
Combined TCBS and CHROMagar Analyses Allow for Basic Identification of Vibrio vulnificus within a 48 h Incubation Period in the Coastal Baltic Sea


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
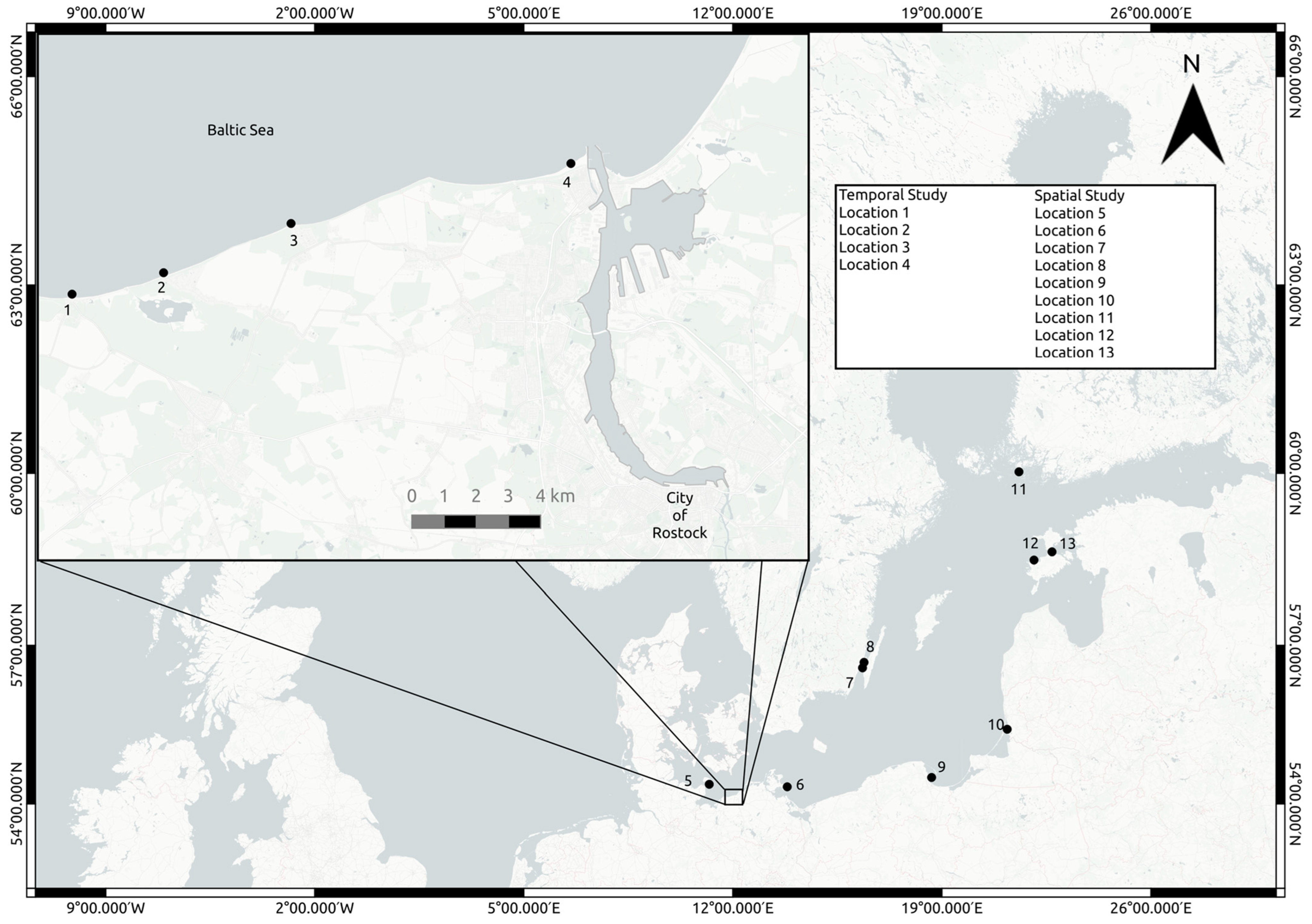
2.1. Sampling Areas
2.2. Environmental Sampling
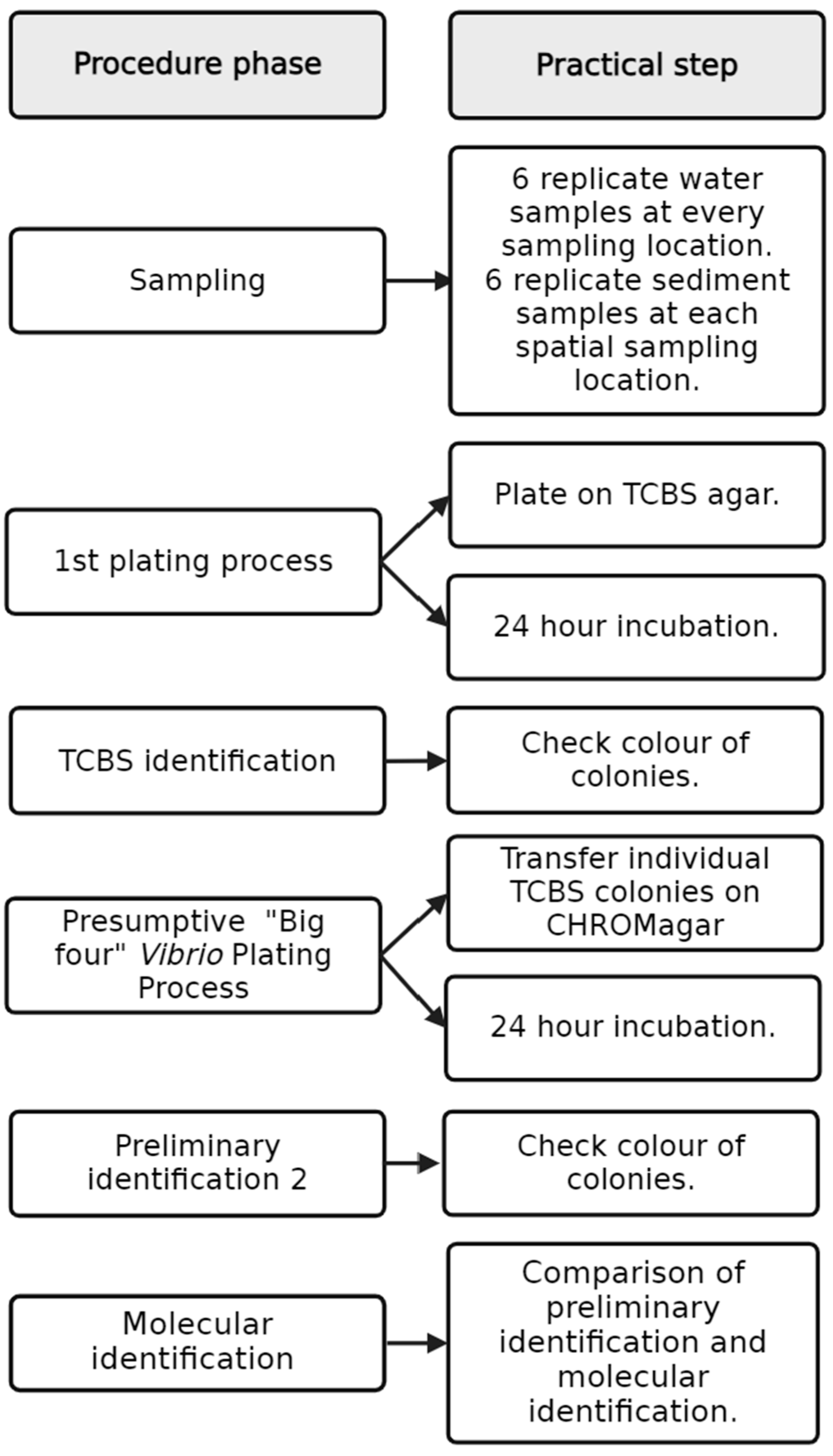
2.3. Vibrio spp. Isolation and Culture-Based Identification
2.4. Molecular Identification of Vibrio spp.
2.5. Statistical Analyses
3. Results
3.1. Environmental Parameters
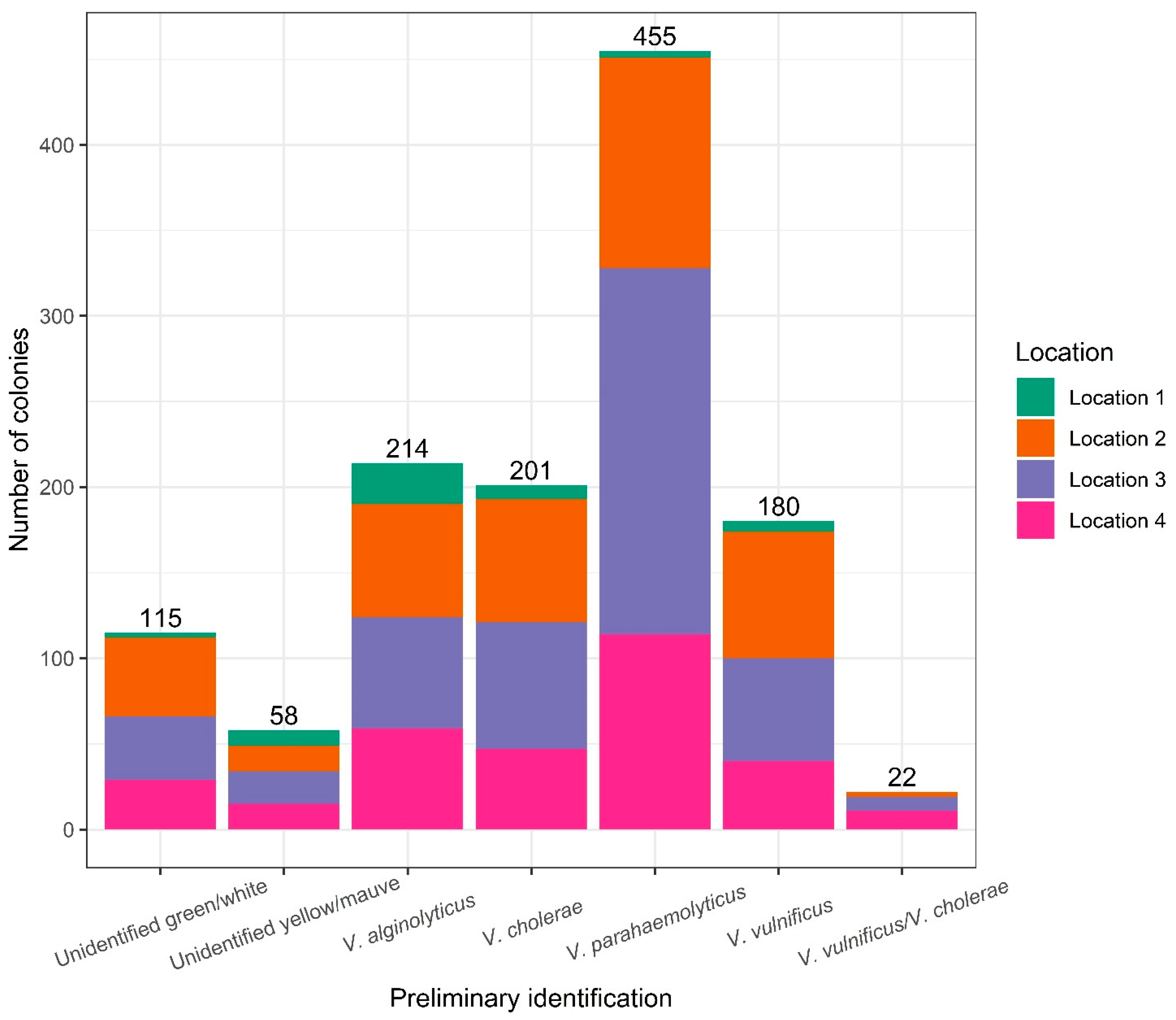
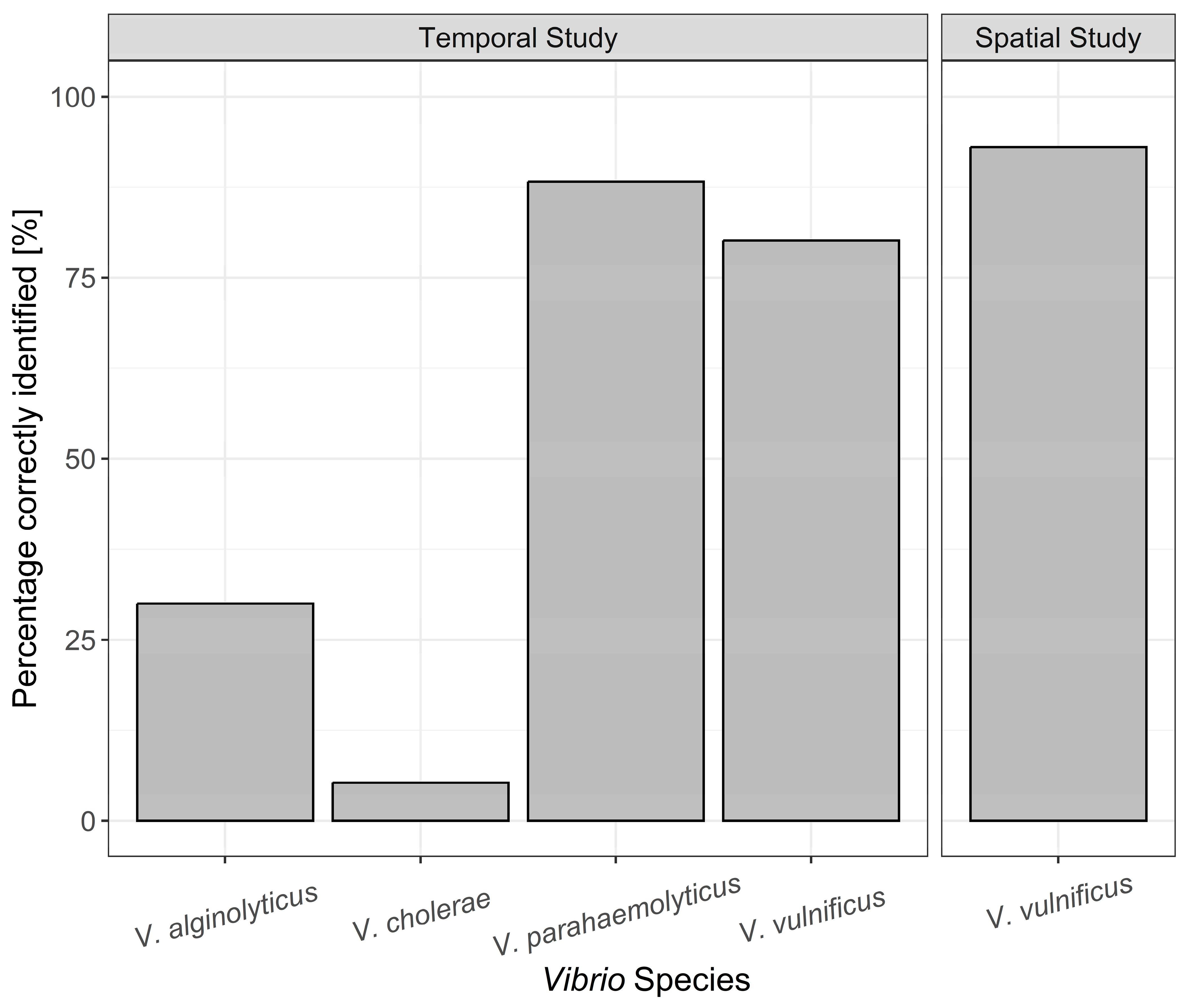
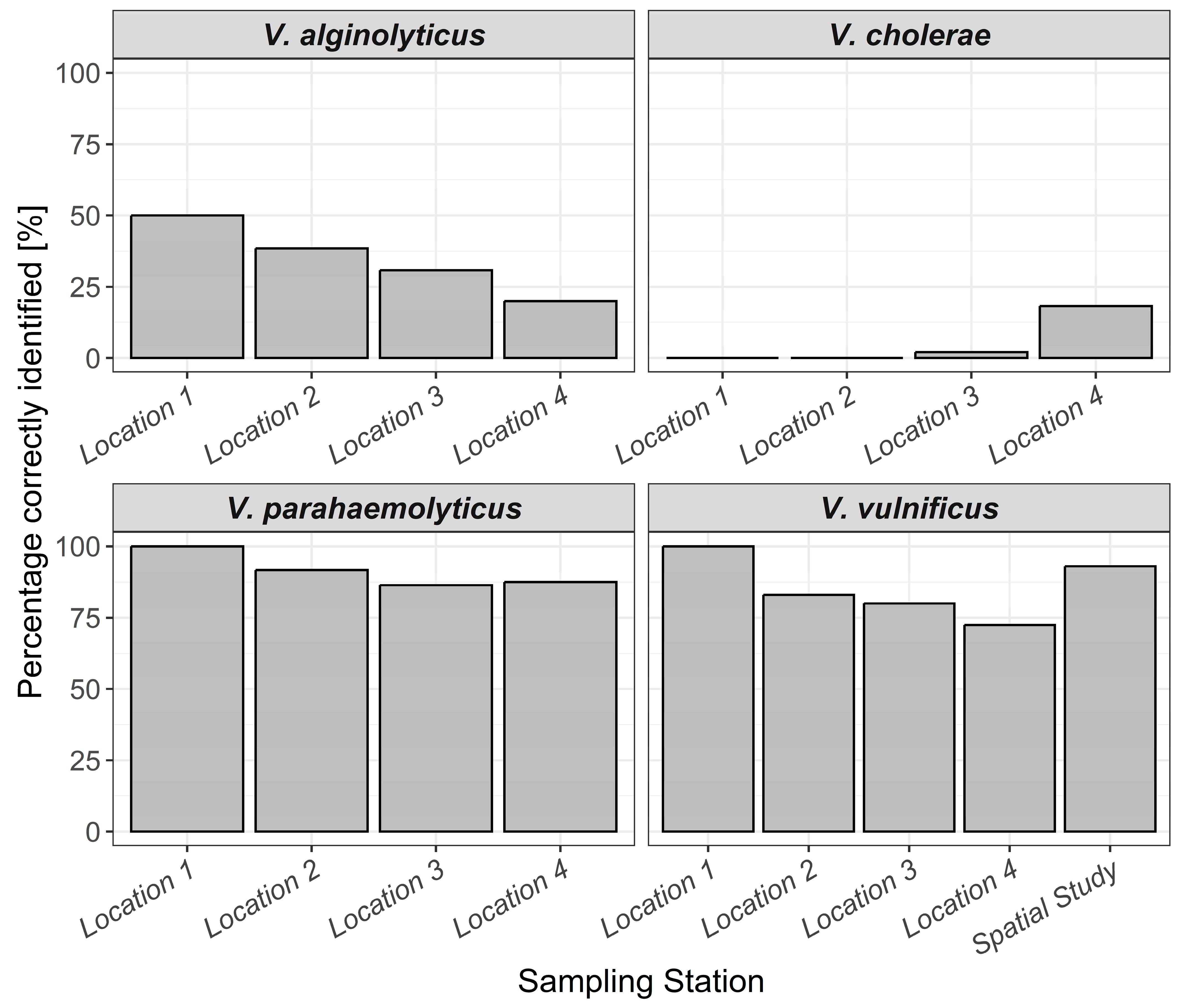
3.2. Identification of Bacterial Isolates on TCBS Agar and CHROMagar™
3.3. Highest Vibrio spp. Abundance per Day
4. Discussion
Spatiotemporal Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Roux, F.; Blokesch, M. Eco-evolutionary dynamics linked to horizontal gene transfer in Vibrios. Annu. Rev. Microbiol. 