
Secondary Metabolites from the Endoparasitic Nematophagous Fungus Harposporium anguillulae YMF 1.01751

State Key Laboratory for Conservation and Utilization of Bio-Resources in Yunnan, Key Laboratory for Southwest Microbial Diversity of the Ministry of Education, Yunnan University, Kunming 650091, China
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(8), 1553; https://doi.org/10.3390/microorganisms10081553
Submission received: 6 July 2022
/
Revised: 28 July 2022
/
Accepted: 29 July 2022
/
Published: 31 July 2022
(This article belongs to the Special Issue Secondary Metabolism of Microorganisms 2.0)
Abstract
:Harposporium anguillulae, an endoparasitic nematophagous fungus (ENF), is a model fungus from which the genus Harposporium was established. It can infect nematodes via ingested conidia. In this paper, the morphology and nematode–fungus interaction between Panagrellus redivivus and H. anguillulae were observed by scanning electron microscopy (SEM). The secondary metabolites of H. anguillulae were also studied. Seven metabolites were purified and identified from an ethyl acetate extract of broth and a methanol extract of mycelium. These include a new polyketone 5-hydroxy-3-(hydroxymethyl)-6-methyl-2H-pyran-2-one (1) and six known metabolites (17R)-17-methylincisterol (2), eburicol (3), ergosterol peroxide (4), terpendole C (5), (3β,5α,9β,22E)-3,5-dihydroxy-ergosta-7,22-dien-6-one (6), and 5α,6β-epoxy-(22E,24R)-ergosta-8,22-diene- 3β,7α-diol (7). These metabolites were assayed for their activity against plant root-knot nematode, Meloidogyne incognita, and the results showed that terpendole C (5) had weak nematicidal activity but also that other compounds did not have evident activity at a concentration of 400 μg mL−1. Compound 1 exhibited an attractive effect towards P. redivivus.
1. Introduction
The diseases caused by plant parasitic nematodes lead to huge economic losses to crops every year [1]. Traditional chemical pesticides to control nematode diseases have irreversible damaged the ecological environment, so biological control has gradually become a hot topic of research. Nematophagous fungi play an important role as the main source of nematode biological control management [2]. Nematophagous fungi include trapping fungi, endoparasitic nematophagous fungi (ENF), opportunistic fungi, and toxic fungi [3]. Among them, ENF are a type of nematophagous fungi that can parasitize nematodes through special spores.
Fungi of the genus Harposporium are a class of endoparasitic nematophagous fungi that can produce conidia to parasitize nematodes, and their unique spore morphology provides more possibilities for them to exhibit nematicidal effects [4]. Harposporium was established by Lohde (1874) [5], and the genus is an important group of endoparasitic nematophagous fungi. Harposporium species are widely distributed and can be isolated from soil and substrate nematodes [6,7]. Up to now, 31 species of Harposporium have been reported all over the world [8]. In addition to producing infection conidia, some species can also produce arthroconidia, chlamydospores, and accessory conidia [9]. In studies of Harposporium metabolites, only one paper has reported a new furan, harposporin A, along with a known aureonitol that was found in Harposporium sp. YMF 1.01735 [10].
H. anguillulae was the first fungus described as a parasite of nematodes. It is also the model fungus from which the genus Harposporium was established. It can infect nematodes by producing abundant ingested conidia. The process from ingesting conidia to infecting nematodes is divided into three processes: ingestion, germination, and penetration. H. anguillulae can form spores and chlamydospores in water–agar petri dishes. In an investigation of phospholipids on growth and sporulation of H. anguillulae, the result showed that the mycelial growth and sporulation of the species was not affected by the addition of phospholipids [11]. H. anguillulae can significantly reduce the free living stages of trichostrongylid nematodes, indicating that the species could be a candidate for the development of a biocontrol agent for trichostrongylid nematodes [12].
In the present investigation, research on H. anguillulae mainly focuses on strain isolation, identification, and pathogenicity to nematodes, in addition to studying secondary metabolites, which have not been reported for H. anguillulae. In this study, the strain H. anguillulae YMF 1.01751 was selected as the research object for its morphology and secondary metabolites, in which seven compounds, including one new polyketone, were identified.
2. Materials and Methods
2.1. Normal Materials
H. anguillulae YMF 1.01751 was isolated from soil in Yunnan Province, China, and is now deposited in the microbial library of the germplasm bank of wild species from Southwest China, Yunnan University, Kunming, China. It was stored in glycerol at –80 °C.
The plant root-knot nematode Meloidogyne incognita was obtained from the roots of tomatoes grown in E’shan County in Yunnan Province. The method of obtaining M. incognita is referred to in the literature [13].
Panagrellus redivivus: An appropriate amount of oat (25 g) and water (60 mL) was added to a 250 mL conical flask for sterilization. After cooling, an appropriate amount of P. redivivus seeds were picked and inoculated into oat medium and cultured at 25 °C for one week. The culture and preparation of P. redivivus is referred to in the literature [14].
2.2. General Experimental Instruments
Optical rotation was measured with a Jasco DIP-370 digital polarimeter (Tokyo, Japan). Ultraviolet (UV) spectra were recorded on a Shimadzu UV-2401PC spectrophotometer (Kyoto, Japan). Nuclear magnetic resonance (NMR) spectra were measured on an Avance III-600 spectrometer (Bruker Biospin, Rheinstetten, Germany). Electrospray ionization mass spectrometry (ESI–MS) spectra were recorded on a Thermo high-resolution Q Exactive Focus mass spectrometer (Thermo, Bremen, Germany). Column chromatography was performed on silica gel G (200–300 mesh, Qingdao Marine Chemical Inc., Qingdao, China) and Sephadex LH-20 (Amersham Biosciences, Piscataway, NJ, USA) columns.
