
Natural Killer Cells and Cytotoxic T Cells: Complementary Partners against Microorganisms and Cancer

 , ,
, ,
Abstract
:1. Introduction
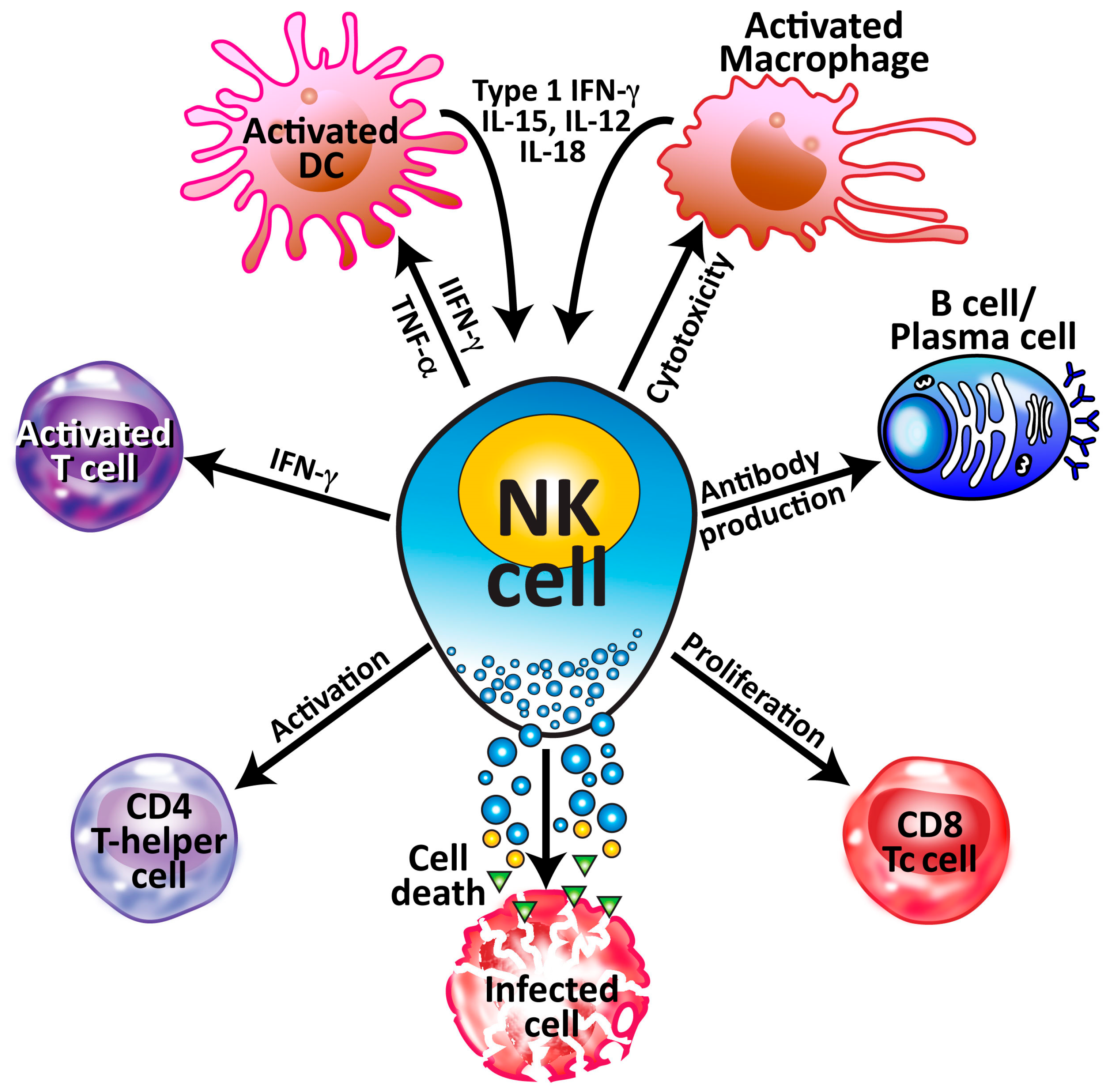
2. Natural Killer (NK) Cells and Their Subpopulations
2.1. What Are CD56+CD16dim NK Cells?
2.2. What Are CD56dimCD16+ NK Cells?
2.3. What Are CD57+ Cells?
2.4. What Are CD57+CD16+ NK Cells?
- Elevation in CD57+CD16+ cells is an indication of persistent antigenic stimulation, especially from viruses. Human CMV is one of the clearest examples of infection driving NK cell differentiation, particularly driving the expansion of NKG2C+ NK cells, which preferentially acquire CD57. Mature CD57+ NK cells expand as a consequence of lifetime exposure to infections, including HBV, HCV, EBV, CMV, hantavirus, and HIV. Their increase is an indication of ongoing or previous viral infection. Elevation in the levels of CD57+ NK cells observed in patients with chronic viral infection correlates with slower disease progression [48,50].
3. Cytotoxic CD8+ T Cells or Cytotoxic T Lymphocytes (CD8+, CTL, TC)
3.1. Important CD8+ T-Cell Subsets
3.2. What Are CD3+CD16+CD56+ NKT Cells?
3.3. How the Innate and Adaptive Immune Systems Work Together to Protect the Body against Different Pathogens
- In the absence of a functional innate and adaptive immune response, pathogens in infected cells or in their tissue environment grow exponentially. This exponential growth of the pathogens may result in serious inflammation, a cytokine storm, and death.
- Upon the detection of pathogens by antigen-presenting T-helper, NK cells, cytotoxic T cells and NKT cells become activated and produce significant amounts of IFN-γ within minutes. The job of interferon is to stop the spread of the pathogens before the NK cells and cytotoxic T cells arrive at the battleground.
- This production of IFN-γ by different cells inhibits pathogen replication, resulting in the flattening of the pathogen curve until the NK cells arrive at the site of the infection.
- The arrival of CD56+ and CD16+ NK cells at the infected area not only stops the replication of pathogens but also causes a significant decline in their numbers by killing the pathogens and the pathogen-infected cells.
- To finish the job, CD57+ and CD57+CD16+ cells, which are highly cytotoxic, are now summoned to join the operation in order to cleanse the body of the pathogen’s remnants, and, thus, the immune system prevails against infections.
- All these steps of pathogen elimination are part of the innate immune response in which the responding cytotoxic activity is carried out within minutes to hours. However, if the NK cell and its subpopulations fail to destroy the pathogen entirely, the immune system must call on additional resources.
- These are the complementary components of adaptive immunity, such as cytotoxic T cells, and their hybrids with CD57, the CD57+CD8+ cytotoxic T cells, will arrive at the pathogen-infected area within a few days of infection.
4. Mechanisms by Which NK Cells Defend the Body against Microbes
4.1. The Direct Antimicrobial Pathway
4.2. The Indirect Antimicrobial Pathway
5. How NK Cells and Cytotoxic T Cells Recognize and Eliminate Viral-Infected and Tumor Cells
5.1. How NK Cells Eliminate Target Cells
5.1.1. Stage 1—Target Cell Identification
5.1.2. Stage 2—Formation of Immunological Synapse
5.1.3. Stage 3—NK-Cell-Induced Target Cell Death
5.1.4. Stage 4—Post-Apoptosis Detachment of the Killer Cell
5.2. How CD8+ T Cells Eliminate Target Cells
6. Potential Clinical and Therapeutic Applications of NK and CD8+ Cells
- One such approach is the use of NK cells for cancer immunotherapy. Advances in viral transduction and electroporation now allow for detailed characterization of genetically modified NK cells and provide a better understanding of how these cells can be used clinically to optimize their capacity to induce tumor regression in vivo through approaches such as improving NK-cell persistence via autocrine IL-2 and IL-15 stimulation, enhancing tumor targeting by silencing inhibitory NK-cell receptors such as NKG2A, and redirecting tumor killing via chimeric antigen receptors [260]. A review by Vishwashrao et al. focused on the use of CARs, BiKEs, and TriKEs to engineer NK cells for tumor immunotherapy. CARs are chimeric antigen receptors that can improve the cytotoxicity of effector cells by specifically redirecting their destructive capability towards a defined target antigen on a tumor or transformed cell. The addition of CAR expression to existing NK-cell-activating receptors could enhance NK cells’ ability to eliminate targeted tumor cells, especially in solid tumors often resistant to NK-cell-mediated killing. BiKEs and TriKEs are bispecific and trispecific killer engagers that can further enhance cytotoxic killing and cytokine production [261]. In a very recent review, Maia et al. described how genetic modifications can enhance NK cells’ tumor-targeting capability, cytotoxicity, persistence, and tumor infiltration, and also prevent exhaustion, as well as detailed both non-viral (electroporation, lipid nanoparticles, lipofection, DNA transposons) and viral (lentivirus, gamma retrovirus, adeno-associated virus) technologies for genetically modifying NK cells [262].
- The second approach is therapeutic manipulation of NK cells in infectious diseases. The anticancer effect of NK cells is already being investigated in multiple clinical trials, but relatively little is known about the therapeutic utilization of NK cells in patients suffering from infectious complications. In vivo data clearly show that NK cells are active against viral, bacterial, and fungal pathogens, and the adoptive transfer of NK cells seems to be a promising methodology. However, although animal studies have been encouraging, it should be remembered that purposeful activation of the host immune system has been shown to have severe complications in humans. Again, more research is needed to fully characterize the optimal patient population, the best time point of the therapy, and the best approach to generating NK cells for immunotherapy [21].
- The third approach is genetic manipulation of NKT cells in autoimmunity. Several autoimmune diseases show defects in the functionality of NKT cells. Attempts have been made to utilize molecules’ ability to activate NKT cells. For instance, in mice, injection of the glycolipid α-GalCer led to upregulation of iNKT cells, which, in turn, led to the activation of NK cells through IFN-γ produced by the iNKT cells. However, further research is needed as repeated injections seemed to exacerbate some autoimmune diseases, and tests in humans seemed to be less effective than in mice. On the other hand, increasing the number of iNKT cells through adoptive transfer seemed to be safe and well-tolerated in human trials. Another method tested is the promotion of self-ligands that activate iNKT cells [263].
- A fourth approach is the therapeutic transfer of ex vivo expanded ILC2s to induce tumor cell death. In a major breakthrough reported by Li et al. [264], the researchers found that as a member of the cytolytic immune effector cell family, human ILC2s secrete granzymes that lyse tumor cells by inducing pyroptosis and/or apoptosis of tumor cells. Unlike CAR T-cell therapy, which requires specific characteristics, ILC2 can be isolated from healthy donors, which is a huge advantage. Thus, in their experiments, Li et al. isolated cells from the blood and expanded them by 2000-fold in culture. The expanded ILC2s were exogenously administered to patients with leukemia, and to solid tumor models in vitro and in vivo. Their findings suggested the potential usefulness of ex vivo-expanded human ILC2s as an adoptive cell strategy for cancer immunotherapy.