2018, 72, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Oliver, J.D.; Alam, M.; Ali, A.; Waldor, M.K.; Qadri, F.; Martinez-Urtaza, J. Vibrio spp. infections. Nat. Rev. Dis. Primers 2018, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Potdukhe, T.V.; Caffrey, J.M.; Rothfus, M.J.; Daniel, C.E.; Swords, M.E.; Albrecht, B.B.; Jeffrey, W.H.; Waidner, L.A. Viable putative Vibrio vulnificus and parahaemolyticus in the Pensacola and Perdido Bays: Water column, sediments, and invertebrate biofilms. Front. Mar. Sci. 2021, 8, 645755. [Google Scholar] [CrossRef]
- Ceccarelli, D.; Amaro, C.; Romalde, J.L.; Suffredini, E.; Vezzulli, L. Vibrio species. In Food Microbiology; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2019; pp. 347–388. [Google Scholar]
- Sampaio, A.; Silva, V.; Poeta, P.; Aonofriesei, F. Vibrio spp.: Life strategies, ecology, and risks in a changing environment. Diversity 2022, 14, 97. [Google Scholar] [CrossRef]
- Thompson, F.L.; Austin, B.; Swings, J. The Biology of Vibrios; ASM Press: Washington, DC, USA, 2006. [Google Scholar]
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-cholera Vibrios: The microbial barometer of climate change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Crim, S.M.; Iwamoto, M.; Huang, J.Y.; Griffin, P.M.; Gilliss, D.; Cronquist, A.B.; Cartter, M.; Tobin-D’Angelo, M.; Blythe, D.; Smith, K.; et al. Incidence and trends of infection with pathogens transmitted commonly through food--Foodborne Diseases Active Surveillance Network, 10 U.S. sites, 2006–2013. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 328–332. [Google Scholar]
- Baker-Austin, C.; Trinanes, J.A.; Taylor, N.G.H.; Hartnell, R.; Siitonen, A.; Martinez-Urtaza, J. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Clim. Chang. 2013, 3, 73–77. [Google Scholar] [CrossRef]
