
Type I Cystatin Derived from Cysticercus pisiformis—Stefins, Suppresses LPS-Mediated Inflammatory Response in RAW264.7 Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Production of Recombinant Cpstefin and Removal of Endotoxin
2.2. Cell Culture and Treatments
2.3. Cell Viability Assay
2.4. Evaluation of NO by Griess Test
2.5. ELISA Test Cytokine Secretion Levels
2.6. Detection of Transcript Levels of Cytokines by Reverse Transcription Real-Time PCR
2.7. Statistical Analysis
3. Results and Analysis
3.1. Expression and Purification of rCpStefin Proteins
3.2. CpStefin Has No Effect on LPS-Induced Activity in RAW264.7 Cells
3.3. rCpStefin Inhibits LPS-Induced NO Production in RAW264.7 Cells
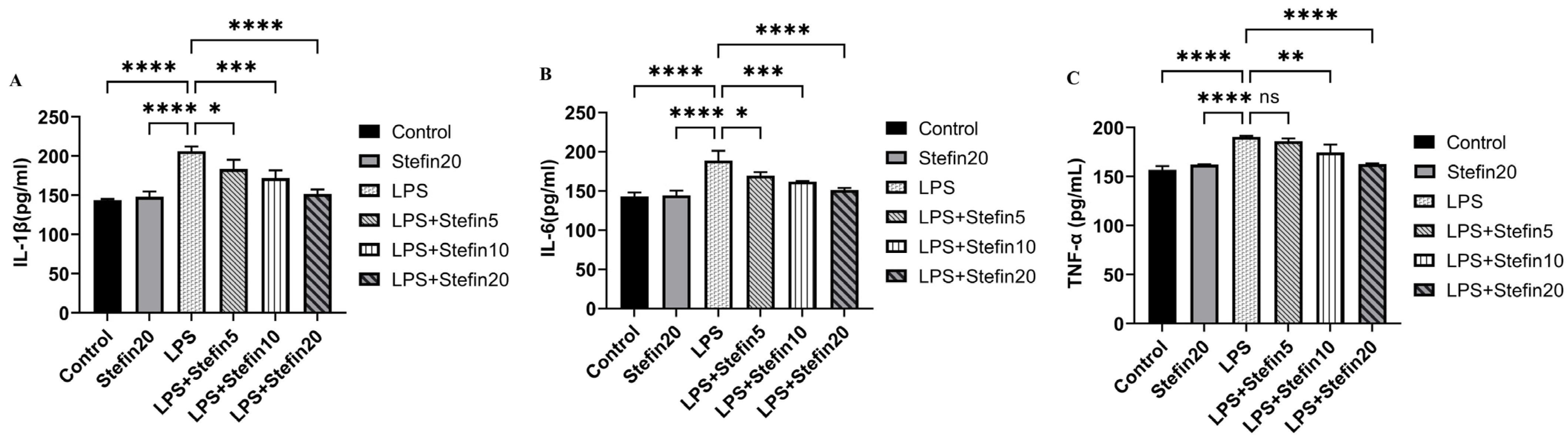
3.4. rCpStefin Inhibits LPS-Induced Cytokine Production by RAW264.7
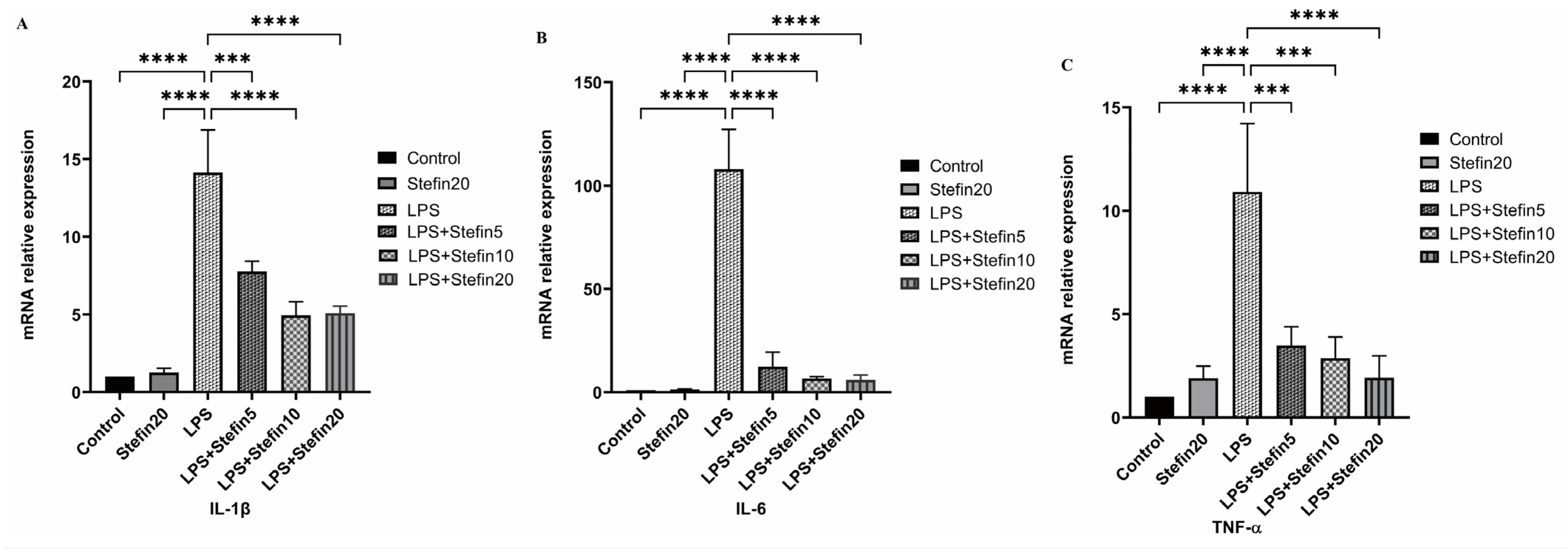
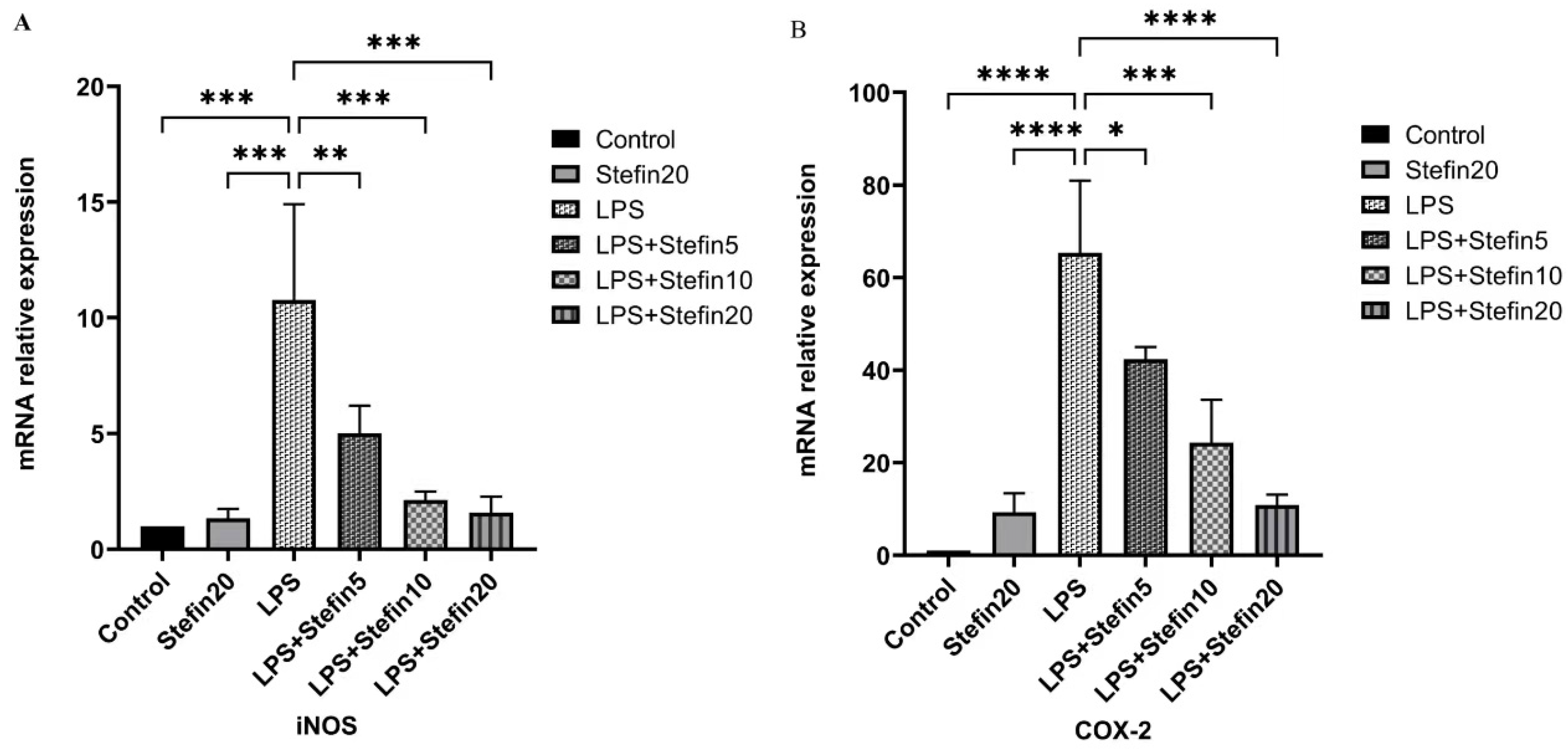
3.5. rCpStefin Inhibits mRNA Levels of Cytokine Production by RAW264.7 under LPS Induction
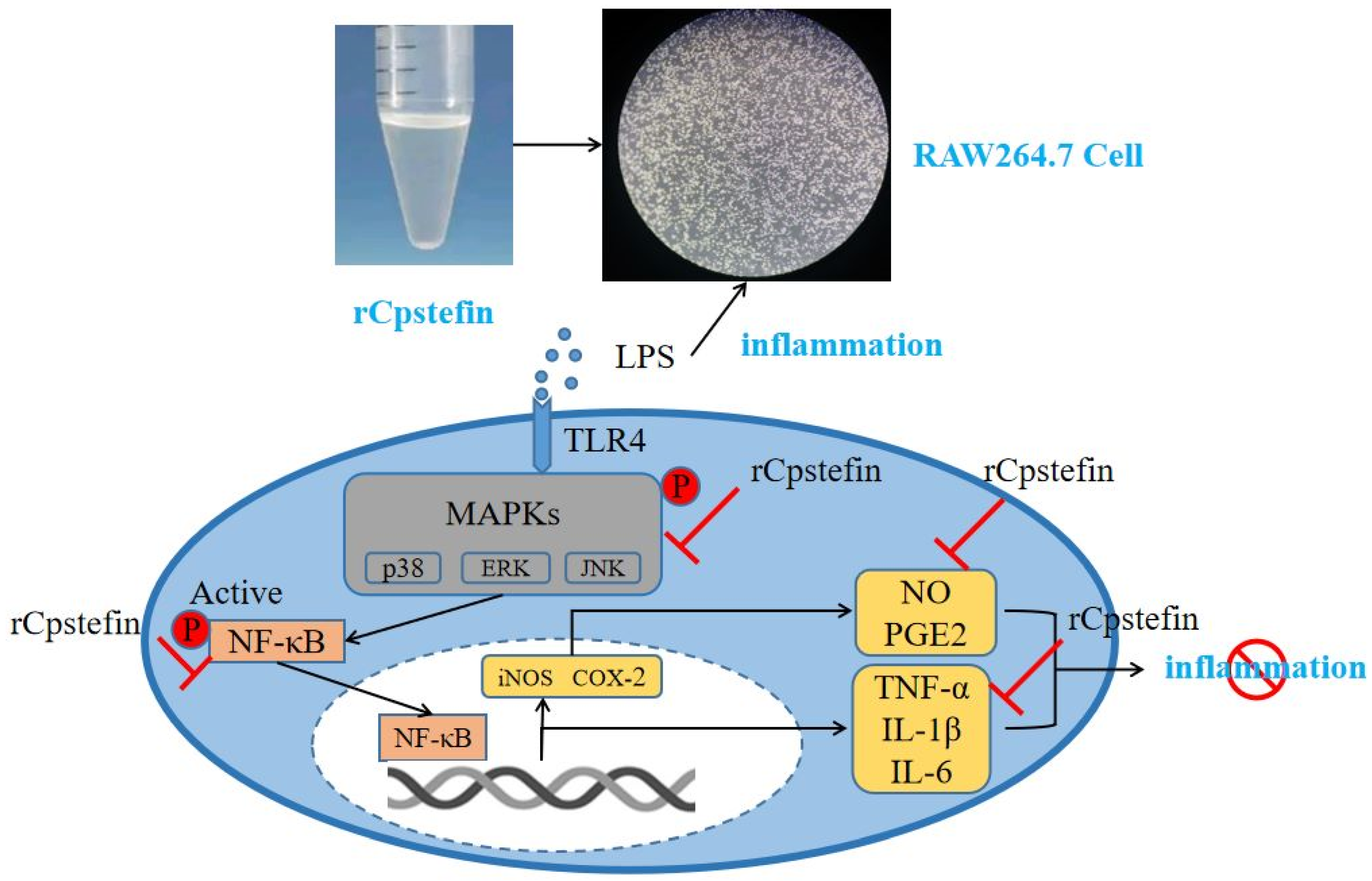
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Toral-Bastida, E.; Garza-Rodriguez, A.; Jimenez-Gonzalez, D.E.; Garcia-Cortes, R.; Avila-Ramirez, G.; Maravilla, P.; Flisser, A. Development of Taenia pisiformis in golden hamster (Mesocricetus auratus). Parasites Vectors 2011, 4, 147. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wang, L.; Liu, T.; Li, Y.; Zhang, S.; Li, H.; Luo, X. Identification and Expression Profiling of Circulating MicroRNAs in Serum of Cysticercus pisiformis—Infected Rabbits. Genes 2021, 12, 1591. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Guo, Z.; Duo, H.; Fu, Y.; Peng, M.; Shen, X.; Nonaka, N. Survey on helminths in the small intestine of wild foxes in Qinghai, China. J. Vet. Med. Sci. 2013, 75, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S. Comparative Transcriptomic Analysis of the Larval and Adult Stages of Taenia pisiformis. Genes 2019, 10, 507. [Google Scholar] [CrossRef] [PubMed]
