
Ligand-Free Silver Nanoparticles: An Innovative Strategy against Viruses and Bacteria
 , , , ,
, , , ,  , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Methods for Production of AgNPs
2.2. PLAL-AgNP Characterization
2.2.1. Total X-ray Fluorescence
2.2.2. Transmission Electron Microscopy (TEM) Analysis
2.2.3. Zeta Potential
2.2.4. UV-Vis
2.2.5. Determination of AgNP Concentration Nanoparticle Tracking Analysis (NTA)
2.3. Cell Lines and Viral Strains
2.4. Bacteria and Growth Conditions
2.5. Cell Viability
2.6. Antiviral Activity
2.7. Molecular Analysis
2.8. Antibacterial Activity
2.8.1. Microdilution Assay
2.8.2. Time–Killing Analysis
2.8.3. Biofilm Inhibition and Degradation
2.9. Scanning Electron Microscopy (SEM) Analysis
2.10. Statistical Analysis
3. Results and Discussion
3.1. PLAL-AgNP Characterization
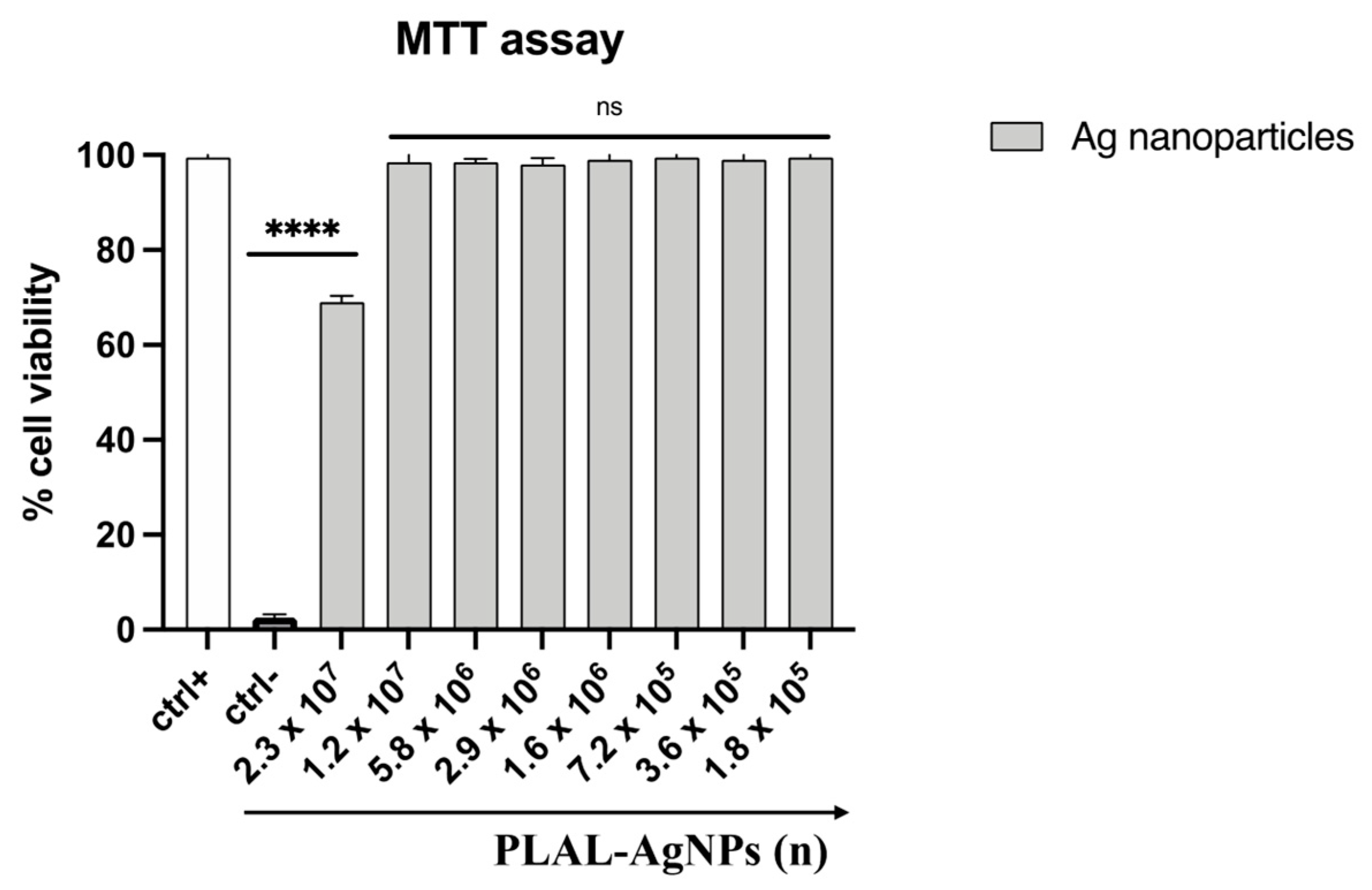
3.2. PLAL-AgNP Cytotoxicity Evaluation
3.3. Evaluation of Antiviral Activity
3.3.1. HSV-1
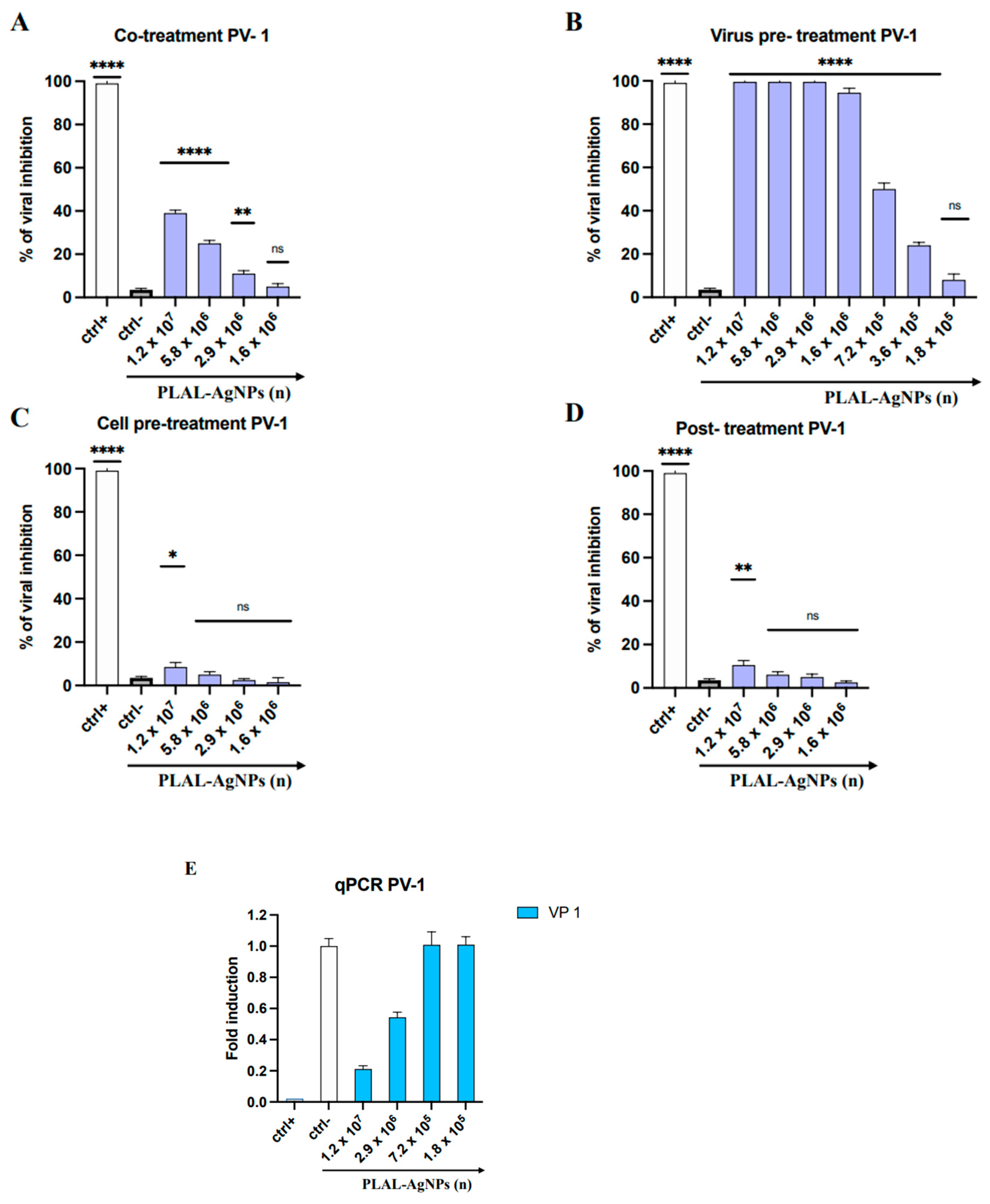
3.3.2. PV-1

3.4. Antibacterial Activity
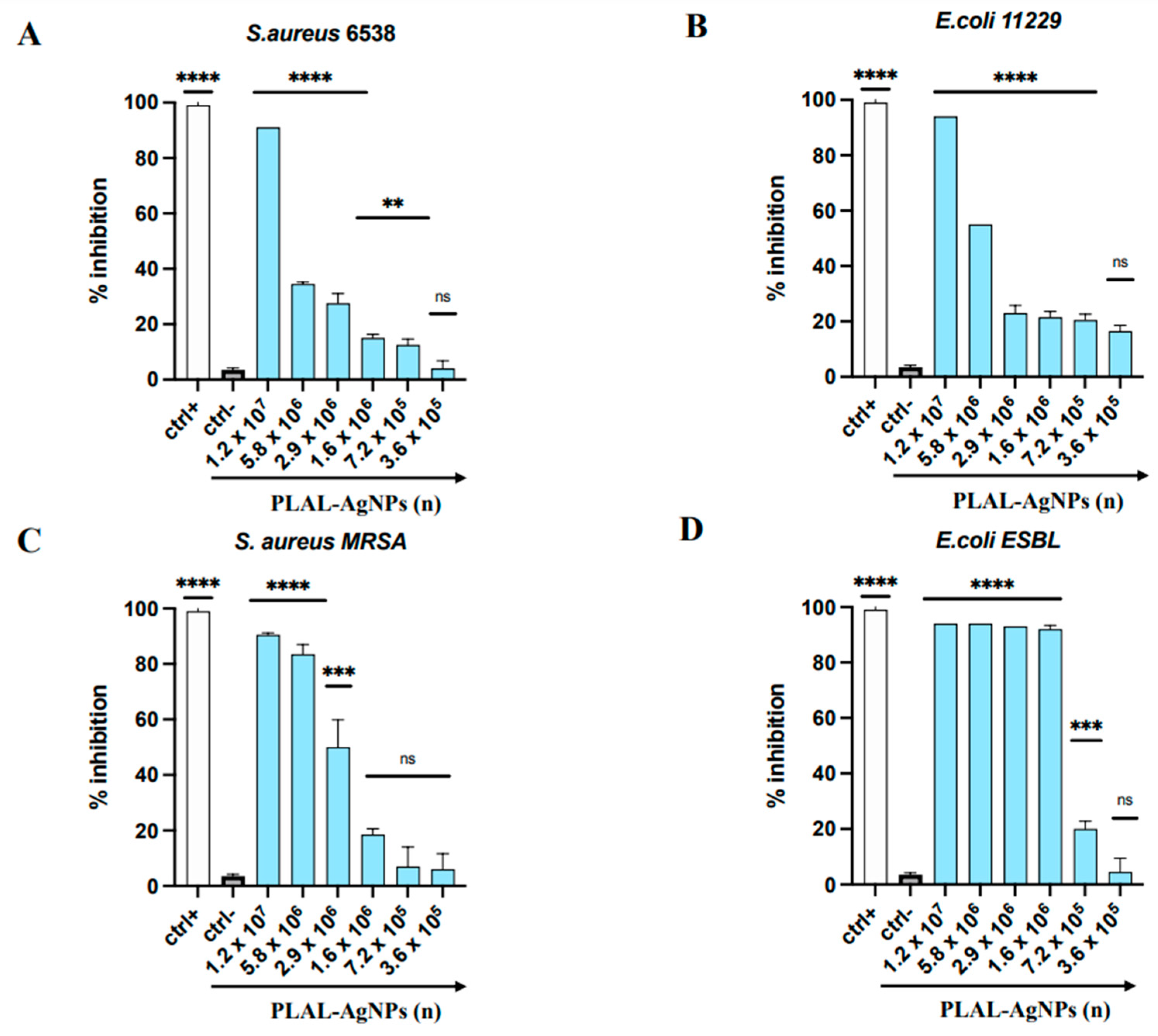
3.4.1. MIC and Time–Killing Assay
3.4.2. SEM Analysis
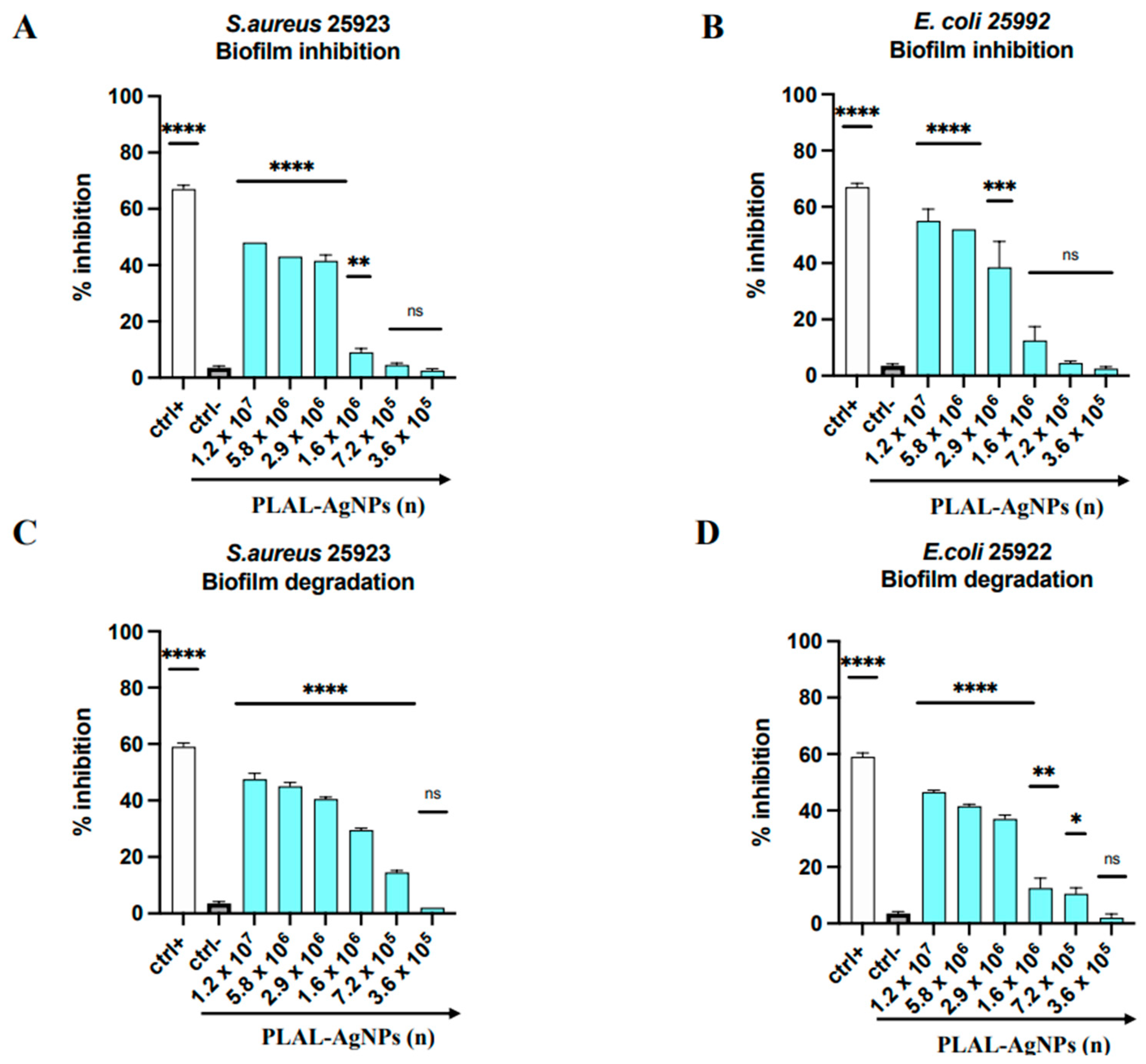