- Jones, M.K.; Oliver, J.D. Vibrio vulnificus: Disease and pathogenesis. Infect. Immun. 2009, 77, 1723–1733. [Google Scholar] [CrossRef]
- Amato, E.; Riess, M.; Thomas-Lopez, D.; Linkevicius, M.; Pitkänen, T.; Wołkowicz, T.; Rjabinina, J.; Jernberg, C.; Hjertqvist, M.; MacDonald, E.; et al. Epidemiological and microbiological investigation of a large increase in vibriosis, northern Europe, 2018. Euro Surveill. 2022, 27, 2101088. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trinanes, J.A.; Salmenlinna, S.; Löfdahl, M.; Siitonen, A.; Taylor, N.G.H.; Martinez-Urtaza, J. Heat wave-associated Vibriosis, Sweden and Finland, 2014. Emerg. Infect. Dis. 2016, 22, 1216–1220. [Google Scholar] [CrossRef]
- Brehm, T.T.; Berneking, L.; Sena Martins, M.; Dupke, S.; Jacob, D.; Drechsel, O.; Bohnert, J.; Becker, K.; Kramer, A.; Christner, M.; et al. Heatwave-associated Vibrio infections in Germany, 2018 and 2019. Euro Surveill. 2021, 26, 2002041. [Google Scholar] [CrossRef]
- Oliver, J.D. The biology of Vibrio vulnificus. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef]
- Maar, M.; Møller, E.F.; Larsen, J.; Madsen, K.S.; Wan, Z.; She, J.; Jonasson, L.; Neumann, T. Ecosystem modelling across a salinity gradient from the North Sea to the Baltic Sea. Ecol. Model. 2011, 222, 1696–1711. [Google Scholar] [CrossRef]
- Belkin, I.M. Rapid warming of large marine ecosystems. Prog. Oceanogr. 2009, 81, 207–213. [Google Scholar] [CrossRef]
- Frank, C.; Littman, M.; Alpers, K.; Hallauer, J. Vibrio vulnificus wound infections after contact with the Baltic Sea, Germany. Euro Surveill. 2006, 11, E060817.1. [Google Scholar] [CrossRef]
- New World Encyclopedia. Baltic Sea—New World Encyclopedia, 2023UTC. Available online: https://www.newworldencyclopedia.org/p/index.php?title=Baltic_Sea&oldid=1122453 (accessed on 15 January 2024).
