
Priestia megaterium ASC-1 Isolated from Pickled Cabbage Ameliorates Hyperuricemia by Degrading Uric Acid in Rats
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of the Bacterial Strain with UA-Degrading Ability
2.2. Determination of Uric Acid Degradation Ability of Strains In Vitro
2.3. 16S rRNA Gene Sequencing and Phylogenetic Analysis
2.4. Experimental Design Based on the Animal Model
2.5. Colonization of Priestia Megaterium ASC-1
2.6. Determination of UA in Rats
2.7. Determination of Short-Chain Fatty Acids (SCFAs) in Rat Feces
2.8. Tissues Collection and Biomarker Measurements
2.9. Degradation Ability of UA by Rats’ Fecal Flora
2.10. DNA Extraction and 16S rRNA Gene Amplification
2.11. Statistical Analysis
3. Results
3.1. Isolation of UA-Degrading Strains and Product Identification
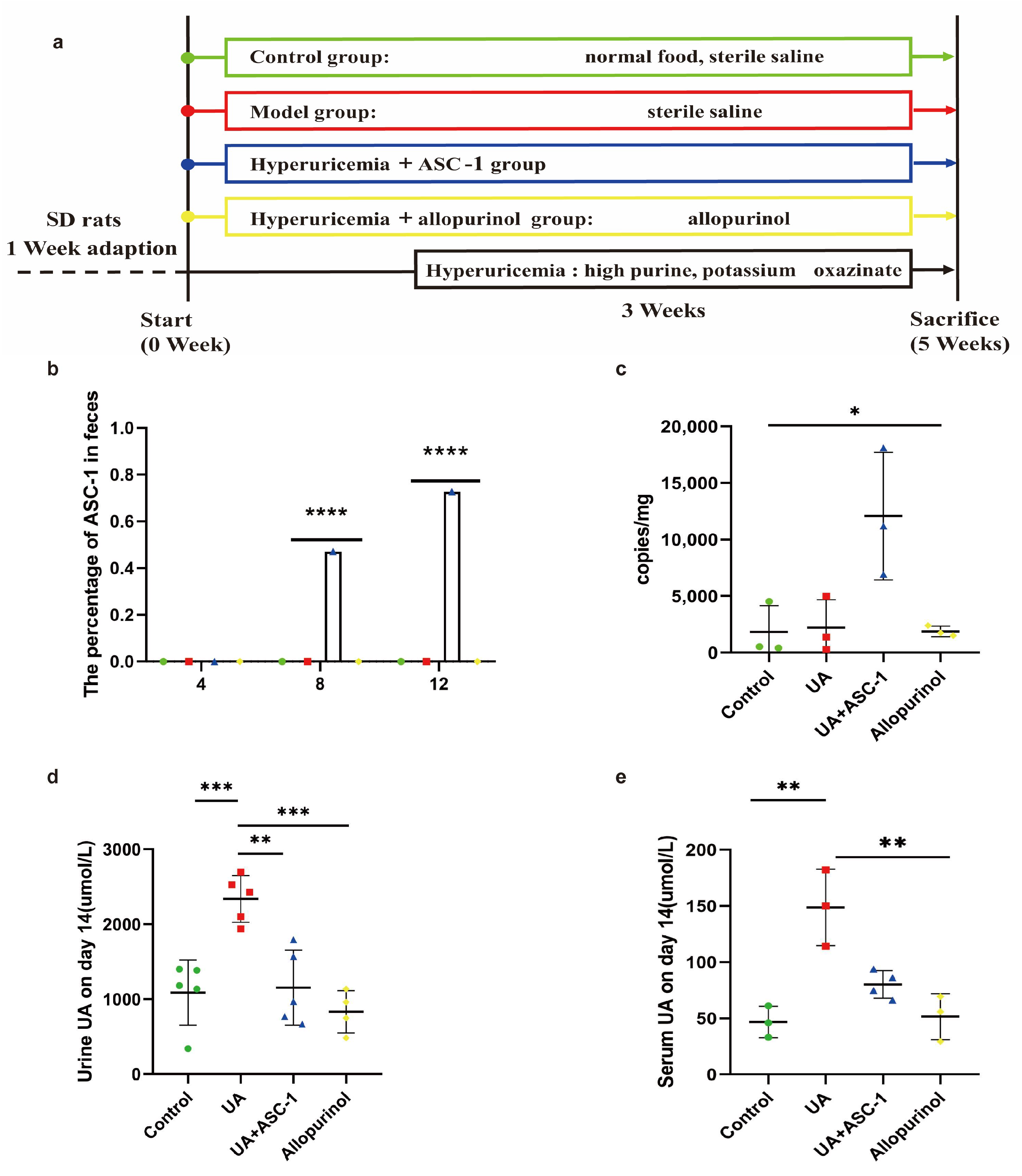
3.2. ASC-1 Decreases UA Level in Hyperuricemic Rats
3.3. ASC-1 Strain Attenuated UA-Induced Oxidative Stress and Inflammation
3.4. ASC-1 Regulated UA-Induced Gut Microbiota Dysbiosis in Hyperuricemic Rats
4. Discussion
4.1. ASC-1 Efficiently Degraded Uric Acid and Inosine
4.2. ASC-1 Alleviated Inflammation and Oxidative Stress in Hyperuricemia Rats
4.3. ASC-1 Suppressed Elevated XOD Activity in the Livers of Hyperuricemia Rats
4.4. ASC-1 Restores the Gut Microbiota and SCFA Production in Hyperuricemia Rats
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keebaugh, A.C.; Thomas, J.W. The Evolutionary Fate of the Genes Encoding the Purine Catabolic Enzymes in Hominoids, Birds, and Reptiles. Mol. Biol. Evol. 2010, 27, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.K.; Atkinson, K.; Karlson, E.W.; Willett, W.; Curhan, G. Purine-Rich Foods, Dairy and Protein Intake, and the Risk of Gout in Men. N. Engl. J. Med. 2004, 350, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Karantas, I.D.; Miliotou, A.N.; Siafaka, P.I. An Updated Review for Hyperuricemia and Gout Management; Special Focus on the Available Drug Delivery Systems and Clinical Trials. Curr. Med. Chem. 2023, 31. [Google Scholar] [CrossRef]
- Johnson, R.J.; Bakris, G.L.; Borghi, C.; Chonchol, M.B.; Feldman, D.; Lanaspa, M.A.; Merriman, T.R.; Moe, O.W.; Mount, D.B.; Sanchez Lozada, L.G.; et al. Hyperuricemia, Acute and Chronic Kidney Disease, Hypertension, and Cardiovascular Disease: Report of a Scientific Workshop Organized by the National Kidney Foundation. Am. J. Kidney Dis. 2018, 71, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Roddy, E.; Doherty, M. Gout. Epidemiology of Gout. Arthritis Res. Ther. 2010, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Fan, J.; Huang, Z.; Zhou, D.; Wang, X. Causal Relationship between Gut Microbiota and Gout: A Two-Sample Mendelian Randomization Study. Nutrients 2023, 15, 4260. [Google Scholar] [CrossRef]