3.5. Antibiofilm Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Van Seventer, J.M.; Hochberg, N.S. Principles of Infectious Diseases: Transmission, Diagnosis, Prevention, and Control. In International Encyclopedia of Public Health; Elsevier: Amsterdam, The Netherlands, 2017; pp. 22–39. ISBN 978-0-12-803708-9. [Google Scholar]
- Ismahene, Y. Infectious Diseases, Trade, and Economic Growth: A Panel Analysis of Developed and Developing Countries. J. Knowl. Econ. 2022, 13, 2547–2583. [Google Scholar] [CrossRef]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef]
- Jiang, Y.-C.; Feng, H.; Lin, Y.-C.; Guo, X.-R. New strategies against drug resistance to herpes simplex virus. Int. J. Oral Sci. 2016, 8, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Anton-Vazquez, V.; Mehra, V.; Mbisa, J.L.; Bradshaw, D.; Basu, T.N.; Daly, M.-L.; Mufti, G.J.; Pagliuca, A.; Potter, V.; Zuckerman, M. Challenges of aciclovir-resistant HSV infection in allogeneic bone marrow transplant recipients. J. Clin. Virol. 2020, 128, 104421. [Google Scholar] [CrossRef]
- Chayavichitsilp, P.; Buckwalter, J.V.; Krakowski, A.C.; Friedlander, S.F. Herpes Simplex. Pediatr. Rev. 2009, 30, 119–130. [Google Scholar] [CrossRef]
- Cunningham, A.L.; Diefenbach, R.J.; Miranda-Saksena, M.; Bosnjak, L.; Kim, M.; Jones, C.; Douglas, M.W. The Cycle of Human Herpes Simplex Virus Infection: Virus Transport and Immune Control. J. Infect. Dis. 2006, 194, S11–S18. [Google Scholar] [CrossRef]
- Marjomäki, V.; Kalander, K.; Hellman, M.; Permi, P. Enteroviruses and coronaviruses: Similarities and therapeutic targets. Expert Opin. Ther. Targets 2021, 25, 479–489. [Google Scholar] [CrossRef]
- Chen, B.-S.; Lee, H.-C.; Lee, K.-M.; Gong, Y.-N.; Shih, S.-R. Enterovirus and Encephalitis. Front. Microbiol. 2020, 11, 261. [Google Scholar] [CrossRef]
- Available online: https://www.who.int/news-room/fact-sheets/detail/poliomyelitis (accessed on 10 March 2024).
