
Roles of Hcp2, a Hallmark of T6SS2 in Motility, Adhesive Capacity, and Pathogenicity of Vibrio alginolyticus

1
Guangdong Provincial Key Laboratory of Aquatic Animal Disease Control and Healthy Culture, College of Fishery, Guangdong Ocean University, Zhanjiang 524088, China
2
Key Laboratory of Diseases Controlling for Aquatic Economic Animals of Guangdong Higher Education Institutions, College of Fishery, Guangdong Ocean University, Zhanjiang 524088, China
3
Central People’s Hospital of Zhanjiang, Zhanjiang 524045, China
*
Authors to whom correspondence should be addressed.
Microorganisms 2023, 11(12), 2893; https://doi.org/10.3390/microorganisms11122893
Submission received: 15 November 2023
/
Revised: 28 November 2023
/
Accepted: 29 November 2023
/
Published: 30 November 2023
(This article belongs to the Special Issue Vibrio Virulence)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The type VI secretion system (T6SS) is a large secretory device, widely found in Gram-negative bacteria, which plays important roles in virulence, bacterial competition, and environmental adaptation. Vibrio alginolyticus (V. alginolyticus) is an opportunistic pathogen that causes vibriosis in aquaculture animals. V. alginolyticus possesses two type VI secretion systems (named the T6SS1 and T6SS2), but their functions remain largely unclear. In this paper, the roles of the core component of the T6SS2 cluster of V. alginolyticus HY9901, hemolysin-coregulated protein2 coding gene hcp2, are reported. Deletion of hcp2 clearly impaired the swarming motility, adhesive capacity, and pathogenicity of V. alginolyticus against zebrafish. Furthermore, transmission electron microscopy (TEM) found that the abnormal morphology of flagellum filament in the hcp2 mutant strain could be partially restored by hcp2 complementarity. By proteomic and RT-qPCR analysis, we confirmed that the expression levels of flagellar flagellin and assembly-associated proteins were remarkably decreased in an hcp2 mutant strain, compared with the wild-type strain, and could be partially restored with a supply of hcp2. Accordingly, hcp2 had a positive influence on the transcription of flagellar regulons rpoN, rpoS, and fliA; this was verified by RT-qPCR. Taken together, these results suggested that hcp2 was involved in mediating the motility, adhesion, and pathogenicity of Vibrio alginolyticus through positively impacting its flagellar system.
1. Introduction
The type VI secretion system (T6SS), an inverted bacteriophage-like complex that was first defined in Vibrio cholerae [1], plays key roles in the infection of eukaryotic cells [2] and bacterial competition [3,4]. V. alginolyticus has two T6SS gene clusters (T6SS1 and T6SS2) [5] and encodes homologs of 13 characteristic core proteins [6,7]. One of these components, hemolysin coregulated protein (Hcp), is the hallmark of the T6SS and serves as a key effector for bacterial pathogenicity [8,9,10]. Although Hcp proteins share a highly similar structure in various bacteria, the functions of Hcp proteins are distinct. Recently, it is reported that Hcp is involved in mediating T6SS biofilm formation, bacterial adhesion, virulent regulation, environmental stress adaption, etc. [11,12,13]. More importantly, there are reports showing that the deletion of the T6SS genomic island (GI) or its other core components, such as Hcp, will result in less-motile, lower-virulence bacteria than wild-type strains by regulating the flagellar system at the levels of transcription and translation in several species [14,15,16,17].
Bacterial motility relies on the polar flagellum, which is composed of three major structural components: the basal body, the hook, and the filament [18]. Polar flagellar biosynthesis and assembly is a tightly transcriptional hierarchy process regulated by four promoters, named Class I~IV in the Vibrio species [19]. During flagellar assembly, flagellar gene expression is controlled by several master regulons. RpoN (σ54), a type of transcriptional regulatory factor, is widely identified in pathogenic bacteria. RpoN can bind with core RNA polymerase (RNAP) and regulate gene transcription [20,21]. Recent reports have shown that RpoN is essential for biofilm detachment, flagellar formation, motility, and virulence of V. alginolyticus and other pathogens [22,23,24]. RpoN also affects the flagellar biogenesis-dependent motility in Vibrio spp. by regulating RpoN-dependent gene transcription [19]. Furthermore, the Class IV flagellar genes’ transcription of Vibrio spp. are regulated by RpoN-dependent activation of alternative sigma factors FliA (σ28) [25]. Another necessary sigma factor for flagellum regulation is the general stress response regulator RpoS (σ38), which is considered as the antagonist of RpoN and negatively controls flagellar genes’ expression and motility in bacteria [26,27,28].
V. alginolyticus is one of the major pathogens that leads to the vibriosis of aquatic animals and severe food safety concerns. However, except for its mediation of interbacterial competition [5,7], other roles of the Vibrio alginolyticus T6SS are still largely unclear. In this study, the impacts of hcp2 on bacterial swarming motility, adhesive ability, and virulence against zebrafish were analyzed by constructing an hcp2 mutant strain of V. alginolyticus HY9901. Furthermore, the proteomic and real-time fluorescence quantitative results confirmed the positive roles of hcp2 on flagellar assembly. These data will be beneficial to further explore the pathogenicity of V. alginolyticus.
2. Materials and Methods
2.1. Bacterial Strains, Media, Plasmids, and Zebrafish Lines and Maintenance
The bacterial strains, plasmids, and zebrafish lines used in this study are shown in Supplementary Material Table S1. Vibrio alginolyticus strain HY9901 was kept in our laboratory [29]. V. alginolyticus was grown in tryptic soy broth (TSB, Huankai, Guangzhou, China) at 28 °C. Escherichia coli S17 strains were cultured in Luria broth (LB, Huankai, Guangzhou, China) at 37 °C. Adult AB zebrafish line purchased from China Zebrafish Resource Center (CZRC) was maintained under standard conditions [30]. Handling of zebrafish complied with “The Legislation of Guangdong Laboratory Animal Management Regulations (2010)”.
2.2. Construction and Physiology Analysis of the hcp2 Mutation
The primers for the construction of the hcp2 mutation were designed based on the sequence of hcp2 gene of V. alginolyticus strain HY9901 cloned by our laboratory (Supplementary Material Table S2). The PCR-amplified DNA fragment used for constructing the in-frame deletion mutation of hcp2 was generated by overlap PCR [31]. Briefly, two PCR fragments were obtained from V. alginolyticus strain HY9901 genomic DNA with the primer pairs of hcp2-MF1/ hcp2-MR1 and hcp2-MF2 /hcp2-MR2 by using PrimeSTAR Max DNA Polymerase (TaKaRa, Kusatsu City, Japan). The resulting products generated a 707 bp fragment containing the DNA upstream of hcp2 and a 650 bp fragment containing the DNA downstream of hcp2. A 20 bp overlap in the sequences permitted amplification of a 1357 bp product containing a deletion from nucleotides 58 to 447 of hcp2 during a second PCR with primers of hcp2-MF1 and hcp2-MR2. The resulting product was linked into the suicide plasmid pLP12 by Exnase II recombinase (ClonExpress II, Vazyme, Nanjing, China) and the recombinant product was transformed into E. coli DH5α cultured with LB solid media containing 20 μg/mL chloramphenicol and 0.3% D-Glucose. The recombinant suicide plasmid pLP12-hcp2 was confirmed by PCR with primers of pLP-UF/pLP-UR and subsequently transformed into E. coli β2163. E. coli β2163 containing the plasmid pLP12-hcp2 was conjugated with wild-type V. alginolyticus strain HY9901 and co-incubated with LB plate containing 0.3 mM diaminopimelic acid (DAP) and 0.3% D-Glucose. Recipient cells were plated on LB supplemented with 0.3% D-Glucose and the antibiotics chloramphenicol to select the clone pLP12-hcp2 that had integrated the vector by homologous recombination. Antibiotic-resistant colonies were isolated, identified with the primers of hcp2-TF/pLP-UTR, grown in LB to the late logarithmic phase, and plated on counter-selective LB containing 0.4% (w/v) L-arabinose. The selected colonies were confirmed by PCR using the primers of hcp2-TF/ hcp2-TR and direct sequencing.
Growth ability and swarming motility assay were measured according to the method of Zhou et al. [32] and Moisi et al. [33], respectively, with a slight modification. Briefly, to measure the growth level of bacteria in TSB, overnight cultures of V. alginolyticus were adjusted to OD600 of 0.5 and subsequently inoculated into TSB with a dilution rate of 1:100 (v/v). Samples were removed every 1 h, and the optical density was measured at 600 nm. To measure swarming motility, the colony of V. alginolyticus was spotted on TSA plate containing 0.3% agar and 3% sodium chloride. The plate was analyzed after at least 12 h of incubation at 28 °C.
