.; et al. The commensal Streptococcus salivarius K12 downregulates the innate immune responses of human epithelial cells and promotes host-microbe homeostasis. Infect. Immun. 2008, 76, 4163–4175. [Google Scholar] [CrossRef] [Green Version]
- Gueniche, A.; Benyacoub, J.; Philippe, D.; Bastien, P.; Kusy, N.; Breton, L.; Blum, S.; Castiel-Higounenc, I. Lactobacillus paracasei CNCM I-2116 (ST11) inhibits substance P-induced skin inflammation and accelerates skin barrier function recovery in vitro. Eur. J. Dermatol. 2010, 20, 731–737. [Google Scholar] [CrossRef]
- Pavicic, T.; Wollenweber, U.; Farwick, M.; Korting, H.C. Anti-microbial and -inflammatory activity and efficacy of phytosphingosine: An in vitro and in vivo study addressing acne vulgaris. Int. J. Cosmet. Sci. 2007, 29, 181–191. [Google Scholar] [CrossRef]
- Di Marzio, L.; Cinque, B.; De Simone, C.; Cifone, M.G. Effect of the lactic acid bacterium Streptococcus thermophilus on ceramide levels in human keratinocytes in vitro and stratum corneum in vivo. J. Investig. Dermatol. 1999, 113, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Al-Ghazzewi, F.H.; Tester, R.F. Effect of konjac glucomannan hydrolysates and probiotics on the growth of the skin bacterium Propionibacterium acnes in vitro. Int. J. Cosmet. Sci. 2010, 32, 139–142. [Google Scholar] [CrossRef]
- Lopes, E.G.; Moreira, D.A.; Gullón, P.; Gullón, B.; Cardelle-Cobas, A.; Tavaria, F.K. Topical application of probiotics in skin: Adhesion, antimicrobial and antibiofilm in vitro assays. J. Appl. Microbiol. 2017, 122, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Chae, M.; Kim, B.J.; Na, J.; Kim, S.Y.; Lee, J.O.; Kim, Y.J.; Lee, E.; Cho, D.; Roh, J.; Kim, W. Antimicrobial activity of Lactiplantibacillus plantarum APsulloc 331261 and APsulloc 331266 against pathogenic skin microbiota. Front. Biosci. (Elite Ed.) 2021, 13, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Espinoza-Monje, M.; Campos, J.; Alvarez Villamil, E.; Jerez, A.; Dentice Maidana, S.; Elean, M.; Salva, S.; Kitazawa, H.; Villena, J.; García-Cancino, A. Characterization of Weissella viridescens UCO-SMC3 as a Potential Probiotic for the Skin: Its Beneficial Role in the Pathogenesis of Acne Vulgaris. Microorganisms 2021, 9, 1486. [Google Scholar] [CrossRef]
- Siver, R. Lactobacillus for the control of acne. J. Med. Soc. N. J. 1961, 59, 52–53. [Google Scholar]
- Kim, J.; Ko, Y.; Park, Y.K.; Kim, N.I.; Ha, W.K.; Cho, Y. Dietary effect of lactoferrin-enriched fermented milk on skin surface lipid and clinical improvement of acne vulgaris. Nutrition 2010, 26, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Cui, W.; Wei, H. Influence of lactoferrin on Propionibacterium acnes-induced inflammation in vitro and in vivo. Dermatol. Ther. 2020, 33, e14483. [Google Scholar] [CrossRef]
- Jung, G.W.; Tse, J.E.; Guiha, I.; Rao, J. Prospective, randomized, open-label trial comparing the safety, efficacy, and tolerability of an acne treatment regimen with and without a probiotic supplement and minocycline in subjects with mild to moderate acne. J. Cutan. Med. Surg. 2013, 17, 114–122. [Google Scholar] [CrossRef]
- Fabbrocini, G.; Bertona, M.; Picazo, Ó.; Pareja-Galeano, H.; Monfrecola, G.; Emanuele, E. Supplementation with Lactobacillus rhamnosus SP1 normalises skin expression of genes implicated in insulin signalling and improves adult acne. Benef. Microbes 2016, 7, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Rahmayani, T.; Putra, I.B.; Jusuf, N.K. The Effect of Oral Probiotic on the Interleukin-10 Serum Levels of Acne Vulgaris. Open Access Maced. J. Med. Sci. 2019, 7, 3249–3252. [Google Scholar] [CrossRef] [Green Version]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Tolino, E.; Skroza, N.; Mambrin, A.; Bernardini, N.; Zuber, S.; Balduzzi, V.; Marchesiello, A.; Proietti, I.; Potenza, C. Novel combination for the treatment of acne differentiated based on gender: A new step towards personalized treatment. G. Ital. Dermatol. Venereol. 2018, 153, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Manzhalii, E.; Hornuss, D.; Stremmel, W. Intestinal-borne dermatoses significantly improved by oral application of Escherichia coli Nissle 1917. World J. Gastroenterol. 2016, 22, 5415–5421. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, F.; Marotta, L.; Mascolo, A.; Amoruso, A.; Pane, M.; Giuliani, G.; Pinto, D. Facial Acne: A Randomized, Double-Blind, Placebo-Controlled Study on the Clinical Efficacy of a Symbiotic Dietary Supplement. Dermatol. Ther. 2022, 12, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Habeebuddin, M.; Karnati, R.K.; Shiroorkar, P.N.; Nagaraja, S.; Asdaq, S.M.B.; Khalid Anwer, M.; Fattepur, S. Topical Probiotics: More Than a Skin Deep. Pharmaceutics 2022, 14, 557. [Google Scholar] [CrossRef]