- Akram, F.; Imtiaz, M.; Haq, I.U. Emergent crisis of antibiotic resistance: A silent pandemic threat to 21st century. Microb. Pathog. 2023, 174, 105923. [Google Scholar] [CrossRef]
- Asokan, G.V.; Ramadhan, T.; Ahmed, E.; Sanad, H. WHO Global Priority Pathogens List: A Bibliometric Analysis of Medline-PubMed for Knowledge Mobilization to Infection Prevention and Control Practices in Bahrain. Oman Med. J. 2019, 34, 184–193. [Google Scholar] [CrossRef]
- Kumar, M.; Sarma, D.K.; Shubham, S.; Kumawat, M.; Verma, V.; Nina, P.B.; Jp, D.; Kumar, S.; Singh, B.; Tiwari, R.R. Futuristic Non-antibiotic Therapies to Combat Antibiotic Resistance: A Review. Front. Microbiol. 2021, 12, 609459. [Google Scholar] [CrossRef] [PubMed]
- Sharmin, S.; Rahaman, M.M.; Sarkar, C.; Atolani, O.; Islam, M.T.; Adeyemi, O.S. Nanoparticles as antimicrobial and antiviral agents: A literature-based perspective study. Heliyon 2021, 7, e06456. [Google Scholar] [CrossRef] [PubMed]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; Rodriguez-Torres, M.D.P.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano based drug delivery systems: Recent developments and future prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef] [PubMed]
- Lamparelli, E.P.; Ciardulli, M.C.; Scala, P.; Scognamiglio, M.; Charlier, B.; Di Pietro, P.; Izzo, V.; Vecchione, C.; Maffulli, N.; Della Porta, G. Lipid nano-vesicles for thyroid hormone encapsulation: A comparison between different fabrication technologies, drug loading, and an in vitro delivery to human tendon stem/progenitor cells in 2D and 3D culture. Int. J. Pharm. 2022, 624, 122007. [Google Scholar] [CrossRef] [PubMed]
- Lamparelli, E.P.; Marino, M.; Szychlinska, M.A.; Della Rocca, N.; Ciardulli, M.C.; Scala, P.; D′Auria, R.; Testa, A.; Viggiano, A.; Cappello, F.; et al. The Other Side of Plastics: Bioplastic-Based Nanoparticles for Drug Delivery Systems in the Brain. Pharmaceutics 2023, 15, 2549. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, I.; Lamparelli, E.P.; Ciardulli, M.C.; Scala, P.; Reverchon, E.; Forsyth, N.; Maffulli, N.; Santoro, A.; Della Porta, G. Supercritical emulsion extraction fabricated PLA/PLGA micro/nano carriers for growth factor delivery: Release profiles and cytotoxicity. Int. J. Pharm. 2021, 592, 120108. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Núñez, M.F.; Castillo-López, M.; Sevilla-Castillo, F.; Roque-Reyes, O.J.; Romero-Lechuga, F.; Medina-Santos, D.I.; Martínez-Daniel, R.; Peón, A.N. Nanoparticle-Based Devices in the Control of Antibiotic Resistant Bacteria. Front. Microbiol. 2020, 11, 563821. [Google Scholar] [CrossRef]
- More, P.R.; Zannella, C.; Folliero, V.; Foglia, F.; Troisi, R.; Vergara, A.; Franci, G.; De Filippis, A.; Galdiero, M. Antimicrobial Applications of Green Synthesized Bimetallic Nanoparticles from Ocimum Basilicum. Pharmaceutics 2022, 14, 2457. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jun, B.-H. Silver Nanoparticles: Synthesis and Application for Nanomedicine. Int. J. Mol. Sci. 2019, 20, 865. [Google Scholar] [CrossRef]
- Thambirajoo, M.; Maarof, M.; Lokanathan, Y.; Katas, H.; Ghazalli, N.F.; Tabata, Y.; Fauzi, M.B. Potential of Nanoparticles Integrated with Antibacterial Properties in Preventing Biofilm and Antibiotic Resistance. Antibiotics 2021, 10, 1338. [Google Scholar] [CrossRef]
- Rodriguez-Garraus, A.; Azqueta, A.; Vettorazzi, A.; López de Cerain, A. Genotoxicity of Silver Nanoparticles. Nanomaterials 2020, 10, 251. [Google Scholar] [CrossRef] [PubMed]
