
Engineering a Lactobacillus Lysine Riboswitch to Dynamically Control Metabolic Pathways for Lysine Production in Corynebacterium glutamicum
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Media
2.2. DNA Manipulation
2.3. Identification of Lysine Riboswitches in Lb. plantarum
2.4. Construction of Plasmids
2.5. Fluorescence Measurement
2.6. Screening Engineered Lb. plantarum Lysine Riboswitches
2.7. Construction of C. glutamicum Lysine Riboswitch-Controlled Mutants
2.8. Enzyme Activity Assay
2.9. Fermentation and Analytical Methods
2.10. Statistics Analysis
3. Results
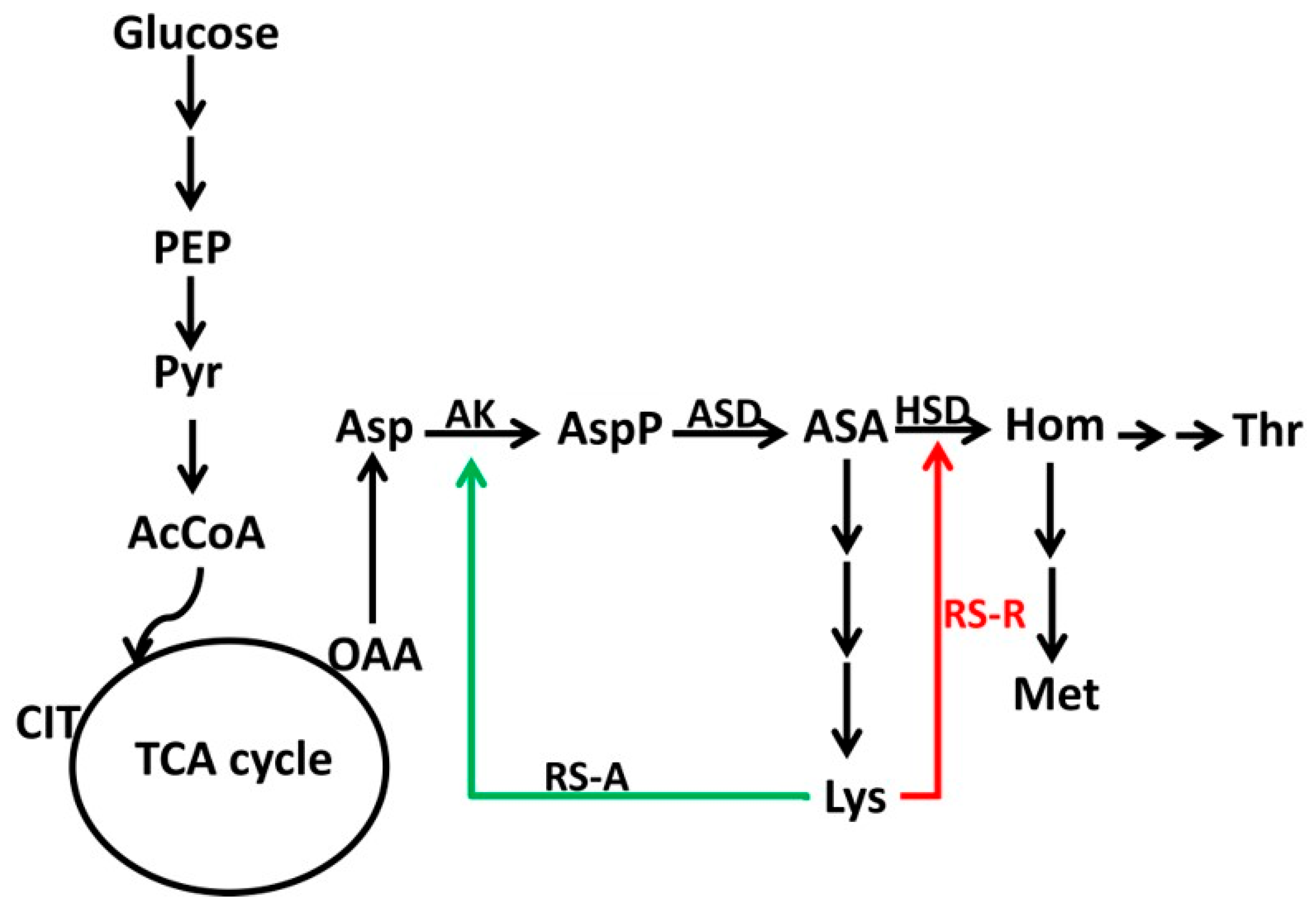
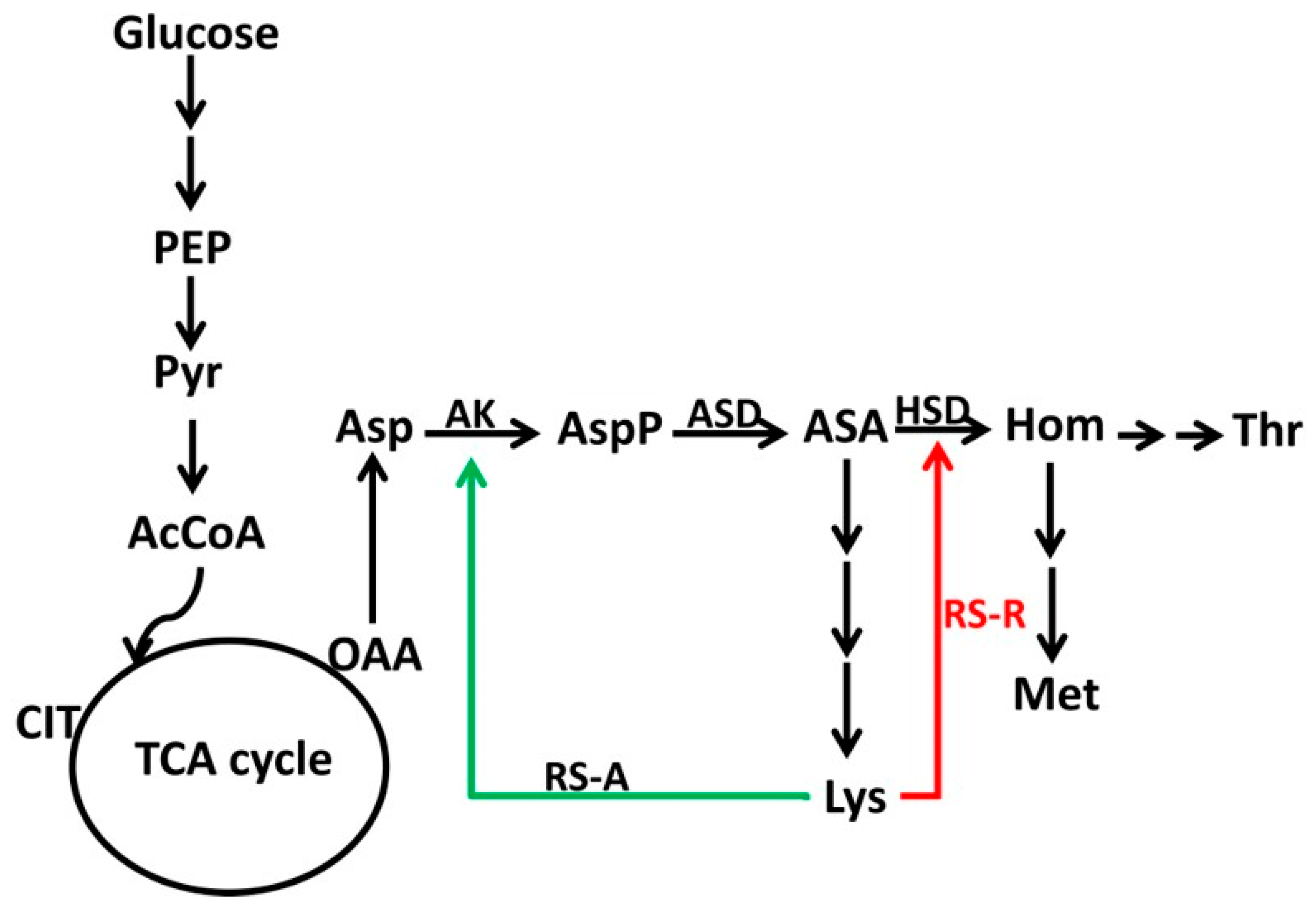
3.1. A Putative Lactobacillus Lysine Riboswitch Controls a Lysine Transport Protein
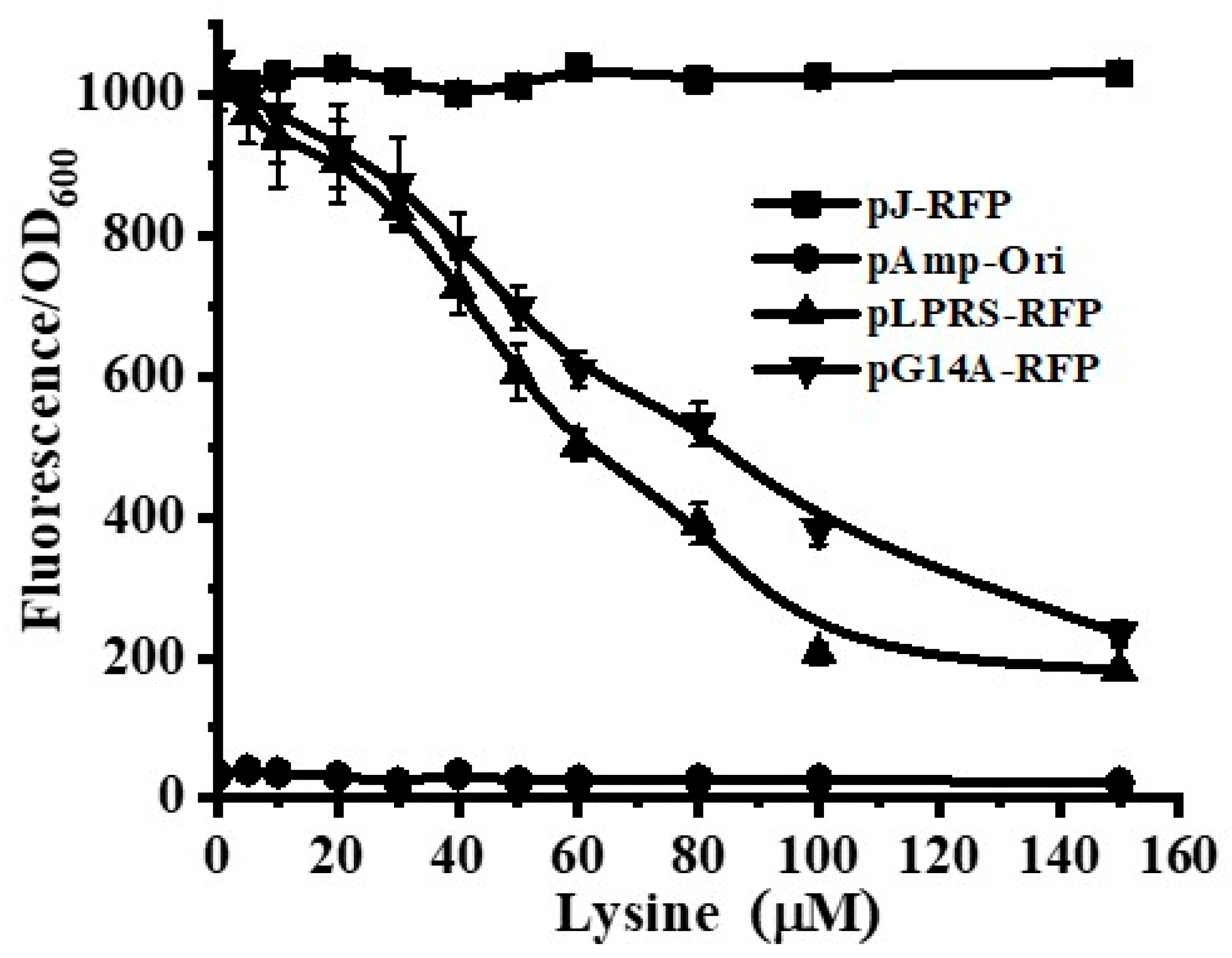
3.2. Construction of Lactobacillus Lysine Riboswitch-RFP Sensor
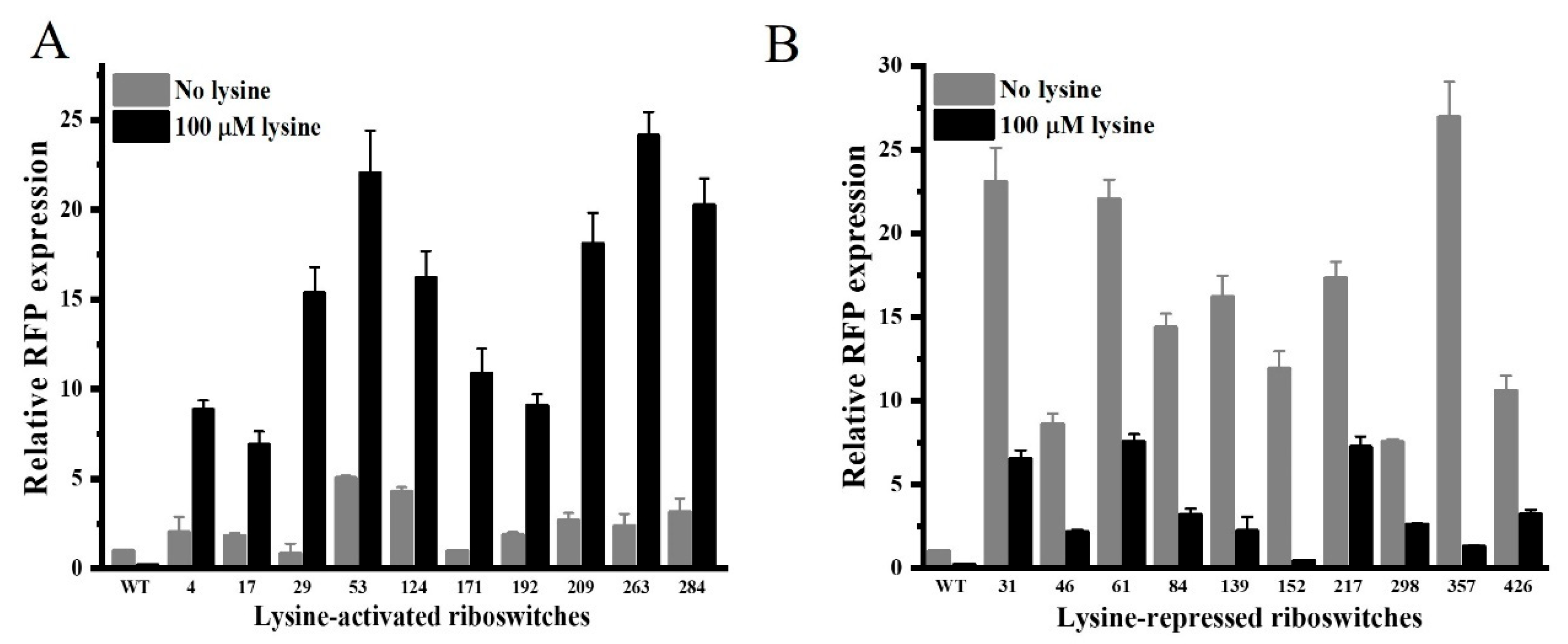
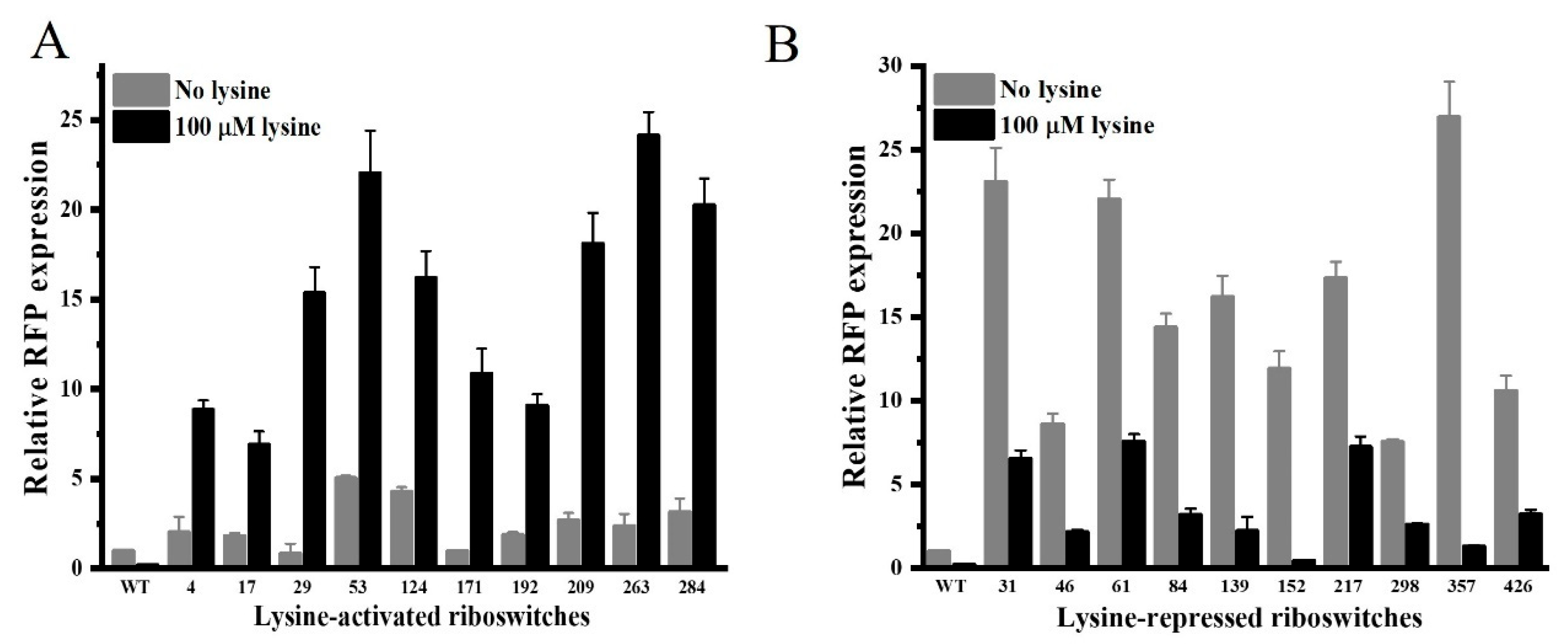
3.3. Selection of Engineered Lysine Riboswitches with Optimized Properties
3.4. Construction of a Riboswitch-Based Mutant in C. glutamicum
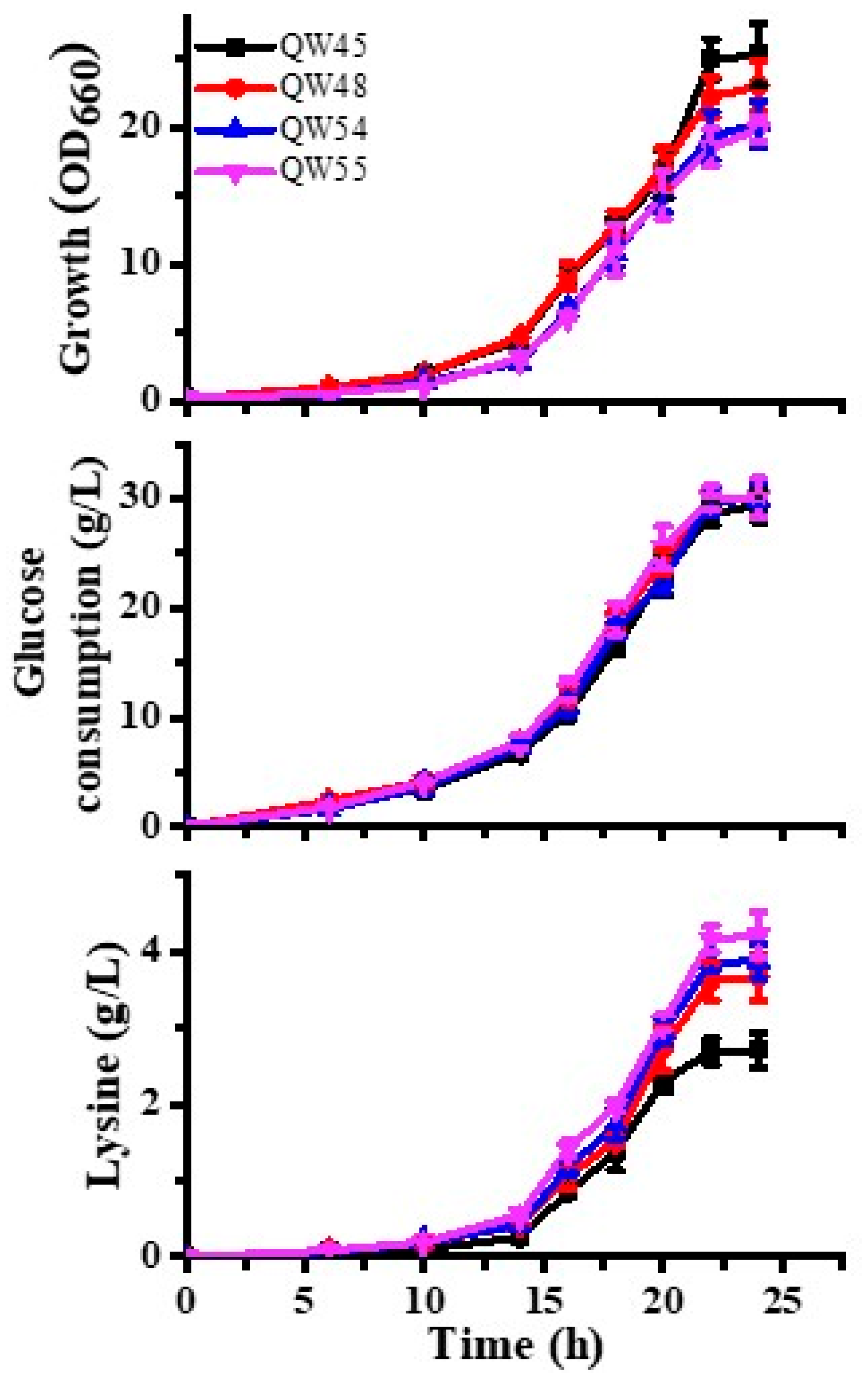
3.5. Lysine Production Enhanced by Riboswitch-Controlled Mutants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bédard, A.S.V.; Hien, E.D.M.; Lafontaine, D.A. Riboswitch regulation mechanisms: RNA, metabolites and regulatory proteins. BBA Gene Regul. Mech. 2020, 1863, 194501. [Google Scholar] [CrossRef]