- Ahn, S.O.; Ohtomo, S.; Kiyokawa, J.; Nakagawa, T.; Yamane, M.; Lee, K.J.; Kim, K.H.; Kim, B.H.; Tanaka, J.; Kawabe, Y.; et al. Stronger Uricosuric Effects of the Novel Selective URAT1 Inhibitor UR-1102 Lowered Plasma Urate in Tufted Capuchin Monkeys to a Greater Extent than Benzbromarone. J. Pharmacol. Exp. Ther. 2016, 357, 157–166. [Google Scholar] [CrossRef]
- Sundy, J.S.; Hershfield, M.S. Uricase and Other Novel Agents for the Management of Patients with Treatment-Failure Gout. Curr. Rheumatol. Rep. 2007, 9, 258–264. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Liao, W.; Huang, J.; Liu, Y.; Li, Z.; Tang, J. Gut Microbiota Remodeling: A Promising Therapeutic Strategy to Confront Hyperuricemia and Gout. Front. Cell. Infect. Microbiol. 2022, 12, 935723. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Y.; Zhong, H.; Chen, F.; Regenstein, J.; Hu, X.; Cai, L.; Feng, F. The Gut Microbiota as a Target to Control Hyperuricemia Pathogenesis: Potential Mechanisms and Therapeutic Strategies. Crit. Rev. Food Sci. Nutr. 2022, 62, 3979–3989. [Google Scholar] [CrossRef]
- Wu, Y.; Ye, Z.; Feng, P.; Li, R.; Chen, X.; Tian, X.; Han, R.; Kakade, A.; Liu, P.; Li, X. Limosilactobacillus fermentum JL-3 Isolated from “Jiangshui” Ameliorates Hyperuricemia by Degrading Uric Acid. Gut Microbes 2021, 13, 1897211. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, X.; Zhang, L.; Meng, F.; Zhou, L.; Pang, X.; Lu, Z.; Lu, Y. Lacticaseibacillus rhamnosus Fmb14 Prevents Purine Induced Hyperuricemia and Alleviate Renal Fibrosis through Gut-Kidney Axis. Pharmacol. Res. 2022, 182, 106350. [Google Scholar] [CrossRef]
- Yamada, N.; Saito-Iwamoto, C.; Nakamura, M.; Soeda, M.; Chiba, Y.; Kano, H.; Asami, Y. Lactobacillus gasseri PA-3 Uses the Purines IMP, Inosine and Hypoxanthine and Reduces Their Absorption in Rats. Microorganisms 2017, 5, 10. [Google Scholar] [CrossRef]
- Caliceti, C.; Calabria, D.; Roda, A.; Cicero, A. Fructose Intake, Serum Uric Acid, and Cardiometabolic Disorders: A Critical Review. Nutrients 2017, 9, 395. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Liu, Y.; Fan, Y.; Wu, D.; Meng, Y.; Qin, M. Agents for the Treatment of Gout: Current Advances and Future Perspectives. J. Med. Chem. 2023, 66, 14474–14493. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, L.; Zhang, Y.; Zeng, C. Recent Advances in Fructose Intake and Risk of Hyperuricemia. Biomed. Pharmacother. 2020, 131, 110795. [Google Scholar] [CrossRef]
- Jan, T.; Negi, R.; Sharma, B.; Kour, D.; Kumar, S.; Rai, A.K.; Rustagi, S.; Singh, S.; Sheikh, M.A.; Kumar, K.; et al. Diversity, Distribution and Role of Probiotics for Human Health: Current Research and Future Challenges. Biocatal. Agric. Biotechnol. 2023, 53, 102889. [Google Scholar] [CrossRef]
- Mazziotta, C.; Tognon, M.; Martini, F.; Torreggiani, E.; Rotondo, J.C. Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health. Cells 2023, 12, 184. [Google Scholar] [CrossRef]
- Alizadeh Behbahani, B.; Jooyandeh, H.; Hojjati, M.; Ghodsi Sheikhjan, M. Evaluation of Probiotic, Safety, and Anti-Pathogenic Properties of Levilactobacillus brevis HL6, and Its Potential Application as Bio-Preservatives in Peach Juice. LWT 2024, 191, 115601. [Google Scholar] [CrossRef]