2.3. Virulence Determination
The wild-type and hcp2 mutation of V. alginolyticus were prepared as follows. Briefly, V. alginolyticus strains were cultured at 28 °C in TSB medium for 18 h. The overnight cultures were adjusted to OD600 of 0.5, subsequently inoculated into TSB with a dilution rate of 1:100 (v/v), and incubated at 28 °C with rotation until they reached the exponential phase. Then, the number of colonies were determined by flat colony counting method. For larval infection assay, 4 dpf (days post-fertilization) zebrafish were randomly transferred into 6-well plates. Each well contained 8~10 fish with 10 mL embryo medium, and the fish were immersed in 108 CFU of V. alginolyticus strains; then, bacteria and fish were kept at 28 °C for 2~3 d. From 5 dpf, all the fish were fed by paramecium to maintain normal growth. The control larvae were raised normally without V. alginolyticus strains. For adult zebrafish challenged assay, 7-month-old fish were randomly transferred into 4 groups and were infected by intraperitoneal injection of 108, 107, 106, and 105 CFU of V. alginolyticus strains, respectively. As a control, the fish were injected with sterilized 10% (g/mL) NaCl. All the adult fish were kept at 28 °C for 12 hpi~14 hpi (hours post-injection), and mortality was recorded. Medium lethal dose (LD50) was used to estimate the bacterial virulence in zebrafish via LD50 data processing software 1.01 published by Blue Cosmos Studio.
2.4. Adhesion and Intestinal Colonization Assays
The overnight cultures were adjusted to OD600 of 0.5, subsequently inoculated into TSB with a dilution rate of 1:100 (v/v), and incubated at 28 °C with a rotation speed of 125 rpm until they reached the exponential phase. Plate colony counting method was used to determine bacterial concentration (CFU/mL). The cultures were centrifuged at 4000 rpm for 2 min, and the cells were resuspended with 5 ug/mL CellTracker™ CM-DiI dye (C7000, Invitrogen, Waltham, MA, USA) in 10% NaCl. Bacterial suspension was incubated at 37 °C for 5 min then at 4 °C for 15 min (low-temperature incubation helps to reduce bacterial endocytosis of dyes and facilitates the labeling of plasma membranes). After labelling was finished, bacteria were washed twice and resuspended in 10% NaCl. For adhesion experiment, 7~10 of 5 dpf transgenic zebrafish Tg (coro1a: eGFP) were co-incubated with 107 and 108 CFU of dye-marked bacteria at 28° for 42 hpi (hours post-immersion). Similarly, 5 dpf AB line zebrafish were challenged by 108 CFU dye-labelled Vibrio alginolyticus for 3 dpi (days post-immersion) and 5 dpi, respectively. Subsequently, the zebrafish were washed with PBS 3 times and imaged by a Nikon upright fluorescence microscope (Ni-E, Tokyo, Japan).
2.5. Construction of hcp2 Complementary Strain and Electron Microscopy Imaging
To create a complemental plasmid for the hcp2 mutation, the hcp2 gene was amplified by primers of comhcp2-F and comhcp2-R (Supplementary Material Table S2), and then the fragment obtained was cloned into PstI and XhoI sites of the pBBR1-MCS1 to construct pBBR1-MCS1-hcp2. The resulting plasmid was first transformed into E. coli DH5α and plasmid DNA was isolated. The correct pBBR1-MCS1-hcp2 plasmid was transformed into E. coli S17-1 λpir. S17-1 λpir harboring the plasmid pBBR1-MCS1-hcp2 was conjugated with hcp2 mutation and co-grown with TSA overnight without antibiotics. The mixed culture was plated on TCBS with chloramphenicol, which only permitted vibrio spp. proliferation. The selected colonies were confirmed further by PCR analysis and sequencing. The constructing method of wild-type HY9901 and hcp2 mutation harboring the empty vector pBBR1-MCS1 was the same as the complementary strain.
To observe flagellar morphology, V. alginolyticus strains of overnight cultures were negatively stained with 1% phosphotungstic acid (pH 7.4) on a Formvar carbon-coated grid and scanned by a transmission electron microscope (JEM-1400, Tokyo, Japan) [34]. Flagellar filament width was measured on electron micrographs recorded at a magnification of 30,000. For the phenotype observed, zebrafish were embedded with 1% (m/v) low-melting point agarose and imaged with a Nikon upright fluorescence microscope (Ni-E, Japan).
2.6. Proteomics Analysis of the Proteins Associated with Flagellum Assembly
2.6.1. Protein Extraction, Digestion, TMT/iTRAQ Labeling, and HPLC Fractionation
For protein extraction, after overnight cultures were adjusted to OD600 of 0.5, wild-type HY9901 and Δhcp2 strains were inoculated into TSB with a dilution of 1:100 for 18 h of incubation, respectively, and then the cells were collected by centrifugation at 4000× g for 5 min at 4 °C. The sediments were sonicated in lysis buffer (8 M urea, 1% protease inhibitor cocktail). The remaining debris was removed by centrifugation at 12,000× g at 4 °C for 10 min. The supernatant was collected, and the protein concentration was determined with BCA kit according to the manufacturer’s instructions. For digestion, the protein solution was reduced with 5 mM dithiothreitol for 30 min at 56 °C and alkylated with 11 mM iodoacetamide for 15 min at room temperature in darkness. The protein sample was then diluted by adding 100 mM TEAB to urea concentration less than 2 M. Trypsin was added at 1:50 trypsin-to-protein mass ratio for the first digestion overnight and 1:100 trypsin-to-protein mass ratio for a second 4 h digestion. After trypsin digestion, the peptide was desalted by Strata X C18 SPE column (Phenomenex, Hong Kong, China) and vacuum dried. The peptide was reconstituted in 0.5 M TEAB and processed according to the manufacturer’s protocol for TMT kit/iTRAQ kit. The peptide mixtures were then incubated for 2 h at room temperature and pooled, desalted, and dried by vacuum centrifugation. For purification, the tryptic peptides were fractionated into fractions by high pH reverse-phase HPLC using Agilent 300Extend C18 column (5 μm particles, 4.6 mm ID, 250 mm length) (Santa Clara, CA, USA). Then, the peptides were combined into 18 fractions and dried by vacuum centrifuging.
2.6.2. LC-MS/MS Analysis
The tryptic peptides were dissolved in 0.1% formic acid (solvent A) and directly loaded onto a homemade reversed-phase analytical column (15 cm length, 75 μm i.d.). The gradient comprised an increase from 6% to 23% solvent B (0.1% formic acid in 98% acetonitrile) over 26 min, 23% to 35% in 8 min, rose to 80% in 3 min, and then was maintained at 80% for the last 3 min. All this occurred at a constant flow rate of 400 nL/min on an EASY-nLC 1000 UPLC system. The peptides were subjected to NSI source followed by tandem mass spectrometry (MS/MS) in Q ExactiveTM Plus (Thermo, Waltham, MA, USA) coupled online to the UPLC. The electrospray voltage applied was 2.0 kV. The m/z scan range was 350 to 1800 for the full scan, and intact peptides were detected in the Orbitrap at a resolution of 70,000. Peptides were then selected for MS/MS using NCE setting of 28 and the fragments were detected in the Orbitrap at a resolution of 17,500. A data-dependent procedure that alternated between one MS scan followed by 20 MS/MS scans with 15.0 s dynamic exclusion was carried out. Automatic gain control (AGC) was set at 5 × 104. The fixed first mass was set as 100 m/z.
2.6.3. Volcano Plot Analysis
To analyze the differential expression of flagellar proteins, a volcano plot was generated based on the proteome data. The threshold of fold change (FC) ratio and p-value were set as above 1.5 and less than 0.05, respectively, which were considered as statistically different. The flagellum protein level between wild-type HY9901 and Δhcp2 was quantified by the LFQ intensity value from proteome data.
2.7. Real-Time Quantitative PCR (RT-qPCR)
For bacteria, adjusted to OD600 of 0.5 in overnight cultures, V. alginolyticus strain HY9901 harboring the empty vector PBBR1-MCS1 (HY9901-ev), HY9901 Δhcp2 strain harboring the empty vector (Δhcp2-ev), and Δhcp2 complementary strain (Δhcp2-com), respectively, were inoculated on TSB with a dilution of 1:100 and incubated at 28 °C with agitation for 18 h. Total RNA was extracted and purified using RNAprep Pure Cell/Bacteria Kit (TIANGEN, Beijing, China) according to the manufacturer’s instructions. Reverse transcription PCR was conducted by using a PrimeScript™ RT reagent Kit (Takara, Kusatsu City, Japan) with 500 ng of total RNA in each reaction according to the manufacturer’s instructions. RT-qPCR was performed using LightCycler 480 detection system (Roche, Penzberg, Germany) with a PerfectStart® Green qPCR SuperMix (+Dye I) (TransGen, Beijing, China).
Zebrafish were challenged with V. alginolyticus for 43 h at the beginning of 4 dpf; then, the larvae were collected and washed with PBS thrice. Total zebrafish RNA was extracted using Trizol regent (Invitrogen, Waltham, MA, USA) and then was reverse transcribed into cDNA with PrimeScript RT Reagent Kit (TaKaRa, Kusatsu City, Japan) according to the manufacturer’s instructions. RT-qPCR was performed using a LightCycler 480 detection system (Roche, Penzberg, Germany) with a PerfectStart® Green qPCR SuperMix (+Dye I) (TransGen, Beijing, China). All the sequences of RT-qPCR primers are listed in Supplementary Material Table S2.