- Bastet, L.; Dubé, A.; Massé, E.; Lafontaine, D.A. New insights into riboswitch regulation mechanisms. Mol. Microbiol. 2011, 80, 1148–1154. [Google Scholar] [CrossRef] [PubMed]
- Serganov, A.; Patel, D.J. Amino acid recognition and gene regulation by riboswitches. BBA Gene Regul. Mech. 2009, 1789, 592–611. [Google Scholar] [CrossRef] [PubMed]
- Suddala, K.C.; Cabello-Villegas, J.; Michnicka, M.; Marshall, C.; Nikonowicz, E.P.; Walter, N.G. Hierarchical mechanism of amino acid sensing by the T-box riboswitch. Nat. Commun. 2018, 9, 1896. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.G.; Milito, A.; Yang, J.S.; Jang, S.; Jung, G.Y. Riboswitch-guided chalcone synthase engineering and metabolic flux optimization for enhanced production of flavonoids. Metab. Eng. 2023, 75, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Page, K.; Liu, J. Engineering riboswitch-based whole cell biosensors in Escherichia coli. FASEB J. 2018, 32, 143–152. [Google Scholar] [CrossRef]
- Zhou, L.B.; Ren, J.; Li, Z.D.; Nie, J.L.; Wang, C.; Zeng, A.P. Characterization and engineering of a Clostridium glycine riboswitch and its use to control a novel metabolic pathway for 5-aminolevulinic acid production in Escherichia coli. ACS Synth. Biol. 2019, 8, 2327–2335. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, H.; Dworkin, M.; Falkow, S.; Rosenberg, E.; Schleifer, K.; Stackebrandt, E. Amino acid production. Prokaryotes 2006, 1, 756–765. [Google Scholar]