7. Summary and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Greenberg, A.H.; Hudson, L.; Shen, L.; Roitt, I.M. Antibody-dependent cell mediated cytotoxicity due to a “Null” Lymphoid Cell. Nat. New Biol. 1973, 242, 111–113. [Google Scholar] [CrossRef] [PubMed]
- Cózar, B.; Greppi, M.; Carpentier, S.; Narni-Mancinelli, E.; Chiossone, L.; Vivier, E. Tumor-infiltrating natural killer cells. Cancer Discov. 2021, 11, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, A.H. The origins of the NK cell, or a Canadian in King Ivan’s court. Clin. Investig. Med. 1994, 17, 626–631. [Google Scholar]
- Greenberg, A.H.; Playfair, J.H.L. Spontaneously arising cytotoxicity to the P-815-Y mastocytoma in NZB mice. Clin. Exp. Immunol. 1974, 16, 99–110. [Google Scholar] [PubMed]
- Kiessling, R.; Klein, E.; Wigzell, H. “Natural” killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype. Eur. J. Immunol. 1975, 5, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Cantor, H.; Boyse, E.A. Functional subclasses of T-lymphocytes bearing different Ly antigens. I. The generation of functionally distinct T-cell subclasses is a differentiative process independent of antigen. J. Exp. Med. 1975, 141, 1376–1389. [Google Scholar] [CrossRef] [PubMed]
- Shiku, H.; Kisielow, P.; Bean, M.A.; Takahashi, T.; Boyse, E.A.; Oettgen, H.F.; Old, L.J. Expression of T-cell differentiation antigens on effector cells in cell-mediated cytotoxicity in vitro. Evidence for functional heterogeneity related to the surface phenotype of T cells. J. Exp. Med. 1975, 141, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Kisielow, P.; Hirst, J.A.; Shiku, H.; Beverley, P.C.; Hoffman, M.K.; Boyse, E.A.; Oettgen, H.F. Ly antigens as markers for functionally distinct subpopulations of thymus-derived lymphocytes of the mouse. Nature 1975, 253, 219–220. [Google Scholar] [CrossRef]
- Masopust, D.; Vezys, V.; Wherry, E.J.; Ahmed, R. A brief history of CD8 T cells. Eur. J. Immunol. 2007, 37, S103–S110. [Google Scholar] [CrossRef]
- Vivier, E. The discovery of innate lymphoid cells. Nat. Rev. Immunol. 2021, 21, 616. [Google Scholar] [CrossRef]
- Spits, H.; Artis, D.; Colonna, M.; Diefenbach, A.; Di Santo, J.P.; Ebert, G.; Koyasu, S.; Locksley, R.M.; McKenzie, A.N.J.; Mebius, R.E.; et al. Innate lymphoid cells—A proposal for uniform nomenclature. Nat. Rev. Immunol. 2013, 13, 145–149. [Google Scholar] [CrossRef]
- Mjosberg, J.; Roncaralo, M.G.; Blom, B. Hergen Spits—A legend at the top of his career. Allergy 2021, 76, 1925–1928. [Google Scholar] [CrossRef] [PubMed]
- Bozzano, F.; Perrone, C.; Moretta, L.; De Maria, A. NK cell precursors in human bone marrow in health and inflammation. Front. Immunol. 2019, 10, 02045. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.M.; Adams, N.M.; Geary, C.D.; Weizman, O.E.; Rapp, M.; Pritykin, Y.; Leslie, C.S.; Sun, J.C. Epigenetic control of innate and adaptive immune memory. Nat. Immunol. 2018, 19, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Mujal, A.M.; Delconte, R.B.; Sun, J.C. Natural killer cells: From innate to adaptive features. Annu. Rev. Immunol. 2021, 39, 417–447. [Google Scholar] [CrossRef]
- Sun, J.C.; Beilke, J.N.; Lanier, L.L. Adaptive immune features of natural killer cells. Nature 2009, 457, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Santosa, E.K.; Kim, H.; Rückert, T.; Le Luduec, J.-B.; Abbasi, A.J.; Wingert, C.K.; Peters, L.; Frost, J.N.; Hsu, K.C.; Romagnani, C.; et al. Control of nutrient uptake by IRF4 orchestrate innate immune memory. Nat. Immunol. 2023, 24, 1685–1697. [Google Scholar] [CrossRef]
- Basilio-Queiros, D.; Mischak-Weissinger, E. Natural killer cells—From innate cells to the discovery of adaptability. Front. Immunol. 2023, 14, 1172437. [Google Scholar] [CrossRef]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef]
- Orange, J.S. Natural killer cell deficiency. J. Allergy Clin. Immunol. 2013, 132, 515–526. [Google Scholar] [CrossRef]
- Schmidt, S.; Tramsen, L.; Rais, B.; Ulrich, E.; Lehrnbecher, T. Natural killer cells as a therapeutic tool for infectious diseases—Current status and future perspectives. Oncotarget 2018, 9, 20891–20897. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Ulrich, E.; Bochennek, K.; Zimmermann, S.-Y.; Lehrnbrecher, T. Role of natural killer cells in antibacterial immunity. Expert Rev. Hematol. 2016, 9, 1119–1127. [Google Scholar] [CrossRef]
- Paul, S.; Lal, G. The molecular mechanism of natural killer cells function and its importance in cancer immunotherapy. Front. Immunol. 2017, 8, 1124. [Google Scholar] [CrossRef] [PubMed]
- Lanier, L.L. Up on the tightrope: Natural killer cell activation and inhibition. Nat. Immunol. 2008, 9, 495–502. [Google Scholar] [CrossRef]
- Mody, C.H.; Ogbomo, H.; Xiang, R.F.; Kyei, S.K.; Feehan, D.; Islam, A.; Li, S.S. Microbial killing by NK cells. J. Leukoc. Biol. 2019, 105, 1285–1296. [Google Scholar] [CrossRef]
- Lodoen, M.B.; Lanier, L.L. Natural killer cells as an initial defense against pathogens. Curr. Opin. Immunol. 2006, 18, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Garcia-Penarrubia, P.; Koster, F.T.; Kelley, R.O.; McDowell, T.D.; Bankhurst, A.D. Antibacterial activity of human natural killer cells. J. Exp. Med. 1989, 169, 99–113. [Google Scholar] [CrossRef]
- Ogbomo, H.; Mody, C.H. Granule-dependent natural killer cell cytotoxicity to fungal pathogens. Front. Immunol. 2016, 7, 692. [Google Scholar] [CrossRef]
- Stuehler, C.; Kuenzli, E.; Jaeger, V.K.; Baettig, V.; Ferracin, F.; Rajacic, Z.; Kaiser, D.; Bernardini, C.; Forrer, P.; Weiiser, M.; et al. Immune reconstitution after allogeneic hematopoietic stem cell transplantation and association with occurrence and outcome of invasive aspergillosis. J. Infect. Dis. 2015, 212, 959–967. [Google Scholar] [CrossRef]
- Tagliabue, A.; Nencioni, L.; Villa, L.; Boraschi, D. Genetic control of an in vitro- natural cell-mediated activity against Salmonella typhimurium by intestinal and splenic lymphoid cells in mice. Clin. Exp. Immunol. 1984, 56, 531–536. [Google Scholar] [PubMed]
- Klimpel, G.R.; Niesel, D.W.; Klimpel, K.D. Natural cytotoxic effector cell activity against Shigella flexneri-induced HeLa cells. J. Immunol. 1986, 136, 1081–1086. [Google Scholar] [CrossRef]
- Lu, C.C.; Wu, T.S.; Hsu, Y.J.; Chang, C.J.; Lin, C.S.; Chia, J.H.; Wu, T.L.; Huang, T.T.; Martgel, J.; Ojcius, D.M.; et al. NK cells kill mycobacteria directly by releasing perforin and granulysin. J. Leukoc. Biol. 2014, 96, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- Small, C.L.; McCormick, S.; Gill, N.; Kugathasan, K.; Santosuosso, M.; Donaldson, N.; Heinrichs, D.E.; Ashkar, A.; Xing, Z. NK cells play a critical protective role in host defense against acute extracellular Staphyulococcus aureus bacterial infection in the lung. J. Immunol. 2008, 180, 5558–5568. [Google Scholar] [CrossRef] [PubMed]
- Naper, C.; Shegarfi, H.; Inngjerdingen, M.; Rolstad, B. The role of natural killer cells in the defense against Listeria monocytogenes lessons from a rat model. J. Innate Immun. 2011, 3, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Feenan, D.D.; Jamil, K.; Polyal, M.J.; Ogbomo, H.; Hasell, M.; Li, S.S.; Xiang, R.F.; Parkins, M.; Trapani, J.A.; Harrison, J.J.; et al. Natural killer cells kill extracellular Pseudomonas aeruginosa using contact-dependent release of granzymes B and H. PLoS Pathog. 2022, 18, e1010325. [Google Scholar] [CrossRef]
- Voigt, J.; Kurzai, O. The role of natural killer cells in the immune response against Candida albicans. Mycoses 2011, 54, 413–414. [Google Scholar]
- Li, S.S.; Ogbomo, H.; Mansour, M.K.; Xiang, R.F.; Szabo, L.; Munro, F.; Mukherjee, P.; Mariuzza, R.A.; Amrein, M.; Vyas, J.M.; et al. Identification of the fungal ligand-triggering cytotoxic PRR-mediated NK cell killing of Cryptococcus and Candida. Nat. Commun. 2018, 9, 751. [Google Scholar] [CrossRef]
- Ma, L.L.; Wang, C.L.; Neely, G.G.; Epelman, S.; Krensky, A.M.; Mody, C.H. NK cells use perforin rather than granulysin for anticryptococcal activity. J. Immunol. 2004, 173, 3357–3365. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Tramsen, L.; Hanisch, M.; Latgé, J.P.; Huenecke, S.; Koehl, U.; Lehrnbecher, T. Human natural killer cells exhibit direct activity against Aspergillus fumigatus hyphae, but not against resting conidia. J. Infect. Dis. 2011, 203, 430–435. [Google Scholar] [CrossRef]
- Bouzani, M.; Ok, M.; McCormick, A.; Ebel, F.; Kurzai, O.; Morton, C.O.; Einsele, H.; Loeffler, J. Human NK cells display important antifungal activity against Aspergillus fumigatus, which is directly mediated by IFN-γ release. J. Immunol. 2011, 187, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, B.E.; Murphy, J.W. In vitro effects of natural killer cells against Paracoccidioides brasiliensis yeast phase. Infect. Immun. 1984, 46, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Petkus, A.F.; Baum, L.L. Natural killer cell inhibition of young spherules and endospores of Coccidioides immitis. J. Immunol. 1987, 139, 3107–3111. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, C.M.; Williams, C.B.; Calderon, V.E.; Huante, M.B.; Moen, S.T.; Popov, V.L.; Baze, W.B.; Peterson, J.W.; Endsley, J.J. Antibacterial role for natural killer cells in host defense to Bacillus anthracis. Infect. Immun. 2012, 80, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.N.; Zheng, M.; Ruan, S.; Kolls, J.; D’Souza, A.; Shellito, J.E. Memory CD4+ T cells are required for optimal NK cell effector functions against the opportunistic fungal pathogen Pneumocystis murina. J. Immunol. 2013, 190, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Pierce, S.; Geanes, E.S.; Bradley, T. Targeting natural killer cells for improved immunity and control of the adaptive immune response. Front. Cell. Infect. Microbiol. 2020, 10, 231. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Xu, W. Cluster of differentiation. In Comprehensive Biomedical Physics; Brahme, A., Ed.; Elsevier: Stockholm, Sweden, 2014; Volume 10, pp. 91–114. [Google Scholar]
- Nielsen, C.M.; White, M.J.; Goodier, M.R.; Riley, E.M. Functional significance of CD57 expression on human NK cells and relevance to disease. Front. Immunol. 2013, 4, 422. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, O.A.; Gonzalez-Hinojosa, M.D.R.; Arakawa-Hoyt, J.S.; Millan, A.J.; Gotthardt, D.; Nabekura, T.; Lanier, L.L. The CD16 and CD32b Fc-gamma receptors regulate antibody-mediated responses in mouse natural killer cells. J. Leukoc. Biol. 2023, 113, 27–40. [Google Scholar] [CrossRef]
- Lopez-Vergés, S.; Milush, J.M.; Pandey, S.; York, V.A.; Arakawa-Hoyt, J.; Pircher, H.; Norris, P.J.; Nixon, D.F.; Lanier, L.L. CD57 defines a functionally distinct population of mature NK cells in the human CD56dimCD16+ NK-cell subset. Blood 2010, 116, 3865–3874. [Google Scholar] [CrossRef] [PubMed]
- Björkström, N.K.; Strunz, B.; Ljungren, H.-G. Natural killer cells in antiviral immunity. Nat. Rev. Immunol. 2022, 22, 112–123. [Google Scholar] [CrossRef]
- Ozcimen, E.E.; Kiyici, H.; Uckuyu, A.; Yanik, F.F. Are CD57+ natural killer cells really important in early pregnancy failure? Arch. Gynecol. Obstet. 2009, 279, 493–497. [Google Scholar] [CrossRef]