2.8. Statistical Analysis
The results are presented as means ± standard error (mean ± SEM). All the experiments were conducted at least 3 times. Two-tailed Student’s t-test (for two groups) or one-way analysis of variance (ANOVA followed by Tukey’s test for three or more groups) were applied to determine statistical significance with Prism 5 software (GraphPad Software). Confidence intervals of 95% for data analyses were acquired for all experiments. p-value less than 0.05 (p < 0.05) was considered significant.
3. Results
3.1. Loss of hcp2 Reduced the Swarming Motility of Vibrio alginolyticus
Hcp, a hallmark component of the T6SS, was initially identified as a secreted protein of Vibrio cholerae [35]. The current study revealed that Hcp also functions as an exported effector and chaperone of Type VI secretion substrates in Gram-negative bacteria [2,10,36]. Hcp has been proven to be involved in bacterial motility, adherence, and cytotoxicity to host cells by regulating the flagellar system in various pathogens [12,13,15,16]. However, for Vibrio alginolyticus, the roles of Hcp on bacterial motility remains unknow. Using the suicide plasmid-mediated homologous recombination, a hcp2 deletion mutation (Δhcp2) was constructed. Using a housekeeping gene, 16s RNA, as control for normalization, the RT-PCR result confirmed that the hcp2 transcription level of the mutation was clearly decreased compared to HY9901 wild-type (Figure 1A). The mutant showed a similar growth level to the wild-type strain cultured in TSB (Figure 1B). Meanwhile, notable impairment of the swarming motility was observed in the Δhcp2 of V. alginolyticus compared with wild-type HY9901 (Figure 1C,D; p < 0.05). To further clarify the effect of hcp2 on motility, a Δhcp2 complementary strain was constructed and confirmed by RT-qPCR (Figure 1E,F). As expected, supplementation with hcp2 partially restored the swarming ability in the Δhcp2 strain (Figure 1G).
3.2. Attenuated Pathogenicity of Δhcp2 Strain against Zebrafish
As mentioned above, Hcp family proteins have been proved to participate in modulating pathogenicity in many pathogens. To compare the virulent differences between the wild-type strain HY9901 and the Δhcp2 strain, larval and adult zebrafish were challenged with V. alginolyticus. After 43 hpi (hours post-immersion) of 4 dpf AB larvae, more severe edema in the zebrafish larval belly (red asterisk) and gut (red arrow) were observed compared to after the zebrafish were challenged by the wild-type strain then immersed in Δhcp2 counterparts (Figure 2A). And then, around 69 hpi, those larvae infected by the Δhcp2 strain had a lower mortality rate compared to those fish challenged by wild-type bacteria (Figure 2B). Consistently, a red and swollen phenotype on the belly was observed in adult zebrafish injected with V. alginolyticus, and IC50 value was significantly increased in the Δhcp2 infection group (Figure 2C,D).
3.3. Reduced Adhesive Ability of Δhcp2 Strain on the Zebrafish larval Surface and Intestinal Tract
Motility is a fundamental function of bacteria and is responsible for the initial colonization to the host. Before that, adhesive capacity on the host is the first event in bacterial infection. To clarify that the attenuated virulence of the Δhcp2 strain was partially related to reduced adhesion, a comparison of adhesive capacity on zebrafish was performed between Δhcp2 and HY9901 wild-type strains. We found a clear decrease of red dye-labelled Vibrio alginolyticus of the Δhcp2 strain on the whole zebrafish trunk surface and tail at 5 dpf after 42 hpi treatment (Figure 3A vs. Figure 3B, Figure 3C vs. Figure 3D). Interestingly, the dye-labelled strains seemed to be surrounded by GFP-labelled macrophage in the zebrafish intestinal canal (white rectangle in Figure 3), which indicated that the phenotype of edema from zebrafish abdomen and gut displayed in Figure 2 was indeed caused by Vibrio alginolyticus infection. To further confirm whether the Δhcp2 strain was more easily excreted from the intestinal canal of the zebrafish as its motility was impaired, 5 dpf AB larvae were infected with dye-labelled Vibrio alginolyticus. Consistent with what we observed in a transgenic zebrafish assay, the hcp2 mutant bacteria were weakened in colonization (Figure 3E–I’) and easily excreted to the outside of zebrafish canal by the intestinal movement (white arrow in Figure 3).
3.4. A Malformed Morphology of Flagellar Filaments in hcp2 Mutation of Vibrio alginolyticus
The bacterial polar flagellum is essential for mediating motility, adhesion, and pathogenicity. To further verify whether hcp2 influenced the flagellar morphology and resulted in the disablement of motility, negative-staining TEM scanning was performed. As shown in Figure 4B, a severely hollow-like structure of flagellar filament was observable in the Δhcp2-ev strain compared with the HY9901-ev strain (Figure 4A), and this morphology of filament could be rescued partially by the complement of hcp2 in the mutant strain (Figure 4C). Then, the outer diameter of each flagellar filament was measured based on the TEM data. The result showed that the outer diameter of filament of the Δhcp2-ev mutation was significantly enlarged (as shown in Figure 4D, the mean ± standard error of filament width in each of the strain Δhcp2-ev was 49.5 ± 2.5 nm vs. 36.48 ± 1.5 nm in wild-type HY9901-ev; p < 0.001). Nevertheless, there was no difference in the outer diameter of filament between the Δhcp2-ev and the Δhcp2-com strain (mean ± standard error of the filament width in Δhcp2-ev bacteria were 49.5 ± 2.5 nm vs. 48.51 ± 0.66 nm in the Δhcp2-com strain; p > 0.05).
3.5. Expression and Transcription Levels Analysis of Flagellin and Flagellum Export Apparatus by Quantitative Proteomics and RT-qPCR, Respectively
Emerging studies have demonstrated the potential regulatory role of the T6SS on the bacterial flagella system [37]. A study of Citrobacter freundii [15], Erwinia amylovora [38], Pseudomonas genus [39], and Vibrionaceae [40] recorded the involvement of the T6SS in modulating flagellar gene expression and secretion. Consider that the normal assembly of polar flagella is crucial for motile flagellated bacteria [41]. Thus, the effect of hcp2 on bacterial flagellar assembly was assessed. Our previous studies confirmed the flagellin of V. alginolyticus HY9901 are composed of three flagellum genes encoded FlaA, FlaB, and FlaC [42,43]. Moreover, flagellin from V. alginolyticus HY9901 has been proven to be one of the important factors for bacterial intestinal colonization [44]. By proteomic analysis, the differential expression proteins were significantly enriched in the processes of bacterial-type flagellum organization, assembly, and flagellum-dependent cell motility (red rectangle in Figure 5A). All the identified proteins associated with flagellar assembly were analyzed by volcano plot, in which the differential proteins were marked (Figure 5B). The intracellular protein levels of flagellin FlaA, FlaB, and FlaC were remarkably decreased in the Δhcp2 strain (Figure 5B,C). Furthermore, the expression levels of flagellar type III secretion system (fT3SS)-associated protein FliH, MS ring protein FliF, and flagellum hook protein FlgE were also decreased in the mutant strain (Figure 5D). In RT-qPCR assay, the transcriptional levels of flaA, flaB, and flaC genes were significantly reduced in the Δhcp2 strain and were partially restored by supplying hcp2 (Figure 5E).