- Liu, J.; Xu, J.Z.; Rao, Z.M.; Zhang, W.G. Industrial production of L-lysine in Corynebacterium glutamicum: Progress and prospects. Microbiol. Res. 2022, 262, 127101. [Google Scholar] [CrossRef]
- Wittmann, C.; Becker, J. The L-lysine story: From metabolic pathways to industrial production. In Amino Acid Biosynthesis~Pathways, Regulation and Metabolic Engineering; Springer: Berlin/Heidelberg, Germany, 2007; pp. 39–70. [Google Scholar]
- Bommareddy, R.R.; Chen, Z.; Rappert, S.; Zeng, A.P. A de novo NADPH generation pathway for improving lysine production of Corynebacterium glutamicum by rational design of the coenzyme specificity of glyceraldehyde 3-phosphate dehydrogenase. Metab. Eng. 2014, 25, 30–37. [Google Scholar] [CrossRef]
- Hoffmann, S.L.; Kohlstedt, M.; Jungmann, L.; Hutter, M.; Poblete-Castro, I.; Becker, J.; Wittmann, C. Cascaded valorization of brown seaweed to produce L-lysine and value-added products using streamlined by systems metabolic engineering. Metab. Eng. 2021, 67, 293–307. [Google Scholar] [CrossRef]
- Wu, W.J.; Zhang, Y.; Liu, D.H.; Chen, Z. Efficient mining of natural NADH-utilizing dehydrogenases enables systematic cofactor engineering of lysine synthesis pathway of Corynebacterium glutamicum. Metab. Eng. 2019, 52, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Shin, H.; Lee, S.M.; Um, Y.; Woo, H.M. RNA-guided single/double gene repressions in using an efficient CRISPR interference and its application to industrial strain. Microb. Cell Fact. 