

Investigating Bio-Inspired Degradation of Toxic Dyes Using Potential Multi-Enzyme Producing Extremophiles



Abstract
:1. Introduction
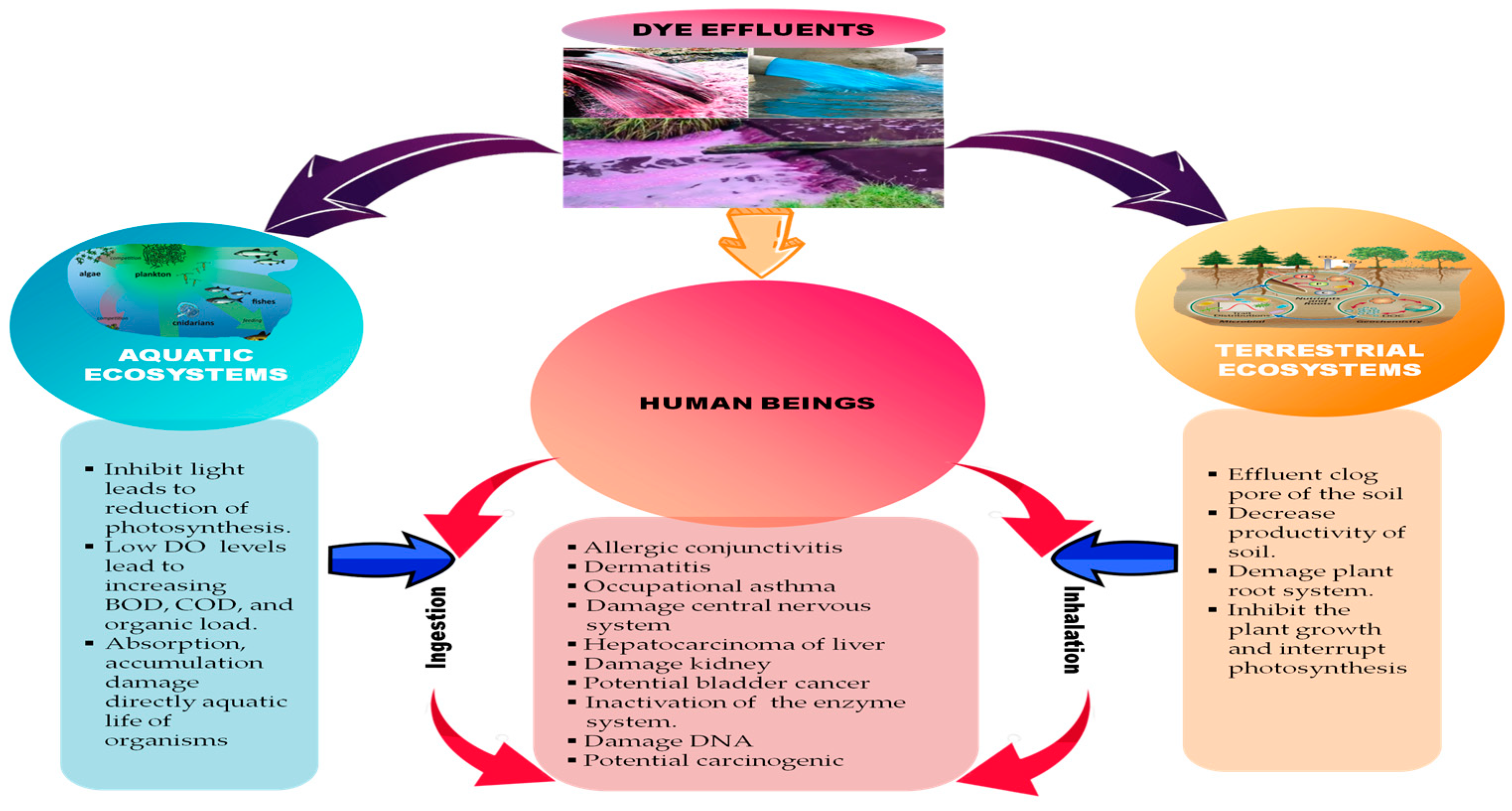
2. Toxicity and the Negative Impact of Dyes on Human and Environmental Ecosystems
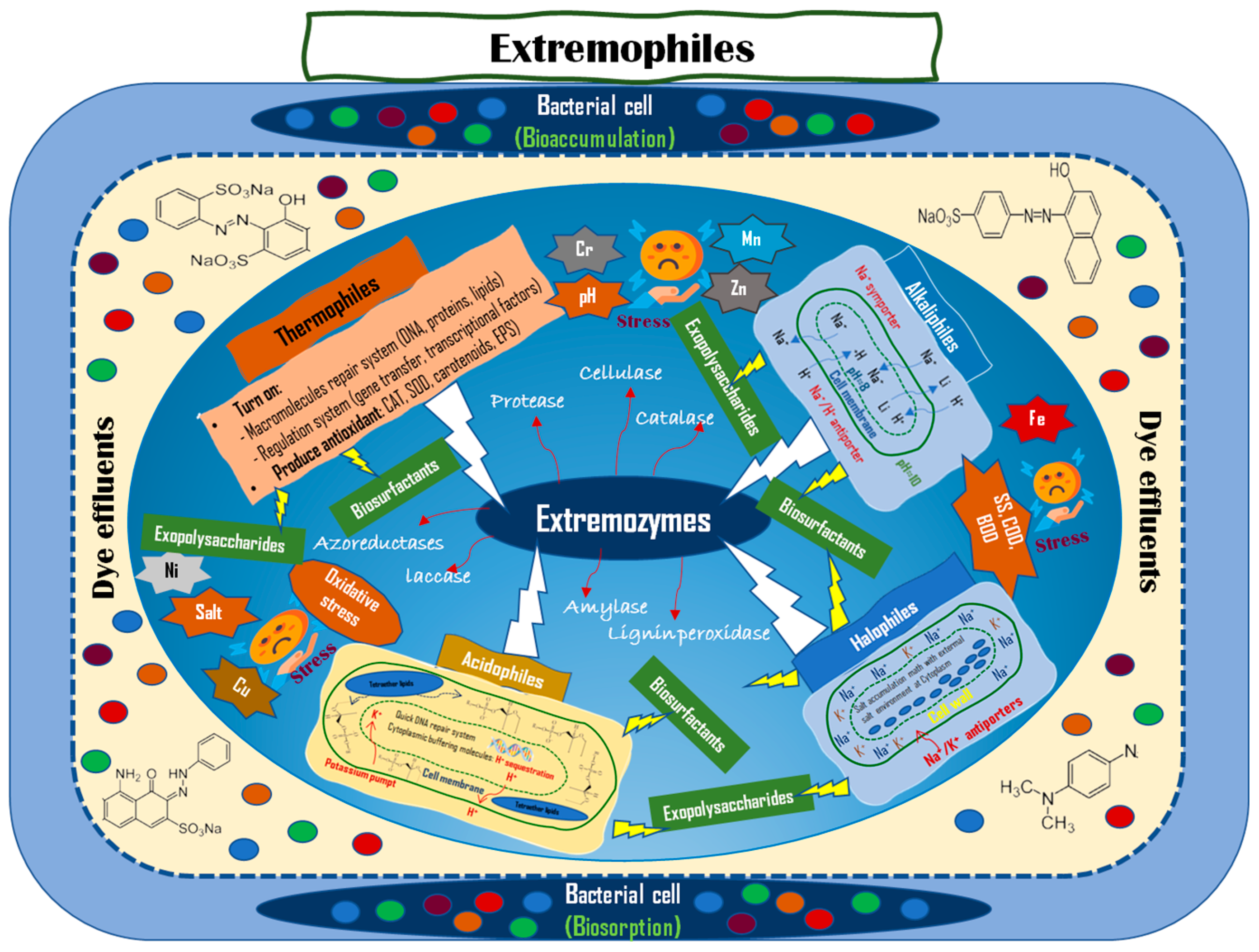
3. The Role of Polyextremophilic Bacteria in Dye Degradation
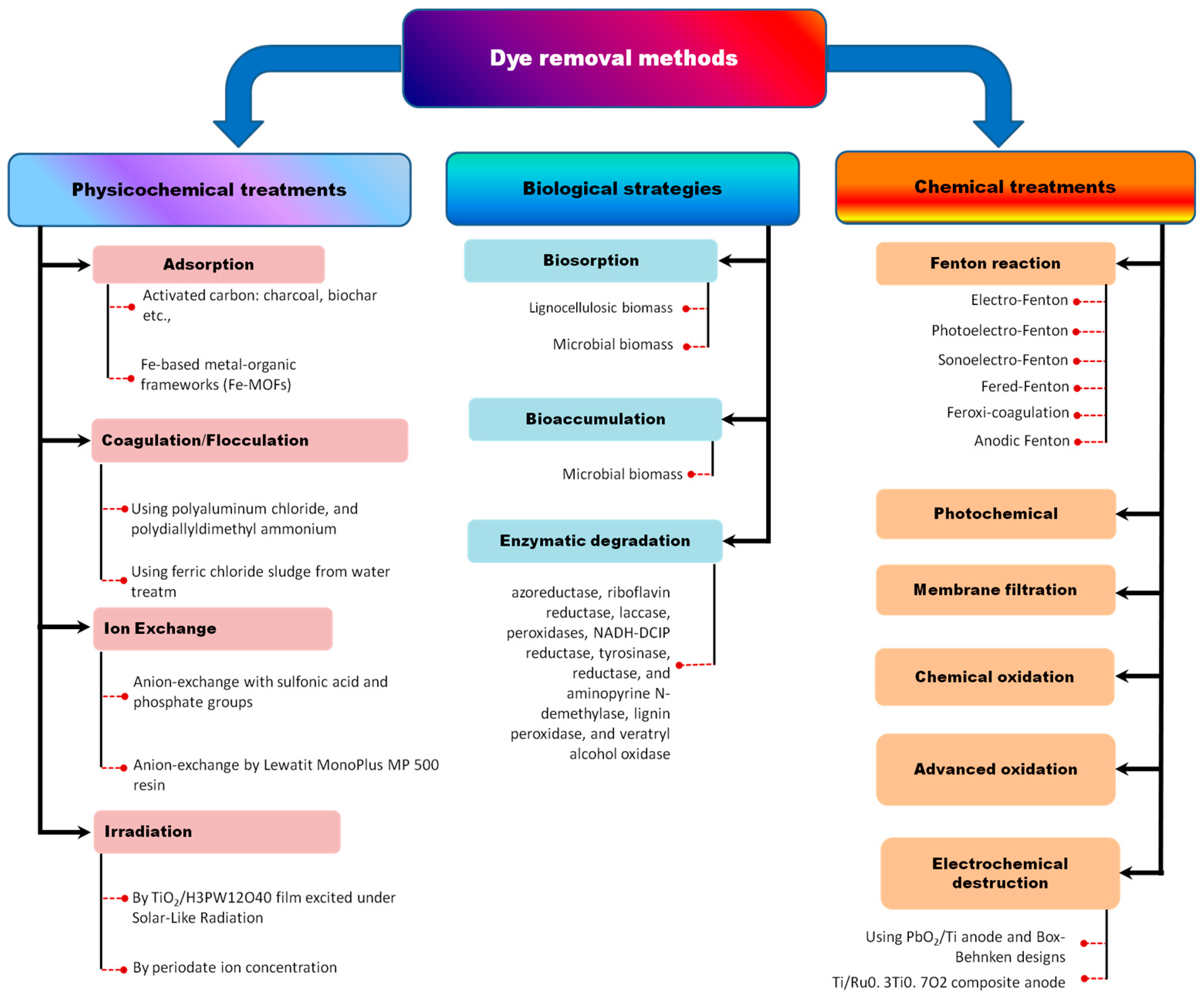
4. Enzyme-Linked Bioremediation of Dyes
5. Discussion and Future Prospective
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhatia, S.C. Pollution Control in Textile Industry; WPI Publishing: New York, NY, USA, 2017. [Google Scholar]
- Patila, S.M.; Suryavanshib, M.V.; Chandanshivec, V.V.; Kuradea, M.B.; Govindwara, S.P.; Jeona, B.H. Regeneration of textile wastewater deteriorated microbial diversity of soil microcosm through bioaugmentation. J. Chem. Eng. 2020, 380, 122533. [Google Scholar] [CrossRef]
- Hassan, M.M.; Carr, C.M. A critical review on recent advancements of the removal of reactive dyes from dyehouse effluent by ion-exchange adsorbents. Chemosphere 2018, 209, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Patel, V.R.; Khan, R.; Bhatt, N. Cost-effective in-situ remediation technologies for complete mineralization of dyes contaminated soils. Chemosphere 2020, 243, 125253. [Google Scholar] [CrossRef]
- Singh, R.L.; Singh, P.K.; Singh, R.P. Enzymatic decolorization and degradation of azo dyes—A review. Int. Biodeterior. Biodegrad. 2015, 104, 21–31. [Google Scholar] [CrossRef]
- Vikrant, K.; Giri, B.S.; Raza, N.; Roy, K.; Kim, K.-H.; Rai, B.N.; Singh, R.S. Recent advancements in bioremediation of dye: Current status and challenges. Bioresour. Technol. 2018, 253, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Banerjee, A.; Halder, U.; Biswas, R.; Bandopadhyay, R. Degradation of Synthetic Azo Dyes of Textile Industry: A Sustainable Approach Using Microbial Enzymes. Water. Conserv. Sci. Eng. 2017, 2, 121–131. [Google Scholar] [CrossRef]
- Meerbergen, K.; Willems, K.A.; Dewil, R.; Van Impe, J.; Appels, L.; Lievens, B. Isolation and screening of bacterial isolates from wastewater treatment plants to decolorize azo dyes. J. Biosci. Bioeng. 2018, 125, 448–456. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Biodegradation of Methylene Blue using a novel Lignin peroxidase enzyme producing bacteria, named Bacillus sp. React3, as a promising candidate for dye-contaminated wastewater treatment. Fermentation 2022, 8, 190. [Google Scholar] [CrossRef]
- Shaul, G.M.; Holdsworth, T.J.; Dempsey, C.R.; Dostal, K.A. Fate of water soluble azo dyes in the activated sludge process. Chemosphere 1991, 22, 107–119. [Google Scholar] [CrossRef]
- Le Borgne, S.; Paniagua, D.; Vazquez-Duhalt, R. Biodegradation of organic pollutants by halophilic bacteria and archaea. J. Mol. Microbiol. Biotechnol. 2008, 15, 74–92. [Google Scholar] [CrossRef]
- Ning, X.-A.; Lin, M.-Q.; Shen, L.-Z.; Zhang, J.-H.; Wang, J.-Y.; Wang, Y.-J.; Yang, Z.-Y.; Liu, J.-Y. Levels, composition profiles and risk assessment of polycyclic aromatic hydrocarbons (PAHs) in sludge from ten textile dyeing plants. Environ. Res. 2014, 132, 112–118. [Google Scholar] [CrossRef]
- Ning, X.-A.; Liang, J.-Y.; Li, R.-J.; Hong, Z.; Wang, Y.-J.; Chang, K.-L.; Zhang, Y.-P.; Yang, Z.-Y. Aromatic amine contents, component distributions and risk assessment in sludge from 10 textile-dyeing plants. Chemosphere 2015, 134, 367–373. [Google Scholar] [CrossRef]
- Kumar, M.; Leon, V.; De Sisto Materano, A.; Ilzins, O.A. A halotolerant and thermotolerant bacillus sp. degrades hydrocarbons and produces tensio-active emulsifying agent. World J. Microbiol. Biotech. 2007, 23, 211–220. [Google Scholar] [CrossRef]