- Peyri, J. Topical bacteriotherapy of the skin. J. Cutan. Dis. 1912, 30, 688–689. [Google Scholar]
- Kang, S.; Cho, S.; Chung, J.H.; Hammerberg, C.; Fisher, G.J.; Voorhees, J.J. Inflammation and extracellular matrix degradation mediated by activated transcription factors nuclear factor-kappaB and activator protein-1 in inflammatory acne lesions in vivo. Am. J. Pathol. 2005, 166, 1691–1699. [Google Scholar] [CrossRef]
- AOBiome Therapeutics. Available online: https://www.aobiome.com/pressreleases/aobiome-therapeutics-reports-positive-efficacy-results-from-phase-2b-clinical-trial-of-ammonia-oxidizing-bacteria-aob-for-the-treatment-of-acne-vulgaris (accessed on 3 May 2022).
- Sathikulpakdee, S.; Kanokrungsee, S.; Vitheejongjaroen, P.; Kamanamool, N.; Udompataikul, M.; Taweechotipatr, M. Efficacy of probiotic-derived lotion from Lactobacillus paracasei MSMC 39-1 in mild to moderate acne vulgaris, randomized controlled trial. J. Cosmet. Dermatol. 2022; in press. [Google Scholar] [CrossRef]
- Dall’Oglio, F.; Nasca, M.R.; Fiorentini, F.; Micali, G. Diet and acne: Review of the evidence from 2009 to 2020. Int. J. Dermatol. 2021, 60, 672–685. [Google Scholar] [CrossRef]
- Çerman, A.A.; Aktaş, E.; Altunay, İ.K.; Arıcı, J.E.; Tulunay, A.; Ozturk, F.Y. Dietary glycemic factors, insulin resistance, and adiponectin levels in acne vulgaris. J. Am. Acad. Dermatol. 2016, 75, 155–162. [Google Scholar] [CrossRef]
- Smith, T.M.; Gilliland, K.; Clawson, G.A.; Thiboutot, D. IGF-1 induces SREBP-1 expression and lipogenesis in SEB-1 sebocytes via activation of the phosphoinositide 3-kinase/Akt pathway. J. Investig. Dermatol. 2008, 128, 1286–1293. [Google Scholar] [CrossRef] [Green Version]
- Jewell, J.L.; Russell, R.C.; Guan, K.L. Amino acid signalling upstream of mTOR. Nat. Rev. Mol. Cell. Biol. 2013, 14, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Melnik, B. Dietary intervention in acne: Attenuation of increased mTORC1 signaling promoted by Western diet. Dermatoendocrinology 2012, 4, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef] [PubMed]
- Ansaldo, E.; Farley, T.K.; Belkaid, Y. Control of Immunity by the Microbiota. Annu. Rev. Immunol. 2021, 39, 449–479. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Reference | Methodology and Study Population | Key Results |
|---|---|---|
| Loveman 1955 [102]. | A total of 10 patients with severe pustular acne and 10 healthy controls. Stool microbiological culture. | No significant differences were found in a small classic group of intestinal pathogenic bacteria compared to a group of 10 healthy controls |
| Volkova 2001 [103]. | A total of 114 patients with acne. Stool microbiological culture. | 54% of acne patients presented gut dysbiosis. |
| Deng 2018 [104]. | A total of 43 treatment-naïve patients with different degrees of acne versus 43 age- and sex-matched healthy controls. NGS 16S rRNA. | Decreased alpha diversity in acne patients compared to healthy controls. No differences in severity. PCoA and ANOSIM analyses showed two different clusters for cases and controls. LefSe analysis found 38 differences between the groups of acne patients and healthy controls. Decrease in Firmicutes and increase in Bacteroidetes in the group of patients with acne. Decrease in Clostridial families such as Lachnospiraceae and Ruminococcaceae, producers of SCFA in patients with acne. PICRUSt analysis showed an increase in LPS synthesis pathways in acne patients. |