2018, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.H.; Chen, J.Z.; Wang, Y.; Li, M.Y.; Rao, D.M.; Guo, Y.M.; Chen, N.; Zheng, P.; Sun, J.B.; Ma, Y.H. Metabolic engineering of by synthetic small regulatory RNAs. J. Ind. Microbiol. Biot. 2019, 46, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer-Sancar, K.; Mentz, A.; Rückert, C.; Kalinowski, J. Comprehensive analysis of the transcriptome using an improved RNAseq technique. BMC Genom. 2013, 14, 888. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, N.; Tanaka, Y.; Inui, M.; Yukawa, H. The physiological role of riboflavin transporter and involvement of FMN-riboswitch in its gene expression in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2014, 98, 4159–4168. [Google Scholar] [CrossRef] [PubMed]
- Reich, S.J.; Goldbeck, O.; Lkhaasuren, T.; Weixler, D.; Weiss, T.; Eikmanns, B.J. C-di-AMP is a second messenger in that regulates expression of a cell wall-related peptidase via a riboswitch. Microorganisms 2023, 11, 296. [Google Scholar] [CrossRef]
- Caron, M.P.; Bastet, L.; Lussier, A.; Simoneau-Roy, M.; Massé, E.; Lafontaine, D.A. Dual-acting riboswitch control of translation initiation and mRNA decay. Proc. Natl. Acad. Sci. USA 2012, 109, E3444–E3453. [Google Scholar] [CrossRef]
- Takemoto, N.; Tanaka, Y.; Inui, M. Rho and RNase play a central role in FMN riboswitch regulation in. Nucleic Acids Res. 2015, 43, 520–529. [Google Scholar] [CrossRef]