- Gahlout, M.; Gupte, S.; Gupte, A. Optimization of culture condition for enhanced decolorization and degradation of azo dye reactive violet 1 with concomitant production of ligninolytic enzymes by Ganoderma cupreum AG-1.3. Biotechnology 2013, 3, 143–152. [Google Scholar] [CrossRef]
- Ergun, S.O.; Urek, R.O. Production of ligninolytic enzymes by solid state fermentation using Pleurotus ostreatus. Ann. Agrar. Sci. 2017, 15, 273–277. [Google Scholar] [CrossRef]
- Khaled, J.M.; Alyahya, S.A.; Govindan, R.; Chelliah, C.K.; Maruthupandy, M.; Alharbi, N.S.; Kadaikunnan, S.; Issac, R.; Murugan, S.; Li, W.J. Laccase producing bacteria influenced the high decolorization of textile azo dyes with advanced study. Environ. Res. 2022, 207, 112211. [Google Scholar] [CrossRef]
- Jojy, J.; Ramadoss, D.; Raman, H.K.; Peter, D.M.; Dharani, G.; Amit, K. Bio-decolorization of synthetic dyes by a halophilic bacterium Salinivibrio sp. Front. Microbiol. 2020, 11, 594011. [Google Scholar]
- Amoozegar, M.A.; Hajighasemi, M.; Hamedi, J.; Asad, S.; Ventosa, A. Azo dye decolorization by halophilic and halotolerant microorganisms. Ann. Microbiol. 2011, 61, 217–230. [Google Scholar] [CrossRef]
- Tian, F.; Wang, Y.; Guo, G.; Ding, K.; Yang, F.; Wang, H.; Cao, Y.; Liu, C. Enhanced azo dye biodegradation at high salinity by a halophilic bacterial consortium. Bioresour. Technol. 2021, 326, 124749. [Google Scholar] [CrossRef] [PubMed]
- John, J.; Siva, V.; Kumari, R.; Arya, A.; Kumar, A. Unveiling cultivable and uncultivable halophilic bacteria inhabiting Marakkanam saltpan, India and their potential for biotechnological applications. Geomicrobiology 2020, 37, 691–701. [Google Scholar] [CrossRef]
- Nzila, A. Current status of the degradation of aliphatic and aromatic petroleum hydrocarbons by Thermophilic microbes and Future Perspectives. Int. J. Environ. Res. Public Health 2018, 15, 2782. [Google Scholar] [CrossRef] [PubMed]
- Baker, B.A.; Tawabini, B.; Nazal, M.; AlThaqfi, J.; Khalil, A. Efficiency of thermophilic bacteria in wastewater treatment. Arab. J. Sci. Eng. 2021, 46, 123–128. [Google Scholar] [CrossRef]
- Dutta, S.; Bhattacharjee, J. A comparative study between physicochemical and biological methods for effective removal of textile dye from wastewater. In Development in Wastewater Treatment Research and Processes; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1–21. [Google Scholar]
- Parmar, S.; Daki, S.; Bhattacharya, S.; Shrivastav, A. Microorganism: An ecofriendly tool for waste management and environmental safety. In Development in Wastewater Treatment Research and Processes; Elsevier: Amsterdam, The Netherlands, 2022; pp. 175–193. [Google Scholar]
- Ao, S.; Zayed, T. Impact of sewer overflow on public health: A comprehensive scientometric analysis and systematic review. Environ. Res. 2022, 203, 111609. [Google Scholar]
- Gallo, G.; Puopolo, R.; Carbonaro, M.; Maresca, E.; Fiorentino, G. Extremophiles, a nifty tool to face environmental pollution: From exploitation of metabolism to genome engineering. Int. J. Environ. Res. Public Health 2021, 18, 5228. [Google Scholar] [CrossRef] [PubMed]
- Patil, R.; Zahid, M.; Govindwar, S.; Khandare, R.; Vyavahare, G.; Gurav, R.; Desai, N.; Pandit, S.; Jadhav, J. Constructed wetland: A promising technology for the treatment of hazardous textile dyes and effluent. In Development in Wastewater Treatment Research and Processes; Elsevier: Amsterdam, The Netherlands, 2022; pp. 173–198. [Google Scholar]
- Elgarahy, A.M.; Elwakeel, K.Z.; Mohammad, S.H.; Elshoubaky, G.A. A critical review of biosorption of dyes, heavy metals and metalloids from wastewater as an efficient and green process. Clean. Eng. Technol. 2021, 4, 100209. [Google Scholar] [CrossRef]
- Rawat, D.; Sharma, R.S.; Karmakar, S.; Arora, L.S.; Mishra, V. Ecotoxic potential of a presumably non-toxic azo dye. Ecotoxicol. Environ. Saf. 2018, 148, 528–537. [Google Scholar] [CrossRef]