2.3. The Morphology and Pathogenicity against Nematodes of H. anguillulae YMF 1.01751 Observed by Scanning Electron Microscopy
H. anguillulae YMF 1.01751 was cultured on improved potato dextrose agar (PDA) (200 g potato, 20 g glucose, 3 g KH2PO4, 1.5 g MgSO4, 5 g yeast extract, 2 g peptone, and 15 g agar) plates for 14 days. P. redivivus was cultured in oats for about 7 days, and the oats containing nematodes were placed into three layers of sterilized lens paper, which were then placed into sterile water to allow the nematodes separate from the oats. The nematodes were prepared as a suspension in water corresponding to about 100 nematodes/10 μL.
After growth of H. anguillulae YMF 1.01751 on improved PDA plates for 14 days, 20 μL P. redivivus suspension was added to the cultured strains for interaction. Samples were taken after 12, 24, 36, 48, 60, and 72 h of nematode–fungus coculture on plates. These samples were cut into small pieces and fixed for 30 min in 4% glutaraldehyde, then dehydrated with 30%, 50%, 70%, 80%, and 90% ethanol for 15 min each, respectively, and then dehydrated twice with 100% ethanol for 15 min each time, then, after that, in ethanol/isoamyl acetate (1:1, v/v) and in 100% isoamyl acetate liquor for 10 min, respectively. The samples were freeze-dried, gilded, and observed by scanning electron microscopy (SEM) [15].
2.4. Extraction and Isolation of Metabolites from H. anguillulae YMF 1.01751
H. anguillulae YMF 1.01751 was grown on 50 L optimized liquid medium (200 g potato, 20 g glucose, 3 g KH2PO4, 1.5 g MgSO4, 5 g yeast extract, 2 g peptone) at 25 °C for 21 days. The mycelia and fermentation broth were separated using 5 layers of gauze. The fermentation broth was concentrated to 2 L under vacuum and then transferred to a 5 L separation funnel. The crude extracts (10.3 g) extracted with ethyl acetate were fractionated with a silica gel G (200–300 mesh) column eluted using a petroleum ether–ethyl acetate (100:1-0:100) solvent system followed by trichloromethane–acetone (7:3) and trichloromethan–methanol (7:3) to yield 10 fractions (T1751-1~T1751-10). Dried mycelia were soaked in methanol for 5 days, and the metabolites were repeatedly extracted 3 times. The methanol phase was evaporated in vacuo to provide the mycelium extracts (50 g). The extracts were fractionated with a silica gel G (200–300 mesh) column eluted with a petroleum ether–acetone (100:1-0:100) gradient solvent system followed by ethyl acetate–methanol (10:1-0:1) to yield 16 fractions (B1751-1~B1751-16).
T1751-7 (920 mg) was subjected to a Sephadex LH-20 column eluted with methanol to yield 8 fractions, namely 7-1~7-8. Fraction 7-4 (350 mg) was purified by a silica gel column eluted with ethyl acetate–acetone (20:1-8:1) to produce 6 portions (7-4-1~7-4-6). Compound 1 (5 mg) was obtained from 7-4-1 (12 mg) with a methanol Sephadex LH-20 column. B1751-2 (205 mg) was subjected to a Sephadex LH-20 column eluted with methanol to yield 2-1 (52 mg) and 2-2 (30 mg). Fraction 2-1 was submitted to a silica gel column eluted with ethyl acetate–acetone (150:1) to obtain 2 fractions (2-1-1~2-1-2). These two fractions were each purified by an acetone Sephadex LH-20 column to obtain compounds 2 (6 mg) and 3 (3 mg). B1751-4 (97 mg) was submitted to a silica gel column eluted with ethyl acetate–acetone (120:1-60:1) to produce 3 fractions (4-1~4-3). Fraction 4-3 (9.6 mg) was isolated by a silica gel column eluted with dichloromethane–acetone (50:1-40:1) to yield 2 fractions (4-3-1~4-3-2). Compound 4 (8 mg) was obtained from 4-3-1 (8.5 mg) by a Sephadex LH-20 column eluted with trichloromethane–methanol (1:1, v/v).
B1751-8 (161 mg) was subjected to a methanol Sephadex LH-20 column to yield 3 fractions, namely 8-1~8-3. Fraction 8-3 (129 mg) was separated by a silica gel column eluted with ethyl acetate–acetone (50:1-5:1) to produce 4 portions (8-3-1~8-3-4). Fraction 8-3-3 (48 mg) was purified by a Sephadex LH-20 column eluted with methanol to yield 2 fractions (8-3-3-1 and 8-3-3-2). Fraction 8-3-3-1 (40 mg) was isolated by a silica gel column eluted with dichloromethane–acetone (50:1-5:1) and then purified on a methanol Sephadex LH-20 column to provide compound 5 (4 mg). B1751-11 (30 mg) was subjected to a methanol Sephadex LH-20 column to yield 3 fractions, namely 11-1~11-3. Fraction 11-3 was isolated on a silica gel column eluted with petroleum ether–acetone (10:1-3:1) to produce 3 fractions (11-3-1~11-3-3). Compounds 6 (1.7 mg) and 7 (2 mg) were obtained from 11-3-1 and 11-3-3 with a methanol Sephadex LH-20 column respectively.
2.5. Nematicidal and Chemotaxis Activity of Metabolites
The tested compounds (1, 2, 3, and 5) were dissolved in methanol and diluted with sterile water. Approximately 200 M. incognita nematodes J2s were added to each plate, and the final concentration of compounds was 400 µg mL−1. M. incognita was considered dead when no movement occurred after touching it with a needle [16]. The mortality was calculated at different times, and the experiment was repeated 3 times.