- Lu, Z.; Tian, Y.; Bai, Z.; Liu, J.; Zhang, Y.; Qi, J.; Jin, M.; Zhu, J.; Li, X. Increased oxidative stress contributes to impaired peripheral CD56dimCD57+ NK cells from patients with systemic lupus erythematosus. Arthritis Res. Ther. 2022, 24, 48. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Coleman, R.A.; Alber, C.; Ballas, Z.K.; Waldschmidt, T.J.; Mortari, F.; LaBrecque, D.R.; Cook, R.T. TH1 cytokine response of CD57+ T-cell subsets in healthy controls and patients with alcoholic liver disease. Alcohol 2001, 24, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.C.; Jung, M.K.; Yu, H.T.; Kwon, J.S.; Kwak, J.E.; Park, S.H.; Kim, I.C.; Park, M.S.; Lee, S.K.; Choi, S.W.; et al. Increased frequency of CD4+CD57+ senescent T cells in patients with newly diagnosed acute heart failure: Exploring new pathogenic mechanisms with clinical relevance. Sci. Rep. 2019, 9, 12887. [Google Scholar] [CrossRef]
- Kiu, B.; Yang, G.X.; Sun, Y.; Tomiyama, T.; Zhang, W.; Leung, P.S.C.; He, X.S.; Dhaliwal, S.; Invernizzi, P.; Gershwin, M.E.; et al. Decreased CD57 expression of natural killer cells enhanced cytotoxicity in patients with primary sclerosing cholangitis. Front. Immunol. 2022, 13, 912961. [Google Scholar] [CrossRef]
- Künemund, V.; Jungalwala, F.B.; Fischer, G.; Chou, D.K.; Keilhauer, G.; Schachner, M. The L2/HNK-1 carbohydrate of neural cell adhesion molecules is involved in cell interactions. J. Cell Biol. 1988, 106, 123–213. [Google Scholar] [CrossRef] [PubMed]
- Vesely, M.D.; Kershaw, M.H.; Schreiber, R.D.; Smyth, M.J. Natural innate and adaptive immunity to cancer. Annu. Rev. Immunol. 2011, 29, 235–271. [Google Scholar] [CrossRef] [PubMed]
- Strioga, M.; Pasukoniene, V.; Characiejus, D. CD8+CD28− and CD8+CD57+ T cells and their role in health and disease. Immunology 2011, 134, 17–32. [Google Scholar] [CrossRef]
- Characiejus, D.; Pasukoniene, V.; Kazlauskaite, N.; Valuckas, K.P.; Petraitis, T.; Mauricas, M.; Den Otter, W. Predictive value of CD8highCD57+ lymphocyte subset in interferon therapy of patients with renal cell carcinoma. Anticancer Res. 2002, 22, 3679–3683. [Google Scholar] [PubMed]
- Characiejus, D.; Pasukoniene, V.; Jonusauskaite, R.; Azlauskaite, N.; Aleknavicius, E.; Mauricas, M.; Den Otter, W. Peripheral blood CD8highCD57+ lymphocyte levels may predict outcome in melanoma patients treated with adjuvant interferon-alpha. Anticancer Res. 2008, 28, 1139–1142. [Google Scholar] [PubMed]
- Akagi, J.; Baba, H. Prognostic value of CD57(+) T lymphocytes in the peripheral blood of patients with advanced gastric cancer. Int. J. Clin. Oncol. 2008, 13, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Sze, D.M.; Brown, R.D.; Yuen, E.; Gibson, J.; Ho, J.; Raitakari, M.; Basten, A.; Joshua, D.E.; de St Groth, B.F. Clonal cytotoxic T cells in myeloma. Leuk. Lymphoma 2003, 44, 1667–1674. [Google Scholar] [CrossRef]
- Van den Hove, L.E.; Vandenberghe, P.; Van Gool, S.W.; Ceuppens, J.L.; Demuynck, H.; Verhoef, G.E.; Boogaerts, M.A. Peripheral blood lymphocyte subset shifts in patients with untreated hematological tumors: Evidence for systemic activation of the T cell compartment. Leuk. Res. 1998, 22, 175–184. [Google Scholar] [CrossRef]
- Atayar, C.; Poppema, S.; Visser, L.; van den Berg, A. Cytokine gene expression profile distinguishes CD4+/CD57+ T cells of the nodular lymphocyte predominance type of Hodgkin’s lymphoma from their tonsillar counterparts. J. Pathol. 2006, 208, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Serrano, D.; Monteiro, J.; Allen, S.L.; Kolitz, J.; Schulman, P.; Lichtman, S.M.; Buchbinder, A.; Vinciguerra, V.P.; Chiorazzi, N.; Gregersen, P.K. Clonal expansion within the CD4+CD57+ and CD8+CD57+ T cell subsets in chronic lymphocytic leukemia. J. Immunol. 1997, 158, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Di Girolamo, W.; Coronato, S.; Portiansky, E.; Laguens, G. Profile of immune cells in lymph nodes draining human malignant tumors. Medicina 2008, 68, 423–427. [Google Scholar] [PubMed]
- Muntasell, A.; Servitjia, S.; Cabo, M.; Bermejo, B.; Pérez-Buira, S.; Rojo, F.; Costa-Garcia, M.; Arpi, O.; Moraru, M.; Serrano, L.; et al. High numbers of circulating CD57+ NK cells associate with resistance to HER2- specific therapeutic antibodies in HER2+ primary breast cancer. Cancer Immunol. Res. 2019, 7, 1280–1292. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, N.; Bagheri, S.; Akbarzadeh, A. Prognostic implication of CD57, CD16 and TGF-β expression in oral squamous cell carcinoma. J. Oral Pathol. Med. 2016, 45, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Stabile, H.; Fionda, C.; Gismondi, A.; Santoni, A. Role of distinct natural killer cell subsets in anticancer response. Front. Immunol. 2017, 8, 293. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, P.K.; Betts, M.R.; Price, D.A.; Gostick, E.; Horton, H.; Roederer, M.; De Rosa, S.C. The cytolytic enzymes granyzme A, granzyme B, and perforin: Expression patterns, cell distribution, and their relationship to cell maturity and bright CD57 expression. J. Leukoc. Biol. 2009, 85, 88–97. [Google Scholar] [CrossRef]
- Hazeldine, J.; Lord, J.M. The impact of ageing on natural killer cell function and potential consequences for health in older adults. Ageing Res. Rev. 2013, 12, 1069–1078. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.J.; Ruiz, N.L.; Leon, M.L.; Enriquez, L.M.; Velasquez, M.P.M.; Aguirre, J.P.O.; Bohorquez, O.M.R.; Vargas, E.A.V.; Hernandez, E.D.; Lopez, C.A.P. Immunosenescence study of T cells: A systematic review. Front. Immunol. 2021, 11, 604591. [Google Scholar] [CrossRef] [PubMed]
- Giscombe, R.; Wang, X.B.; Kakoulidou, M.; Lefvert, A.K. Characterization of the expanded T-cell populations in patients with Wegener’s granulomatosis. J. Intern. Med. 2006, 260, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Ratts, R.B.; Karandikar, N.J.; Hussain, R.Z.; Choy, J.; Northrop, S.C.; Lovett-Racke, A.E.; Racke, M.K. Phenotypic characterization of autoreactive T cells in multiple sclerosis. J. Neuroimmunol. 2006, 178, 100–110. [Google Scholar] [CrossRef]
- Mikulkova, Z.; Praksova, P.; Stourac, P.; Bednarik, J.; Strajtova, L.; Pacasova, R.; Belobradkova, J.; Dite, P.; Michalek, J. Numerical defects in CD8+CD28- T-suppressor lymphocyte population in patients with type 1 diabetes mellitus and multiple sclerosis. Cell. Immunol. 2010, 262, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhong, W.; Lu, X.; Shi, B.; Zhu, Y.; Chen, L.; Zhang, G.; Zhang, X. Association of Graves’ disease and prevalence of circulating IFN-gamma-producing CD28(-) T cells. J. Clin. Immunol. 2008, 28, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.C.; Lawson, T.M.; Vedhara, K.; Moss, P.A.; Lehner, P.J.; Borysiewicz, L.K. CD8high+(CD57+) T cells in patients with rheumatoid arthritis. Arthritis Rheum. 1997, 40, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Miyake, S.; Kondo, T.; Terao, K.; Hatakenaka, M.; Hashimoto, S.; Yamamura, T. Natural killer type2 bias in remission of multiple sclerosis. J. Clin. Investig. 2001, 107, R23–R29. [Google Scholar] [CrossRef]
- Kastrukoff, L.F.; Morgan, N.G.; Zecchini, D.; White, R.; Petkau, A.J.; Satoh, J.; Paty, D.W. A role for natural killer cells in the immunopathogenesis of multiple sclerosis. J. Neuroimmunol. 1998, 86, 123–133. [Google Scholar] [CrossRef]
- Aramaki, T.; Ida, H.; Izumi, Y.; Fujikawa, K.; Huang, M.; Arima, K.; Tamai, M.; Kamachi, M.; Nakamura, H.; Kawakami, A.; et al. A significantly impaired natural killer cell activity due to a low activity on a per-cell basis in rheumatoid arthritis. Mod. Rheumatol. 2009, 19, 245–252. [Google Scholar] [CrossRef]
- Izumi, Y.; Ida, H.; Huang, M.; Iwanaga, N.; Tanaka, F.; Aratake, K.; Arima, K.; Tamai, M.; Kamachi, M.; Nakamura, H.; et al. Characterization of peripheral natural killer cells in primary Sjogren’s syndrome: Impaired NK cell activity and low NK cell number. J. Lab. Clin. Med. 2006, 147, 242–249. [Google Scholar] [CrossRef]
- Park, Y.W.; Kee, S.J.; Cho, Y.N.; Lee, E.H.; Lee, H.Y.; Kim, E.M.; Shin, M.H.; Park, J.J.; Kim, T.J.; Lee, S.S.; et al. Impaired differentiation and cytotoxicity of natural killer cells in systemic lupus erythematosus. Arthritis Rheum. 2009, 60, 1753–1763. [Google Scholar] [CrossRef]
- Ciampolillo, A.; Guastamacchia, E.; Amati, L.; Magrone, T.; Munno, I.; Jirillo, E.; Triggiani, V.; Fallacara, R.; Tafaro, E. Modifications of the immune responsiveness in patients with autoimmune thyroiditis: Evidence for a systemic immune alteration. Curr. Pharm. Des. 2003, 9, 1946–1950. [Google Scholar] [CrossRef]
- Cameron, A.L.; Kirby, B.; Griffiths, C.E. Circulating natural killer cells in psoriasis. Br. J. Dermatol. 2003, 149, 160–164. [Google Scholar] [CrossRef]
- O’Gorman, M.; Smith, R.; Garrison, A.; Shamiyeh, E.; Pachman, L. Lymphocyte subsets in peripheral blood from newly diagnosed, untreated patients with juvenile dermatomyositis (JDM) are associated with disease activity scores (DAS). Arthritis Rheum. 2002, 46, S490. [Google Scholar]
- Wouters, C.H.P.; Ceuppens, J.L.; Stevens, E.A.M. Different circulating lymphocyte profiles in patients with different subtypes of juvenile idiopathic arthritis. Clin. Exp. Rheumatol. 2002, 20, 239–248. [Google Scholar] [PubMed]
- Wehrmann, W.; Reinhold, U.; Kukel, S.; Franke, N.; Uerlich, M.; Kreysel, H.W. Selective alterations in natural killer cell subsets in patients with atopic dermatitis. Int. Arch. Allergy Appl. Immunol. 1990, 92, 318–322. [Google Scholar] [CrossRef]
- Matsumura, G. Leu7(HNK-1)-positive cells in peripheral blood and natural killer cell activity in patients with atopic dermatitis. Nihon Hifuka Gakkai Zasshi 1990, 100, 57–62. [Google Scholar]
- Struyf, N.J.; Snoeck, H.W.; Bridts, C.H.; De Clerck, L.S.; Stevens, W.J. Natural killer cell activity in Sjogren’s syndrome and systemic lupus erythematosus: Stimulation with interferons and interleukin-2 and correlation with immune complexes. Ann. Rheum. Dis. 1990, 49, 690–693. [Google Scholar] [CrossRef]
- Antonaci, S.; Polignano, A.; Ottolenghi, A.; Tortorella, C.; Schena, F.P. Redistribution of natural killer (NK) cell frequency and NK cytotoxic activity in primary IgA nephropathy. Cytobios 1992, 69, 27–34. [Google Scholar]
- Batista, M.D.; Ho, E.L.; Kuebler, P.J.; Milush, J.M.; Lanier, L.L.; Kallas, E.G.; York, V.A.; Chang, D.; Liao, W.; Unemori, P.; et al. Skewed distribution of natural killer cells in psoriasis skin lesions. Exp. Dermatol. 2013, 22, 64–66. [Google Scholar] [CrossRef]
- Imai, R.; Miura, J.; Numata, K.; Aikawa, Y.; Takamori, K.; Ogawa, H. Analysis of T cell, activated T cell and NK cell subsets in peripheral blood lymphocytes from patients with alopecia areata. In Trends in Human Hair Growth and Alopecia Research; Van Neste, D., Lachapelle, J.M., Antoine, J.L., Eds.; Springer: Berlin/Heidelberg, Germany, 1989; pp. 299–304. [Google Scholar] [CrossRef]
- Stricker, R.B.; Winger, E.E. Decreased CD57 lymphocyte subset in patients with chronic Lyme disease. Immunol. Lett. 2001, 76, 43–48. [Google Scholar] [CrossRef]
- Stricker, R.B.; Burrascano, J.; Winger, E. Longterm decrease in the CD57 lymphocyte subset in a patient with chronic Lyme disease. Ann. Agric. Environ. Med. 2002, 9, 111–113. [Google Scholar]
- Marques, A.; Brown, M.R.; Fleisher, T.A. Natural killer cell counts are not different between patients with post-Lyme disease syndrome and controls. Clin. Vaccine Immunol. 2009, 16, 1249–1250. [Google Scholar] [CrossRef]
- Laboratory Tests and Practices That Are Not Currently Recommended. Available online: https://www.cdc.gov/lyme/diagnosistesting/labtest/otherlab/index.html (accessed on 27 December 2023).