- Khan, S.; Malik, A. Toxicity evaluation of textile effluents and role of native soil bacterium in biodegradation of a textile dye. Environ. Sci. Pollut. Res. 2018, 25, 4446–4458. [Google Scholar] [CrossRef]
- Copaciu, F.; Opris¸, O.; Coman, V.; Ristoiu, D.; Niinemets, Ü.; Copolovici, L. Diffuse water pollution by anthraquinone and azo dyes in environment importantly alters foliage volatiles, carotenoids and physiology in wheat (Triticum aestivum). Water. Air. Soil. Pollut. 2013, 224, 1478. [Google Scholar] [CrossRef]
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nature. 2001, 409, 1092–1101. [Google Scholar] [CrossRef]
- Ajaz, M.; Rehman, A.; Khan, Z.; Nisar, M.A.; Hussain, S. Degradation of azo dyes by Alcaligenes aquatilis 3c and its potential use in the wastewater treatment. AMB Express 2019, 9, 64. [Google Scholar] [CrossRef]
- Sridharan, R.; Krishnaswamy, V.; Archana, K.M.; Rajagopal, R.; Kumar Thirumal, D.; Priya Doss George, C. Integrated approach on azo dyes degradation using laccase enzyme and Cul nanoparticle. SN Appl. Sci. 2021, 3, 370. [Google Scholar] [CrossRef]
- Aragaw, T.A.; Bogale, F.M.; Gessesse, A. Adaptive Response of thermophiles to redox stress and their role in the process of dye degradation from textile industry wastewater. Front. Physiol. 2022, 13, 1220. [Google Scholar] [CrossRef]
- Bedekar, P.A.; Saratale, R.G.; Saratale, G.D.; Govindwar, S.P. Oxidative stress response in dye degrading bacterium Lysinibacillus sp. RGS exposed to Reactive Orange 16, degradation of RO16 and evaluation of toxicity. Environ. Sci. Pollut. Res. 2014, 21, 11075–11085. [Google Scholar] [CrossRef]
- Tapadar, S.; Tripathi, D.; Pandey, S.; Goswami, K.; Bhattacharjee, A.; Das, K.; Palwan, E.; Rani, M.; Kumar, A. Role of extremophiles and extremophilic proteins in industrial waste treatment. In Removal of Emerging Contaminants through Microbial Processes; Shah, M.P., Ed.; Springer: Singapore, 2021; pp. 217–235. [Google Scholar]
- Russell, N.J. Membrane components and cold sensing. In Psychrophiles: From Biodiversity to Biotechnology; Springer: Berlin/Heidelberg, Germany, 2008; pp. 177–190. [Google Scholar]
- Bar Dolev, M.; Braslavsky, I.; Davies, P.L. Ice-binding proteins and their function. Annu. Rev. Biochem. 2016, 85, 515–542. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.; Bag, S.K.; Das, S.; Harvill, E.T.; Dutta, C. Molecular signature of hypersaline adaptation: Insights from genome and proteome composition of halophilic prokaryotes. Genome Biol. 2008, 9, R70. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Dopson, M. Life in acid: pH homeostasis in acidophiles. Trends. Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef]
- Sarma, J.; Sengupta, A.; Laskar, M.K.; Sengupta, S.; Tenguria, S.; Kumar, A. Chapter 13—Microbial adaptations in extreme environmental conditions. In Developments in Applied Microbiology and Biotechnology. Bacterial Survival in the Hostile Environment; Kumar, A., Tenguria, S., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 193–206. [Google Scholar]
- Horikoshi, K. Alkaliphiles. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2004, 80, 166–178. [Google Scholar] [CrossRef]
- Maksimainen, M.; Paavilainen, S.; Hakulinen, N.; Rouvinen, J. Structural analysis, enzymatic characterization, and catalytic mechanisms of β-galactosidase from Bacillus circulans sp. alkalophilus. FEBS J. 2012, 279, 1788–1798. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.S.; Nair, A.; Nivetha, K.; More, V.S.; Anantharaju, K.S.; More, S.S. Chapter 7—Molecular adaptations in proteins and enzymes produced by extremophilic microorganisms. In Extremozymes and Their Industrial Applications; Arora, N.K., Agnihotri, S., Mishra, J., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 205–230. [Google Scholar]