The nematode chemotaxis assay for compounds 1, 2, 3, and 5 was conducted using the four-point plate method [17]. The compounds’ concentrations were set to 4, 8, 16, and 32 µg. Approximately 200 worms were added to the center of the petri dish. Compounds and control (methanol) were added. After 2 h at 25 °C, the worms were counted under the microscope, and the chemotaxis index (CI) was calculated.
3. Results
3.1. Scanning Electron Microscopy Observation of Interactions between H. anguillulae YMF 1.01751 and P. redivivus
After growth on the improved PDA medium for two weeks at 28 °C, the hyphae of H. anguillulae YMF 1.01751 had overspread 6 cm on the plate. The mycelia of H. anguillulae YMF 1.01751 were septate and colorless. SEM results show that the phialides (sporogenous cell) of H. anguillulae YMF 1.01751 were globose or flask-shaped, and there was a conidial peduncle on the tip of phialides. Conidia were colorless, curved, bow-shaped, and had hooked ends (Figure 1A–D).
H. anguillulae produces ingested conidia to infect nematodes. While studying the infection of P. redivivus by H. anguillulae YMF 1.01751, approximately 200 worms were added to each plate. In the process of infesting nematodes, the conidia were swallowed by nematodes, after which the spores underwent germination to produce hyphae, which penetrated the body wall of the nematodes and stuck there for further growth to produce new conidia again. If nematodes were present, a new round of the infestation process could be carried out.
Endoparasitic nematophagous fungi can infect nematodes by producing adhesive conidia or ingested conidia. Adhesive conidia were specifically identified and adhered to the body wall, head, and genital pore of the nematode, then grew and reproduced using nematodes. Previous studies have shown that treating the nematode epidermis with pronease E can cause the conidia to fail to adsorb, which leads to the inference that nematode surface proteins play a key role in the inter action process of fungi and nematodes. Similarly, trypsin treatment for conidia eventually resulted in a reduction in the ability of the conidia to adsorb onto nematodes. It has been speculated that the interaction process of nematode and adhesive conidia is a protein-with-protein process of interaction [18]. The process by which ingested conidia infect nematodes is the same as the interaction between H. anguillulae YMF 1.01751 and nematodes, which is divided into swallowing conidia, parasitism, germination, and penetration. This interaction begins when the nematode ingests conidia [9]. In addition to being swallowed, some spores will attach to the surface of the nematode (Figure 2A). The conidia began to germinate and the hyphae grew through nematode body about 48 h after being swallowed (Figure 2B), and some nematode had already begun to die. Subsequently, a lot of hyphae would continue to grow through the body wall (Figure 2C,D) until the nematode mass was finally surrounded (Figure 2E,F), and new conidia were then produced. A new round of infestation was performed in the case of nematodes.
3.2. Structure Identification
Seven compounds (1–7) were isolated from an ethyl acetate extract of broth and a methanol extract of the mycelium of H. anguillulae YMF 1.01751. Their structures were identified by nulear magnetic resonance (NMR) and mass spectrometry (MS) data.
Compound 1 was obtained as colorless solid. The molecular formula of compound 1 is C7H8O4 (157.0494 [M + H]+, Calcd. 157.0495), which was calculated with positive ion mode high-resolution mass spectrometry. It contains four unsaturation degrees. According to 13C-nulear magnetic resonance and distortionless enhancement by polarization transfer (DEPT) (Table 1) data, compound 1 contains one methyl, one oxygen-substituted methylene, one methine, and four quaternary carbons. Based on two-dimensional nuclear magnetic resonance (2D-NMR) data, H-6 (δH 2.36) is correlated with C-5 (δc 141.9) and C-4 (δc 148.7), H-3 (δH 7.59) is related with C-4 (δc 148.7), C-1 (δc 168.2) and C-2 (δc 146.3), and H-7 (δH 3.79) is correlated with C-2 (δc 146.3). Thus, compound 1 was identified as 5-hydroxy-3-(hydroxymethyl)-6-methyl-2H-pyran-2-one, and its structure is shown in Figure 3.
Compounds 2–7 were determined to be (17R)-17-methylincisterol (2) [19], eburicol (3) [20], ergosterol peroxide (4) [21], terpendole C (5) [22], (3β,5α,9β,22E)-3,5-dihydroxy-ergosta-7,22-dien-6-one (6) [23] and 5α,6β-epoxy-(22E,24R)-ergosta-8,22-diene-3β,7α-diol (7) [24]. Their structures are shown in Figure 3.
3.3. The Nematicidal and Chemotaxis Activity of Compounds
Compounds 1, 2, 3, and 5 were tested for their nematicial activity against root-knot nematode M. incognita. The results showed that terpendole C (5) caused 37.4% mortality of M. incognita at 400 μg mL−1 during 48 h. Other compounds have no evident nematicidal activity at the same concentration. Terpendole C (5) is an indole diterpene. This type of compound has been reported to have nematicidal activity. A similar compound, gymnoascole acetate, was obtained from fungus Gymnoascus reessii za-30 and caused 100% paralysis of M. incognita at 36 μg mL−1 for 24 h [25].