- Khairani, I.Y.; Mínguez-Vega, G.; Doñate-Buendía, C.; Gökce, B. Green nanoparticle synthesis at scale: A perspective on overcoming the limits of pulsed laser ablation in liquids for high-throughput production. Phys. Chem. Chem. Phys. 2023, 25, 19380–19408. [Google Scholar] [CrossRef] [PubMed]
- Sportelli, M.C.; Izzi, M.; Volpe, A.; Clemente, M.; Picca, R.A.; Ancona, A.; Lugarà, P.M.; Palazzo, G.; Cioffi, N. The Pros and Cons of the Use of Laser Ablation Synthesis for the Production of Silver Nano-Antimicrobials. Antibiotics 2018, 7, 67. [Google Scholar] [CrossRef] [PubMed]
- Lazar, O.A.; Nikolov, A.S.; Moise, C.C.; Enachescu, M. Pulsed Laser Ablation in Liquids for Fabrication of Noble Metal Nanostructures. In Laser Ablation - Applications and Modeling; IntechOpen: London, England, 2023. [Google Scholar] [CrossRef]
- Ahmad, S.A.; Das, S.S.; Khatoon, A.; Ansari, M.T.; Afzal, M.; Hasnain, S.; Nayak, A.K. Bactericidal activity of silver nanoparticles: A mechanistic review. Mater. Sci. Energy Technol. 2020, 3, 756–769. [Google Scholar] [CrossRef]
- Tsuji, T.; Iryo, K.; Watanabe, N.; Tsuji, M. Preparation of silver nanoparticles by laser ablation in solution: Influence of laser wavelength on particle size. Appl. Surf. Sci. 2002, 202, 80–85. [Google Scholar] [CrossRef]
- Morone, M.V.; Dell’Annunziata, F.; Giugliano, R.; Chianese, A.; De Filippis, A.; Rinaldi, L.; Gambardella, U.; Franci, G.; Galdiero, M.; Morone, A. Pulsed Laser Ablation of Magnetic Nanoparticles as a Novel Antibacterial Strategy against Gram Positive Bacteria. Appl. Surf. Sci. Adv. 2022, 7, 100213. [Google Scholar] [CrossRef]
- Brignati, M.J.; Loomis, J.S.; Wills, J.W.; Courtney, R.J. Membrane Association of VP22, a Herpes Simplex Virus Type 1 Tegument Protein. J. Virol. 2003, 77, 4888–4898. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Zannella, C.; Folliero, V.; Di Girolamo, R.; Bajardi, F.; Chianese, A.; Altucci, L.; Damasco, A.; Del Sorbo, M.R.; Imperatore, C.; et al. Combating Actions of Green 2D-Materials on Gram Positive and Negative Bacteria and Enveloped Viruses. Front. Bioeng. Biotechnol. 2020, 8, 569967. [Google Scholar] [CrossRef]
- Dhaka, A.; Mali, S.C.; Sharma, S.; Trivedi, R. A review on biological synthesis of silver nanoparticles and their potential applications. Results Chem. 2023, 6, 101108. [Google Scholar] [CrossRef]
- Sharma, A.; Sagar, A.; Rana, J.; Rani, R. Green synthesis of silver nanoparticles and its antibacterial activity using fungus Talaromyces purpureogenus isolated from Taxus baccata Linn. Micro Nano Syst. Lett. 2022, 10, 2. [Google Scholar] [CrossRef]
- Zannella, C.; Chianese, A.; De Bernardo, M.; Folliero, V.; Petrillo, F.; De Filippis, A.; Boccia, G.; Franci, G.; Rosa, N.; Galdiero, M. Ophthalmic Solutions with a Broad Antiviral Action: Evaluation of Their Potential against Ocular Herpetic Infections. Microorganisms 2022, 10, 1728. [Google Scholar] [CrossRef] [PubMed]
- Stelitano, D.; Franci, G.; Chianese, A.; Galdiero, S.; Morelli, G.; Galdiero, M. HSV Membrane Glycoproteins, Their Function in Viral Entry and Their Use in Vaccine Studies. In Amino Acids, Peptides and Proteins; Ryadnov, M., Hudecz, F., Eds.; Royal Society of Chemistry: Cambridge, UK, 2019; Volume 43, pp. 14–43. ISBN 978-1-78801-367-3. [Google Scholar]