- Srivastava, A.; Dangi, L.K.; Kumar, S.; Rani, R. Microbial decolorization of Reactive Black 5 dye by Bacillus albus DD1 isolated from textile water effluent: Kinetic, thermodynamics & decolorization mechanism. Heliyon 2022, 8, e08834. [Google Scholar]
- Guadie, A.; Tizazu, S.; Melese, M.; Guo, W.; Ngo, H.H.; Xia, S. Biodecolorization of textile azo dye using Bacillus sp. strain CH12 isolated from alkaline lake. Biotechnol. Rep. 2017, 15, 92–100. [Google Scholar] [CrossRef]
- Unnikrishnan, S.; Khan, M.H.; Ramalingam, K. Dye-tolerant marine Acinetobacter baumannii-mediated biodegradation of reactive red. Water Sci. Eng. 2018, 11, 265–275. [Google Scholar] [CrossRef]
- Dos Santos, A.B.; Traverse, J.; Cervantes, F.J.; Van Lier, B. Enhancing the electron transfer capacity and subsequent color removal in bioreactors by applying thermophilic anaerobic treatment and redox mediators. Biotechnol. Bioengineer. 2005, 89, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Giovanella, P.; Vieira, G.A.L.; Ramos Otero, I.V.; Pais Pellizzer, E.; de Jesus Fontes, B.; Sette, L.D. Metal and organic pollutants bioremediation by extremophile microorganisms. J. Hazard. Mater. 2020, 382, 121024. [Google Scholar] [CrossRef] [PubMed]
- Aydin, M.; Ağaoğlu, A.; Baris, Ö. Azo dye decolorization by using four different psychrotolerant bacillus species. NE. Sci. 2021, 6, 19–29. [Google Scholar] [CrossRef]
- Khalid, A.; Kausar, F.; Arshad, M.; Mahmood, T.; Ahmed, I. Accelerated decolorization of reactive azo dyes under saline conditions by bacteria isolated from Arabian seawater sediment. Appl. Microbiol. Biotechnol. 2012, 96, 1599–1606. [Google Scholar] [CrossRef]
- Mishra, S.; Nayak, J.K.; Maiti, A. Bacteria-mediated bio-degradation of reactive azo dyes coupled with bio-energy generation from model wastewater. Clean. Technol. Environ. Policy 2020, 22, 651–667. [Google Scholar] [CrossRef]
- Takkar, S.; Tyagi, B.; Kumar, N.; Kumari, T.; Iqbal, K.; Varma, A.; Thakur, I.S.; Mishra, A. Biodegradation of methyl red dye by a novel actinobacterium Zhihengliuella sp. ISTPL4: Kinetic studies, isotherm and biodegradation pathway. Environ. Technol. Innov. 2022, 26, 102348. [Google Scholar] [CrossRef]
- Zhuang, M.; Sanganyado, E.; Zhang, X.; Xu, L.; Zhu, J.; Liu, W.; Song, H. Azodye degrading bacteria tolerant to extreme conditions inhabit nearshore ecosystems:Optimization and degradation pathways. J. Environ. Manage. 2020, 261, 110222. [Google Scholar] [CrossRef]
- Kamal, I.M.; Abdeltawab, N.F.; Ragab, Y.M.; Farag, M.A.; Ramadan, M.A. Biodegradation, decolorization, and detoxification of Di-azo dye direct Red 81 by Halotolerant, alkali-thermo-tolerant bacterial mixed cultures. Microorganisms 2022, 10, 994. [Google Scholar] [CrossRef]
- Mirbolooki, H.; Amirnezhad, R.; Pendashteh, A.R. Treatment of high saline textile wastewater by activated sludge microorganisms. J. Appl. Res. Technol. 2017, 15, 167–172. [Google Scholar] [CrossRef]
- Gianolini, J.E.; Britos, C.N.; Mulreedy, C.B.; Trelles, J.A. Hyperstabilization of a thermophile bacterial laccase and its applicationfor industrial dyes degradation. 3 Biotech 2020, 10, 288. [Google Scholar] [CrossRef]
- Chen, G.; An, X.; Li, H.; Lai, F.; Yuan, E.; Xia, X.; Zhang, Q. Detoxification of Azo Dye Direct Black G by Thermophilic Anoxybacillus Sp. PDR2 and its application potential in bioremediation. Ecotoxicol. Environ. Saf. 2021, 214, 112084. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Wang, Y.; Yang, G.; Luo, H.; Zhuang, L.; Yu, Z.; Zhou, S. Complete genome sequence of the dissimilatory azo reducing thermophilic bacterium Novibacillus Thermophiles SG-1. J. Biotechnol. 2018, 284, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Guo, X.; Qian, D.K.; Sun, T.; Zhang, W.; Dai, K.; Zeng, R.J. Decolorization of Acid Orange 7 by extreme-thermophilic mixed culture. Bioresour. Technol. 2019, 291, 121875. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Geng, Z.Q.; Dai, K.; Guo, X.; Zeng, R.J.; Fang Zhang, F. Decoupling mechanism of Acid Orange 7 decolorization and sulfate reduction by a Caldanaerobacter dominated extreme-thermophilic consortium. J. Hazard. Mater. 2021, 419, 126498. [Google Scholar] [CrossRef]