- Lantos, P.M.; Rumbaugh, J.; Bockenstedt, L.K.; Falck-Ytter, Y.T.; Aguero-Rosenfeld, M.E.; Auwaeerter, P.G.; Baldwin, K.; Bannuru, R.R.; Belani, K.K.; Bowie, W.R.; et al. Clinical practice guidelines by the Infectious Diseases Society of America (IDSA), American Academy of Neurology (AAN), and American College of Rheumatology (ACR): 2020 guidelines for the prevention, diagnosis and treatment of Lyme disease. Clin. Infect. Dis. 2021, 72, e1–e48. [Google Scholar] [CrossRef]
- Mygland, A.; Ljostad, U.; Fingerle, V.; Rupprecht, T.; Schmutzhard, E.; Steiner, I. EFNS guidelines on the diagnosis and management of European Lyme neuroborreliosis. Eur. J. Neurol. 2010, 17, 8–16. [Google Scholar] [CrossRef] [PubMed]
- The Royal College of Pathologists of Australasia. Position Statement: Diagnostic Laboratory Testing for Lyme Disease (or Similar Syndromes) in Australia and New Zealand; Microbiol AC, 1/2014; 2022. Available online: https://www.rcpa.edu.au/Library/College-Policies/Position-Statements/Diagnostic-Laboratory-testing-for-Borreliosis-Lyme (accessed on 27 December 2023).
- Galán, M.; Vigón, L.; Fuentes, D.; Murciano-Antón, M.A.; Casado-Fernández, G.; Domínguez-Mateos, S.; Mateos, E.; Ramos-Martín, F.; Planelles, V.; Torres, M.; et al. Persistent overactive cytotoxic immune response in a Spanish cohort of individuals with long COVID: Identification of diagnostic biomarkers. Front. Immunol. 2022, 13, 848886. [Google Scholar] [CrossRef]
- Di Vito, C.; Calcaterra, F.; Colaniz, N.; Terzoli, S.; Voza, A.; Mikulak, J.; Della Bella, S.; Mavilio, D. Natural killer cells in SARS-CoV-2 infection: Pathophysiology and therapeutic implications. Front. Immunol. 2022, 13, 888248. [Google Scholar] [CrossRef] [PubMed]
- Clavarino, G.; Leroy, C.; Epaulard, O.; Raskovalova, T.; Vilotitch, A.; Pernollet, M.; Dumestre-Pérard, C.; Defendi, F.; Le Maréchal, M.; Le Gouellec, A.; et al. Fine analysis of lymphocyte subpopulations in SARS-CoV-2 infected patients: Differential profiling of patients with severe outcome. Front. Immunol. 2022, 13, 889613. [Google Scholar] [CrossRef] [PubMed]
- Varchetta, S.; Mele, D.; Oliviero, B.; Mantovani, S.; Ludovisi, S.; Cerino, A.; Bruno, R.; Castelli, A.; Mosconi, M.; Vecchia, M.; et al. Unique immunological profile in patients with COVID-19. Cell. Mol. Immunol. 2021, 18, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Maucourant, C.; Filipovic, I.; Ponzetta, A.; Aleman, S.; Cornillet, M.; Hertwig, L.; Strunz, B.; Lentini, A.; Reinius, B.; Brownlie, D.; et al. Natural killer cell immunotypes related to COVID-19 disease severity. Sci. Immunol. 2020, 5, eabd6832. [Google Scholar] [CrossRef]
- Li, H.; Geng, L.; Cao, Z.; Jin, Z.; Yan, Q.; Long, X.; Zhang, X.; Meng, D.; Jiang, Y.; Li, W.; et al. CD56brightCD16− to CD57+CD56dimCD16+ NK cell ratio discriminates disease activity and renal involvement in patients with systemic lupus erythematosus. Clin. Exp. Rheumatol. 2023, 41, 1768–1776. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Travers, P.; Walport, M.; Shlomchik, M.J. Immunobiology: The Immune System in Health and Disease, 5th ed.; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Cassioli, C.; Baldari, C.T. The expanding arsenal of cytotoxic T cells. Front. Immunol. 2022, 13, 883010. [Google Scholar] [CrossRef]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8+ T cells in cancer and cancer immunotherapy. Br. J. Cancer 2021, 124, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Jaime-Sanchez, P.; Uranga-Murillo, I.; Aguilo, N.; Khouili, S.C.; Arias, M.A.; Sancho, D.; Pardo, J. Cell death induced by cytotoxic CD8+ T cells is immunogenic and primes caspase-3-dependent spread immunity against endogenous tumor antigens. J. Immunother. Cancer 2020, 8, e000528. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.; Huang, J. CD8+ T cells and NK cells: Parallel and complementary soldiers of immunotherapy. Curr. Opin. Chem. Eng. 2018, 19, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Chijioke, O.; Landtwing, V.; Münz, C. NK cell influence on the outcome of primary Epstein-Barr virus infection. Front. Immunol. 2016, 7, 323. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, S.P.H.; Pardiek, I.N.; Lanfermeijer, J.; Sauce, D.; Klenerman, P.; van Baarle, D.; Arens, R. The hallmarks of CMV-specific CD8 T-cell differentiation. Med. Microbiol. Immunol. 2019, 208, 365–373. [Google Scholar] [CrossRef]
- Bandres, E.; Merino, J.; Vasquez, B.; Inoges, S.; Moreno, C.; Subira, M.L.; Sanchez-Ibarrola, A. The increase of IFN-γ production through aging correlates with the expanded CD8+highCD28-CD57+ subpopulation. Clin. Immunol. 2000, 96, 230–235. [Google Scholar] [CrossRef]
- Sato, M.; Takemura, M.; Shinohe, R.; Yamamoto, Y.; Saito, K. A study of natural killer cell subtractions in Abatacept therapy for rheumatoid arthritis. In Proceedings of the ACR/ARHP Annual Meeting, San Diego, CA, USA, 25–30 October 2013. Abstract 1385. [Google Scholar]
- Huang, B.; Liu, R.; Wang, P.; Yuan, Z.; Yang, J.; Xiong, H.; Zhang, N.; Huang, Q.; Fu, X.; Sun, W.; et al. CD8+CD57+ T cells exhibit distinct features in human non-small cell lung cancer. J. Immunother. Cancer 2020, 8, e000639. [Google Scholar] [CrossRef]
- Olloquequi, J.; Montes, J.F.; Prats, A.; Rodríguez, E.; Montero, M.A.; García-Valero, J.; Ferrer, J. Significant increase of CD57+ cells in pulmonary lymphoid follicles of COPD patients. Eur. Resp. J. 2011, 37, 289–298. [Google Scholar] [CrossRef]
- Mendes, A.V.; Kallas, E.G.; Benard, G.; Pannuti, C.S.; Menezes, R.; Dulley, F.L.; Evans, T.G.; Salomao, R.; Machado, C.M. Impact of cytomegalovirus and grafts versus host disease on the dynamics of CD57+CD28−CD8+ T cells after bone marrow transplant. Clinics 2008, 63, 667–676. [Google Scholar] [CrossRef]
- Björkström, N.K.; Beziat, V.; Cichocki, F.; Liu, L.L.; Levine, J.; Larsson, S.; Koup, R.A.; Anderson, S.K.; Ljunggren, H.G.; Malmberg, K.J. CD8 T cells express randomly selected KIRs with distinct specificities compared with NK cells. Blood 2012, 120, 3455–3465. [Google Scholar] [CrossRef]
- Koh, J.Y.; Shin, E.C. KIRs mark killers suppressing autoimmunity. Immunity 2022, 55, 735–737. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.K.; Rujkijyanont, P.; Neale, G.; Yang, J.; Bari, R.; Das Gupta, N.; Holladay, M.; Rooney, B.; Leung, W. Multiplex and genome-wide analyses reveal distinctive properties of KIR+ and CD56+ T cells in human blood. J. Immunol. 2013, 191, 1625–1636. [Google Scholar] [CrossRef]
- Campilo, J.A.; Martinez-Escribano, J.A.; Moya-Quiles, M.R.; Marin, L.A.; Muro, M.; Guerra, N.; Parrado, A.; Campos, M.; Frias, J.F.; Minguela, A.; et al. Increased expression of the natural killer cell inhibitory receptor CD94/NKG2A and CD158b on circulating and lesional T cells in patients with chronic plaque psoriasis. Br. J. Dermatol. 2006, 155, 318–324. [Google Scholar]
- Li, J.; Zaslavsky, M.; Su, Y.; Guo, J.; Sikora, M.I.; van Unen, V.; Christophersen, A.; Chiou, S.H.; Chen, I.; Li, J.; et al. KIR+CD8+ T cells suppress pathogenic T cells and are active in autoimmune diseases and COVID-19. Sci. Immunol. 2022, 376, eabi9591. [Google Scholar] [CrossRef]
- Paris-Muñoz, A.; León-Triana, O.; Pérez-Martinez, A.; Barber, D.F. Helios as a potential biomarker in systemic lupus erythematosus and new therapies based on immunosuppressive cells. Int. J. Mol. Sci. 2024, 25, 452. [Google Scholar] [CrossRef]
- Hill, T.M.; Bezbradica, J.S.; Van Kaer, L.; Joyce, S. CD1d-Restricted Natural Killer T Cells; John Wiley & Sons, Ltd.: Chichester, UK, 2016. [Google Scholar] [CrossRef]
- Almeida, J.S.; Casanova, J.M.; Santos-Rosa, M.; Tarazona, R.; Solana, R.; Rodrigues-Santos, P. Natural killer T-like cells: Immunobiology and role in disease. Int. J. Mol. Sci. 2023, 24, 2743. [Google Scholar] [CrossRef]
- Liu, X.; Li, L.; Si, F.; Huang, L.; Zhao, Y.; Zhang, C.; Hoft, D.F.; Peng, G. NK and NKT cells have distinct properties and functions in cancer. Oncogene 2021, 40, 4521–4537. [Google Scholar] [CrossRef]
- Jerud, E.S.; Bricard, G.; Porcxelli, S.A. CD1d-restricted natural killer T cells: Roles in tumor immunosurveillance and tolerance. Transfus. Med. Hemother. 2006, 33, 18–36. [Google Scholar] [CrossRef]
- Krijgsman, D.; Hokland, M.; Kuppen, P.J.K. The role of natural killer T cells in cancer—A phenotypical and functional approach. Front. Immunol. 2018, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Metelitsa, L.S.; Wu, H.W.; Wang, H.; Yang, Y.; Warsi, Z.; Asgharzadeh, S.; Groshen, S.; Wilson, S.B.; Seeger, R.C. Natural killer T cells infiltrate neuroblastomas expressing the chemokine CCL2. J. Exp. Med. 2004, 199, 1213–1221. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.S.; Mao, F.Y.; Zhao, Y.L.; Wang, T.T.; Chen, N.; Zhang, J.Y.; Cheng, P.; Li, W.-H.; Lu, Y.-P.; Teng, Y.-S.; et al. Altered phenotypic and functional characteristics of CD3+CD56+ NKT-like cells in human gastric cancer. Oncotarget 2016, 7, 55222–55230. [Google Scholar] [CrossRef] [PubMed]
- Molling, J.W.; Kolgen, W.; Langendijk, J.A.; van der Vliet, H.J.; Boomsma, M.F.; Kruizenga, H.; Smorenberg, C.H.; Molenkamp, B.G.; Langendeijk, J.A.; Leemans, C.R.; et al. Peripheral blood IFN-gamma-secreting Valpha24+Vbeta11+ NKT cell numbers are decreased in cancer patients independent of tumor type or tumor load. Int. J. Cancer 2005, 116, 87–93. [Google Scholar] [CrossRef]