- Du, Y.; Xu, W.; Wu, T.; Li, H.; Hu, X.; Chen, J. Enhancement of Growth, Survival, Immunity and Disease Resistance in Litopenaeus vannamei, by the Probiotic, Lactobacillus plantarum Ep-M17. Fish Shellfish Immunol. 2022, 129, 36–51. [Google Scholar] [CrossRef]
- Tang, H.; Zhou, T.; Jin, W.; Zong, S.; Mamtimin, T.; Salama, E.-S.; Jeon, B.-H.; Liu, P.; Han, H.; Li, X. Tumor-Targeting Engineered Probiotic Escherichia coli Nissle 1917 Inhibits Colorectal Tumorigenesis and Modulates Gut Microbiota Homeostasis in Mice. Life Sci. 2023, 324, 121709. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Peng, Z.; Zheng, W.; Yang, S.; Wu, M.; Liu, K.; Xiao, M.; Huang, T.; Xie, M.; Xiong, T. Probiotic-Fermented Tomato Alleviates High-Fat Diet-Induced Obesity in Mice: Insights from Microbiome and Metabolomics. Food Chem. 2024, 436, 137719. [Google Scholar] [CrossRef]
- Rudzki, L.; Ostrowska, L.; Pawlak, D.; Małus, A.; Pawlak, K.; Waszkiewicz, N.; Szulc, A. Probiotic Lactobacillus plantarum 299v Decreases Kynurenine Concentration and Improves Cognitive Functions in Patients with Major Depression: A Double-Blind, Randomized, Placebo Controlled Study. Psychoneuroendocrinology 2019, 100, 213–222. [Google Scholar] [CrossRef]
- Zarezadeh, M.; Musazadeh, V.; Ghalichi, F.; Kavyani, Z.; Nasernia, R.; Parang, M.; Jamilian, P.; Jamilian, P.; Fakhr, L.; Ostadrahimi, A.; et al. Effects of Probiotics Supplementation on Blood Pressure: An Umbrella Meta-Analysis of Randomized Controlled Trials. Nutr. Metab. Cardiovasc. Dis. 2023, 33, 275–286. [Google Scholar] [CrossRef]
- Nelson, K.; Wysocki, J. Benefits of Probiotic Consumption on Chronic Kidney Disease. J. Ren. Nutr. 2020, 30, e35–e36. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Y.; Sailike, J.; Sun, X.; Abuduwaili, N.; Tuoliuhan, H.; Yusufu, M.; Nabi, X. Fourteen Composite Probiotics Alleviate Type 2 Diabetes through Modulating Gut Microbiota and Modifying M1/M2 Phenotype Macrophage in Db/Db Mice. Pharmacol. Res. 2020, 161, 105150. [Google Scholar] [CrossRef]
- Chai, Z.; Yan, Y.; Zan, S.; Meng, X.; Zhang, F. Probiotic-Fermented Blueberry Pomace Alleviates Obesity and Hyperlipidemia in High-Fat Diet C57BL/6J Mice. Food Res. Int. 2022, 157, 111396. [Google Scholar] [CrossRef] [PubMed]
- Maky, M.A.; Ishibashi, N.; Nakayama, J.; Zendo, T. Characterization of the Biosynthetic Gene Cluster of Enterocin F4-9, a Glycosylated Bacteriocin. Microorganisms 2021, 9, 2276. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.M.; Zhang, Y. Effects of Lactic Acid Bacteria on Nitrite Degradation during Pickle Fermentation. Adv. Mater. Res. 2013, 781–784, 1656–1660. [Google Scholar] [CrossRef]