- Samorek-Pieróg, M.; Karamon, J.; Brzana, A.; Bilska-Zając, E.; Zdybel, J.; Cencek, T. Molecular Confirmation of Massive Taenia pisiformis Cysticercosis in One Rabbit in Poland. Pathogens 2021, 10, 1029. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.H.M.; Lee, S.; Huh, J.-E.; Perumalsamy, H.; Balusamy, S.R.; Kim, Y.-J. Fermentation of Moringa oleifera Lam. using Bifidobacterium animalis subsp. lactis enhances the anti-inflammatory effect in RAW 264.7 macrophages. J. Funct. Foods 2023, 109, 105752. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New Adventures of an Old Flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef]
- Shou, J.; Kong, X.; Wang, X.; Tang, Y.; Wang, C.; Wang, M.; Zhang, L.; Liu, Y.; Fei, C.; Xue, F.; et al. Tizoxanide Inhibits Inflammation in LPS-Activated RAW264.7 Macrophages via the Suppression of NF-kappaB and MAPK Activation. Inflammation 2019, 42, 1336–1349. [Google Scholar] [CrossRef]
- Li, W.X. Canonical and non-canonical JAK–STAT signaling. Trends Cell Biol. 2008, 18, 545–551. [Google Scholar] [CrossRef]
- Lee, H.H.; Jang, E.; Kang, S.Y.; Shin, J.S.; Han, H.S.; Kim, T.W.; Lee, D.H.; Lee, J.H.; Jang, D.S.; Lee, K.T. Anti-inflammatory potential of Patrineolignan B isolated from Patrinia scabra in LPS-stimulated macrophages via inhibition of NF-kappaB, AP-1, and JAK/STAT pathways. Int. Immunopharmacol. 2020, 86, 106726. [Google Scholar] [CrossRef]
- Zhao, X.L.; Yu, L.; Zhang, S.D.; Ping, K.; Ni, H.Y.; Qin, X.Y.; Zhao, C.J.; Wang, W.; Efferth, T.; Fu, Y.J. Cryptochlorogenic acid attenuates LPS-induced inflammatory response and oxidative stress via upregulation of the Nrf2/HO-1 signaling pathway in RAW 264.7 macrophages. Int. Immunopharmacol. 2020, 83, 106436. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Lee, E.K.; Gong, S.Y.; Sohng, J.K.; Kang, Y.J.; Jung, H.J. Sparassis crispa exerts anti-inflammatory activity via suppression of TLR-mediated NF-kappaB and MAPK signaling pathways in LPS-induced RAW264.7 macrophage cells. J. Ethnopharmacol. 2019, 231, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Strandwitz, P. Neurotransmitter modulation by the gut microbiota. Brain Res. 2018, 1693, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Bach, J.F. The hygiene hypothesis in autoimmunity: The role of pathogens and commensals. Nat. Rev. Immunol. 2018, 18, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Strachan, D.P. Family size, infection and atopy: The first decade of the “hygiene hypothesis”. Thorax 2000, 55 (Suppl. 1), S2–S10. [Google Scholar] [CrossRef] [PubMed]
- Massicot, J.G.; Cohen, S.G. Epidemiologic and socioeconomic aspects of allergic diseases. J. Allergy Clin. Immunol. 1986, 78 Pt 2, 954–958. [Google Scholar] [CrossRef] [PubMed]
- Santiago, H.C.; Bennuru, S.; Boyd, A.; Eberhard, M.; Nutman, T.B. Structural and immunologic cross-reactivity among filarial and mite tropomyosin: Implications for the hygiene hypothesis. J. Allergy Clin. Immunol. 2011, 127, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Su, Z.; Piccirillo, C.; Stevenson, M.M. Impairment of dendritic cell function by excretory-secretory products: A potential mechanism for nematode-induced immunosuppression. Eur. J. Immunol. 2007, 37, 1887–1904. [Google Scholar] [CrossRef]
- Langelaar, M.; Aranzamendi, C.; Franssen, F.; Van Der Giessen, J.; Rutten, V.; van der Ley, P.; Pinelli, E. Suppression of dendritic cell maturation by Trichinella spiralis excretory/secretory products. Parasite Immunol. 2009, 31, 641–645. [Google Scholar] [CrossRef]
- Falcón, C.; Carranza, F.; Martínez, F.F.; Knubel, C.P.; Masih, D.T.; Motrán, C.C.; Cervi, L. Excretory-secretory products (ESP) from Fasciola hepatica induce tolerogenic properties in myeloid dendritic cells. Vet. Immunol. Immunopathol. 2010, 137, 36–46. [Google Scholar] [CrossRef]