- Molling, J.W.; Langius, J.A.; Langendijk, J.A.; Leemans, C.R.; Bontkes, H.J.; van der Vliet, H.J.; von Blomberg, M.E.; Scheper, R.J.; van den Eertwegh, A.J.M. Low levels of circulating invariant natural killer T cells predict poor clinical outcome in patients with head and neck squamous cell carcinoma. J. Clin. Oncol. 2007, 25, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Klatka, J.; Grywalska, E.; Wasiak, M.; Markowicz, J.; Trojanowski, P.; Olszanski, W.; Rolinski, J. The percentage of iNKT cells among other immune cells at various clinical stages of laryngeal cancer. Postepy Hig. Med. Dosw. 2016, 70, 392–399. [Google Scholar] [CrossRef]
- Guven, H.; Gilljam, M.; Chambers, B.J.; Ljunggren, H.G.; Christensson, B.; Kimby, E.; Dilber, M.S. Expansion of natural killer (NK) and natural killer-like T (NKT)-cell populations derived from patients with B-chronic lymphocytic leukemia (B-CLL): A potential source for cellular immunotherapy. Leukemia 2003, 17, 1973–1980. [Google Scholar] [CrossRef]
- Kumar, A.; Suryadevara, N.; Hill, T.M.; Bezbradeica, J.S.; Van Kaer, L.; Joyce, S. Natural killer T cells: An ecological evolutionary developmental biology perspective. Front. Immunol. 2017, 8, 1858. [Google Scholar] [CrossRef]
- Wu, L.; Van Kaer, L. Natural killer T cells in health and disease. Front. Biosci. 2011, 3, 236–251. [Google Scholar] [CrossRef]
- Wu, L.; Van Kaer, L. Natural killer T cells and autoimmune disease. Curr. Mol. Med. 2009, 9, 4–14. [Google Scholar] [CrossRef]
- Chan, A.C.; Serwecinska, L.; Cochrane, A.; Harrison, L.C.; Godfrey, D.I.; Berzins, S.P. Immune characterization of an individual with an exceptionally high natural killer T cell frequency and her immediate family. Clin. Exp. Immunol. 2009, 156, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, X.; Wang, M.; Zou, Q.; Zhao, S.; Sun, B.; Xu, L.; Jiang, Y. Increased numbers of NK cells, NKT-like cells, and NK inhibitory receptors in peripheral blood of patients with chronic obstructive pulmonary disease. Clin. Dev. Immunol. 2013, 2013, 721782. [Google Scholar] [CrossRef] [PubMed]
- Krijgsman, D.; de Vries, N.L.; Skovbo, A.; Andersen, M.N.; Swets, M.; Bastiaannet, E.; Vahrmeijer, A.L.; van de Velde, C.J.H.; Heemskerk, M.H.M.; Hokland, M.; et al. Characterization of circulating T-, NK-, and NKT cell subsets in patients with colorectal cancer: The peripheral blood immune cell profile. Cancer Immunol. Immunother. 2019, 68, 1011–1024. [Google Scholar] [CrossRef] [PubMed]
- Sokhatska, O.; Padrão, E.; Sousa-Pinto, B.; Beltrão, M.; Mota, P.C.; Melo, N.; Delgado, L.; Morais, A. NK and NKT cells in the diagnosis of diffuse lung diseases presenting with a lymphocytic alveolitis. BMC Pulm. Med. 2019, 19, 39. [Google Scholar] [CrossRef] [PubMed]
- Marrero, I.; Ware, R.; Kumar, V. Type II NKT cells in inflammation, autoimmunity, microbial immunity, and cancer. Front. Immunol. 2015, 6, 316. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhao, X.; Wang, Z.; Wang, J.; Sun, H.; Hu, Y. High circulating CD3+CD56+CD16+ natural killer-like T cell levels predict a better IVF treatment outcome. J. Reprod. Immunol. 2013, 97, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Michel, T.; Théresine, M.; Andrés, E.; Hentges, F.; Zimmer, J. CD56bright natural killer (NK) cells: An important NK cell subset. Immunology 2009, 126, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Azuz-Lieberman, A.; Markel, G.; Mizrahi, S.; Gazit, R.; Hannah, J.; Achdout, H.; Gruda, R.; Katz, G.; Arnon, T.I.; Battat, S.; et al. The involvement of NK cells in ankylosing spondylitis. Int. Immunol. 2005, 17, 837–845. [Google Scholar] [CrossRef]
- Marcenaro, E.; Notarangelo, L.D.; OIrange, J.S.; Vivier, E. Editorial: NK cell subsets in health and disease: New developments. Front. Immunol. 2017, 8, 1363. [Google Scholar] [CrossRef]
- Nitta, T.; Yagita, H.; Sato, K.; Okumura, K. Involvement of CD56 (NKH-1/Leu-19 antigen) as an adhesion molecule in natural killer-target cell interaction. J. Exp. Med. 1989, 170, 1757–1761. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Fehniger, T.A.; Turner, S.C.; Chen, K.S.; Ghaheri, B.A.; Ghayur, T.; Carson, W.E.; Caligiuri, M.A. Human natural killer cells: A unique innate immunoregulatory role for the CD56(bright) subset. Blood 2001, 97, 3146–3151. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Almehmadi, M.; Flanagan, B.F.; Khan, N.; Alomar, S.; Christmas, S.E. Increased numbers and functional activity of CD56+ T cells in healthy cytomegalovirus positive subjects. Immunology 2014, 142, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Khakoo, S.I.; Thio, C.L.; Martin, M.P.; Brooks, C.R.; Gao, X.; Astemborski, J.; Cheng, J.; Goedert, J.J.; Vlahov, D.; Hilgartner, M.; et al. HLA and NK cell inhibitory receptor genes in resolving hepatitis C virus infection. Science 2004, 305, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Schafer, J.L.; Li, H.; Evans, T.I.; Estes, J.D.; Reeves, R.K. Accumulation of cytotoxic CD16+ NK cells in simian immunodeficiency virus-infected lymph nodes associated with in situ differentiation and functional anergy. J. Virol. 2015, 89, 6887–6894. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.; Strauss-Albee, D.M.; Leipold, M.; Kubo, J.; Nemat-Gorgani, M.; Dogan, O.C.; Dekker, C.L.; Mackey, S.; Maecker, H.; Swan, G.E.; et al. Genetic and environmental determinants of human NK cell diversity revealed by mass cytometry. Sci. Transl. Med. 2013, 5, 208ra145. [Google Scholar] [CrossRef] [PubMed]
- Müller-Durovic, B.; Grählert, J.; Devine, O.P.; Akbar, A.N.; Hess, C. CD56-negative NK cells with impaired effector function expand in CMV and EBV co-infected healthy donors with age. Aging 2019, 11, 724–740. [Google Scholar] [CrossRef]
- Mavillo, D.; Lombardo, G.; Benjamin, J.; Kim, D.; Follman, D.; Marcenaro, E.; O’Shea, M.A.; Kinter, A.; Kovacs, C.; Moretta, A.; et al. Characterization of CD56-/CD16+ natural killer (NK) cells: A highly dysfunctional NK subset expanded in HIV-infected viremic individuals. Proc. Natl. Acad. Sci. USA 2005, 102, 2886–2891. [Google Scholar] [CrossRef]
- Gonzalez, V.D.; Falconer, K.; Björkström, N.K.; Blom, K.G.; Weiland, O.; Ljunggren, H.-G.; Alaeus, A.; Sandberg, J.K. Expansion of functionally skewed CD56-negative NK cells in chronic hepatitis C virus infection: Correlation with outcome of pegylated IFN-a and Ribavirin treatment. J. Immunol. 2009, 183, 6612–6618. [Google Scholar] [CrossRef]
- Grier, J.T.; Forbes, L.R.; Monaco-Shawver, L.; Oshinsky, J.; Atkinson, T.P.; Moody, C.; Pandey, R.; Campbell, K.S.; Orange, J.S. Human immunodeficiency-causing mutation defines CD16 in spontaneous NK cell cytotoxicity. J. Clin. Investig. 2012, 122, 3769–3780. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.S.; Eberhard, J.M.; Keudel, P.; Bollmann, B.A.; Ballmaier, M.; Bhatnagar, N.; Zielinska-Skowronek, M.; Schmidt, R.E.; Meyer-Olson, D. HIV infection is associated with a preferential decline in less-differentiated CD56dim CD16+ NK cells. J. Virol. 2010, 84, 1183–1188. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.S.; Ahmad, F.; Eberhard, J.M.; Bhatnagar, N.; Bollmann, B.A.; Keudel, P.; Ballmaier, M.; Zielinska-Skowronek, M.; Schmidt, R.E.; Meyer-Olson, D. Loss of CCR7 expression on CD56bright NK cells is associated with a CD56dimCD16+ NK cell-like phenotype and correlates with HIV viral load. PLoS ONE 2012, 7, e44820. [Google Scholar] [CrossRef] [PubMed]
- Björkström, N.K.; Ljunggren, H.G.; Sandberg, J.K. CD56 negative NK cells: Origin, function, and role in chronic viral disease. Trend Immunol. 2010, 31, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Björkström, N.K.; Riese, P.; Heuts, F.; Andersson, S.; Fauriat, C.; Ivarsson, M.A.; Björklund, A.T.; Flodström-Tullberg, M.; Michaëlsson, J.; Rottenberg, M.E.; et al. Expression patterns of NKG2A, KIR, and CD57 define a process of CD56dim NK-cell differentiation uncoupled from NK-cell education. Blood 2010, 116, 3853–3864. [Google Scholar] [CrossRef] [PubMed]
- Phetsouphanh, C.; Aldridge, D.; Marchi, E.; Munier, C.M.L.; Meyerowitz, J.; Murray, L.; Van Vuuren, C.; Goedhals, D.; Fidler, S.; Kelleher, A.; et al. Maintenance of functional CD57+ cytolytic CD4+ T cells in HIV+ elite controllers. Front. Immunol. 2019, 10, 1844. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Vergès, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.-M.; Norris, P.J.; et al. Expansion of a unique CD57⁺NKG2Chi natural killer cell subset during acute human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725–14732. [Google Scholar] [CrossRef]
- Gratama, J.W.; Kluin-Nelemans, H.C.; Langelaar, R.A.; den Ottolander, G.J.; Stijnen, T.; D’Amaro, J.; Torensma, R.; Tanke, H.J. Flow cytometric and morphologic studies of HNK1+ (Leu 7+) lymphocytes in relation to cytomegalovirus carrier status. Clin. Exp. Immunol. 1988, 74, 190–195. [Google Scholar]