- Zhang, X.; Han, J.; Zheng, X.; Yan, J.; Chen, X.; Zhou, Q.; Zhao, X.; Gu, Q.; Li, P. Use of Lactiplantibacillus plantarum ZJ316 as a Starter Culture for Nitrite Degradation, Foodborne Pathogens Inhibition and Microbial Community Modulation in Pickled Mustard Fermentation. Food Chem. X 2022, 14, 100344. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, S.; Liu, W.; Chang, L.; Zhu, X.; Mu, G.; Qian, F. Probiotic Properties of Lactobacillus paraplantarum LS-5 and Its Effect on Antioxidant Activity of Fermented Sauerkraut. Food Biosci. 2023, 52, 102489. [Google Scholar] [CrossRef]
- Xiao, Y.; Zhang, C.; Zeng, X.; Yuan, Z. Microecological Treatment of Hyperuricemia Using Lactobacillus from Pickles. BMC Microbiol. 2020, 20, 195. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yang, D.; Mei, L.; Yuan, L.; Xie, A.; Yuan, J. Screening and Characterization of Purine Nucleoside Degrading Lactic Acid Bacteria Isolated from Chinese Sauerkraut and Evaluation of the Serum Uric Acid Lowering Effect in Hyperuricemic Rats. PLoS ONE 2014, 9, e105577. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, F.; Wu, Q.; Jin, L.; Wang, Q. The identification of Five Bacillus Species Used in Environment Microbe Agentia with Realtime PCR. Chin. J. Health Lab. Technol. 2010, 20, 246–248+296. [Google Scholar]
- Jeong, J.; Mun, S.; Oh, Y.; Cho, C.-S.; Yun, K.; Ahn, Y.; Chung, W.-H.; Lim, M.Y.; Lee, K.E.; Hwang, T.S.; et al. A qRT-PCR Method Capable of Quantifying Specific Microorganisms Compared to NGS-Based Metagenome Profiling Data. Microorganisms 2022, 10, 324. [Google Scholar] [CrossRef] [PubMed]
- Arya, G.; Petronella, N.; Crosthwait, J.; Carrillo, C.D.; Shwed, P.S. Draft Genome Sequence of Bacillus megaterium Type Strain ATCC 14581. Genome Announc. 2014, 2, e01124-14. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.-Y.; Chien, Y.-J.; Lin, Y.-H.; Lin, Y.-H.; Chan, S.-T.; Hu, W.-C.; Wu, H.-F.; Chiang, C.-F.; Hsu, C.-L. Supplementation of Lactobacillus plantarum (TCI227) Prevented Potassium-Oxonate-Induced Hyperuricemia in Rats. Nutrients 2022, 14, 4832. [Google Scholar] [CrossRef] [PubMed]
- Griebsch, A.; Zöllner, N. Effect of Ribomononucleotides Given Orally on Uric Acid Production in Man. In Purine Metabolism in Man; Sperling, O., De Vries, A., Wyngaarden, J.B., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 1974; Volume 41, pp. 443–449. ISBN 978-1-4757-1435-7. [Google Scholar]
- Li, S.; Liu, X.; Jia, X.; Fang, M.; Yang, Q.; Gong, Z. Assessment of the Temporal Trend and Daily Profiles of the Dietary Purine Intake among Chinese Residents during 2014 to 2021. Front. Nutr. 2023, 10, 1259053. [Google Scholar] [CrossRef] [PubMed]
- Koguchi, T. Modification of Dietary Habits for Prevention of Gout in Japanese People: Gout and the Japanese Diet. Am. J. Health Res. 2021, 9, 117. [Google Scholar] [CrossRef]
- Yamada, N.; Saito, C.; Murayama-Chiba, Y.; Kano, H.; Asami, Y.; Itoh, H. Lactobacillus gasseri PA-3 Utilizes the Purines GMP and Guanosine and Decreases Their Absorption in Rats. Nucleosides Nucleotides Nucleic Acids 2018, 37, 307–315. [Google Scholar] [CrossRef]