- Weinstock, J.V.; Elliott, D.E. Helminths and the IBD hygiene hypothesis. Inflamm. Bowel Dis. 2009, 15, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.J. The potential impact of early exposures to geohelminth infections on the development of atopy. Clin. Rev. Allergy Immunol. 2004, 26, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Smits, H.H.; Everts, B.; Hartgers, F.C.; Yazdanbakhsh, M. Chronic helminth infections protect against allergic diseases by active regulatory processes. Curr. Allergy Asthma Rep. 2010, 10, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Steinfelder, S.; O’Regan, N.L.; Hartmann, S. Diplomatic Assistance: Can Helminth-Modulated Macrophages Act as Treatment for Inflammatory Disease? PLoS Pathog. 2016, 12, e1005480. [Google Scholar] [CrossRef] [PubMed]
- Chantree, P.; Tarasuk, M.; Prathaphan, P.; Ruangtong, J.; Jamklang, M.; Chumkiew, S.; Martviset, P. Type I Cystatin Derived from Fasciola gigantica Suppresses Macrophage-Mediated Inflammatory Responses. Pathogens 2023, 12, 395. [Google Scholar] [CrossRef] [PubMed]
- Khatri, V.; Chauhan, N.; Kalyanasundaram, R. Parasite Cystatin: Immunomodulatory Molecule with Therapeutic Activity against Immune Mediated Disorders. Pathogens 2020, 9, 431. [Google Scholar] [CrossRef] [PubMed]
- Keppler, D. Towards novel anti-cancer strategies based on cystatin function. Cancer Lett. 2006, 235, 159–176. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, T.; Rausch, S.; Steinfelder, S.; Klotz, C.; Hepworth, M.R.; Kühl, A.A.; Burda, P.C.; Lucius, R.; Hartmann, S. A novel regulatory macrophage induced by a helminth molecule instructs IL-10 in CD4+ T cells and protects against mucosal inflammation. J. Immunol. 2015, 194, 1555–1564. [Google Scholar] [CrossRef] [PubMed]