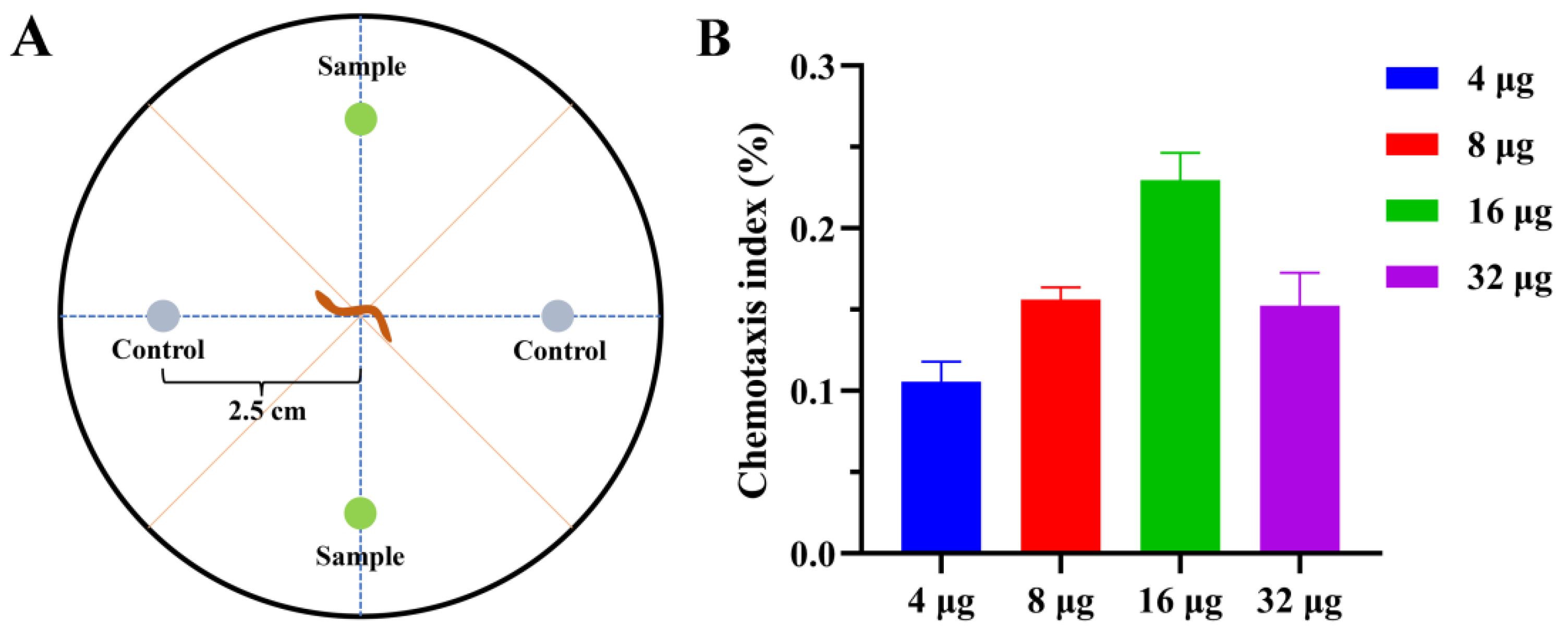
In the chemotaxis assay, the new polyketone 1 exhibited an attractive effect towards P. redivivus. With the concentration increased (4, 8, 16 µg), the attraction activity of compound 1 was enhanced. At 16 µg, it showed a chemotaxis index of 0.21 after 2 h. The attractive effect of compound 1 decreased with increasing concentration (32 µg) after 2 h (Figure 4). The other tested metabolites (1, 2, and 3,) displayed no pronounced activity.
4. Discussion
In this study, the morphology and pathogenicity of H. anguillulae YMF 1.01751 was observed by scanning electron microscopy. It produces abundant crescent-shaped conidia. This kind of long and curved spore is characteristic of Harposporium. Most Herposporiums infect nematodes by ingested spores that germinate inside the nematode and produce septate and branching hyphae that absorb the nutritious of the nematode. At last, the hyphae break out through the nematode cuticle [26].
The fungal metabolites were studied. Seven metabolites, include a new polyketide compound 5-hydroxy-3-(hydroxymethyl)-6-methyl-2H-pyran-2-one (1), were isolated from a fermented extract of H. anguillulae YMF 1.01751. Polyketide 1 exhibited an attractive activity towards P. redivivus. This is the first report about the genus Harposporium secreting small molecular metabolites to attract nematodes. Some biocontrol potential microorganisms can attract nematodes and further kill them. Bacillus nematocida B16 produces 2-heptanone to attract nematodes, subsequently killing them through protease secretion [27].
Other structures of the known metabolites are triterpene, indole diterpene, and steroid. (17R)-17-Methylincisterol (2) was identified from Dictyonella incisa for the first time as a new natural product. It is believed that the biosynthesis pathway of this compound might begin with the unique oxidation process of cholestatrien-3β-ols in vivo [15]. In addition, the compound can also be obtained from the fungus Aspergillus terreus TZS-201607 and has certain cytotoxic activity against A549, MCF-7, and THP-1 with IC50 values of 48.9, 16.8, and 23.6 μM, respectively [28]. Compounds 4, 6, and 7 are sterols. Steroids are a kind of important natural organic compounds that are widespread in biological tissues. With cyclopentane polyhydrophenanthrene as the mother nucleus, steroids can be divided into many types according to their substituents, double-bond positions or stereoconfigurations. Steroids have a variety of biological activities because of their structural specificity, such as antitumor, anti-inflammatory, antibacterial, antiviral, and enzyme-inhibitory activities [29]. Compound 6 showed no activity in an experiment evaluating neutrophil elastase (HNE) activity [30]. Compound 7 exists in a variety of fungi and has moderate cytotoxic activity on tumor cells such as A549 (non-small cell lung adenocarcinoma), SK-OV-3 (ovarian cancer), SK-MEL-2 (skin melanoma), XF498 (central nervous system), and HCT15 (colon) cultured in vitro. In addition, compound 7 can inhibit the growth of the hypocotyl of lettuce and promote the growth of lettuce root at the same concentration [31].