- Ikeda, M.; Ohnishi, J.; Hayashi, M.; Mitsuhashi, S. A genome-based approach to create a minimally mutated, strain for efficient L-lysine production. J. Ind. Microbiol. Biot. 2006, 33, 610–615. [Google Scholar] [CrossRef]
- Chen, Z.; Rappert, S.; Zeng, A.P. Rational design of allosteric regulation of homoserine dehydrogenase by a nonnatural inhibitor L-lysine. ACS Synth. Biol. 2015, 4, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.B.; Zeng, A.P. Exploring lysine riboswitch for metabolic flux control and improvement of L-lysine synthesis in Corynebacterium glutamicum. ACS Synth. Biol. 2015, 4, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.B.; Zeng, A.P. Engineering a lysine-ON riboswitch for metabolic control of lysine production in Corynebacterium glutamicum. ACS Synth. Biol. 2015, 4, 1335–1340. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Fowler, C.C.; Brown, E.D.; Li, Y.F. Using a riboswitch sensor to examine coenzyme B12 metabolism and transport in Escherichia coli. Chem. Biol. 2010, 17, 756–765. [Google Scholar] [CrossRef]
- Schafer, A.; Tauch, A.; Jager, W.; Kalinowski, J.; Thierbach, G.; Puhler, A. Small mobilizable multipurpose cloning vectors derived from the Escherichia Coli plasmids pk18 and pk19 selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Rodionov, D.A.; Vitreschak, A.G.; Mironov, A.A.; Gelfand, M.S. Regulation of lysine biosynthesis and transport genes in bacteria: Yet another RNA riboswitch? Nucleic Acids Res. 2003, 31, 6748–6757. [Google Scholar] [CrossRef] [PubMed]