| Yan 2018 [108]. | A total of 31 mild-moderate acne patients and 31 age- and sex-matched healthy controls. NGS 16S rRNA. | No differences were observed regarding alpha diversity between cases with acne and controls. PCA did not show different clusters. Gut microbiota between cases and controls was quite similar. Loss in Bifidobacterium and a slight gain in Proteobacteria in patients with acne. |
| Thompson 2020 [110]. | Eight patients with acne before and after receiving minocycline as treatment and eight healthy age- and sex-matched controls. NGS 16S rRNA. | Not significant differences in alpha diversity were between acne patients before and after antibiotic therapy. Gut microbiota of patients with acne before taking antibiotics compared to healthy controls showed a decrease in Lactobacillus iners, Lactobacillus zeae, and Bifidobacterium animalis. After antibiotic treatment, gut microbiota of acne patients showed decreases in Lactobacillus salivarius, Bifidobacterium adolescentis, Bifidobacterium pseudolongum, Bifidobacterium breve, and Akkermansia mucinophila compared to healthy controls. Bacteroidetes increase in acne patients after treatment. Decrease in Firmicutes/Bacteroidetes ratio. |
| Rainer 2021 [111]. | Eight subjects with acne after 4 weeks treatment with minocycline and eight age-, race-, and sex-matched healthy controls. NGS 16S rRNA. | Firmicutes/Bacteroidetes ratio was higher in controls and lower in patients with acne after treatment Slightly higher baseline Firmicutes/Bacteroidetes ratio was observed in patients with less-severe acne compared to those with greater severity. |
| Huang 2021 [105]. | A total of 86 study subjects: 26 men with acne + 26 healthy control men and 17 women with acne + 17 healthy control women. NGS 16S rRNA and fecal metabolomic study. | Alpha diversity was lower in men with acne compared to control men. No significant differences were found between women with acne and control women. ANOSIM test showed four statistically significant clusters among the four study groups: Firmicutes and Bacteroidetes were significantly increased and decreased, respectively, in men with acne versus control men. These differences were not found in women with acne versus control women. At the genus level, decreases were observed in men with acne compared to control men from 18 genera. Many genera were SCFA producers. In women with acne compared to control women, an increase in Clostridium sensu stricto and a decrease in Oscillibacter and Odoribacter were observed. Men with acne tended to have impaired fatty acid metabolism while women with acne tended to have impaired amino acid metabolism. |
| Reference | Study | Probiotic | Key Results |
|---|---|---|---|
| Bowe [112]. | In vitro | Streptococcus salivarius | Bacteriocin inhibited C. acnes growth. |
| Oh [113]. | In vitro | Lactococcus sp. HY 449 | Bacteriocin inhibited C. acnes growth. |
| Deidda [114]. | In vitro | Lactobacillus salivarius LS03 | Bacteriocin inhibited C. acnes growth. |
| Lee [109]. | In vitro | Bifidobacterium adolescentis | Antimicrobial activity against C. acnes and Staphylococcus aureus. |
| Wang [115]. | In vitro | Staphylococcus epidermidis | Production of succinic acid through glycerol fermentation. |
| Cosseau [116]. | In vitro | Streptococcus salivarius K12 | Anti-inflammatory response; modulation of genes associated with epithelial adhesion. |
| Gueniche [117]. | In vitro | Lactobacillus paracasei CNCM I-2126 | Improvement of skin barrier function. |
| Al-Ghazzewi [120]. | In vitro | L. casei NCFB 161, L. acidophilus NCFB 1748, L. plantarum DSM 12028, L. gasseri NCFB 2233, and Lactococcus lactis NCIMB 66 plus glucomannan hydrolysates of Amorphophallus konjac | Inhibition of C. acnes growth. |
| Lopes [121]. | In vitro | Several Bifidobacterium and Lactobacillus strains | Adherence to keratin; inhibition of biofilm formation of pathogenic bacteria; limited ability to adhere to C. acnes. |
| Chae [122]. | In vitro | L. plantarum APsulloc 331261 and APsulloc 331266 | Inhibition of skin pathogen growth. |
| Espinoza-Monje [123]. | In vitro and murine model | Weissella viridescens UCO_SMC3 | Inhibition of C. acnes growth; anti-inflammatory effects. |
| Siver [124]. | Clinical trial | L. acidophilus and L. bulgaricus (oral) | A total of 300 acne patients; 2 weeks of treatment. Clinical improvement in 80% of acne patients. |