- Rajashekhar, K.K.; Neelagund, S.; Mahadevan, G.D. Novel Geobacillus thermoleovorans KNG 112 Thermophilic bacteria from Bandaru hot spring: A potential producer of thermostable enzymes. Asian J. Pharm. Clin. Res. 2020, 13, 134–141. [Google Scholar]
- Rajashekarappa, K.K.; Mahadevan, G.D.; Neelagund, S.E.; Sathynarayana, M.; Vijaya, D.; Mulla, S.I. Decolorization of amaranth RI and fast red E azo dyes by thermophilic Geobacillus thermoleovorans KNG 112. J. Chem. Technol. Biotechnol. 2021, 97, 482–489. [Google Scholar] [CrossRef]
- Sharma, N.; Leung, I.K. Novel thermophilic bacterial laccase for the degradation of Aromatic organic pollutants. Front. Chem. 2021, 9, 711345. [Google Scholar] [CrossRef]
- Navas, L.E.; Carballo, R.; Levin, L.; Berretta, M.F. Fast decolorization of azo dyes in alkaline solutions by a thermostable metal-tolerant bacterial laccase and proposed degradation pathways. Extremophiles 2020, 24, 705–719. [Google Scholar] [CrossRef]
- Ibrahim, H.K.; Mahdi, M.S.; Ameen, R.S. Study the ability of the newly isolated thermophilic bacterium (Geobacillus Thermoleovorans) Strain Ir1 (Jq912239) in decolonization and degradation of Azo dyes (Methylene Blue and Acid Orange G). Int. J. Drug Deliv. Technol. 2021, 11, 429–433. [Google Scholar]
- Miao, L.-L.; Hou, Y.-J.; Fan, H.-X.; Qu, J.; Qi, C.; Liu, Y.; Li, D.L.; Liu, Z.P. Molecular structural basis for the cold adaptedness of the psychrophilic β-glucosidase BglU in Micrococcus antarcticus. Appl. Environ. Microbiol. 2016, 82, 2021–2030. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elnaby, H.M.; Abou-Elela, G.M.; Ghozlan, H.A.; Hussein, S.H.; Sabry, O.A. Characterization and bioremediation potential of marine Psychrobacter species. Egypt. J. Aquat. Res. 2016, 42, 193–203. [Google Scholar] [CrossRef]
- Lu, L.; Zhao, M.; Wang, T.-N.; Zhao, L.-Y.; Du, M.-H.; Li, T.-L.; Li, D.-B. Characterization and dye decolorization ability of an alkaline resistant and organic solvents tolerant laccase from Bacillus licheniformis LS04. Bioresour. Technol. 2012, 115, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Neetha, J.; Sandesh, K.; Girish Kumar, K.; Chidananda, B.; Ujwal, P. Optimization of Direct Blue-14 dye degradation by Bacillus fermus (Kx898362) an alkaliphilic plant endophyte and assessment of degraded metabolite toxicity. J. Hazard. Mater. 2019, 364, 742–751. [Google Scholar]
- Bhattacharya, A.; Goyal, N.; Gupta, A. Degradation of azo dye methyl red by alkaliphilic, halotolerant Nesterenkonia lacusekhoensis EMLA3: Application in alkaline and salt-rich dyeing effluent treatment. Extremophiles 2017, 21, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Krithika, A.; Gayathri, K.V.; Kumar, D.T.; Doss, C. Mixed azo dyes degradation by an intracellular azoreductase enzyme from alkaliphilic Bacillus subtilis: A molecular docking study. Arch. Microbiol. 2021, 203, 3033–3044. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.K.; Bijan Choudhury, B. Synthetic dyes degradation using lignolytic enzymes produced from Halopiger aswanensis strain ABC_IITR by Solid State Fermentation. Chemosphere 2021, 273, 129671. [Google Scholar] [CrossRef]
- Tian, F.; Guo, G.; Zhang, C.; Yang, F.; Hu, Z.; Liu, C.; Wang, S.W. Isolation, cloning and characterization of an azoreductase and the effect of salinity on its expression in a halophilic bacterium. Int. J. Biol. Macromol. 2019, 123, 1062–1069. [Google Scholar] [CrossRef]