- Serganov, A.; Huang, L.L.; Patel, D.J. Structural insights into amino acid binding and gene control by a lysine riboswitch. Nature 2008, 455, 1263-U76. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Sengupta, S. Riboswitch Scanner: An efficient pHMM-based web-server to detect riboswitches in genomic sequences. Bioinformatics 2016, 32, 776–778. [Google Scholar] [CrossRef]
- Gruber, A.R.; Lorenz, R.; Bernhart, S.H.; Neuböock, R.; Hofacker, I.L. The Vienna RNA Websuite. Nucleic Acids Res. 2008, 36, W70–W74. [Google Scholar] [CrossRef]
- Johnson, P.Z.; Simon, A.E. RNAcanvas: Interactive drawing and exploration of nucleic acid structures. Nucleic Acids Res. 2023, 51, W501–W508. [Google Scholar] [CrossRef]
- Muranaka, N.; Yokobayashi, Y. Posttranscriptional signal integration of engineered riboswitches yields band-pass output. Angew. Chem. Int. Ed. 2010, 49, 4653–4655. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Bommareddy, R.R.; Frank, D.; Rappert, S.; Zeng, A.P. Deregulation of feedback inhibition of phosphoenolpyruvate carboxylase for improved lysine production in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2014, 80, 1388–1393. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.Y.; Zhao, Y.; Zhao, J.X.; Wang, X.Y. Characterization of aspartate kinase and homoserine dehydrogenase from Corynebacterium glutamicum IWJ001 and systematic investigation of L-isoleucine biosynthesis. J. Ind. Microbiol. Biot. 2016, 43, 873–885. [Google Scholar] [CrossRef]