Terpendole C (5) is an indole diterpene, and many indole diterpenoids have been reported to be metabolites of fungi. Terpendole C (5) showed acyl-CoA: cholesterol acyltransferase (ACAT)-inhibitory activity, with an IC50 value of 2.1 μM, in an vitro assay [32]. Terpendole C (5) can specifically inhibit the synthesis of cholesterol esters in peritoneal macrophages by inhibiting cholesterol acyltransferase activity [33]. In addition to enzyme-inhibitory activity, the compound also has cytotoxic activity and antibacterial activity. It has a cytotoxic effect on NCI-H187 and Vero cells, with IC50 values of 35.69 μM and 23.56 μM, respectively, and it has anti-Mycobacterium tuberculosis H37Ra activity, with an MIC value of 25.00 μg mL−1 [34]. According to reports, some indole diterpenes have nematicidal activity. For example, gymnoascole acetate, from G. reessii za-30, has activity against M. incognita, with an EC50 value of 47.5 μg mL−1 for 24 h [25]. This type of metabolite may be a potential biocontrol resource for nematodes. In this study, compound 5 showed weak activity against nematodes. In the isolation process of metabolites from H. anguillulae YMF 1.01751, some other indole diterpenes had not been identified due to their presence in minor amounts. These compounds that have not been isolated may be potential active substances. Therefore, the metabolites of H. anguillulae deserve further study.
H. anguillulae is a typical endoparasitic fungus of nematodes. It infects nematodes by producing ingested conidia. This is the first report on the metabolites of the species. The results state that terpendole C (5) has weak nematicidal activity against root-knot nematode M. incognita, and polyketide 1 exhibites an attractive effect towards the nematode. This result indicated that some secondary metabolites were involved in the pathogenicity process of H. anguillulae infecting nematodes.
Author Contributions
Conceptualization, Z.D. and G.L.; methodology, Z.D., Y.G. and G.L.; software, Z.D.; validation, P.Z. and G.L.; formal analysis, P.Z.; investigation, Z.D.; resources, G.L.; data curation, Y.G. and P.Z.; writing—original draft preparation, Z.D. and G.L.; writing—review and editing, P.Z. and G.L.; supervision, G.L.; project administration, P.Z. and G.L.; funding acquisition, P.Z. and G.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31860015, 31970060 and 32160012) and projects from the Department of Science and Technology of Yunnan Province (202001BB050061).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to Microbial Library of the Germplasm Bank of wild species from Southwest China for providing Harposporium anguillulae YMF 1.01751.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Elling, A.A. Major emerging problems with minor Meloidogyne species. Phytopathology 2013, 103, 1092–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tranier, M.-S.; Pognant-Gros, J.; Quiroz, R.D.; Gonzalez, C.N.A.; Mateille, T.; Roussos, S. Commercial biological control agents targeted against plant-parasitic root-knot nematodes. Braz. Arch. Biol. Technol. 2014, 57, 831–841. [Google Scholar] [CrossRef] [Green Version]
- Nordbring-Hertz, B.; Jansson, H.B.; Tunlid, A. Nematophagous Fungi. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Shimazu, M.; Glockling, S.L. A new species of Harposporium with two spore types isolated from the larva of a cerambycid beetle. Mycol. Res. 1997, 101, 1371–1376. [Google Scholar] [CrossRef]
- Lohde, G. Ueber einige neue parasitische Pilze. Tagebl. Versamml. Dtsch. Nat. Aerzte 1874, 47, 203–206. [Google Scholar]
- Liu, X.Z.; Zhang, K.Q.; Gao, R.H. Nematophagous fungi: Harposporium species from China. Mycosystema 1997, 03, 189–191. [Google Scholar]
- Tian, C.J.; Chen, J.K.; Zhong, Y. Phylogenetic diversity of microbes and its perspectives in conservation biology. Chin. J. Appl. Ecol. 2003, 14, 609–612. [Google Scholar]
- Liu, Z.Q.; Wan, Y.L.; Hao, Y.E. Research advances on resource and biological control of endoparasitic fungi. Chin. J. Appl. Ecol. 2019, 30, 2129–2136. [Google Scholar]
- Li, X.; Luo, H.; Zhang, K.Q. A new species of Harposporium parasitic on nematodes. Can. J. Bot. 2005, 83, 558–562. [Google Scholar] [CrossRef]
- Zheng, Q.M.; Souvanhnachit, S.; Wang, X.; Li, G.H. A new furan derivative from Harposporium sp. YMF 1.01735. Chem. Nat. Compd. 2020, 56, 845–847. [Google Scholar] [CrossRef]
- Lohmann, U.; Sikora, R.A.; HÖfer, M. Influence of phospholipids on growth, sporulation and virulence of the endoparasitic fungi Drechmeria coniospora, Verticillium balanoides and Harposporium anguillulae in liquid culture. J. Phytopathol. 1989, 125, 139–147. [Google Scholar] [CrossRef]
- Charles, T.P.; Roque, M.V.C.; Santos, C.D.P. Reduction of Haemonchus contortus infective larvae by Harposporium anguillulae in sheep faecal cultures. Int. J. Parasitol. 1996, 26, 509–510. [Google Scholar] [CrossRef]
- Eloh, K.; Demurtas, M.; Mura, M.G.; Deplano, A.; Onnis, V.; Sasanelli, N.; Maxia, A.; Caboni, P. Potent nematicidal activity of maleimide derivatives on Meloidogyne incognita. J. Agric. Food Chem. 2016, 64, 4876–4881. [Google Scholar] [CrossRef]