- Foley, B.; Cooley, S.; Verneris, M.R.; Pitt, M.; Curtsinger, J.; Luo, X.; Lopez-Vergès, S.; Lanier, L.L.; Weisdorf, D.; Miller, J.S. Cytomegalovirus reactivation after allogeneic transplantation promotes a lasting increase in educated NKG2C+ natural killer cells with potent function. Blood 2012, 119, 2665–2674. [Google Scholar] [CrossRef]
- Foley, B.; Cooley, S.; Verneris, M.R.; Curtsinger, J.; Luo, X.; Waller, E.K.; Anasetti, C.; Weisdorf, D.; Miller, J.S. Human cytomegalovirus (CMV)-induced memory-like NKG2C(+) NK cells are transplantable and expand in vivo in response to recipient CMV antigen. J. Immunol. 2012, 189, 5082–5088. [Google Scholar] [CrossRef]
- Wu, Z.; Sinzger, C.; Frascaroli, G.; Reichel, J.; Bayer, C.; Wang, L.; Schirmbeck, R.; Mertens, T. Human cytomegalovirus-induced NKG2Chi CD57hi natural killer cells are effectors dependent on humoral antiviral immunity. J. Virol. 2013, 87, 7717–7725. [Google Scholar] [CrossRef] [PubMed]
- Jergovic, M.; Smithey, M.J.; Nikolich-Zugich, J. Intrinsic and extrinsic contributors to defective CD8+ T cell responses with aging. Exp. Gerontol. 2018, 105, 140–145. [Google Scholar] [CrossRef]
- Mogilenko, D.A.; Shypnov, O.; Andhey, P.S.; Arthur, L.; Swain, A.; Esaulova, E.; Brioschi, S.; Shchukina, I.; Kerndl, M.; Bambouskova, M. Comprehensive profiling of an aging immune system reveals clonal GZMK+ CD8+ T cells as conserved hallmark of inflammaging. Immunity 2021, 54, 99–115.e12. [Google Scholar] [CrossRef] [PubMed]
- Pereira, B.I.; De Maeyer, R.P.H.; Covre, L.P.; Nehar-Belaid, D.; Lanna, A.; Ward, S.; Marches, R.; Chambers, E.S.; Gomes, D.C.O.; Riddell, N.E.; et al. Sestrins induce natural killer function in senescent-like CD8+ T cells. Nat. Immunol. 2020, 21, 684–694. [Google Scholar] [CrossRef]
- Ferreira, D.P.; Silva, J.G.; Wyss, T.; Marraco, S.A.F.; Scarpellino, L.; Charmoy, M.; Maas, R.; Siddiqui, I.; Tang, L.; Joyce, J.A.; et al. Central memory CD8+ T cells derive from stem-like Tcf7hi effector cells in the absence of cytotoxic differentiation. Immunity 2020, 53, 985–1000. [Google Scholar] [CrossRef] [PubMed]
- Younes, S.-A.; Freeman, M.L.; Mudd, J.C.; Shive, C.L.; Reynaldi, A.; Panigrahi, S.; Estes, J.D.; Deleage, C.; Lucero, C.; Anderson, J.; et al. IL-15 promotes activation and expansion of CD8+ T cells in HIV-1 infection. J. Clin. Investig. 2016, 126, 2745–2756. [Google Scholar] [CrossRef] [PubMed]
- Hassouneh, F.; Goldeck, D.; Pera, A.; van Heemst, D.; Slagboom, P.E.; Pawelec, G.; Solana, R. Functional changes of T-cell subsets with age and CMV infection. Int. J. Mol. Sci. 2021, 22, 9973. [Google Scholar] [CrossRef] [PubMed]
- Almanzar, G.; Schmalzing, M.; Trippen, R.; Höfner, K.; Weißbrich, B.; Geissinger, E.; Meyer, T.; Liese, J.; Tomy, H.-P.; Prelog, M. Significant IFNγ responses of CD8+ T cells in CMV-seropositive individuals with autoimmune arthritis. J. Clin. Virol. 2016, 77, 77–84. [Google Scholar] [CrossRef]
- Scotet, E.; Peyrat, M.-A.; Saulquin, X.; Retiere, C.; Couedel, C.; Davodeau, F.; Dulphy, N.; Toubert, A.; Bignon, J.-D.; Lim, A.; et al. Frequent enrichment for CD8 T cells reactive against common herpes viruses in chronic inflammatory lesions: Towards a reassessment of the physiopathological significance of T cell clonal expansions found in autoimmune inflammatory processes. Eur. J. Immunol. 1999, 29, 973–998. [Google Scholar] [CrossRef]
- Schirmer, M.; Goldberger, C.; Würzner, R.; Duftner, C.; Pfeiffer, K.-P.; Clausen, J.; Neumayr, G.; Falkenbach, A. Circulating cytotoxic CD8+ CD28− T cells in ankylosing spondylitis. Arthritis Res. 2002, 4, 71–76. [Google Scholar] [CrossRef]
- Tsukishiro, T.; Donnenberg, A.D.; Whiteside, T.L. Rapid turnover of the CD8(+)CD28(-) T-cell subset of effector cells in the circulation of patients with head and neck cancer. Cancer Immunol. Immunother. 2003, 52, 559–607. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.R.; Chen, B.; Mudd, J.C.; Panigrahi, S.; Shive, C.L.; Sieg, S.F.; Cameron, C.M.; Zidar, D.A.; Funderburg, N.T.; Younes, S.-A.; et al. Inflammescent CX3CR1+CD57+CD8+ T cells are generated and expanded by IL-15. JCI Insight 2020, 5, e132963. [Google Scholar] [CrossRef] [PubMed]
- Cencioni, M.T.; Magliozzi, R.; Nicholas, R.; Ali, R.; Malik, O.; Reynolds, R.; Borsellino, G.; Battistini, L.; Muraro, P.A. Programmed death 1 is highly expressed on CD8+CD57+ T cells in patients with stable multiple sclerosis and inhibits their cytotoxic response to Epstein-Barr virus. Immunology 2017, 152, 660–676. [Google Scholar] [CrossRef] [PubMed]
- Gayoso, I.; Sanchez-Correa, B.; Campos, C.; Alonso, C.; Pera, A.; Casado, J.G.; Morgado, S.; TGarazona, R.; Solana, R. Immunosenescence of human natural killer cells. J. Innate Immun. 2011, 3, 337–443. [Google Scholar] [CrossRef] [PubMed]
- Long, E.O.; Kim, H.S.; Liu, D.; Peterson, M.E.; Rajagopalan, S. Controlling natural killer cell responses: Integration of signals for activation and inhibition. Annu. Rev. Immunol. 2013, 31, 227–258. [Google Scholar] [CrossRef] [PubMed]
- Narni-Mancinelli, E.; Ugolini, S.; Vivier, E. Tuning the threshold of natural killer cell responses. Curr. Opin. Immunol. 2013, 25, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Hershkovitz, O.; Rosental, B.; Rosenberg, L.A.; Navarro-Sanchez, M.E.; Jivov, S.; Zika, A.; Gershoni-Yahalom, O.; Brient-Litzler, E.; Bedouelle, H.; Ho, J.W.; et al. NKp44 receptor mediates interaction of the envelope glycoproteins from the West Nile and dengue viruses with NK cells. J. Immunol. 2009, 183, 2610–2621. [Google Scholar] [CrossRef]
- Mandelboim, O.; Lieberman, N.; Lev, M.; Paul, L.; Arnon, T.I.; Bushkin, Y.; Davis, D.M.; Strominger, J.L.; Yewdell, J.W.; Porgador, A. Recognition of haemagglutinins on virus-infected cells by NKp46 activates lysis by human NK cells. Nature 2001, 409, 1055–1060. [Google Scholar] [CrossRef]
- Jarahian, M.; Fiedler, M.; Cohnen, A.; Djandji, D.; Hämmerling, G.J.; Gati, C.; Cerwenka, S.; Turner, P.C.; Moyer, R.W.; Watzl, C.; et al. Modulation of NKp30- and NKp46-mediated natural killer cell responses by poxviral hemagglutinin. PLoS Pathog. 2011, 7, e1002195. [Google Scholar] [CrossRef]
- Chisholm, S.E.; Reyburn, H.T. Recognition of vaccinia virus-infected cells by human natural killer cells depends on natural cytotoxicity receptors. J. Virol. 2006, 80, 2225–2233. [Google Scholar] [CrossRef]
- Markham, R.B.; Goellner, J.; Pier, G.B. In vitro T cell-mediated killing of Pseudomonas aeruginosa. I. Evidence that a lymphokine mediates killing. J. Immunol. 1984, 133, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Markham, R.B.; Pier, G.B.; Goellner, J.J.; Mizel, S.B. In vitro T cell-mediated killing of Pseudomonas aeruginosa. 2. The role of macrophages and T cell subsets in T cell killing. J. Immunol. 1985, 134, 4112–4117. [Google Scholar] [CrossRef] [PubMed]
- Barrow, A.D.; Martin, C.J.; Colonna, M. The natural cytotoxicity receptors in health and disease. Front. Immunol. 2019, 10, 909. [Google Scholar] [CrossRef] [PubMed]
- Vitenshtein, A.; Charpak-Amikam, Y.; Yamin, R.; Bauman, Y.; Isaacson, B.; Stein, N.; Berhani, O.; Dassa, L.; Gamliel, M.; Gur, C.; et al. NK cell recognition of Candida glabrata through binding of NKp46 andNCR1 to fungal ligands Epa1, Epa6, and Epa7. Cell Host Microbe 2016, 20, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Kyei, S.K.; Timm-McCann, M.; Ogbomo, H.; Jones, G.J.; Shi, M.; Xiang, R.F.; Oykhman, P.; Huston, S.M.; Islam, A.; et al. The NK receptor NKp30 mediates direct fungal recognition and killing and is diminished in NK cells from HIV-infected patients. Cell Host Microbe 2013, 14, 387–397. [Google Scholar] [CrossRef] [PubMed]
- De Maria, A.; Fogli, M.; Costa, P.; Murdaca, G.; Puppo, F.; Mavilio, D.; Moretta, A.; Moretta, L. The impaired NK cell cytolytic function in viremic HIV-1 infection is associated with a reduced surface expression of natural cytotoxicity receptors (NKp46, NKp30 and NKp44). Eur. J. Immunol. 2003, 33, 2410–2418. [Google Scholar] [CrossRef] [PubMed]
- Alves, L.C.; Berger, M.B.; Koutsandreas, T.; Kirschke, N.; Lauer, C.; Spörri, R.; Chatziioannou, A.; Corazza, N.; Krebs, P. Non-apoptotic TRAIL function modulates NK cell activity during viral infection. EMBO Rep. 2020, 21, e48789. [Google Scholar] [CrossRef] [PubMed]
- Brincks, E.L.; Katewa, A.; Kucaba, T.A.; Griffith, T.S.; Legge, K.L. CD8T cells utilize TRAIL to control influenza virus infection. J. Immunol. 2008, 181, 4918–49253. [Google Scholar] [CrossRef]
- Ishikawa, E.; Nakazawa, M.; Yoshinari, M.; Minami, M. Role of tumor necrosis factor-related apoptosis-inducing ligand in immune response toinfluenza virus infection in mice. J. Virol. 2005, 79, 7658–76634. [Google Scholar] [CrossRef]
- Walczak, H. Death receptor-ligand systems in cancer, cell death, and inflammation. Cold Spring Harb. Perspect. Biol. 2013, 5, a0086985. [Google Scholar] [CrossRef]
- Zhou, X.; Jiang, W.; Liu, Z.; Liu, S.; Liang, X. Virus infection and death receptor-mediated apoptosis. Viruses 2017, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Lamhamedi-Cherradi, S.E.; Zheng, S.J.; Maguschak, K.A.; Peschon, J.; Chen, Y.H. Defective thymocyte apoptosis and accelerated autoimmune diseases in TRAIL/mice. Nat. Immunol. 2003, 4, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Song, K.; Chen, Y.; Goke, R.; Wilmen, A.; Seidel, C.; Goke, A.; Hilliard, B.; Chen, Y. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) is an inhibitor of autoimmune inflammation and cell cycle progression. J. Exp. Med. 2000, 191, 1095–1104. [Google Scholar] [CrossRef] [PubMed]