- Khatri, V.; Amdare, N.; Tarnekar, A.; Goswami, K.; Reddy, M.V. Brugia malayi cystatin therapeutically ameliorates dextran sulfate sodium-induced colitis in mice. J. Dig. Dis. 2015, 16, 585–594. [Google Scholar] [CrossRef]
- Ji, P.; Hu, H.; Yang, X.; Wei, X.; Zhu, C.; Liu, J. AcCystatin, an immunoregulatory molecule from Angiostrongylus cantonensis, ameliorates the asthmatic response in an aluminium hydroxide/ovalbumin-induced rat model of asthma. Parasitol. Res. 2015, 114, 613–624. [Google Scholar] [CrossRef]
- Malyala, P.; Singh, M. Endotoxin limits in formulations for preclinical research. J. Pharm. Sci. 2008, 97, 2041–2044. [Google Scholar] [CrossRef] [PubMed]
- Reyes, J.L.; Lopes, F.; Leung, G.; Jayme, T.S.; Matisz, C.E.; Shute, A. Macrophages treated with antigen from the tapeworm Hymenolepis diminuta condition CD25+ T cells to suppress colitis. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 5676–5689. [Google Scholar] [CrossRef] [PubMed]
- Rolot, M.; Dewals, B.G. Macrophage Activation and Functions during Helminth Infection: Recent Advances from the Laboratory Mouse. J. Immunol. Res. 2018, 2018, 2790627. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Hu, Y.; Zhao, Y.; Cheng, M.; Qin, H.; Cheng, T.; Wang, Q.; Peng, X.; Zhang, X. Curcumin Attenuates Titanium Particle-Induced Inflammation by Regulating Macrophage Polarization In Vitro and In Vivo. Front. Immunol. 2017, 8, 55. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.C.; Yang, G.; Lai, C.S.; Weerawatanakorn, M.; Pan, M.H. Chemopreventive Effects of Phytochemicals and Medicines on M1/M2 Polarized Macrophage Role in Inflammation-Related Diseases. Int. J. Mol. Sci. 2018, 19, 2208. [Google Scholar] [CrossRef]