- Ziert, C. Metabolic engineering of Corynebacterium glutamicum for the production of L-aspartate and its derivatives β-Alanine and ectoine. BMC 2014, 12, 110. [Google Scholar] [CrossRef]
- Li, J.; Ge, Y.; Zadeh, M.; Curtiss, R.; Mohamadzadeh, M. Regulating vitamin B12 biosynthesis via the riboswitch in strain UF1. Proc. Natl. Acad. Sci. USA 2020, 117, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Dwidar, M.; Seike, Y.; Kobori, S.; Whitaker, C.; Matsuura, T.; Yokobayashi, Y. Programmable artificial cells using histamine-responsive synthetic riboswitch. J. Am. Chem. Soc. 2019, 141, 11103–11114. [Google Scholar] [CrossRef]
- Jang, S.; Jang, S.; Im, D.K.; Kang, T.J.; Oh, M.K.; Jung, G.Y. Artificial caprolactam-specific riboswitch as an intracellular metabolite sensor. ACS Synth. Biol. 2019, 8, 1276–1283. [Google Scholar] [CrossRef]
- Yang, J.; Seo, S.W.; Jang, S.; Shin, S.I.; Lim, C.H.; Roh, T.Y.; Jung, G.Y. Synthetic RNA devices to expedite the evolution of metabolite-producing microbes. Nat. Commun. 2013, 4, 1413. [Google Scholar] [CrossRef]
- Zhang, Y.; Shang, X.L.; Lai, S.J.; Zhang, G.Q.; Liang, Y.; Wen, T.Y. Development and application of an arabinose-inducible expression system by facilitating inducer uptake in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2012, 78, 5831–5838. [Google Scholar] [CrossRef]