- Dyer, H.C.; Boddy, L.; Preston-Meek, C.M. Effect of the nematode Panagrellus redivivus on growth and enzyme production by Phanerochaete velutina and Stereum hirsutum. Mycol. Res. 1992, 96, 1019–1028. [Google Scholar] [CrossRef]
- Wan, J.; Dai, Z.B.; Zhang, K.Q.; Li, G.H.; Zhao, P.J. Pathogenicity and metabolites of endoparasitic nematophagous fungus Drechmeria coniospora YMF 1.01759 against nematodes. Microorganisms 2021, 9, 1735. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Yu, C.; Shao, Z.; Cai, M.; Li, G.; Zheng, L.; Yu, Z.; Zhang, J. Identification and characterization of nematicidal volatile organic compounds from deep-sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef] [Green Version]
- Hsueh, Y.P.; Gronquist, M.R.; Schwarz, E.M.; Nath, R.D.; Lee, C.H.; Gharib, S.; Schroeder, F.C.; Sternberg, P.W. Nematophagous fungus Arthrobotrys oligospora mimics olfactory cues of sex and food to lure its nematode prey. eLife 2017, 6, e20023. [Google Scholar] [CrossRef]
- Jansson, H.B. Adhesion to nematodes of conidia from the nematophagous fungus Drechmeria coniospora. J. Gen. Microbiol. 1993, 139, 1899–1906. [Google Scholar] [CrossRef] [Green Version]
- Ciminiello, P.; Fattorusso, E.; Magno, S.; Mangoni, A.; Pansini, M. Incisterols, a new class of highly degraded sterols from the marine sponge Dictyonella incisa. Cheminform 1990, 21, 3505–3509. [Google Scholar] [CrossRef]
- Wu, F.E.; Koike, K.; Nikaido, T.; Ishii, K.; Ohmoto, T.; Ikeda, K. Terpenoids and flavonoids from Arenaria kansuensis. Chem. Pharm. Bull. 1990, 38, 2281–2282. [Google Scholar] [CrossRef] [Green Version]
- Yue, J.M.; Chen, S.N.; Lin, Z.W. Sterols from the fungus Lactarium volemus. Phytochemistry 2001, 51, 801–806. [Google Scholar] [CrossRef]
- Huang, X.H.; Nishida, H.; Tomoda, H.; Tabata, N.; Shiomi, K.; Yang, D.J.; Takayanagi, H.; Omura, S. Terpendoles, novel ACAT inhibitors produced by Albophoma yamanashiensis. II. Structure elucidation of terpendoles A, B, C and D. J. Antibiot. 1995, 48, 5–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.P.; Xu, J.; Yue, J.M. Sterols from the fungus Catathelasma imperiale. Chin. J. Chem. 2003, 21, 1390–1394. [Google Scholar] [CrossRef]
- Kwon, H.C.; Sang, D.Z.; Cho, S.Y.; Sang, U.C.; Kang, R.L. Cytotoxic ergosterols from Paecilomyces sp. J300. Arch. Pharm. Res. 2002, 25, 851–855. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Meyer, S.L.F.; Chitwood, D.J.; Chauhan, K.R.; Dong, D.; Zhang, T.T.; Li, J.; Liu, W.C. New nematotoxic indoloditerpenoid produced by Gymnoascus reessii za-130. J. Agric. Food Chem. 2017, 65, 3127–3132. [Google Scholar] [CrossRef] [PubMed]
- Glockling, S.L. Eelworm-eaters the Harposporium and the host. Mycologist 1993, 7, 139–142. [Google Scholar] [CrossRef]
- Niu, Q.H.; Huang, X.W.; Zhang, L.; Xu, J.P.; Yang, D.M.; Wei, K.B.; Niu, X.M.; An, Z.Q.; Bennett, J.W.; Zou, C.G.; et al. A Trojan horse mechanism of bacterial pathogenesis against nematodes. Proc. Natl. Acad. Sci. USA 2010, 107, 16631–16636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Chai, H.; Jia, L.; Shao, L.; Liu, X.; Chen, Y. Antitumor secondary metabolites of an endophytic fungus Aspergillus terreus TZS-201607 from Pseudostellaria heterophylla. Nat. Prod. Res. Dev. 2021, 33, 1156–1164. [Google Scholar]
- Gaulin, E.; Bottin, A.; Dumas, B. Sterol biosynthesis in oomycete pathogens. Plant Signal. Behav. 2010, 5, 258–260. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.-S.; Bae, K.; Yoo, J.K.; Ryoo, I.-J.; Kim, B.Y.; Ahn, J.S.; Yoo, I.-D. Inhibition of human neutrophil elastase by ergosterol derivatives from the mycelium of Phellinus linteus. J. Antibiot. 2012, 65, 437–440. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Kobori, H.; Kawaide, M.; Suzuki, T.; Choi, J.-H.; Yasuda, N.; Noguchi, K.; Matsumoto, T.; Hirai, H.; Kawagishi, H. Isolation of bioactive steroids from the Stropharia rugosoannulata mushroom and absolute configuration of strophasterol B. Biosci. Biotechnol. Biochem. 2013, 77, 1779–1781. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.H.; Tomoda, H.; Nishida, H.; Masuma, R.; Omura, S. Terpendoles, novel ACAT inhibitors produced by Albophoma yamanashiensis. I. Production, isolation and biological properties. J. Antibiot. 1995, 48, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Uchida, R.; Kim, Y.P.; Namatame, I.; Tomoda, H.; Omura, S. Sespendole, a new inhibitor of lipid droplet synthesis in macrophages, produced by Pseudobotrytis terrestris FKA-25. J. Antibiot. 2006, 59, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Sadorn, K.; Saepua, S.; Punyain, W.; Saortep, W.; Choowong, W.; Rachtawee, P.; Pittayakhajonwut, P. Chromanones and aryl glucoside analogs from the entomopathogenic fungus Aschersonia confluens BCC53152. Fitoterapia 2020, 144, 104606. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Morphology of H. anguillulae YMF 1.01751. (A–C) Phialides produced by H. anguillulae YMF 1.01751 (red arrow: phialides). (D) Conidia produced by H. anguillulae YMF 1.01751 (orange arrow: conidia).