- Li, Y.; Xie, Z.; Wang, Y.; Hu, H. Macrophage M1/M2 polarization in patients with pregnancy-induced hypertension. Can. J. Physiol. Pharmacol. 2018, 96, 922–928. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.Q.; Li, A.; Yang, X.; Xiao, X.; Hu, R.; Wang, T.W.; Dou, X.Y.; Yang, D.J.; Dong, Z. Paeoniflorin exerts neuroprotective effects by modulating the M1/M2 subset polarization of microglia/macrophages in the hippocampal CA1 region of vascular dementia rats via cannabinoid receptor 2. Chin. Med. 2018, 13, 14. [Google Scholar] [CrossRef]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef]
- Xie, H.; Chu, L.; Wu, L.Q.; Fan, X.Y.; Wang, P.; Ma, S.Y. Polarization of bone marrow-derived macrophages induced by recombinant Trichinella spiralis cysteine protease inhibitors in vitro. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 2020, 32, 181–186. [Google Scholar]
- Wang, L.; Liu, T.; Chen, G.; Li, Y.; Zhang, S.; Mao, L.; Liang, P.; Fasihi Harandi, M.; Li, T.; Luo, X. Exosomal microRNA let-7-5p from Taenia pisiformis Cysticercus Prompted Macrophage to M2 Polarization through Inhibiting the Expression of C/EBP δ. Microorganisms 2021, 9, 1403. [Google Scholar] [CrossRef]
- Gwon, Y.G.; Rod-In, W.; Lee, H.J.; Lee, S.M.; Shin, I.S.; Park, W.J. Inhibitory effects of Oncorhynchus mykiss lipids in LPS-induced RAW264.7 cells via suppression of NF-κB and MAPK pathways. Fish Shellfish Immunol. 2024, 144, 109266. [Google Scholar] [CrossRef] [PubMed]
- González-Rubio, G.; Sellers-Moya, Á.; Martín, H.; Molina, M. A walk-through MAPK structure and functionality with the 30-year-old yeast MAPK Slt2. Int. Microbiol. 2021, 24, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Kim, S.Y.; Kim, K.M.; Mony, T.J.; Bae, H.J.; Kim, M.S. Fermented Sprouts of Codonopsis lanceolata Suppress LPS-Induced Inflammatory Responses by Inhibiting NF-κB Signaling Pathway in RAW 264.7 Macrophages and CD1 Mice. Pharmaceutics 2023, 15, 1793. [Google Scholar] [CrossRef] [PubMed]