- Dotiwala, F.; Mulik, S.; Polidoro, R.B.; Ansara, J.A.; Burleigh, B.A.; Walch, M.; Gazzinelli, R.T.; Lieberman, J. Killer lymphocytes use granulysin, perforin and granzymes to kill intracellular parasites. Nat. Med. 2016, 22, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Rasi, V.; Hameed, O.A.; Matthey, P.; Bera, S.; Grandgenett, D.P.; Salentinig, S.; Walch, M.; Hoft, D.F. Improved purification of human granzyme A/B and granulysin using a mammalian expression system. Front. Immunol. 2022, 13, 830290. [Google Scholar] [CrossRef] [PubMed]
- Oykhman, P.; Timm-McCann, M.; Xiang, R.F.; Islam, A.; Li, S.S.; Stack, D.; Huston, S.M.; Ma, L.L.; Mody, C.H. Requirement and redundancy of the Src family kinases Fyn and Lyn in perforin-dependent killing of Cryptococcus neoformans by NK cells. Infect. Immun. 2013, 81, 3912–3922. [Google Scholar] [CrossRef] [PubMed]
- Braedel, S.; Radsak, M.; Einsele, H.; Latgé, J.-P.; Michan, A.; Loeffler, J.; Haddad, Z.; Grigoleit, U.; Schild, H.; Hebart, H. Aspergillus fumigatus antigens activate innate immune cells via toll-like receptors 2 and 4. Br. J. Haematol. 2004, 125, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Becker, I.; Salaiza, N.; Aguirre, M.; Delgado, J.; Carrillo-Carrasco, N.; Kobeh, L.G.; Ruiz, A.; Cervantes, R.; Torres, A.P.; Cabrera, N.; et al. Leishmania lipophosphoglycan (LPG) activates NK cells through toll-like receptor-2. Mol. Biochem. Parasitol. 2003, 130, 65–74. [Google Scholar] [CrossRef]
- Stevenson, M.M.; Tam, M.F.; Wolf, S.F.; Sher, A. IL-12-induced protection against blood-stage Plasmodium chabaudi AS requires IFN-gamma and TNF-alpha and occurs via a nitric oxide-dependent mechanism. J. Immunol. 1995, 155, 2545–2556. [Google Scholar] [CrossRef]
- de Jong, M.F.; Alto, N.M. Cooperative immune suppression by Escherichia coli and Shigella effector proteins. Infect. Immun. 2018, 86, e00560-17. [Google Scholar] [CrossRef]
- Paczosa, M.K.; Meczas, J. Klebsiella pneumoniae: Going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef]
- Rosbjerg, A.; Genster, N.; Pilely, K.; Garred, P. Evasion mechanisms used by pathogens to escape the lectin complement pathway. Front. Microbiol. 2017, 8, 868. [Google Scholar] [CrossRef]
- Chan, C.J.; Smyth, M.J.; Martinet, L. Molecular mechanisms of natural killer cell activation in response to cellular stress. Cell Death Diff. 2014, 21, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.; Stegmann, K.A.; Riley, E.M. Activation of natural killer cells during microbial infections. Front. Immunol. 2011, 2, 88. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.; Vidal, S.M. Mechanisms of natural killer evasion through viral adaptation. Ann. Rev. Immunol. 2020, 38, 511–539. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.-T.; Carisey, A.F.; Orange, J.S. Measurement of lytic granule convergence after formation of an NK cell immunological synapse. Methods Mol. Biol. 2017, 1584, 497–515. [Google Scholar] [CrossRef] [PubMed]
- Topham, N.J.; Hewitt, E.W. Natural killer cell cytotoxicity: How do they pull the trigger? Immunology 2009, 128, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Volpe, E.; Sambucci, M.; Battistini, L.; Borsellino, G. Fas-Fas ligand: Checkpoint of T cell functions in multiple sclerosis. Front. Immunol. 2016, 7, 382. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, E.E.; Huse, M. How cytotoxic T cells release their dying targets. Nat. Immunol. 2023, 24, 1413–1414. [Google Scholar]
- Sanchez, E.E.; Tello-Lafoz, M.; Guo, A.J.; de Jesus, M.; Elbanna, Y.A.; Winer, B.Y.; Budhu, S.; Chan, E.; Rosiek, E.; Kondo, T.; et al. Apoptotic contraction drives target cell release by cytotoxic T cells. Nat. Immunol. 2023, 24, 1434–1442. [Google Scholar] [CrossRef]
- Leichner, T.; Kambayashi, T. White blood cells and lymphoid tissue. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Balint, S.; Muller, S.; Fischer, R.; Kessler, B.M.; Harkiolaki, M.; Valitutti, S.; Dustin, M.L. Supramolecular attack particles are autonomous killing entities released from cytotoxic T cells. Science 2020, 368, 897–901. [Google Scholar] [CrossRef] [PubMed]
- Dhatchinamoorthy, K.; Colbert, J.D.; Rock, K.L. Cancer immune evasion through loss of MHC Class I antigen presentation. Front. Immunol. 2021, 12, 636568. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Watzl, C. Serial killing of tumor cells by human natural killer cells—Enhancement by therapeutic antibodies. PLoS ONE 2007, 2, e326. [Google Scholar] [CrossRef] [PubMed]
- Prager, I.; Liesche, C.; van Ooijen, H.; Urlaub, D.; Verron, Q.; Sandström, N.; Fasbender, F.; Claus, M.; Eils, R.; Beaudouin, J.; et al. NK cells switch from granzyme B to death receptor-mediated cytotoxicity during serial killing. J. Exp. Med. 2019, 216, 2113–2127. [Google Scholar] [CrossRef]
- Vojdani, A. A potential link between environmental triggers and autoimmunity. Autoimmune Dis. 2014, 2014, 437231. [Google Scholar] [CrossRef]
- Robertson, M.J.; Schacterle, R.S.; Mackin, G.A.; Wilson, S.N.; Bloomingdale, K.L.; Ritz, J.; Komaroff, A.L. Lymphocyte subset differences in patients with chronic fatigue syndrome, multiple sclerosis and major depression. Clin. Exp. Immunol. 2005, 141, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Erdag, G.; Schaefer, J.T.; Smolkin, M.E.; Deacon, D.H.; Shea, S.M.; Dengel, L.T.; Patterson, J.W.; Slingluff, C.L., Jr. Immunotype and immunohistologic characteristics of tumor-inflitrating immune cells are associated with clinical outcome in metastatic melanoma. Cancer Res. 2012, 72, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Tan, S.; Tao, L.; Gong, J.; Chang, Y.; Wang, Y.; Fan, P.; He, D.; Ruan, Y.; Qiu, W. Intestinal barrier breakdown and mucosal microbiota disturbance in neuromyelitis optical spectrum disorders. Front. Immunol. 2020, 11, 2101. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of immune response in patients with Coronavirus 19 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Sakuragi, T.; Yamada, H.; Haraguchi, A.; Kai, K.; Fukushi, J.I.; Ikemura, S.; Akasaki, Y.; Fujiwara, T.; Tsushima, H.; Tsutsui, T.; et al. Autoreactivity of peripheral helper T-cells in the joints of rheumatoid arthritis. J. Immunol. 2021, 206, 2045–2051. [Google Scholar] [CrossRef]
- Malkiel, S.; Jeganathan, V.; Wolfson, S.; Orduño, N.M.; Mafrasco, E.; Aranow, C.; Mackay, M.; Gregersen, P.K.; Diamond, B. Checkpoints for autoreactive B cells in the peripheral blood of lupus patients assessed by flow cytometry. Arth. Rheumatol. 2016, 68, 2210–2220. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Blish, C.A. Defining the role of natural killer cells in COVID-19. Nat. Immunol. 2023, 24, 1628–1638. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.S.; Ryan, P.L.; Bergmeier, L.A.; Fortune, F. Circulating NK cells and their subsets in Behçet’s disease. Clin. Exp. Immunol. 2017, 188, 311–322. [Google Scholar] [CrossRef]
- Atlasy, N.; Bujko, A.; Baekkevold, E.S.; Brazda, P.; Janssen-Megens, E.; Lundin, K.E.A.; Jahnsen, J.; Jahnsen, F.L.; Stunnenberg, H.G. Single cell transciptomic analysis of the immune cell compartment in the human small intestine and in Celiac disease. Nat. Commun. 2022, 13, 4920. [Google Scholar] [CrossRef] [PubMed]
- Marafini, I.; Monteleone, I.; Di Fusco, D.; Sedda, S.; Cupi, M.L.; Fina, D.; Paoluzi, A.O.; Pallone, F.; Monteleone, G. Celiac disease-related inflammation is marked by reduction of Nkp44/Nkp46-double positive natural killer cells. PLoS ONE 2016, 11, e0155103. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Intraepithelial lymphocyte normal cut-off level in celiac disease: The debate continues. Int. J. Celiac Dis. 2016, 4, 4–6. [Google Scholar] [CrossRef]
- Heuser, G.; Vojdani, A. Enhancement of natural killer cell activity and T and B cell function by buffered vitamin C in patients exposed to toxic chemicals: The role of protein kinase-C. Immunopharmacol. Immunotoxicol. 1997, 19, 291–312. [Google Scholar] [CrossRef]
- Vojdani, A.; Ghoneum, M.; Brautbar, N. Immune alteration associated with exposure to toxic chemicals. Toxicol. Ind. Health 1992, 8, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Arowolo, O.; Pobezinsky, L.; Suvorov, A. Chemical exposures affect innate immune response to SARS-CoV-2. Int. J. Mol. Sci. 2021, 22, 12474. [Google Scholar] [CrossRef]
- Matos, L.C.; Batista, P.; Monteiro, N.; Ribeiro, J.; Cipriano, M.A.; Henriques, P.; Girão, F.; Carvalho, A. Lymphocyte subsets in alcoholic liver disease. World J. Hepatol. 2013, 5, 46–55. [Google Scholar] [CrossRef]
- Medina-Avila, Z.; Martínez-Castillo, M.; Santana-Vargas, D.; Bejar, Y.; de la Tijera, F.H.; Pérez-Hernández, J.L.; Kershenobich, D.; Gutiérrez-Reyes, G. Lymphocyte profile on patients with chronic and acute alcohol consumption. Ann. Hepatol. 2022, 27, 100637. [Google Scholar] [CrossRef]