- Mutanda, I.; Li, J.H.; Xu, F.L.; Wang, Y. Recent advances in metabolic engineering, protein engineering, and transcriptome-guided insights toward synthetic production of taxol. Front. Bioeng. Biotechnol. 2021, 9, 632269. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Lysine Yield a | Final Growth (OD660) | Specific Activity of Enzyme (μmol min−1 mg−1) | |
|---|---|---|---|---|
| Aspartate Kinase (AK III) | Homoserine Dehydrogenase (HSD) | |||
| QW45 | 0.113 ± 0.009 | 25.32 ± 2.29 | 0.837 ± 0.064 | 0.915 ± 0.023 |
| QW48 | 0.15 ± 0.012 | 23.07 ± 1.82 | 0.973 ± 0.004 | / |
| QW54 | 0.159 ± 0.008 | 20.24 ± 1.66 | / | 0.782 ± 0.036 |
| QW55 | 0.173 ± 0.011 | 19.86 ± 0.85 | 0.919 ± 0.015 | 0.803 ± 0.022 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.; Geng, F.; Shen, J.; Zhu, P.; Lu, Z.; Zhou, L.; Lu, F. Engineering a Lactobacillus Lysine Riboswitch to Dynamically Control Metabolic Pathways for Lysine Production in Corynebacterium glutamicum. Microorganisms 2024, 12, 606. https://doi.org/10.3390/microorganisms12030606
Jiang Q, Geng F, Shen J, Zhu P, Lu Z, Zhou L, Lu F. Engineering a Lactobacillus Lysine Riboswitch to Dynamically Control Metabolic Pathways for Lysine Production in Corynebacterium glutamicum. Microorganisms. 2024; 12(3):606. https://doi.org/10.3390/microorganisms12030606
Chicago/Turabian StyleJiang, Qingwei, Feng Geng, Juan Shen, Ping Zhu, Zhaoxin Lu, Libang Zhou, and Fengxia Lu. 2024. "Engineering a Lactobacillus Lysine Riboswitch to Dynamically Control Metabolic Pathways for Lysine Production in Corynebacterium glutamicum" Microorganisms 12, no. 3: 606. https://doi.org/10.3390/microorganisms12030606