3.6. The Transcriptions of Sigma Factors rpoN and fliA Controlling Flagellin Biosynthesis and Export Were Positively Influenced by hcp2 in Vibrio alginolyticus
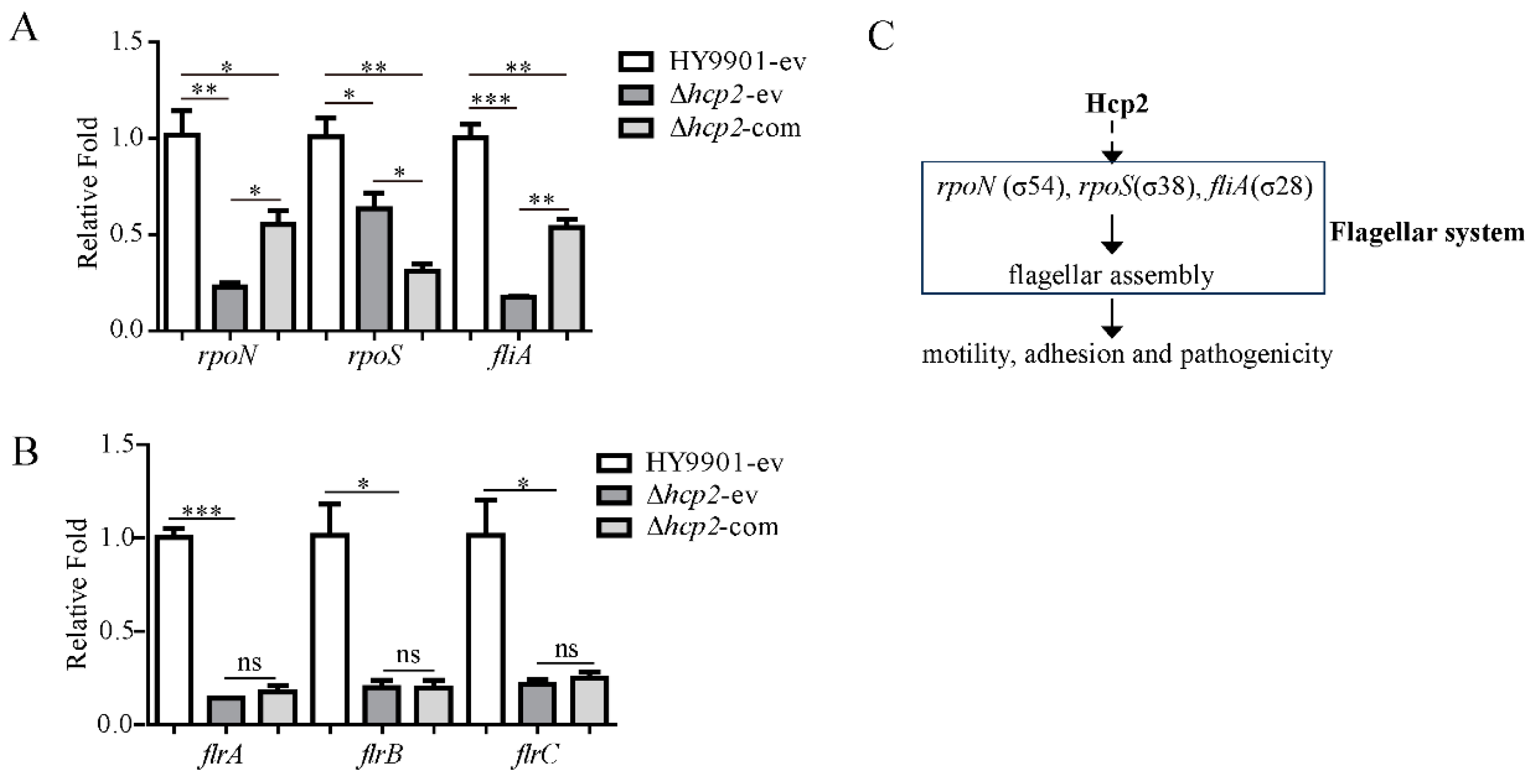
In bacteria, sigma factors RpoN, RpoS, and FliA control flagellar gene transcriptional activation [41,45], but RpoS and RpoN are reciprocal function factors for controlling flagellar gene expression in Escherichia coli and Edwardsiella tarda [27,46,47]. FlrA is another important master regulon that initiates flagellar gene expression in an RpoN-dependent manner, which is required for proper flagellation and includes the polar flagellar number and morphology related to bacterial motility [48]. Additionally, another study found that flagellin mutants or flrA-, flrB-, or flrC-silenced strains affect filament morphology and the swimming, adhesion, and biofilm formation ability for its pathogenesis in Stenotrophomonas maltophilia and Vibrio alginolyticus [49,50]. Hence, we investigated whether the transcriptional levels of these flagellar regulons were affected by hcp2. The RT-qPCR result showed that hcp2 deletion resulted in transcriptional reduction of sigma factors rpoN, fliA, and rpoS. Their mRNA levels were partially restored in the Δhcp2 complementary strain (Figure 6A). Meanwhile, there was no significant transcriptional restoration of the regulators flrA, flrB, and flrC in the complementary strain, though their mRNA levels were reduced in hcp2 mutation (Figure 6B), which indicated that the synthesis and export of flagellin FlaA, FlaB, FlaC were mainly modulated by rpoN and fliA. This finding was consistent with previous reports. Taken together, the results suggested that hcp2 played positive and vital roles on bacterial motility, adhesive ability, and pathogenicity mainly through affecting the transcription of sigma factors in Vibrio alginolyticus HY9901 (Figure 6C).
4. Conclusions
As the hallmark component of the T6SS, hemolysin-coregulated protein (Hcp) is required for functional T6SS, which has a regulatory role in flagellar assembly and bacterial pathogenesis in Gram-negative bacteria. In this study, the roles of hcp2 in Vibrio alginolyticus HY9901 were investigated by constructing a mutant strain with an in-frame deletion of the hcp2 gene. The results suggested that Vibrio alginolyticus Hcp2 protein was involved in bacterial motility ability, adhesive capacity, and virulence through affecting the expression levels of flagellin and flagellar export proteins. Further RT-qPCR results indicated that hcp2 had a positive influence on the transcriptional levels of flagellar assembly-associated sigma factors. flrA transcription was not clearly restored in the Δhcp2 complementary strain, suggestion that the effect of Hcp2 on the motility was independent of the FlrA-RpoN pathway and implying the possible involvement of other regulatory mechanisms in flrA expression in V. alginolyticus. For example, the two-component system, a crucial bacterial signal transduction system for modulating gene expression (in genes such as flrA, flrB, and flrC) to maintain the balance in the amount of regulator input and output via feedback mechanisms, may be involved [51]. A previous study has shown a novel role for Hcp which directly interacts with the RpoN-dependent T6SS regulator VasH to quickly adjust T6SS expression in several pathogens [52]. Therefore, there might be unrecognized Hcp2–protein interaction for mediating flagellar regulons’ expression in Vibrio alginolyticus. This study strengthened the evidence for the regulatory role of the T6SS on flagellar assembly and expanded our understanding of the pathogenesis of Vibrio alginolyticus.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms11122893/s1. Table S1: Plasmids and bacterial strains used in this study; Table S2: Sequences of primers used in this study.
Author Contributions
J.J. and J.C.: conceptualization. S.W., J.T. and B.W.: methodology, investigation, and data curation. S.W. and J.C.: formal analysis and validation. J.J. and J.C.: resources. J.J. and J.C.: supervision and project administration. J.J.: funding acquisition. S.W. and J.C.: writing—original draft preparation. J.C.: writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (U20A2065, 32002426, 32073006); Natural Science Foundation of Guangxi Province (2020GXNSFAA297243); Natural Science Foundation of Guangdong Province (2022A1515010553); and Competitive Allocation Project of Special Fund for Science and Technology Development in Zhanjiang City (2022A01090).
Data Availability Statement
All data in this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Kwaku Amoah for critical reading and grammar correction of the manuscript, Wenxin Liu (Central People’s Hospital of Zhanjiang) for the guide of experiment methods, and Sijie Wang (Affiliated Hospital of Guangdong Medical University) for assistance in the TEM experiment.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Leiman, P.G.; Basler, M.; Ramagopal, U.A.; Bonanno, J.B.; Sauder, J.M.; Pukatzki, S.; Burley, S.K.; Almo, S.C.; Mekalanos, J.J. Type VI secretion apparatus and phage tail-associated protein complexes share a common evolutionary origin. Proc. Natl. Acad. Sci. USA 2009, 106, 4154–4159. [Google Scholar] [CrossRef] [PubMed]
- Ho, B.T.; Dong, T.G.; Mekalanos, J.J. A view to a kill: The bacterial type VI secretion system. Cell Host Microbe 2014, 15, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Basler, M.; Ho, B.T.; Mekalanos, J.J. Tit-for-tat: Type VI secretion system counterattack during bacterial cell-cell interactions. Cell 2013, 152, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Coulthurst, S.J. The Type VI secretion system—A widespread and versatile cell targeting system. Res. Microbiol. 2013, 164, 640–654. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhou, X.; Ma, Y.; Zhou, M.; Waldor, M.K.; Zhang, Y.; Wang, Q. Serine/threonine kinase PpkA coordinates the interplay between T6SS2 activation and quorum sensing in the marine pathogen Vibrio alginolyticus. Environ. Microbiol. 2018, 20, 903–919. [Google Scholar] [CrossRef]
- Sheng, L.; Lv, Y.; Liu, Q.; Wang, Q.; Zhang, Y. Connecting type VI secretion, quorum sensing, and c-di-GMP production in fish pathogen Vibrio alginolyticus through phosphatase PppA. Vet. Microbiol. 2013, 162, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Salomon, D.; Klimko, J.A.; Trudgian, D.C.; Kinch, L.N.; Grishin, N.V.; Mirzaei, H.; Orth, K. Type VI Secretion System Toxins Horizontally Shared between Marine Bacteria. PLoS Pathog. 2015, 11, e1005128. [Google Scholar] [CrossRef] [PubMed]
- Durand, E.; Nguyen, V.S.; Zoued, A.; Logger, L.; Péhau-Arnaudet, G.; Aschtgen, M.S.; Spinelli, S.; Desmyter, A.; Bardiaux, B.; Dujeancourt, A.; et al. Biogenesis and structure of a type VI secretion membrane core complex. Nature 2015, 523, 555–560. [Google Scholar] [CrossRef]