- Schütt, E.M.; Hundsdörfer, M.A.J.; von Hoyningen-Huene, A.J.E.; Lange, X.; Koschmider, A.; Oppelt, N. First steps towards a near real-time modelling system of Vibrio vulnificus in the Baltic Sea. Int. J. Environ. Res. Public Health 2023, 20, 5543. [Google Scholar] [CrossRef] [PubMed]
- Choopun, N.; Louis, V.; Huq, A.; Colwell, R.R. Simple procedure for rapid identification of Vibrio cholerae from the aquatic environment. Appl. Environ. Microbiol. 2002, 68, 995–998. [Google Scholar] [CrossRef] [PubMed]
- Di Pinto, A.; Terio, V.; Novello, L.; Tantillo, G. Comparison between thiosulphate-citrate-bile salt sucrose (TCBS) agar and CHROMagar Vibrio for isolating Vibrio parahaemolyticus. Food Control 2011, 22, 124–127. [Google Scholar] [CrossRef]
- Tamura, K.; Shimada, S.; Prescott, L.M. Vibrio agar: A new plating medium for isolation of Vibrio cholerae. Jpn. J. Med. Sci. Biol. 1971, 24, 125–127. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.C.; Froelich, B.; Oliver, J.D. A new culture-based method for the improved identification of Vibrio vulnificus from environmental samples, reducing the need for molecular confirmation. J. Microbiol. Methods 2013, 93, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.D. Chapter 17 Culture media for the isolation and enumeration of pathogenic Vibrio species in foods and environmental samples. In Rapid Methods in Food Microbiology; Hope, C.F.A., Adams, M.R., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 249–269. ISBN 0079-6352. [Google Scholar]
- Gyraite, G.; Katarzyte, M.; Schernewski, G. First findings of potentially human pathogenic bacteria Vibrio in the south-eastern Baltic Sea coastal and transitional bathing waters. Mar. Pollut. Bull. 2019, 149, 110546. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gil, B.; Roque, A. Isolation, Enumeration, and Preservation of the Vibrionaceae. In The Biology of Vibrios; Thompson, F.L., Austin, B., Swings, J., Eds.; ASM Press: Washington, DC, USA, 2006; pp. 13–26. ISBN 9781683671787. [Google Scholar]
- Messelhäusser, U.; Colditz, J.; Thärigen, D.; Kleih, W.; Höller, C.; Busch, U. Detection and differentiation of Vibrio spp. in seafood and fish samples with cultural and molecular methods. Int. J. Food. Microbiol. 2010, 142, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Tarr, C.L.; Patel, J.S.; Puhr, N.D.; Sowers, E.G.; Bopp, C.A.; Strockbine, N.A. Identification of Vibrio isolates by a multiplex PCR assay and rpoB sequence determination. J. Clin. Microbiol. 2007, 45, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Schirmeister, F.; Dieckmann, R.; Bechlars, S.; Bier, N.; Faruque, S.M.; Strauch, E. Genetic and phenotypic analysis of Vibrio cholerae non-O1, non-O139 isolated from German and Austrian patients. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, S.; Herrig, I.; Wesp, J.; Stiedl, J.; Reifferscheid, G.; Strauch, E.; Alter, T.; Brennholt, N. Prevalence and distribution of potentially human pathogenic Vibrio spp. on German North and Baltic Sea coasts. Front. Cell Infect. Microbiol. 2022, 12, 846819. [Google Scholar] [CrossRef]
- Lotz, M.J.; Tamplin, M.L.; Rodrick, G.E. Thiosulfate-citrate-bile salts-sucrose agar and its selectivity for clinical and marine vibrio organisms. Ann. Clin. Lab. 1983, 13, 45–48. [Google Scholar]
- Froelich, B.A.; Weiss, M.J.; Noble, R.T. The evaluation of four recent culture-based methods for the isolation and enumeration of Vibrio vulnificus bacteria from oyster meat. J. Microbiol. Methods 2014, 97, 1–5. [Google Scholar] [CrossRef]
- Froelich, B.A.; Williams, T.C.; Noble, R.T.; Oliver, J.D. Apparent loss of Vibrio vulnificus from North Carolina oysters coincides with a drought-induced increase in salinity. Appl. Environ. Microbiol. 2012, 78, 3885–3889. [Google Scholar] [CrossRef]