- Zhao, X.; Peng, F.; Liu, Z.; Peng, Z.; Guan, Q.; Cai, P.; Xiong, S.; Yu, Q.; Xie, M.; Xiong, T. Lactic Acid Bacteria with Anti-Hyperuricemia Ability: Screening in Vitro and Evaluating in Mice. Food Biosci. 2023, 52, 102411. [Google Scholar] [CrossRef]
- Cao, X.; Cai, J.; Zhang, Y.; Liu, C.; Song, M.; Xu, Q.; Liu, Y.; Yan, H. Biodegradation of Uric Acid by Bacillus Paramycoides-YC02. Microorganisms 2023, 11, 1989. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Liu, Q.; Hao, H.; Bu, Y.; Tian, X.; Wang, T.; Yi, H. Lactobacillus paracasei X11 Ameliorates Hyperuricemia and Modulates Gut Microbiota in Mice. Front. Immunol. 2022, 13, 940228. [Google Scholar] [CrossRef] [PubMed]
- Nganou-Gnindjio, C.N.; Tsasse Fokou, J.F.; Nkeck, J.R.; Pieme, C.A.; Doualla, M.S. Investigation of Malondialdehyde, Catalase, Glutathione Peroxydase and FRAP as Biomarker for Oxidative Stress Linked to Hyperuricemia in a Sub-Saharan Population with Heart Failure. Arch. Cardiovasc. Dis. Suppl. 2023, 15, 122. [Google Scholar] [CrossRef]
- Nascimento, G.V.R.D.; Balbi, A.L.; Ponce, D.; Abrão, J.M.G. Early Initiation of Dialysis: Mortality and Renal Function Recovery in Acute Kidney Injury Patient. J. Bras. Nefrol. 2012, 34, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.-Y.; Ning, X.-Y.; Chen, Y.-Q.; Han, S.-J.; Chi, P.; Zhu, S.-N.; Yue, Y. Predictive Value of Serum Creatinine, Blood Urea Nitrogen, Uric Acid, and β 2-Microglobulin in the Evaluation of Acute Kidney Injury after Orthotopic Liver Transplantation. Chin. Med. J. 2018, 131, 1059–1066. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhu, J.; Lin, G.; Gao, K.; Yu, Y.; Chen, S.; Chen, L.; Chen, Z.; Li, L. Probiotic Effects of Lacticaseibacillus rhamnosus 1155 and Limosilactobacillus Fermentum 2644 on Hyperuricemic Rats. Front. Nutr. 2022, 9, 993951. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wang, T.; Liu, Y.; Zhou, W.; Hao, H.; Liu, Q.; Yin, B.; Yi, H. Lactobacillus Fermentum F40-4 Ameliorates Hyperuricemia by Modulating the Gut Microbiota and Alleviating Inflammation in Mice. Food Funct. 2023, 14, 3259–3268. [Google Scholar] [CrossRef]
- Yang, B.; Xin, M.; Liang, S.; Xu, X.; Cai, T.; Dong, L.; Wang, C.; Wang, M.; Cui, Y.; Song, X.; et al. New Insight into the Management of Renal Excretion and Hyperuricemia: Potential Therapeutic Strategies with Natural Bioactive Compounds. Front. Pharmacol. 2022, 13, 1026246. [Google Scholar] [CrossRef]
- Shi, X.; Zhao, T.; Da Silva-Júnior, E.F.; Zhang, J.; Xu, S.; Gao, S.; Liu, X.; Zhan, P. Novel Urate Transporter 1 (URAT1) Inhibitors: A Review of Recent Patent Literature (2020–Present). Expert Opin. Ther. Pat. 2022, 32, 1175–1184. [Google Scholar] [CrossRef]
- Wang, H.; Mei, L.; Deng, Y.; Liu, Y.; Wei, X.; Liu, M.; Zhou, J.; Ma, H.; Zheng, P.; Yuan, J.; et al. Lactobacillus brevis DM9218 Ameliorates Fructose-Induced Hyperuricemia through Inosine Degradation and Manipulation of Intestinal Dysbiosis. Nutrition 2019, 62, 63–73. [Google Scholar] [CrossRef] [PubMed]