- Beckhauser, T.F.; Francis-Oliveira, J.; De Pasquale, R. Reactive Oxygen Species: Physiological and Physiopathological Effects on Synaptic Plasticity. J. Exp. Neurosci. 2016, 10 (Suppl. 1), 23–48. [Google Scholar] [CrossRef] [PubMed]
- Coulthard, L.R.; White, D.E.; Jones, D.L.; McDermott, M.F.; Burchill, S.A. p38MAPK: Stress responses from molecular mechanisms to therapeutics. Trends Mol. Med. 2009, 15, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Bubici, C.; Papa, S. JNK signalling in cancer: In need of new, smarter therapeutic targets. Br. J. Pharmacol. 2014, 171, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, L.; Huang, G.; Zhou, T.; Zhang, X.; Leng, X.; Chen, Z.; Lin, J. Embelia Laeta aqueous extract suppresses acute inflammation via decreasing COX-2/iNOS expression and inhibiting NF-κB pathway. J. Ethnopharmacol. 2021, 281, 114575. [Google Scholar] [CrossRef] [PubMed]
- Nahrevanian, H. Involvement of nitric oxide and its up/down stream molecules in the immunity against parasitic infections. Braz. J. Infect. Dis. 2009, 13, 440–448. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef]
- Elzahhar, P.A.; Orioli, R.; Hassan, N.W.; Gobbi, S.; Belluti, F.; Labib, H.F. Chromone-based small molecules for multistep shutdown of arachidonate pathway: Simultaneous inhibition of COX-2, 15-LOX and mPGES-1 enzymes. Eur. J. Med. Chem. 2024, 266, 116138. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Pu, G.; Wang, L.; Li, Y.; Liu, T.; Li, H.; Zhang, S.; Wang, X.; Liu, X.; Luo, X. Cysticercus pisiformis-derived novel-miR1 targets TLR2 to inhibit the immune response in rabbits. Front. Immunol. 2023, 14, 1201455. [Google Scholar] [CrossRef] [PubMed]
- Ghelani, H.; Khursheed, M.; Adrian, T.E.; Jan, R.K. Anti-Inflammatory Effects of Compounds from Echinoderms. Mar. Drugs 2022, 20, 693. [Google Scholar] [CrossRef] [PubMed]