- Huy, T.Q.; Hien Thanh, N.T.; Thuy, N.T.; Chung, P.V.; Hung, P.N.; Le, A.-T.; Hong Hanh, N.T. Cytotoxicity and Antiviral Activity of Electrochemical—Synthesized Silver Nanoparticles against Poliovirus. J. Virol. Methods 2017, 241, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Sanna, G.; Madeddu, S.; Serra, A.; Collu, D.; Efferth, T.; Hakkim, F.L.; Rashan, L. Anti-Poliovirus Activity of Nerium Oleander Aqueous Extract. Nat. Prod. Res. 2021, 35, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Dove, A.W.; Racaniello, V.R. An Antiviral Compound That Blocks Structural Transitions of Poliovirus Prevents Receptor Binding at Low Temperatures. J. Virol. 2000, 74, 3929–3931. [Google Scholar] [CrossRef]
- Zammuto, V.; Spanò, A.; Agostino, E.; Macrì, A.; De Pasquale, C.; Ferlazzo, G.; Rizzo, M.G.; Nicolò, M.S.; Guglielmino, S.; Gugliandolo, C. Anti-Bacterial Adhesion on Abiotic and Biotic Surfaces of the Exopolysaccharide from the Marine Bacillus licheniformis B3-15. Mar. Drugs 2023, 21, 313. [Google Scholar] [CrossRef]
- Netrusov, A.I.; Liyaskina, E.V.; Kurgaeva, I.V.; Liyaskina, A.U.; Yang, G.; Revin, V.V. Exopolysaccharides Producing Bacteria: A Review. Microorganisms 2023, 11, 1541. [Google Scholar] [CrossRef] [PubMed]
- Siddique, M.H.; Aslam, B.; Imran, M.; Ashraf, A.; Nadeem, H.; Hayat, S.; Khurshid, M.; Afzal, M.; Malik, I.R.; Shahzad, M.; et al. Effect of Silver Nanoparticles on Biofilm Formation and EPS Production of Multidrug-Resistant Klebsiella pneumoniae. Biomed. Res. Int. 2020, 2020, 6398165. [Google Scholar] [CrossRef]
- Montazeri, A.; Salehzadeh, A.; Zamani, H. Effect of silver nanoparticles conjugated to thiosemicarbazide on biofilm formation and expression of intercellular adhesion molecule genes, icaAD, in Staphylococcus aureus. Folia Microbiol. 2020, 65, 153–160. [Google Scholar] [CrossRef]
- Swolana, D.; Kępa, M.; Kruszniewska-Rajs, C.; Wojtyczka, R.D. Antibiofilm Effect of Silver Nanoparticles in Changing the Biofilm-Related Gene Expression of Staphylococcus epidermidis. Int. J. Mol. Sci. 2022, 23, 9257. [Google Scholar] [CrossRef]
- Al-Momani, H.; Almasri, M.; Al Balawi, D.; Hamed, S.; Albiss, B.A.; Aldabaibeh, N.; Ibrahim, L.; Albalawi, H.; Al Haj Mahmoud, S.; Khasawneh, A.I.; et al. The efficacy of biosynthesized silver nanoparticles against Pseudomonas aeruginosa isolates from cystic fibrosis patients. Sci. Rep. 2023, 13, 8876. [Google Scholar] [CrossRef]
- Kraśniewska, K.; Galus, S.; Gniewosz, M. Biopolymers-Based Materials Containing Silver Nanoparticles as Active Packaging for Food Applications—A Review. Int. J. Mol. Sci. 2020, 21, 698. [Google Scholar] [CrossRef] [PubMed]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morone, M.V.; Chianese, A.; Dell’Annunziata, F.; Folliero, V.; Lamparelli, E.P.; Della Porta, G.; Zannella, C.; De Filippis, A.; Franci, G.; Galdiero, M.; et al. Ligand-Free Silver Nanoparticles: An Innovative Strategy against Viruses and Bacteria. Microorganisms 2024, 12, 820. https://doi.org/10.3390/microorganisms12040820
Morone MV, Chianese A, Dell’Annunziata F, Folliero V, Lamparelli EP, Della Porta G, Zannella C, De Filippis A, Franci G, Galdiero M, et al. Ligand-Free Silver Nanoparticles: An Innovative Strategy against Viruses and Bacteria. Microorganisms. 2024; 12(4):820. https://doi.org/10.3390/microorganisms12040820
Chicago/Turabian StyleMorone, Maria Vittoria, Annalisa Chianese, Federica Dell’Annunziata, Veronica Folliero, Erwin Pavel Lamparelli, Giovanna Della Porta, Carla Zannella, Anna De Filippis, Gianluigi Franci, Massimiliano Galdiero, and et al. 2024. "Ligand-Free Silver Nanoparticles: An Innovative Strategy against Viruses and Bacteria" Microorganisms 12, no. 4: 820. https://doi.org/10.3390/microorganisms12040820