- Wang, J.; Brackmann, M.; Castaño-Díez, D.; Kudryashev, M.; Goldie, K.N.; Maier, T.; Stahlberg, H.; Basler, M. Cryo-EM structure of the extended type VI secretion system sheath-tube complex. Nat. Microbiol. 2017, 2, 1507–1512. [Google Scholar] [CrossRef]
- Howard, S.A.; Furniss, R.C.D.; Bonini, D.; Amin, H.; Paracuellos, P.; Zlotkin, D.; Costa, T.R.D.; Levy, A.; Mavridou, D.A.I.; Filloux, A. The Breadth and Molecular Basis of Hcp-Driven Type VI Secretion System Effector Delivery. MBio 2021, 12, e0026221. [Google Scholar] [CrossRef]
- Sana, T.G.; Flaugnatti, N.; Lugo, K.A.; Lam, L.H.; Jacobson, A.; Baylot, V.; Durand, E.; Journet, L.; Cascales, E.; Monack, D.M. Salmonella Typhimurium utilizes a T6SS-mediated antibacterial weapon to establish in the host gut. Proc. Natl. Acad. Sci. USA 2016, 113, E5044–E5051. [Google Scholar] [CrossRef]
- Wang, N.; Liu, J.; Pang, M.; Wu, Y.; Awan, F.; Liles, M.R.; Lu, C.; Liu, Y. Diverse roles of Hcp family proteins in the environmental fitness and pathogenicity of Aeromonas hydrophila Chinese epidemic strain NJ-35. Appl. Microbiol. Biotechnol. 2018, 102, 7083–7095. [Google Scholar] [CrossRef]
- Kalindamar, S.; Abdelhamed, H.; Kordon, A.O.; Pinchuk, L.M.; Karsi, A. Hemolysin Co-regulated Family Proteins Hcp1 and Hcp2 Contribute to Pathogenesis. Front. Vet. Sci. 2021, 8, 681609. [Google Scholar] [CrossRef] [PubMed]
- Guan, J.; Xiao, X.; Xu, S.; Gao, F.; Wang, J.; Wang, T.; Song, Y.; Pan, J.; Shen, X.; Wang, Y. Roles of RpoS in Yersinia pseudotuberculosis stress survival, motility, biofilm formation and type VI secretion system expression. J. Microbiol. 2015, 53, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Hao, S.; Lan, R.; Wang, G.; Xiao, D.; Sun, H.; Xu, J. The Type VI Secretion System Modulates Flagellar Gene Expression and Secretion in Citrobacter freundii and Contributes to Adhesion and Cytotoxicity to Host Cells. Infect. Immun. 2015, 83, 2596–2604. [Google Scholar] [CrossRef]
- Wang, P.; Dong, J.F.; Li, R.Q.; Li, L.; Zou, Q.H. Roles of the Hcp family proteins in the pathogenicity of Salmonella typhimurium 14,028 s. Virulence 2020, 11, 1716–1726. [Google Scholar] [CrossRef]
- Gong, J.S.; Guan, Y.C.; Zhao, Z.L.; Cai, Y.N.; Shan, X.F. Hcp1 regulates flagella of Aeromonas veronii TH0426 to reduce virulence. Aquaculture 2023, 576, 739899. [Google Scholar]
- Chevance, F.F.V.; Hughes, K.T. Coordinating assembly of a bacterial macromolecular machine. Nat. Rev. Microbiol. 2008, 6, 455–465. [Google Scholar] [CrossRef]
- Echazarreta, M.A.; Klose, K.E. Vibrio Flagellar Synthesis. Front. Cell. Infect. Microbiol. 2019, 9, 131. [Google Scholar] [CrossRef] [PubMed]
- Danson, A.E.; Jovanovic, M.; Buck, M.; Zhang, X. Mechanisms of σ54-Dependent Transcription Initiation and Regulation. J. Mol. Biol. 2019, 431, 3960–3974. [Google Scholar] [CrossRef]
- Yu, C.; Yang, F.; Xue, D.; Wang, X.; Chen, H. The Regulatory Functions of σ54 Factor in Phytopathogenic Bacteria. Int. J. Mol. Sci. 2021, 22, 12692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, S.; Ren, W.; Gong, X.; Long, H.; Zhang, X.; Cai, X.; Huang, A.; Xie, Z. Roles of rpoN in biofilm formation of Vibrio alginolyticus HN08155 at different cell densities. Microbiol. Res. 2021, 247, 126728. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Zhang, Y.; Wang, K.; Li, M.; Jiao, X. Characterization of the RpoN regulon reveals the regulation of motility, T6SS2 and metabolism in Vibrio parahaemolyticus. Front. Microbiol. 2022, 13, 1025960. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Guo, X.; Li, J.; Li, Y.; Sun, H.; Li, A.; Cao, B. RpoN is required for the motility and contributes to the killing ability of Plesiomonas shigelloides. BMC Microbiol. 2022, 22, 299. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Tabassum, N.; Anand, R.; Kim, Y.M. Motility of Vibrio spp.: Regulation and controlling strategies. Appl. Microbiol. Biotechnol. 2020, 104, 8187–8208. [Google Scholar] [CrossRef] [PubMed]
- Patten, C.L.; Kirchhof, M.G.; Schertzberg, M.R.; Morton, R.A.; Schellhorn, H.E. Microarray analysis of RpoS-mediated gene expression in Escherichia coli K-12. Mol. Genet. Genom. 2004, 272, 580–591. [Google Scholar] [CrossRef]
- Dong, T.; Yu, R.; Schellhorn, H. Antagonistic regulation of motility and transcriptome expression by RpoN and RpoS in Escherichia coli. Mol. Microbiol. 2011, 79, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Bi, W.; Chen, S.; Zhu, J.; Liu, X. Regulatory function of sigma factors RpoS/RpoN in adaptation and spoilage potential of Shewanella baltica. Food Microbiol. 2021, 97, 103755. [Google Scholar] [CrossRef]
- Cai, S.H.; Wu, Z.H.; Jian, J.C.; Lu, Y.S. Cloning and expression of the gene encoding an extracellular alkaline serine protease from Vibrio alginolyticus strain HY9901, the causative agent of vibriosis in Lutjanus erythopterus (Bloch). J. Fish. Dis. 2007, 30, 493–500. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Danio rerio); University of Oregon Press: Eugene, OR, USA, 2007. [Google Scholar]
- Luo, P.; He, X.; Liu, Q.; Hu, C. Developing Universal Genetic Tools for Rapid and Efficient Deletion Mutation in Vibrio Species Based on Suicide T-Vectors Carrying a Novel Counterselectable Marker, vmi480. PLoS ONE 2015, 10, e0144465. [Google Scholar] [CrossRef]
- Zhou, Z.; Pang, H.; Ding, Y.; Cai, J.; Huang, Y.; Jian, J.; Wu, Z. VscO, a putative T3SS chaperone escort of Vibrio alginolyticus, contributes to virulence in fish and is a target for vaccine development. Fish Shellfish Immunol. 2013, 35, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Moisi, M.; Jenul, C.; Butler, S.M.; New, A.; Tutz, S.; Reidl, J.; Klose, K.E.; Camilli, A.; Schild, S. A novel regulatory protein involved in motility of Vibrio cholerae. J. Bacteriol. 2009, 191, 7027–7038. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Xu, T.; Fossheim, L.E.; Zhang, X.H. FliC, a flagellin protein, is essential for the growth and virulence of fish pathogen Edwardsiella tarda. PLoS ONE 2012, 7, e45070. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.G.; Varcoe, L.T.; Attridge, S.R.; Manning, P.A. Vibrio cholerae Hcp, a secreted protein coregulated with HlyA. Infect. Immun. 1996, 64, 283–289. [Google Scholar] [CrossRef]
- Silverman, J.M.; Agnello, D.M.; Zheng, H.; Andrews, B.T.; Li, M.; Catalano, C.E.; Gonen, T.; Mougous, J.D. Haemolysin coregulated protein is an exported receptor and chaperone of type VI secretion substrates. Mol. Cell 2013, 51, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Bouteiller, M.; Dupont, C.; Bourigault, Y.; Latour, X.; Barbey, C.; Konto-Ghiorghi, Y.; Merieau, A. Flagella: Generalities and Specificities. Int. J. Mol. Sci. 2021, 22, 3337. [Google Scholar] [CrossRef] [PubMed]
- Kamber, T.; Pothier, J.F.; Pelludat, C.; Rezzonico, F.; Duffy, B.; Smits, T.H.M. Role of the type VI secretion systems during disease interactions of Erwinia amylovora with its plant host. BMC Genom. 2017, 18, 628. [Google Scholar] [CrossRef]