- Arias, C.R.; Aznar, R.; Pujalte, M.J.; Garay, E. A comparison of strategies for the detection and recovery of Vibrio vulnificus from marine samples of the western Mediterranean coast. Syst. Appl. Microbiol. 1998, 21, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.R. Global climate and infectious disease: The cholera paradigm. Science 1996, 274, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- DePaola, A.; Nordstrom, J.L.; Bowers, J.C.; Wells, J.G.; Cook, D.W. Seasonal abundance of total and pathogenic Vibrio parahaemolyticus in Alabama oysters. Appl. Environ. Microbiol. 2003, 69, 1521–1526. [Google Scholar] [CrossRef]
- Grimes, D.J.; Johnson, C.N.; Dillon, K.S.; Flowers, A.R.; Noriea, N.F.; Berutti, T. What genomic sequence information has revealed about Vibrio ecology in the ocean—A review. Microb. Ecol. 2009, 58, 447–460. [Google Scholar] [CrossRef]
- Huq, A.; West, P.A.; Small, E.B.; Huq, M.I.; Colwell, R.R. Influence of water temperature, salinity, and pH on survival and growth of toxigenic Vibrio cholerae serovar 01 associated with live copepods in laboratory microcosms. Appl. Environ. Microbiol. 1984, 48, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.N.; Flowers, A.R.; Noriea, N.F.; Zimmerman, A.M.; Bowers, J.C.; DePaola, A.; Grimes, D.J. Relationships between environmental factors and pathogenic Vibrios in the Northern Gulf of Mexico. Appl. Environ. Microbiol. 2010, 76, 7076–7084. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.T. Effect of temperature and salinity on Vibrio (Beneckea) vulnificus occurrence in a Gulf Coast environment. Appl. Environ. Microbiol. 1982, 44, 820–824. [Google Scholar] [CrossRef] [PubMed]
- Stauder, M.; Vezzulli, L.; Pezzati, E.; Repetto, B.; Pruzzo, C. Temperature affects Vibrio cholerae O1 El Tor persistence in the aquatic environment via an enhanced expression of GbpA and MSHA adhesins. Environ. Microbiol. Rep. 2010, 2, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Lobitz, B.; Beck, L.; Huq, A.; Wood, B.; Fuchs, G.; Faruque, A.S.; Colwell, R. Climate and infectious disease: Use of remote sensing for detection of Vibrio cholerae by indirect measurement. Proc. Natl. Acad. Sci. USA 2000, 97, 1438–1443. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, A.M.; DePaola, A.; Bowers, J.C.; Krantz, J.A.; Nordstrom, J.L.; Johnson, C.N.; Grimes, D.J. Variability of total and pathogenic Vibrio parahaemolyticus densities in northern Gulf of Mexico water and oysters. Appl. Environ. Microbiol. 2007, 73, 7589–7596. [Google Scholar] [CrossRef]
- Tamplin, M.; Rodrick, G.E.; Blake, N.J.; Cuba, T. Isolation and characterization of Vibrio vulnificus from two Florida estuaries. Appl. Environ. Microbiol. 1982, 44, 1466–1470. [Google Scholar] [CrossRef]
- Igbinosa, E.O.; Obi, C.L.; Okoh, A.I. Seasonal abundance and distribution of Vibrio species in the treated effluent of wastewater treatment facilities in suburban and urban communities of Eastern Cape Province, South Africa. J. Microbiol. 2011, 49, 224–232. [Google Scholar] [CrossRef]
- Parveen, S.; Hettiarachchi, K.A.; Bowers, J.C.; Jones, J.L.; Tamplin, M.L.; McKay, R.; Beatty, W.; Brohawn, K.; Dasilva, L.V.; DePaola, A. Seasonal distribution of total and pathogenic Vibrio parahaemolyticus in Chesapeake Bay oysters and waters. Int. J. Food Microbiol. 2008, 128, 354–361. [Google Scholar] [CrossRef]
- Ramirez, G.D.; Buck, G.W.; Smith, A.K.; Gordon, K.V.; Mott, J.B. Incidence of Vibrio vulnificus in estuarine waters of the south Texas Coastal Bend region. J. Appl. Microbiol. 2009, 107, 2047–2053. [Google Scholar] [CrossRef]
- Caburlotto, G.; Haley, B.J.; Lleò, M.M.; Huq, A.; Colwell, R.R. Serodiversity and ecological distribution of Vibrio parahaemolyticus in the Venetian Lagoon, Northeast Italy. Environ. Microbiol. Rep. 2010, 2, 151–157. [Google Scholar] [CrossRef]