- Guadie, A.; Gessesse, A.; Xia, S. Halomonas sp. strain A55, a novel dye decolorizing bacterium from dye-uncontaminated Rift Valley Soda lake. Chemosphere 2018, 206, 59–69. [Google Scholar] [CrossRef]
- Hulin, Q.; Fengfei, S.; Aiguo, Y.; Jiaxian, L.; Biyu, W.; Ying, L.; Yunyi, X.; Jinping, H.; Bo, X. Biodegradation and detoxification of Azo dyes by halophilic/halotolerant microflora isolated from the salt fields of tibet autonomous region China. Front. Microbiol. 2022, 13, 877151. [Google Scholar]
- Liu, W.; Liu, C.; Liu, L.; You, Y.; Jiang, J.; Zhou, Z.; Dong, Z. Simultaneous decolorization of sulfonated azo dyes and reduction of hexavalent chromium under high salt condition by a newly isolated salt-tolerant strain Bacillus circulans BWL1061. Ecotoxicol. Environ. Saf. 2017, 141, 9–16. [Google Scholar] [CrossRef]
- Song, X.Y.; Liu, F.J.; Zhou, H.B.; Yang, H.L. Biodegradation of Acid Scarlet 3R by a new salt-tolerant strain Alcaligenes faecalis LJ-3: Character, Enzyme and Kinetics analysis. Chem. Biochem. Eng. Q. 2018, 32, 371–381. [Google Scholar] [CrossRef]
- Franciscon, E.; Grossman, M.; Paschoal, J.A.; Reyes, F.G.; Durrant, L. Decolorization and biodegradation of reactive sulfonated azo dyes by a newly isolated Brevibacterium sp. strain VN-15. Springerplus 2012, 1, 37. [Google Scholar] [CrossRef]
- Masarbo, R.S.; Karegoudar, T. Decolourisation of toxic azo dye Fast Red E by three bacterial strains: Process optimisation and toxicity assessment. Int. J. Environ. Anal. Chem. 2020, 102, 2686–2696. [Google Scholar] [CrossRef]
- Ameenudeen, S.; Unnikrishnan, S.; Ramalingam, K. Statistical optimization for the efficacious degradation of reactive azo dyes using acinetobacter baumannii JC359. J. Environ. Manag. 2021, 279, 111512. [Google Scholar] [CrossRef] [PubMed]
- Nachiyar, C.V.; Rajkumar, G.S. Degradation of a tannery and textile dye, Navitan Fast Blue S5R by Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2003, 19, 609–614. [Google Scholar] [CrossRef]
- Bharagava, R.N.; Mani, S.; Mulla, S.I.; Saratale, G.D. Degradation and decolourization potential of an ligninolytic enzyme producing Aeromonas hydrophila for crystal violet dye and its phytotoxicity evaluation. Ecotoxicol. Environ. Saf. 2018, 156, 166–175. [Google Scholar] [CrossRef]
- Suzuki, H. Remarkable diversification of bacterial azoreductases: Primary sequences, structures, substrates, physiological roles, and biotechnological applications. Appl. Microbiol. Biotechnol. 2019, 103, 3965–3978. [Google Scholar] [CrossRef]
- Ngo, A.C.R.; Tischler, D. Microbial Degradation of Azo Dyes: Approaches and Prospects for a Hazard-Free Conversion by Microorganisms. Int. J. Environ. Res. Public Health 2022, 19, 4740. [Google Scholar] [CrossRef]
- Zahran, S.A.; Ali-Tammam, M.; Hashem, A.M.; Aziz, R.K.; Ali, A.E. Azoreductase activity of dye-decolorizing bacteria isolated from the human gut microbiota. Sci. Rep. 2019, 9, 5508. [Google Scholar] [CrossRef]
- Sharma, V.; Upadhyay, L.S.B.; Vasanth, D. Extracellular thermostable Laccase-like enzymes from Bacillus licheniformis strains: Production, purification and characterization. Appl. Biochem. Microbiol. 2020, 56, 420. [Google Scholar] [CrossRef]
- Edoamodu, C.E.; Nwodo, U.U. Decolourization of synthetic dyes by laccase produced from Bacillus sp. NU2. Biotechnol. Biotechnol. Equip. 2022, 36, 95–106. [Google Scholar] [CrossRef]
- Ashraf, F.; Irfan, M.; Shakir, H.A.; Ali, S.; Khan, M. An overview of production and industrial exploitation of bacterial laccases. Punjab Univ. J. Zool. 2020, 35, 147–156. [Google Scholar] [CrossRef]
- Kadam, S.K.; Tamboli, A.S.; Sambhare, S.B.; Jeon, B.H.; Govindwar, S.P. Enzymatic analysis, structural study and molecular docking of laccase and catalase from B. subtilis SK1 after textile dye exposure. Ecol. Inform. 2018, 48, 269–280. [Google Scholar] [CrossRef]