- Bouteiller, M.; Gallique, M.; Bourigault, Y.; Kosta, A.; Hardouin, J.; Massier, S.; Konto-Ghiorghi, Y.; Barbey, C.; Latour, X.; Chane, A.; et al. Crosstalk between the Type VI Secretion System and the Expression of Class IV Flagellar Genes in the MFE01 Strain. Microorganisms 2020, 8, 622. [Google Scholar] [CrossRef]
- Dong, T.G.; Mekalanos, J.J. Characterization of the RpoN regulon reveals differential regulation of T6SS and new flagellar operons in Vibrio cholerae O37 strain V52. Nucleic Acids Res. 2012, 40, 7766–7775. [Google Scholar] [CrossRef]
- Thormann, K.M.; Beta, C.; Kühn, M.J. Wrapped Up: The Motility of Polarly Flagellated Bacteria. Annu. Rev. Microbiol. 2022, 76, 349–367. [Google Scholar] [CrossRef]
- Liang, H.; Xia, L.; Wu, Z.; Jian, J.; Lu, Y. Expression, characterization and immunogenicity of flagellin FlaC from Vibrio alginolyticus strain HY9901. Fish Shellfish Immunol. 2010, 29, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.Y.; Wu, Z.H.; Jian, J.C.; Liu, Z.H. Construction of a fusion flagellin complex and evaluation of the protective immunity of it in red snapper (Lutjanus sanguineus). Lett. Appl. Microbiol. 2012, 55, 115–121. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Wu, F.; Huang, Y.C.; Jian, J.C.; Cai, S.H. Mechanisms Underlying the Virulence and Intestinal Colonization of the Vibrio alginolyticus HY9901 DctP Protein with Proteomic Analysis. Appl. Biochem. Microbiol. 2023, 59, 646–658. [Google Scholar] [CrossRef]
- Appelt, S.; Heuner, K. The Flagellar Regulon of Legionella-A Review. Front. Cell. Infect. Microbiol. 2017, 7, 454. [Google Scholar] [CrossRef]
- Liu, E.; Ye, J.; Song, S.; Wang, K.; Zhang, Y.; Zhang, H. Impact of co-deficiency of RpoN and RpoS on stress tolerance, virulence and gene regulation in Edwardsiella tarda. J. Basic Microbiol. 2014, 54, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Zhang, X.; Zhang, Y.; Zhu, M.; Yang, P.; Yuan, J.; Xie, Y.; Zhou, T.; Wang, W.; Chen, S.; et al. RpoN-Dependent Direct Regulation of Quorum Sensing and the Type VI Secretion System in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2018, 200, e00205–e00218. [Google Scholar] [CrossRef] [PubMed]
- Homma, M.; Kobayakawa, T.; Hao, Y.; Nishikino, T.; Kojima, S. Function and Structure of FlaK, a Master Regulator of the Polar Flagellar Genes in Marine Vibrio. J. Bacteriol. 2022, 204, e0032022. [Google Scholar] [CrossRef]
- Luo, G.; Huang, L.; Su, Y.; Qin, Y.; Xu, X.; Zhao, L.; Yan, Q. flrA, flrB and flrC regulate adhesion by controlling the expression of critical virulence genes in Vibrio alginolyticus. Emerg. Microbes Infect. 2016, 5, e85. [Google Scholar] [CrossRef]
- Wu, C.M.; Huang, H.H.; Li, L.H.; Lin, Y.T.; Yang, T.C. Molecular Characterization of Three Tandemly Located Flagellin Genes of Stenotrophomonas maltophilia. Int. J. Mol. Sci. 2022, 23, 3863. [Google Scholar] [CrossRef]
- Groisman, E.A. Feedback Control of Two-Component Regulatory Systems. Annu. Rev. Microbiol. 2016, 70, 103–124. [Google Scholar] [CrossRef]
- Manera, K.; Caro, F.; Li, H.; Pei, T.T.; Hersch, S.J.; Mekalanos, J.J.; Dong, T.G. Sensing of intracellular Hcp levels controls T6SS expression in Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2021, 118, e2104813118. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Establishment and physiological analysis of hcp2 mutation. (A) Validation of hcp2 mRNA level in hcp2 mutant strain and wild-type HY9901 by RT-PCR. (B) Growth curve of wild-type HY9901 and Δhcp2 strain. (C) Observation of swarming motility in wild-type HY9901 and Δhcp2 strain. (D) Measurement results of motility growth diameter in wild-type HY9901 and Δhcp2 strain. (E) The result using PCR verifying hcp2 complementary strain of V. alginolyticus. (F) The verification of hcp2 transcription level after recombinant plasmid transformed into Vibrio alginolyticus by RT-qPCR. (G) Validation of swarming motility in wild-type, hcp2 mutation, and complementary strains. M: DL2000 DNA Marker. HY9901-ev: the wild-type strain harboring empty vector pBBR1-MCS1; Δhcp2-ev: the Δhcp2 strain harboring empty vector pBBR1-MCS1; Δhcp2-com: the Δhcp2 strain harboring complementary vector pBBR1-MCS1-hcp2; hcp2-TF/TR: the genotyping result of genomic hcp2 using the hcp2-TF/TR primers; M13F/13R: the genotyping result of plasmid using the M13F/13R primers; Scale bar: 10 mm in (C,G); *** p < 0.001, ** p < 0.01.
Figure 1.
Establishment and physiological analysis of hcp2 mutation. (A) Validation of hcp2 mRNA level in hcp2 mutant strain and wild-type HY9901 by RT-PCR. (B) Growth curve of wild-type HY9901 and Δhcp2 strain. (C) Observation of swarming motility in wild-type HY9901 and Δhcp2 strain. (D) Measurement results of motility growth diameter in wild-type HY9901 and Δhcp2 strain. (E) The result using PCR verifying hcp2 complementary strain of V. alginolyticus. (F) The verification of hcp2 transcription level after recombinant plasmid transformed into Vibrio alginolyticus by RT-qPCR. (G) Validation of swarming motility in wild-type, hcp2 mutation, and complementary strains. M: DL2000 DNA Marker. HY9901-ev: the wild-type strain harboring empty vector pBBR1-MCS1; Δhcp2-ev: the Δhcp2 strain harboring empty vector pBBR1-MCS1; Δhcp2-com: the Δhcp2 strain harboring complementary vector pBBR1-MCS1-hcp2; hcp2-TF/TR: the genotyping result of genomic hcp2 using the hcp2-TF/TR primers; M13F/13R: the genotyping result of plasmid using the M13F/13R primers; Scale bar: 10 mm in (C,G); *** p < 0.001, ** p < 0.01.

Figure 2.
Comparison of virulence on zebrafish between wild-type HY9901 and hcp2 mutation. (A) Zebrafish phenotype after Vibrio alginolyticus immersion for 43 hpi at 4 dpf larvae. (B) Mortality analysis of AB zebrafish larvae after 69 h treatment in uninfected, wild-type HY9901, and Δhcp2 strain groups, respectively. (C) Adult zebrafish phenotype after intraperitoneal injection by wild-type HY9901 or Δhcp2 strain. (D) Comparison of Vibrio alginolyticus LD50 against adult zebrafish; red arrow: gut edema; red asterisk: belly edema. CFU: colony forming unit; dpf: days post-fertilization; hpi: hours post-immersion. The bottom right value in Figure 2A represents the phenotype ratio; n: the number of zebrafish analyzed; *** p < 0.001, ** p < 0.01, * p < 0.05; scale bars: 500 μm in Figure 2A.
Figure 2.
Comparison of virulence on zebrafish between wild-type HY9901 and hcp2 mutation. (A) Zebrafish phenotype after Vibrio alginolyticus immersion for 43 hpi at 4 dpf larvae. (B) Mortality analysis of AB zebrafish larvae after 69 h treatment in uninfected, wild-type HY9901, and Δhcp2 strain groups, respectively. (C) Adult zebrafish phenotype after intraperitoneal injection by wild-type HY9901 or Δhcp2 strain. (D) Comparison of Vibrio alginolyticus LD50 against adult zebrafish; red arrow: gut edema; red asterisk: belly edema. CFU: colony forming unit; dpf: days post-fertilization; hpi: hours post-immersion. The bottom right value in Figure 2A represents the phenotype ratio; n: the number of zebrafish analyzed; *** p < 0.001, ** p < 0.01, * p < 0.05; scale bars: 500 μm in Figure 2A.