- Deter, J.; Lozach, S.; Derrien, A.; Véron, A.; Chollet, J.; Hervio-Heath, D. Chlorophyll a might structure a community of potentially pathogenic culturable Vibrionaceae. Insights from a one-year study of water and mussels surveyed on the French Atlantic coast. Environ. Microbiol. Rep. 2010, 2, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Julie, D.; Solen, L.; Antoine, V.; Jaufrey, C.; Annick, D.; Dominique, H.-H. Ecology of pathogenic and non-pathogenic Vibrio parahaemolyticus on the French Atlantic coast. Effects of temperature, salinity, turbidity and chlorophyll a. Environ. Microbiol. 2010, 12, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Asplund, M.E.; Rehnstam-Holm, A.-S.; Atnur, V.; Raghunath, P.; Saravanan, V.; Härnström, K.; Collin, B.; Karunasagar, I.; Godhe, A. Water column dynamics of Vibrio in relation to phytoplankton community composition and environmental conditions in a tropical coastal area. Environ. Microbiol. 2011, 13, 2738–2751. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Urtaza, J.; Huapaya, B.; Gavilan, R.G.; Blanco-Abad, V.; Ansede-Bermejo, J.; Cadarso-Suarez, C.; Figueiras, A.; Trinanes, J. Emergence of Asiatic Vibrio diseases in South America in phase with El Niño. Epidemiology 2008, 19, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Rehnstam-Holm, A.S.; Godhe, A.; Härnström, K.; Raghunath, P.; Saravanan, V.; Collin, B.; Karunasagar, I. Association between phytoplankton and Vibrio spp. along the southwest coast of India: A mesocosm experiment. Aquat. Microb. Ecol. 2010, 58, 127–139. [Google Scholar] [CrossRef]
- Turner, J.W.; Good, B.; Cole, D.; Lipp, E.K. Plankton composition and environmental factors contribute to Vibrio seasonality. ISME J. 2009, 3, 1082–1092. [Google Scholar] [CrossRef]
- Novriadi, R. Vibriosis in aquaculture. Omni Akuatika 2016, 12, 1–2. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TCBS CFU Colour | CHROM CFU Colour | Vibrio Preliminary Identification |
|---|---|---|
| Green | Blue | V. vulnificus |
| Green | Mauve | V. parahaemolyticus |
| Yellow | Blue | V. cholerae |
| Yellow | White | V. alginolyticus/Other Vibrio spp. |
| Target Species | Primer/Probe | Sequenz (5′–3′) | Localisation |
|---|---|---|---|
| V. cholerae | sodB f | AAGACCTCAACTGGCGGTA | 276–294 |
| sodB r | CAGCAAAAGAACCGAATGCT | 420–401 | |
| sodB TM | Cy5-GCAGGTTTGGAACCACACTT-BHQ-2 | 311–330 | |
| ctx f | AGTTCATTTTGGGGTGCTTG | 369–388 | |
| ctx r | GGAAACCTGCCAATCCATAA | 484–465 | |
| ctx TM | FAM-CATCGTAATAGGGGCTACAGAGA-BHQ-1 | 400–422 | |
| V. parahaemolyticus | toxR f | CCAGAAGCGCCAGTAGTACC | 149–168 |
| toxR r | AAACAGCAGTACGCAAATCG | 262–243 | |
| toxR TM | FAM-TGTGGCTTCTGCTGTGAATC-BHQ-1 | 181–200 | |
| V. vulnificus | vvhA f | ACCAAGTTTGGGGCCTAGAT | 389–408 |
| vvhA r | GCTAAGTTCGCACCACACTG | 506–487 | |
| vvhA TM | Cy5-CCGATCGTTGTTTGACCGTA-BHQ-2 | 440–459 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glackin, C.C.; Dupke, S.; Chandra, T.S.; Riedinger, D.; Labrenz, M. Combined TCBS and CHROMagar Analyses Allow for Basic Identification of Vibrio vulnificus within a 48 h Incubation Period in the Coastal Baltic Sea. Microorganisms 2024, 12, 614. https://doi.org/10.3390/microorganisms12030614
Glackin CC, Dupke S, Chandra TS, Riedinger D, Labrenz M. Combined TCBS and CHROMagar Analyses Allow for Basic Identification of Vibrio vulnificus within a 48 h Incubation Period in the Coastal Baltic Sea. Microorganisms. 2024; 12(3):614. https://doi.org/10.3390/microorganisms12030614
Chicago/Turabian StyleGlackin, Conor Christopher, Susann Dupke, Thota Sharath Chandra, David Riedinger, and Matthias Labrenz. 2024. "Combined TCBS and CHROMagar Analyses Allow for Basic Identification of Vibrio vulnificus within a 48 h Incubation Period in the Coastal Baltic Sea" Microorganisms 12, no. 3: 614. https://doi.org/10.3390/microorganisms12030614