- Panwar, A.; Sheikh, J.N.; Dutta, T. Sustainable denim bleaching by a novel thermostable bacterial laccase. Appl. Biochem. Biotechnol. 2020, 192, 1238–1254. [Google Scholar] [CrossRef]
- Kakkar, P.; Wadhwa, N. Extremozymes used in textile industry. J. Text. Inst. 2021, 113, 2007–2015. [Google Scholar] [CrossRef]
- Sankareswaran, M.; Kaliselvi, K.; Anbalagan, S.; Suganya, M.; Abirami, K. Production, characterization and immobilised dye decolorization of amylase enzyme produced by Bacillus megaterium isolated from soil sample. Int. J. Adv. Res. 2015, 3, 295–305. [Google Scholar]
- Mahdi, M.S.; Ibrahim, H.K.; Nidal, S.; Zbarr, N.S.; Hadi, S.A. Microbiological degradation of food dyes and amylase production, immobilization by Iraqi isolate Anoxybacillus rupiensis strain Ir3 (JQ912241). Int. J. Sci. Res. 2022, 6, 257–262. [Google Scholar]
- Maiti, S.; Kulkarni, K.; Adivarekar, R.V. Biotechnology in textile wet processing. Glo. J. Biomed. Sci. 2018, 2, 7–13. [Google Scholar]
- Raddadi, N.; Cherif, A.; Daffonchio, D.; Neifar, M.; Fava, F. Biotechnological applications of extremophiles, extremozymes and extremolytes. Appl. Microbiol. Biotechnol. 2015, 99, 7907–7913. [Google Scholar] [CrossRef]
- Kavuthodi, B.; Sebastian, D. Review on bacterial production of alkaline pectinase with special emphasis on Bacillus species. Biosci. Biotechnol. Res. Commun. 2018, 11, 18–30. [Google Scholar] [CrossRef]
- Zhu, D.; Adebisi, W.A.; Ahmad, F.; Sethupathy, S.; Danso, B.; Sun, J. Recent Development of Extremophilic Bacteria and Their Application in Biorefinery. Front. Bioeng. Biotechnol. 2020, 12, 483. [Google Scholar] [CrossRef]
- Rathod, M.G.; Pathk, A.P. Efficient decolorization of textile dyes by alkaline protease producing bacterial consortia. Indian J. Mar.Sci. 2018, 47, 1468–1477. [Google Scholar]
- Babu, K.M. Studies on Bioscouring of Cotton. World J. Text. Eng. 2019, 5, 43–52. [Google Scholar]
- Bugg, T.D.H.; Ahmad, M.; Hardiman, E.M.; Singh, R. The emerging role for bacteria in lignin degradation and bio-product formation. Curr. Opin. Biotechnol. 2011, 22, 394–400. [Google Scholar] [CrossRef]
- Zhu, D.; Zhang, P.; Xie, C.; Zhang, W.; Sun, J.; Qian, W.-J.; Yang, B. Biodegradation of alkaline lignin by Bacillus ligniniphilus L1. Biotechnol. Biofuels 2017, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Dumorné, K.; Camacho Córdova, D.; Astorga-Eló, M.; Renganathan, P. Extremozymes: A potential source for industrial applications. J. Microbiol. Biotechnol. 2017, 27, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Takkar, S.; Joshi, N.C.; Shukla, S.; Shukla, K.; Singh, A.; Manikonda, A.; Varma, A. An Integrative approach to study bacterial enzymatic degradation of toxic dyes. Front. Microbiol. 2021, 12, 802544. [Google Scholar] [CrossRef]
- Roy, U.; Manna, S.; Sengupta, S.; Das, P.; Datta, S.; Mukhopadhyay, A.; Bhowal, A. Dye removal using microbial biosorbents. In Green Adsorbents for Pollutant Removal; Crini, G., Lichtfouse, E., Eds.; Springer: Cham, Switzerland, 2018; pp. 253–280. [Google Scholar]
- Pham, V.H.T.; Kim, J.; Shim, J.; Chang, S.; Chung, W. Purification and characterization of strong simultaneous Enzyme production of Protease and α-Amylase from an extremophile-Bacillus sp. FW2 and its possibility in food waste degradation. Fermentation 2022, 8, 12. [Google Scholar] [CrossRef]
- Unuofin, J.O. Treasure from dross: Application of agroindustrial wastes-derived thermo-halotolerant laccases in the simultaneous bioscouring of denim fabric and decolorization of dye bath effluents. Ind. Crops Prod. 2020, 147, 112251. [Google Scholar] [CrossRef]
- van der Zee, F.P.; Villaverde, S. Combined anaerobic-aerobic treatment of azo dyes—A short review of bioreactor studies. Water Res. 2005, 39, 1425–1440. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Group | Name of Bacterial Strain | key Enzymes/Substrates | Type of Dyes | Efficiency | References |