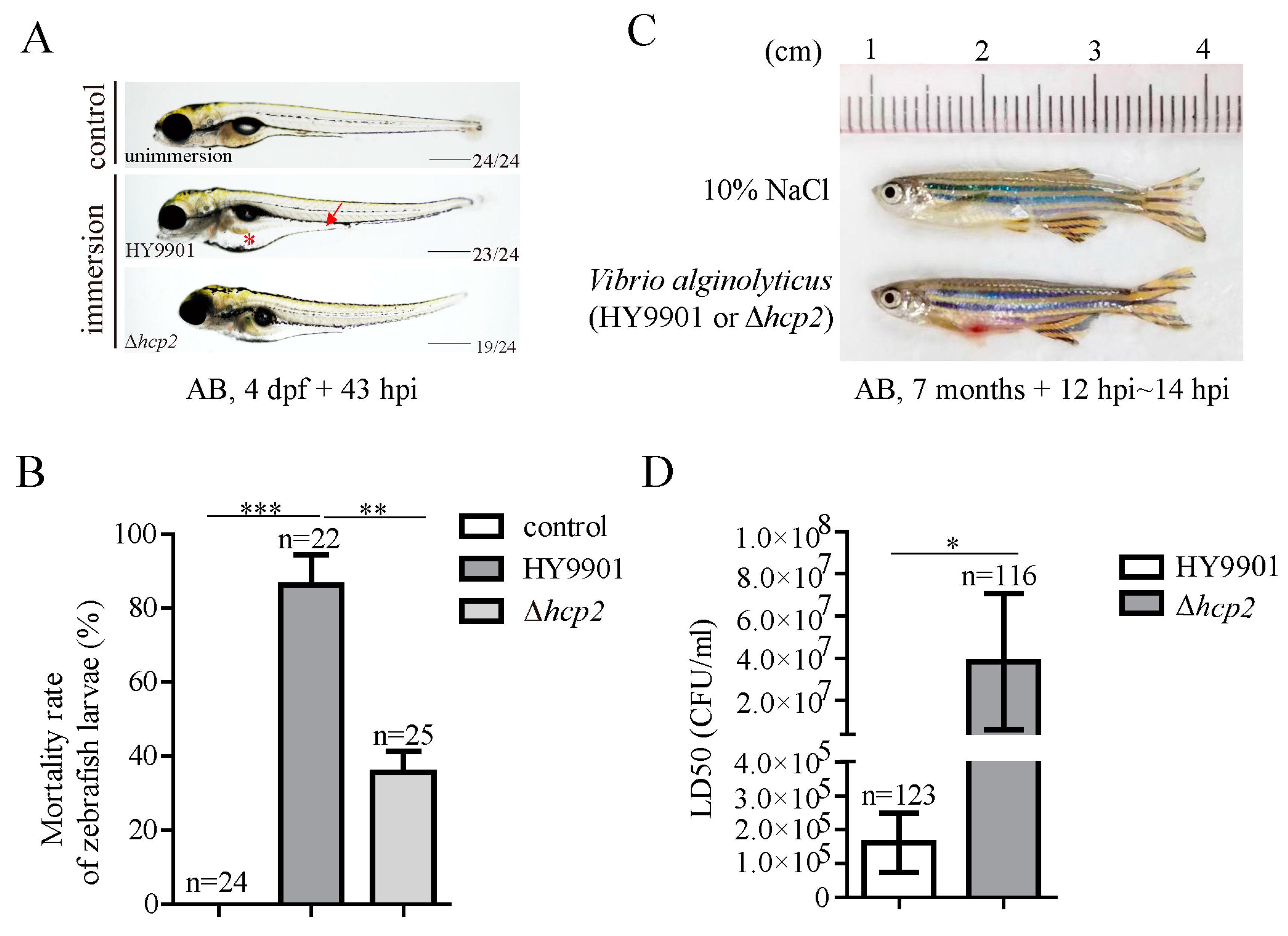
Figure 3.
Adhesive and intestinal colonization capacity of Vibrio alginolyticus against zebrafish. (A–D) Comparison of adhesion between wild-type HY9901 and hcp2 mutant strains on zebrafish larval surface at 5 dpf after 42 hpi. (E–I’) Comparison of colonization ability between wild-type HY9901 and hcp2 mutant strains on zebrafish intestinal canal at 5 dpf after 3 dpi and 5 dpi, respectively. Red font: DiI dye-labelled Vibrio alginolyticus; green font: green fluorescent protein-labelled zebrafish macrophage; white rectangle: zebrafish intestine colonized by Vibrio alginolyticus; white dotted rectangle: zebrafish caudal fin; white arrow: Vibrio alginolyticus outside of zebrafish anus; CFU: colony forming unit; n/n: number of fish with phenotype/number of total imaged fish; dpf: days post-fertilization; hpi or dpi: hours (days) post-immersion; scale bar: 500 μm in (D), 100 μm in (G,I’).
Figure 3.
Adhesive and intestinal colonization capacity of Vibrio alginolyticus against zebrafish. (A–D) Comparison of adhesion between wild-type HY9901 and hcp2 mutant strains on zebrafish larval surface at 5 dpf after 42 hpi. (E–I’) Comparison of colonization ability between wild-type HY9901 and hcp2 mutant strains on zebrafish intestinal canal at 5 dpf after 3 dpi and 5 dpi, respectively. Red font: DiI dye-labelled Vibrio alginolyticus; green font: green fluorescent protein-labelled zebrafish macrophage; white rectangle: zebrafish intestine colonized by Vibrio alginolyticus; white dotted rectangle: zebrafish caudal fin; white arrow: Vibrio alginolyticus outside of zebrafish anus; CFU: colony forming unit; n/n: number of fish with phenotype/number of total imaged fish; dpf: days post-fertilization; hpi or dpi: hours (days) post-immersion; scale bar: 500 μm in (D), 100 μm in (G,I’).

Figure 4.
Imaging of the flagellar filament morphology by electron microscopy. (A–C) Negative-staining EM images of the sheathed polar flagella of wild-type HY9901, Δhcp2 strain, and Δhcp2 complementary strain; (i–iii) are high-magnification scans of the red line-marked regions in each top-left image, respectively. (D) Measurement of outer-diameter of flagellar filaments in wild-type HY9901, Δhcp2 strain, and Δhcp2-complementary strain. HY9901-ev: the wild-type strain harboring empty vector pBBR1-MCS1; Δhcp2-ev: the Δhcp2 strain harboring empty vector pBBR1-MCS1; Δhcp2-com: the Δhcp2 strain harboring complementary vector pBBR1-MCS1-hcp2; n: the number of analyzed bacteria at 30,000× magnification. *** p < 0.001, ** p < 0.01; ns: no significance.
Figure 4.
Imaging of the flagellar filament morphology by electron microscopy. (A–C) Negative-staining EM images of the sheathed polar flagella of wild-type HY9901, Δhcp2 strain, and Δhcp2 complementary strain; (i–iii) are high-magnification scans of the red line-marked regions in each top-left image, respectively. (D) Measurement of outer-diameter of flagellar filaments in wild-type HY9901, Δhcp2 strain, and Δhcp2-complementary strain. HY9901-ev: the wild-type strain harboring empty vector pBBR1-MCS1; Δhcp2-ev: the Δhcp2 strain harboring empty vector pBBR1-MCS1; Δhcp2-com: the Δhcp2 strain harboring complementary vector pBBR1-MCS1-hcp2; n: the number of analyzed bacteria at 30,000× magnification. *** p < 0.001, ** p < 0.01; ns: no significance.
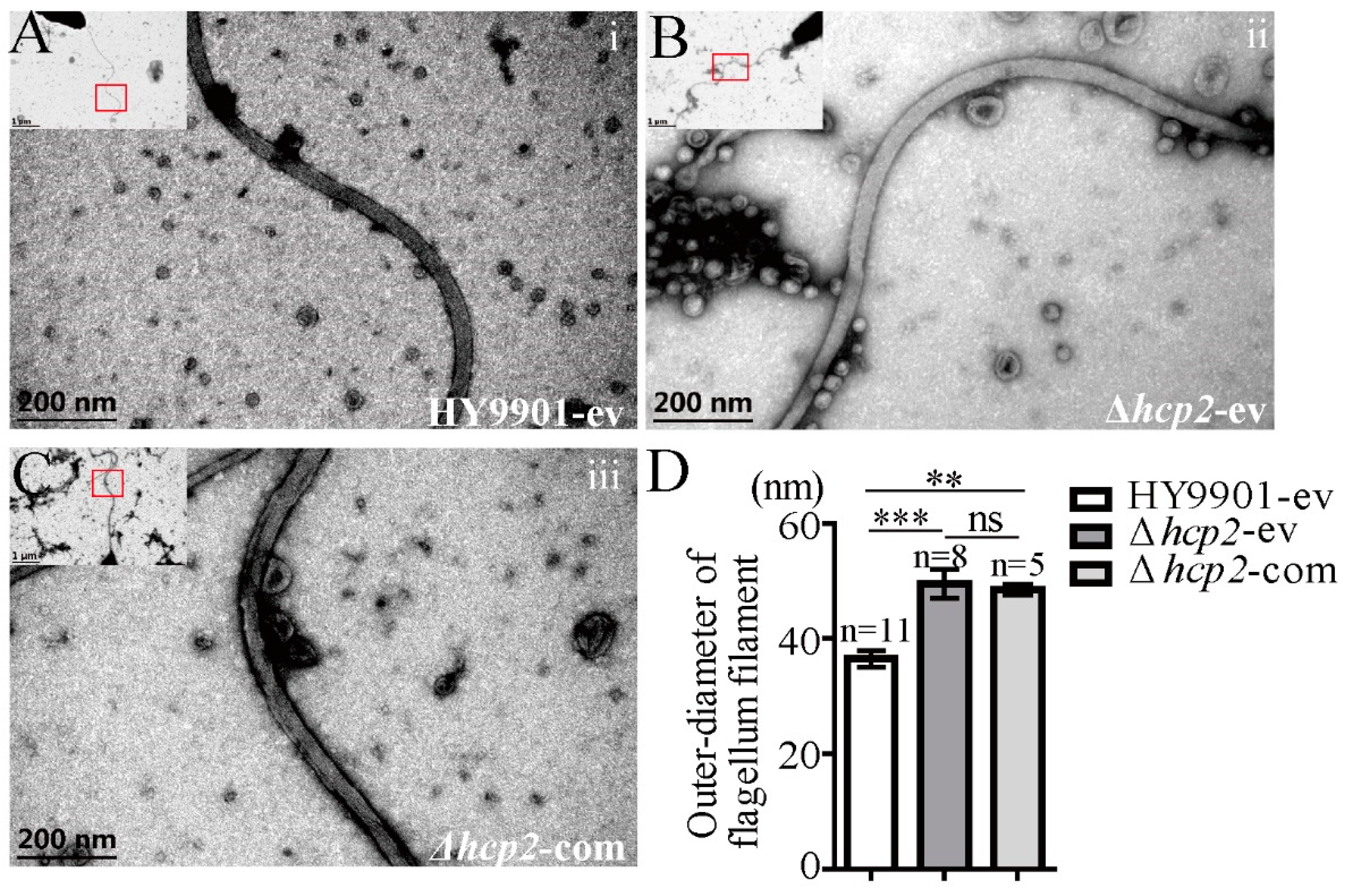
Figure 5.