- Ożarowski, M.; Karpiński, T.M. Extracts and Flavonoids of Passiflora Species as Promising Anti-inflammatory and Antioxidant Substances. Curr. Pharm. Des. 2021, 27, 2582–2604. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, H.; Hu, B.; Yang, L.; Wang, P.; Wang, F.; Meng, X. Coptisine from Coptis chinensis inhibits production of inflammatory mediators in lipopolysaccharide-stimulated RAW 264.7 murine macrophage cells. Eur. J. Pharmacol. 2016, 780, 106–114. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence from 5′ to 3′ | Product Size (bp) |
|---|---|---|
| IL-1β | FW: TGC CAC CCT TTT GAC AGT GAT G | 138 |
| RV: TGA TGT GCT GCT GCG AGA TT | ||
| IL-6 | FW: CCC CAA TTT CCA ATG CTC TCC | 141 |
| RV: CGC ACT AGG TTT GCC GAG TA | ||
| TNF-α | FW: CCC TCA CAC TCA GAT CAT CTT CT | 61 |
| RV: GCT ACG ACG TGG GCT ACA G | ||
| COX-2 | FW: TGT GAC TGT ACC CGG ACT GG | 233 |
| RV: TGC ACA TTG TAA GTA GGT GGA C | ||
| iNOS | FW: CCC TTC CGA AGT TTC TGG CAG CAG | 497 |
| RV: GGC TGT CAG AGC CTC GTG GCT TTG G | ||
| GAPDH | FW: CGA CTT CAA CAG CGA CAC TCA C | 119 |
| RV: CCC TGT TGC TGT AGC CAA ATT C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.; Li, J.; Zhang, L.; Zhao, N.; Sun, X.; Wang, Z. Type I Cystatin Derived from Cysticercus pisiformis—Stefins, Suppresses LPS-Mediated Inflammatory Response in RAW264.7 Cells. Microorganisms 2024, 12, 850. https://doi.org/10.3390/microorganisms12050850
Yang Q, Li J, Zhang L, Zhao N, Sun X, Wang Z. Type I Cystatin Derived from Cysticercus pisiformis—Stefins, Suppresses LPS-Mediated Inflammatory Response in RAW264.7 Cells. Microorganisms. 2024; 12(5):850. https://doi.org/10.3390/microorganisms12050850
Chicago/Turabian StyleYang, Qianqian, Jia Li, Lilan Zhang, Ningning Zhao, Xiaolin Sun, and Zexiang Wang. 2024. "Type I Cystatin Derived from Cysticercus pisiformis—Stefins, Suppresses LPS-Mediated Inflammatory Response in RAW264.7 Cells" Microorganisms 12, no. 5: 850. https://doi.org/10.3390/microorganisms12050850