- Galvin, D.A.; McCrory, C. The role of T-lymphocytes in neuropathic pain initiation, development of chronicity and treatment. Brain Behav. Immun. Health 2021, 18, 100371. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.A.; Fiore, N.T.; van Vreden, C.; Bailey, D.; Santarelli, D.M.; McGuire, H.N.; de St Groth, B.F.; Austin, P.J. Expansion and activation of distinct central memory T lymphocyte subsets in complex regional pain syndrome. J. Neuroinflamm. 2019, 16, 63. [Google Scholar] [CrossRef] [PubMed]
- Pontrelli, P.; Rascio, F.; Castellano, G.; Grandaliano, G.; Gesualdo, L.; Stallone, G. The role of natural killer cells in the immune response in kidney transplantation. Front. Immunol. 2020, 11, 1454. [Google Scholar] [CrossRef] [PubMed]
- Oshida, Y.; Yamanouchi, K.; Hayamizu, S.; Sato, Y. Effect of acute physical exercise on lymphocyte subpopulations in trained and untrained subjects. Int. J. Sports Med. 1988, 9, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Maydych, V.; Claus, M.; Dychus, N.; Ebel, M.; Damaschke, J.; Diestel, S.; Wolf, O.T.; Kleinsorge, T.; Watzl, C. Impact of chronic and acute academic stress on lymphocyte subsets and monocyte function. PLoS ONE 2017, 12, e0188108. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z. Transient stress lymphocytosis uncovers the underlying B-cell lymphoproliferative disorder. Case Rep. Clin. Pathol. 2014, 1, 1. [Google Scholar] [CrossRef]
- Rudak, P.T.; Choi, J.; Parkins, K.M.; Summers, K.L.; Jackson, D.N.; Foster, P.J.; Skaro, A.I.; Leslie, K.; McAlister, V.C.; Kuchroo, V.K.; et al. Chronic stress physically spares but functionally impairs innate-like invariant T cells. Cell Rep. 2021, 35, 108979. [Google Scholar] [CrossRef]
- Snijders, G.; Brouwer, R.; Kemner, S.; Bootsman, F.; Drexhage, H.A.; Hillegers, M.H.J. Genetic and environmental influences on circulating NK and T cells and their relation to bipolar disorder. Int. J. Bipolar Disord. 2019, 7, 4. [Google Scholar] [CrossRef]
- Pietruczuk, K.; Lisowska, K.A.; Grabowski, K.; Landowski, J.; Cubala, W.J.; Witkowski, J.M. Peripheral blood lymphocyte subpopulations in patients with bipolar disorder type II. Sci. Rep. 2019, 9, 5869. [Google Scholar] [CrossRef]
- Sun, C.; Zhao, Z.; Yu, W.; Mo, M.; Song, C.; Si, Y.; Liu, Y. Abnormal subpopulations of peripheral blood lymphocytes are involved in Parkinson’s disease. Ann. Transl. Med. 2019, 7, 637. [Google Scholar] [CrossRef]
- Reagin, K.L.; Funk, K.E. CD8+ T cells pump the brakes on Alzheimer’s disease. Nat. Immunol. 2023, 24, 1597–1598. [Google Scholar] [CrossRef]
- Su, W.; Saravia, J.; Risch, I.; Rankin, S.; Guy, C.; Chapman, N.M.; Shi, H.; Sun, Y.; Kc, A.; Li, W.; et al. CXCR6 orchestrates brain CD8+ T cell residency and limits mouse Alzheimer’s disease pathology. Nat. Immunol. 2023, 24, 1735–1747. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Firulyova, M.; Manis, M.; Herz, J.; Smirnov, I.; Aladyeva, E.; Wang, C.; Bao, X.; Finn, M.B.; Hu, H.; et al. Microglia-mediated T cell infiltration drives neurodegeneration in tauopathy. Nature 2023, 615, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Krechetova, L.V.; Vanko, L.V.; Vtorushina, V.V.; Nikolaeva, M.A.; Inviyaeva, E.V.; Tetruashvili, N.K. Lymphocyte activation in the development of immune tolerance in women with recurrent pregnancy loss. Biochemistry 2020, 85, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Cheung, R.R.; Grinstein, S.; Gelfland, E.W. Volume regulation by human lymphocytes. Identification of differences between the two major lymphocyte subpopulations. J. Clin. Investig. 1982, 70, 632–638. [Google Scholar] [CrossRef]
- Cordone, I.; Masi, S.; Giannarelli, D.; Pasquale, A.; Conti, L.; Telera, S.; Pace, A.; Papa, E.; Marino, M.; de Fabrtiis, P.; et al. Major differences in lymphocyte subpopulations between cerebrospinal fluid and peripheral blood in non-Hodgkin lymphoma without leptomeningeal involvement: Flow cytometry evidence of a cerebral lymphatic system. Front. Oncol. 2021, 11, 85786. [Google Scholar] [CrossRef] [PubMed]
- Vojdani, A.; Mumper, E.; Granpeesheh, D.; Mielke, L.; Traver, D.; Bock, K.; Hirani, K.; Neubrander, J.; Woeller, K.N.; O’Hara, N.; et al. Low natural killer cell cytotoxic activity in autism: The role of glutathione, IL-2 and IL-15. J. Neuroimmunol. 2008, 205, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Enstrom, A.M.; Lit, L.; Onore, C.E.; Gregg, J.P.; Hansen, R.L.; Pessah, I.N.; Hertz-Picciotto, I.; Van de Water, J.A.; Sharp, F.R.; Ashwood, P. Altered gene expression and function of peripheral blood natural killer cells in children with autism. Brain Behav. Immun. 2009, 23, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Saresella, M.; Marventano, I.; Piancone, F.; Bolognesi, E.; Hernis, A.; Zanzottera, M.; La Rosa, F.; Agliardi, C.; Giraldo, S.; Chiappedi, M.; et al. Alterations of natural killer cells activatory molecules phenotype and function in mothers of ASD children: A pilot study. Front. Immunol. 2023, 14, 1190925. [Google Scholar] [CrossRef]
- Childs, R.W.; Berg, M. Bringing natural killer cells to the clinic: Ex vivo manipulation. Henatology Am. Soc. Hematol. Educ. Program 2013, 2013, 234–246. [Google Scholar] [CrossRef]
- Carlsten, M.; Childs, R.W. Genetic manipulation of NK cells for cancer immunotherapy: Techniques and clinical implications. Front. Immunol. 2015, 6, 266. [Google Scholar] [CrossRef] [PubMed]
- Vishwasrao, P.; Hui, S.K.; Smith, D.L.; Khairnar, V. Role of NK cells in cancer and immunotherapy. Onco 2021, 1, 158–175. [Google Scholar] [CrossRef]
- Maia, A.; Tarannum, M.; Romee, R. Genetic manipulation approaches to enhance the clinical application of NK cell-based immunotherapy. Stem Cells Transl. Med. 2023, szad087. [Google Scholar] [CrossRef] [PubMed]
- Simoni, Y.; Diana, J.; Ghazarian, L.; Beaudoin, L.; Lehuen, A. Therapeutic manipulation of natural killer (NK) T cells in autoimmunity: Are we close to reality? Clin. Exp. Immunol. 2012, 171, 8–19. [Google Scholar] [CrossRef]
- Li, Z.; Ma, R.; Tang, H.; Guo, J.; Shah, Z.; Zhang, J.; Liu, N.; Cao, S.; Marcucci, G.; Artis, D.; et al. Therapeutic application of human type 2 innate lymphoid cells via induction of granzyme B-mediated tumor cell death. Cell 2024, 187, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Pesce, S.; Moretta, L.; Moretta, A.; Marcenaro, E. Human NK cell subsets redistribution in pathological conditions: A role for CCR7 receptor. Front. Immunol. 2016, 7, 414. [Google Scholar] [CrossRef]
- Pesce, S.; Trabanelli, S.; Di Vito, C.; Greppi, M.; Obino, V.; Guolo, F.; Minetto, P.; Bozzo, M.; Calvi, M.; Zaghi, E.; et al. Cancer immunotherapy by blocking immune checkpoints on innate lymphocytes. Cancers 2020, 12, 3504. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunophenotype | Cytotoxic Activity | Clinical Significance | References |
|---|---|---|---|
| CD56bright(+)CD16− NK Cell | Weakly cytotoxic Produces limited amounts of cytotoxic molecules | Immunoregulation through production of many cytokines and chemokines | [145,146,147,148,149,150,151,152] |
| CD56−CD16bright(+) NK Cell | Naturally strongly cytotoxic Produces significant amounts of cytotoxic molecules | Protection against microorganisms and cancerous cells | [153,154,155,156,157,158,159,160,161] |
| CD3−CD57bright(+) NK Cell | High cytotoxic potential as a result of producing various cytotoxic molecules | Long-lived cells Immunoprotection against pathogens, especially during aging Deletion of autoreactive T cells Prevention of some autoimmune diseases | [153,154,155,156,157,158,159,160,161,162,163] |
| CD57bright(+) CD16bright(+) NK Cell | Highly cytotoxic Produces significant amounts ofcytotoxic molecules | The most potent cells for combating acute and chronic infections | [69,71,162,163,164,165,166,167,168] |
| CD8+ Cytotoxic T Cell | After activation by APC, becomes highly cytotoxic Produces different cytotoxic molecules | Immune response to bacterial/viral infections and cancer | [113,169,170,171,172,173,174,175,176,177,178] |
| CD8+CD57+ T Cell | Highly cytotoxic Produces significant amounts of cytotoxic molecules | Mediators of adaptive immunity Associated with chronic immune activation Age-related changes in the immune system status | [59,60,61,81,114,179,180,181] |
| CD3+CD56+CD16+ T Cell | Moderately cytotoxic Plays a significant role in immunoregulation | Serves as bridge between innate and adaptive immunity Immune protection against microbial pathogens and cancer Control of autoimmune diseases | [129,136,137,138,139,141,142] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vojdani, A.; Koksoy, S.; Vojdani, E.; Engelman, M.; Benzvi, C.; Lerner, A. Natural Killer Cells and Cytotoxic T Cells: Complementary Partners against Microorganisms and Cancer. Microorganisms 2024, 12, 230. https://doi.org/10.3390/microorganisms12010230
Vojdani A, Koksoy S, Vojdani E, Engelman M, Benzvi C, Lerner A. Natural Killer Cells and Cytotoxic T Cells: Complementary Partners against Microorganisms and Cancer. Microorganisms. 2024; 12(1):230. https://doi.org/10.3390/microorganisms12010230
Chicago/Turabian StyleVojdani, Aristo, Sadi Koksoy, Elroy Vojdani, Mark Engelman, Carina Benzvi, and Aaron Lerner. 2024. "Natural Killer Cells and Cytotoxic T Cells: Complementary Partners against Microorganisms and Cancer" Microorganisms 12, no. 1: 230. https://doi.org/10.3390/microorganisms12010230