Flagellin and flagellum export apparatus-associated genes’ expression of V. alginolyticus by quantitative proteomics and RT-qPCR analysis. (A) The enrichment result of bacterial motility and flagellum assembly-associated biological process (red rectangle) from all differential expression proteins in Δhcp2 strain compared with wild-type HY9901. (B) Volcano graphs for the analysis of the polar flagellum assembly-associated proteins based on proteomics data and differential expression proteins between Δhcp2 strain and wild-type HY9901. (C) Protein quantitative result of flagellins FlaA, FlaB, and FlaC in the wild-type HY9901 and the Δhcp2 strain by LFQ intensity. (D) Protein quantitative result of FliH, FliF, and FlgE in the wild-type HY9901 and the Δhcp2 strain by LFQ intensity. (E) The influence on transcriptional levels of flagellin genes flaA, flaB, and flaC by hcp2 using RT-qPCR analysis. (F) The influence on transcriptional levels of fliH, fliF, and flgE by hcp2 using RT-qPCR analysis. HY9901-ev or Δhcp2-ev: the wild-type strain or hcp2 mutant strain harboring empty vector; Δhcp2-com: the mutant strain harboring hcp2 complementary plasmid. *** p < 0.001, ** p < 0.01, * p < 0.05; ns: no significance.
Figure 5.
Flagellin and flagellum export apparatus-associated genes’ expression of V. alginolyticus by quantitative proteomics and RT-qPCR analysis. (A) The enrichment result of bacterial motility and flagellum assembly-associated biological process (red rectangle) from all differential expression proteins in Δhcp2 strain compared with wild-type HY9901. (B) Volcano graphs for the analysis of the polar flagellum assembly-associated proteins based on proteomics data and differential expression proteins between Δhcp2 strain and wild-type HY9901. (C) Protein quantitative result of flagellins FlaA, FlaB, and FlaC in the wild-type HY9901 and the Δhcp2 strain by LFQ intensity. (D) Protein quantitative result of FliH, FliF, and FlgE in the wild-type HY9901 and the Δhcp2 strain by LFQ intensity. (E) The influence on transcriptional levels of flagellin genes flaA, flaB, and flaC by hcp2 using RT-qPCR analysis. (F) The influence on transcriptional levels of fliH, fliF, and flgE by hcp2 using RT-qPCR analysis. HY9901-ev or Δhcp2-ev: the wild-type strain or hcp2 mutant strain harboring empty vector; Δhcp2-com: the mutant strain harboring hcp2 complementary plasmid. *** p < 0.001, ** p < 0.01, * p < 0.05; ns: no significance.
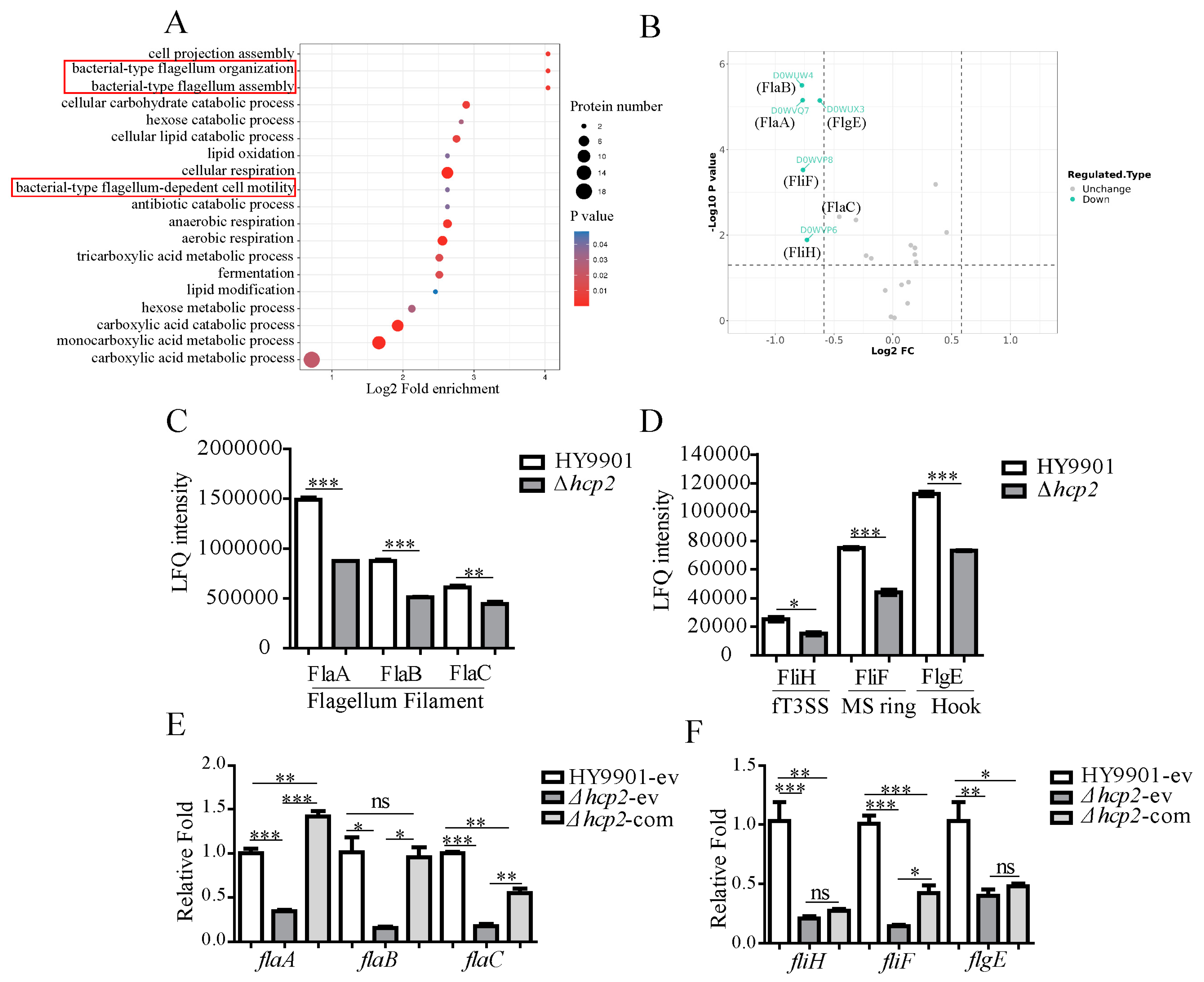
Figure 6.
The modulatory role of V. alginolyticus Hcp2 protein on the flagellar system and pathogenicity. (A) Transcription levels analysis of sigma factors rpoN, rpoS, and fliA in the wild-type, hcp2 mutation, and complementary strain by RT-qPCR. (B) Transcription level analysis of master regulon flrA and its downstream genes flrB and flrC in the wild-type, hcp2 mutation, and complementary strain by RT-qPCR. (C) General view of the roles of Hcp2 protein on flagellar system and pathogenicity of V. alginolyticus. Black dotted arrow: positive impact on flagellar assembly through Hcp2 protein; *** p < 0.001, ** p < 0.01, * p < 0.05; ns: no significance.
Figure 6.
The modulatory role of V. alginolyticus Hcp2 protein on the flagellar system and pathogenicity. (A) Transcription levels analysis of sigma factors rpoN, rpoS, and fliA in the wild-type, hcp2 mutation, and complementary strain by RT-qPCR. (B) Transcription level analysis of master regulon flrA and its downstream genes flrB and flrC in the wild-type, hcp2 mutation, and complementary strain by RT-qPCR. (C) General view of the roles of Hcp2 protein on flagellar system and pathogenicity of V. alginolyticus. Black dotted arrow: positive impact on flagellar assembly through Hcp2 protein; *** p < 0.001, ** p < 0.01, * p < 0.05; ns: no significance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, S.; Tang, J.; Wang, B.; Cai, J.; Jian, J. Roles of Hcp2, a Hallmark of T6SS2 in Motility, Adhesive Capacity, and Pathogenicity of Vibrio alginolyticus. Microorganisms 2023, 11, 2893. https://doi.org/10.3390/microorganisms11122893
AMA Style
Wu S, Tang J, Wang B, Cai J, Jian J. Roles of Hcp2, a Hallmark of T6SS2 in Motility, Adhesive Capacity, and Pathogenicity of Vibrio alginolyticus. Microorganisms. 2023; 11(12):2893. https://doi.org/10.3390/microorganisms11122893
Chicago/Turabian StyleWu, Shuilong, Jufen Tang, Bei Wang, Jia Cai, and Jichang Jian. 2023. "Roles of Hcp2, a Hallmark of T6SS2 in Motility, Adhesive Capacity, and Pathogenicity of Vibrio alginolyticus" Microorganisms 11, no. 12: 2893. https://doi.org/10.3390/microorganisms11122893
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.