Figure 1.
Morphology of H. anguillulae YMF 1.01751. (A–C) Phialides produced by H. anguillulae YMF 1.01751 (red arrow: phialides). (D) Conidia produced by H. anguillulae YMF 1.01751 (orange arrow: conidia).
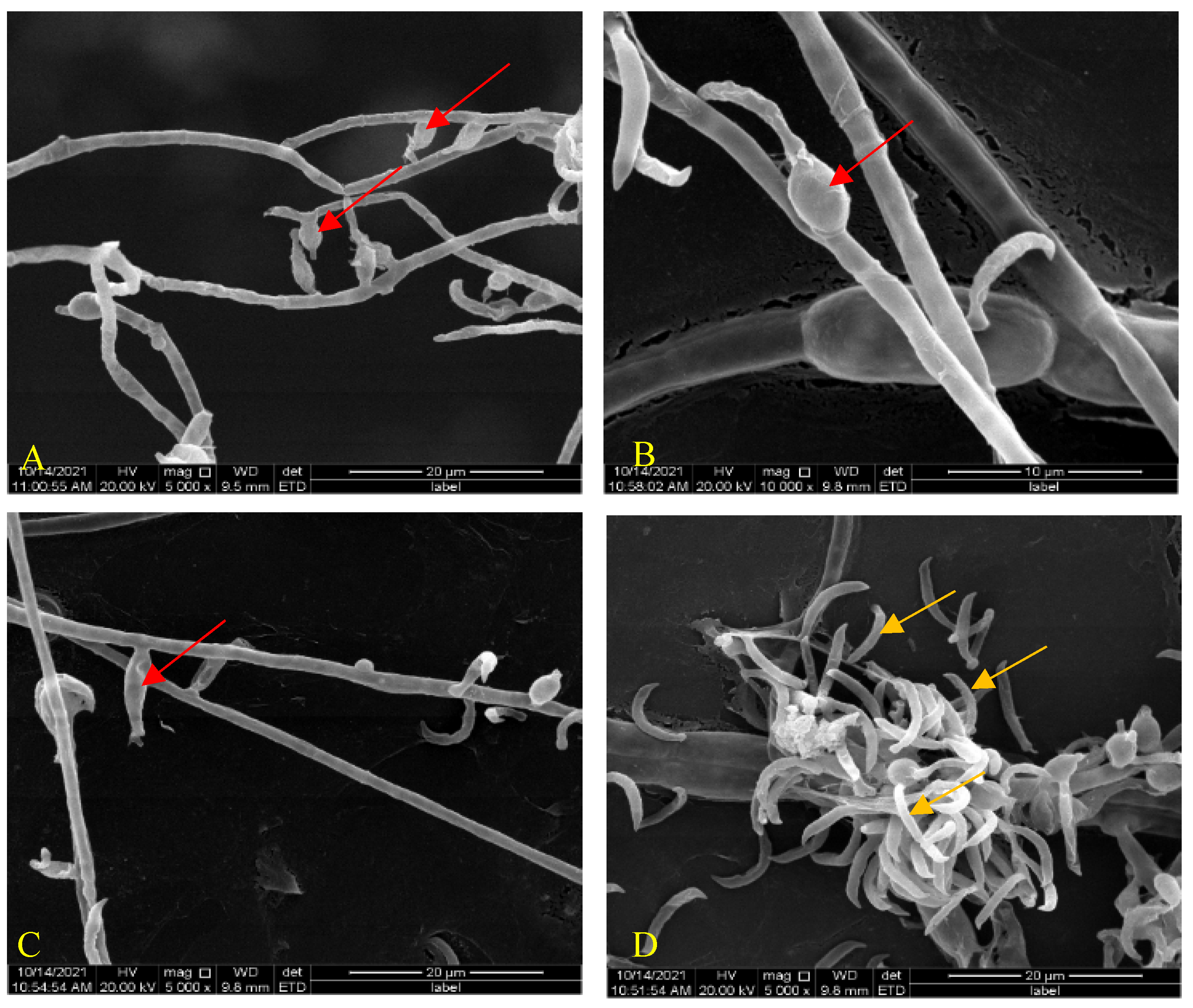
Figure 2.
Scanning electron microscopy observation of the interaction between H. anguillulae YMF 1.01751 and P. redivivus. (A) The coculture of H. anguillulae YMF 1.01751 and P. redivivus. (B) The hyphae begin to grow through nematode body. (C,D) A lot of hyphae continue to grow through the body wall. (E,F) The nematode was finally surrounded by hyphae.
Figure 2.
Scanning electron microscopy observation of the interaction between H. anguillulae YMF 1.01751 and P. redivivus. (A) The coculture of H. anguillulae YMF 1.01751 and P. redivivus. (B) The hyphae begin to grow through nematode body. (C,D) A lot of hyphae continue to grow through the body wall. (E,F) The nematode was finally surrounded by hyphae.