- De Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut Microbiome and Health: Mechanistic Insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ye, J. Commentary: Gut Microbiota Reduce the Risk of Hyperuricemia and Gout in the Human Body. Acta Pharm. Sin. B 2024, 14, 433–435. [Google Scholar] [CrossRef] [PubMed]
- Van Der Hee, B.; Wells, J.M. Microbial Regulation of Host Physiology by Short-Chain Fatty Acids. Trends Microbiol. 2021, 29, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Letchumanan, G.; Abdullah, N.; Marlini, M.; Baharom, N.; Lawley, B.; Omar, M.R.; Mohideen, F.B.S.; Addnan, F.H.; Nur Fariha, M.M.; Ismail, Z.; et al. Gut Microbiota Composition in Prediabetes and Newly Diagnosed Type 2 Diabetes: A Systematic Review of Observational Studies. Front. Cell. Infect. Microbiol. 2022, 12, 943427. [Google Scholar] [CrossRef]
- Bonnechère, B.; Amin, N.; Van Duijn, C. What Are the Key Gut Microbiota Involved in Neurological Diseases? A Systematic Review. Int. J. Mol. Sci. 2022, 23, 13665. [Google Scholar] [CrossRef] [PubMed]
- Takagi, T.; Inoue, R.; Oshima, A.; Sakazume, H.; Ogawa, K.; Tominaga, T.; Mihara, Y.; Sugaya, T.; Mizushima, K.; Uchiyama, K.; et al. Typing of the Gut Microbiota Community in Japanese Subjects. Microorganisms 2022, 10, 664. [Google Scholar] [CrossRef]
- Rodrigues, V.F.; Elias-Oliveira, J.; Pereira, Í.S.; Pereira, J.A.; Barbosa, S.C.; Machado, M.S.G.; Carlos, D. Akkermansia muciniphila and Gut Immune System: A Good Friendship That Attenuates Inflammatory Bowel Disease, Obesity, and Diabetes. Front. Immunol. 2022, 13, 934695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Li, Q.; Cheng, L.; Buch, H.; Zhang, F. Akkermansia muciniphila Is a Promising Probiotic. Microb. Biotechnol. 2019, 12, 1109–1125. [Google Scholar] [CrossRef]
- Zhai, Q.; Feng, S. A next Generation Probiotic, Akkermansia muciniphila. Crit. Rev. Food Sci. Nutr. 2019, 59, 3227–3236. [Google Scholar] [CrossRef]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; De Vos, W.M. Akkermansia muciniphila: Paradigm for next-Generation Beneficial Microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, W.; Bi, S.; Fang, Z.; Iddrisu, L.; Deng, Q.; Sun, L.; Gooneratne, R. Priestia megaterium ASC-1 Isolated from Pickled Cabbage Ameliorates Hyperuricemia by Degrading Uric Acid in Rats. Microorganisms 2024, 12, 832. https://doi.org/10.3390/microorganisms12040832
Zhu W, Bi S, Fang Z, Iddrisu L, Deng Q, Sun L, Gooneratne R. Priestia megaterium ASC-1 Isolated from Pickled Cabbage Ameliorates Hyperuricemia by Degrading Uric Acid in Rats. Microorganisms. 2024; 12(4):832. https://doi.org/10.3390/microorganisms12040832
Chicago/Turabian StyleZhu, Wenjuan, Siyuan Bi, Zhijia Fang, Lukman Iddrisu, Qi Deng, Lijun Sun, and Ravi Gooneratne. 2024. "Priestia megaterium ASC-1 Isolated from Pickled Cabbage Ameliorates Hyperuricemia by Degrading Uric Acid in Rats" Microorganisms 12, no. 4: 832. https://doi.org/10.3390/microorganisms12040832