| Jung [127]. | RCT, open-label | L. acidophilus (5 × 109 CFU/capsule), L. bulgaricus (5 × 109 CFU/capsule) and B. bifidum (20 × 109 CFU/capsule) (oral); two capsules/day | A total of 45 acne patients; three study groups (probiotic, minocycline, probiotic plus minocycline); 12 weeks of treatment. Patients treated with probiotic mixture plus minocycline had significantly better efficacy in terms of total number of lesions. |
| Fabbrocini [128]. | RCT, double-blinded, placebo-controlled | Liquid supplement containing Lactobacillus rhamnosus SP1 at a dose of 3 × 109 CFU/day (oral) | A total of 20 acne patients; 12 weeks of treatment. IGF-1 and FoxO1 gene expression in skin acne areas. Statistically significant reduction in the expression of the IGF-1 gene of 32% and a statistically significant increase in the FoxO1 gene of 65% in probiotic group. Clinical improvement in patients treated with probiotic. |
| Rahmayani [129] | Pre-experimental clinical study with a pretest/posttest | B. lactis W51, B. lactis W52, L. acidophilus W55, L. casei W56, L. salivarius W57, and Lactococcus lactis W58, with total bacterial cells > 108 CFU per sachet (oral); two sachets/day | A total of 30 acne patients; 30 days of treatment. An increase in IL-10 was seen after probiotic mixture treatment. |
| Manzhalii [132]. | RCT, controlled, nonblinded | Escherichia coli Nissle 1917 (oral); one capsule contained 2.5–25 × 109 CFU; two capsules/day | A total of 82 patients with intestinal-borne dermatoses (some of them were diagnosed with acne); 1 month of treatment. Two study groups (patients treated with a conventional topical therapy consisting of ointments containing tetracycline, steroids, and retinoids; and patients treated with conventional topical therapy plus probiotic). A total of 89% of patients treated with E. coli Nissle 1917 improved significantly, while 56% improved in the group treated with only the conventional topical therapy. |
| Rinaldi [133]. | RCT, double-blinded, placebo-controlled | B. breve BR03 (0.5 × 109 CFU), L. casei LC03 (≥0.5 × 109 CFU), and L. salivarius LS03 (≥1.0 × 109 CFU) plus a botanical extract of Solanum melongena and Echinacea (oral); one sachet/day | A total of 114 acne patients. Four study groups (placebo, probiotics, botanical extracts, and probiotics plus botanical extracts); 8 weeks of treatment. A decreased number of acne lesions, rate of desquamation, rate of sebum secretion, and presence of C. acnes was found in patients treated with the probiotic mixture and botanical extract, and a mixture of both. Stronger effects were seen with the probiotic mix plus the botanical extract. |
| Kang [136]. | RCT, double-blinded, placebo-controlled | Concentrated powder lotion obtained from supernatant culture of Enterococcus faecalis SL-5 (topical) | A total of 70 acne patients; 8 weeks of treatment. A decrease in inflammatory lesions was seen. |
| AOBiome [137]. | RCT, double-blinded, placebo-controlled | Nitrosomonas eutropha (topical) | A total of 358 acne subjects; 12 weeks of treatment. A reduction in severity and a trend toward a reduction in inflammatory lesions was seen. |
| Sathikulpakdee [138]. | RCT | Lactobacillus paraceasei MSMC 39-1 (topical) | A total of 104 acne patients; 4 weeks treatment. Topical probiotics vs. 2.5% benzoyl peroxide lotion were compared. Acne lesions and the erythema index were decreased. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Pellicer, P.; Navarro-Moratalla, L.; Núñez-Delegido, E.; Ruzafa-Costas, B.; Agüera-Santos, J.; Navarro-López, V. Acne, Microbiome, and Probiotics: The Gut–Skin Axis. Microorganisms 2022, 10, 1303. https://doi.org/10.3390/microorganisms10071303
Sánchez-Pellicer P, Navarro-Moratalla L, Núñez-Delegido E, Ruzafa-Costas B, Agüera-Santos J, Navarro-López V. Acne, Microbiome, and Probiotics: The Gut–Skin Axis. Microorganisms. 2022; 10(7):1303. https://doi.org/10.3390/microorganisms10071303
Chicago/Turabian StyleSánchez-Pellicer, Pedro, Laura Navarro-Moratalla, Eva Núñez-Delegido, Beatriz Ruzafa-Costas, Juan Agüera-Santos, and Vicente Navarro-López. 2022. "Acne, Microbiome, and Probiotics: The Gut–Skin Axis" Microorganisms 10, no. 7: 1303. https://doi.org/10.3390/microorganisms10071303