|---|---|---|---|---|---|
| Thermophiles | Geobacillus stearothermophilus ATCC 10149 | Extracellular laccase | Remazol Brilliant Blue R | 90% | [59] |
| Anoxybacillus sp. PDR2 | Quinone oxidoreductase | Direct Black G | 90% | [60] | |
| Nivibacillus thermophiles SG-1 | Gene encoding riboflavin biosynthesis protein | Azo dye (Orange I) | 100% | [61] | |
| Consortium of Caldanaerobacter and Pseudomonas | not reported | Acid Orange 7 | 90% | [62] | |
| Caldanaerobacter | Xylose | Acid Orange 7 | 97% | [63] | |
| Geobacillus thermoleovorans KNG 112 | not reported | amaranth R.I and red E azo dyes | ~98% | [64,65] | |
| Geobacillus yumthangensis | Laccase | Organic dyes, including Alizarin, Acid red 27, Congo red, bromophenol blue, Coomassie brilliant blue R-250, Malachite green, and Indigo carmine | ~99% | [66] | |
| Thermus sp. | Laccase | xylidine | 98% | [67] | |
| Geobacillus thermoleovorans | Not reported | Methylene Blue and Acid Orange G | 100% | [68] | |
| Psychrophiles | Micrococcus antarcticus | Psychrophilic β-glucosidase | Starch stain | not reported | [69] |
| Psychrobacter almentarius | Not reported | Reactive Black 5, Reactive Golden Ovifix, and Reactive Blue BRS | ~100% | [53] | |
| Psychrobacter sp. | Not reported | Fast orange, Methanil yellow, and Acid fast red | ~85% | [70] | |
| Zhihengliuella sp. | Lignin peroxidase and laccase | Methyl red | 98.87% | [55] | |
| Bacillus sp. | Azo-reductase | Amido black 10B, Evans blue, Janus green, Methyl orange, Methyl red, and orange G) | ~98% | [52] | |
| Alkaliphiles | Bacillus licheniformis LS04 | Laccase | Reactive black 5 | >80% | [71] |
| Bacillus fermus | Not reported | Direct Blue-14 | >92.76% | [72] | |
| Nesterenkonia lacusekhoensis EMLA3 | Not reported | Methyl red | 97% | [73] | |
| Pseudomonas mendocina | Laccase | Mixture of reactive red (RR), Reactive brown (RB), and Reactive black (RBL) | 58.40% | [35] | |
| Bacillus subtilis | Intracellular azoreductase | Mixed azo dyes | 87.35% | [74] | |
| Halopiger aswanensis | Lignin Peroxidase (LiP), Manganese Peroxidase (MnP), and laccase | Malachite Green, Methyl orange | ~93% | [75] | |
| Halophiles | Halomonas sp | Azoreductase | Acid Brilliant Scarlet GR | [76] | |
| Halomonas sp. strain A55 | Not reported | different dyes | ~100% | [77] | |
| Nesterenkonia lacusekhoensis EMLA3 | Not reported | methyl red | 97% | [73] | |
| Mixture of Enterococcus, unclassified Enterobacteriaceae, Staphylococcus, Bacillus, and Kosakonia. | Laccase, lignin peroxidase, manganese peroxidase, azo reductase, and NADH–DCIP reductase | Congo red, Direct Black G (DBG), Amaranth, Methyl red, and Methyl orange | ~100% | [78] | |
| Bacillus circulans BWL1061 | Azoreductase, NADH-DCIP reductase, and laccase | Methyl orange | >90% | [79] | |
| Alcaligenes faecalis | Azoreductase, laccase and NADH-DCIP (nicotinamide adenine dinucleotide-dichlorophenol indophenols) reductase | Acid Scarlet 3R | ~100% | [80] | |
| Bacillus sp. strain CH12 | Not reported | Reactive Red 239 | ~100% | [48] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, V.H.T.; Kim, J.; Chang, S.; Bang, D. Investigating Bio-Inspired Degradation of Toxic Dyes Using Potential Multi-Enzyme Producing Extremophiles. Microorganisms 2023, 11, 1273. https://doi.org/10.3390/microorganisms11051273
Pham VHT, Kim J, Chang S, Bang D. Investigating Bio-Inspired Degradation of Toxic Dyes Using Potential Multi-Enzyme Producing Extremophiles. Microorganisms. 2023; 11(5):1273. https://doi.org/10.3390/microorganisms11051273
Chicago/Turabian StylePham, Van Hong Thi, Jaisoo Kim, Soonwoong Chang, and Donggyu Bang. 2023. "Investigating Bio-Inspired Degradation of Toxic Dyes Using Potential Multi-Enzyme Producing Extremophiles" Microorganisms 11, no. 5: 1273. https://doi.org/10.3390/microorganisms11051273