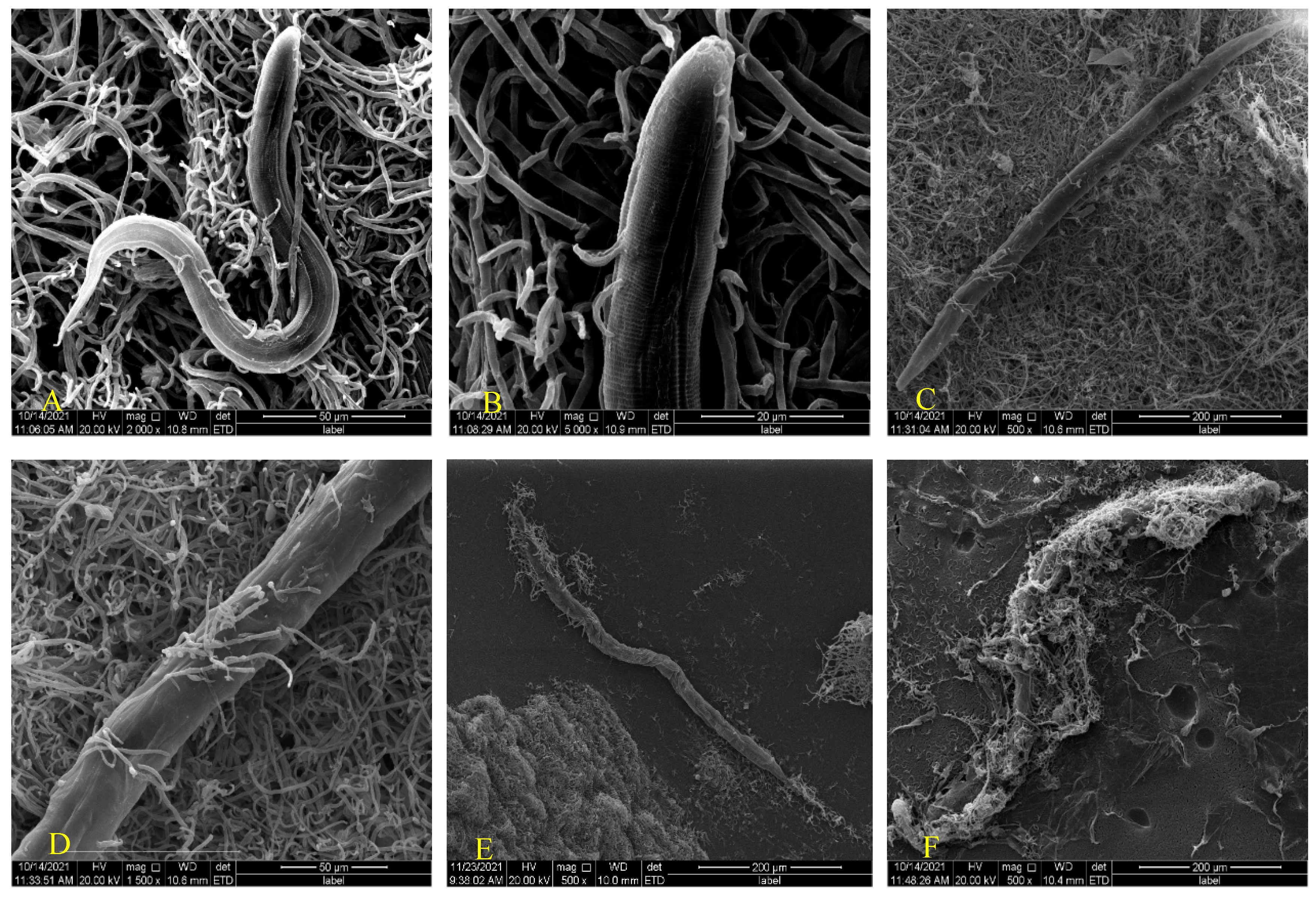
Figure 3.
The compounds isolated from H. anguillulae YMF 1751.

Figure 4.
Effect of compounds on the chemotaxis of P. redivivus. (A) Schematic representation of the quadrant bioassay used to measure the chemotaxis of compounds. (B) Chemotaxis activity of 1 at different concentrations.
Figure 4.
Effect of compounds on the chemotaxis of P. redivivus. (A) Schematic representation of the quadrant bioassay used to measure the chemotaxis of compounds. (B) Chemotaxis activity of 1 at different concentrations.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The nuclear magnetic resonance data of compound 1 in trichloromethane-d (600 MHz).
| Position | 1H | 13C | HMBC |
|---|---|---|---|
| 1 | - | 168.2, s | - |
| 2 | - | 146.3, s | - |
| 3 | 7.59 (1H, s) | 138.3, d | 148.7, 168.2, 146.3 |
| 4 | - | 148.7, s | - |
| 5 | - | 141.9, s | - |
| 6 | 2.36 (3H, s) | 14.6, q | 141.9, 148.7 |
| 7 | 3.79 (2H, s) | 57.0, t | 146.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dai, Z.; Gan, Y.; Zhao, P.; Li, G. Secondary Metabolites from the Endoparasitic Nematophagous Fungus Harposporium anguillulae YMF 1.01751. Microorganisms 2022, 10, 1553. https://doi.org/10.3390/microorganisms10081553
AMA Style
Dai Z, Gan Y, Zhao P, Li G. Secondary Metabolites from the Endoparasitic Nematophagous Fungus Harposporium anguillulae YMF 1.01751. Microorganisms. 2022; 10(8):1553. https://doi.org/10.3390/microorganisms10081553
Chicago/Turabian StyleDai, Zebao, Yang Gan, Peiji Zhao, and Guohong Li. 2022. "Secondary Metabolites from the Endoparasitic Nematophagous Fungus Harposporium anguillulae YMF 1.01751" Microorganisms 10, no. 8: 1553. https://doi.org/10.3390/microorganisms10081553
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.