Towards an Ecological Understanding of Dinoflagellate Cyst Functions

Abstract
:1. Introduction
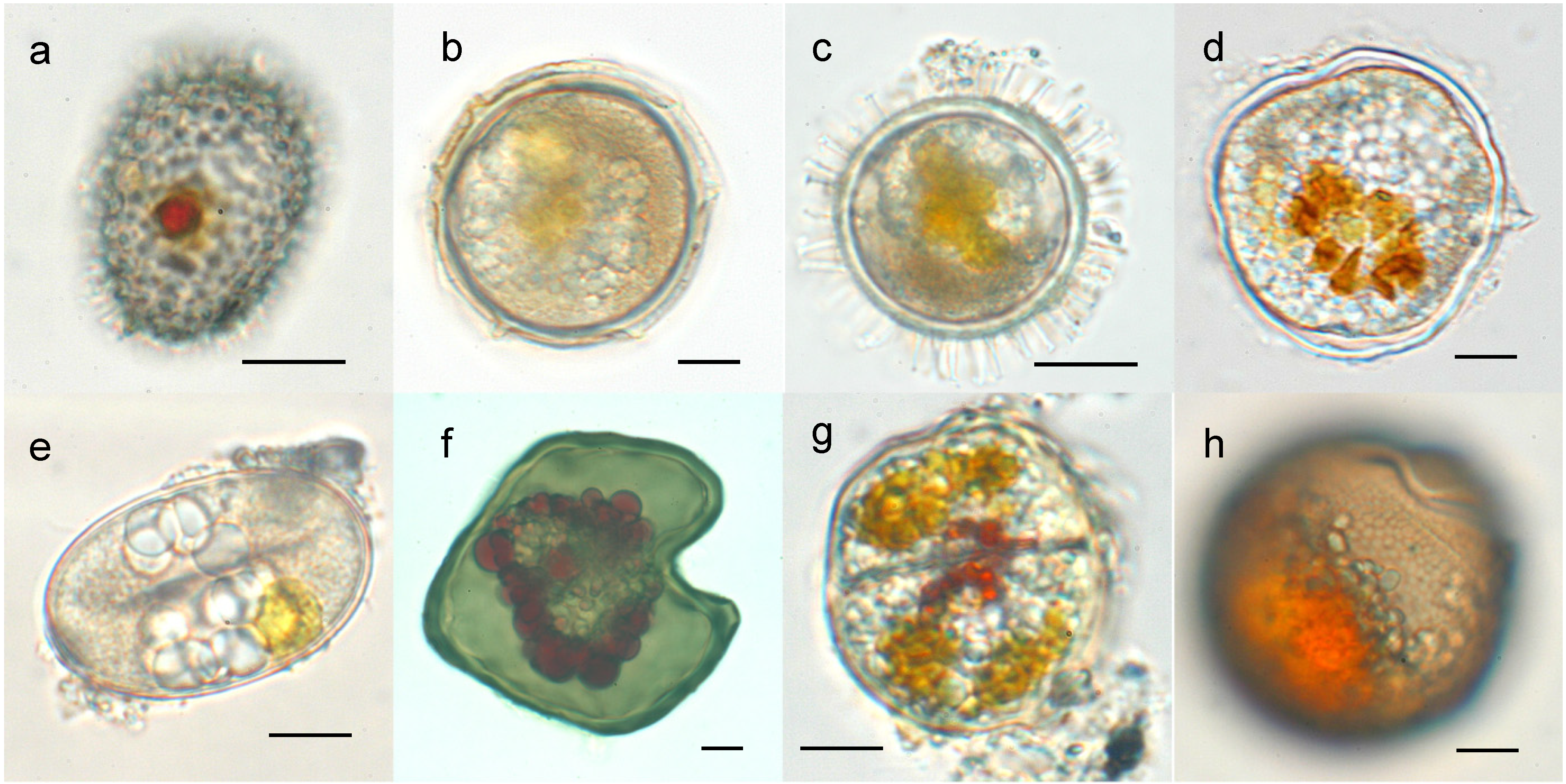
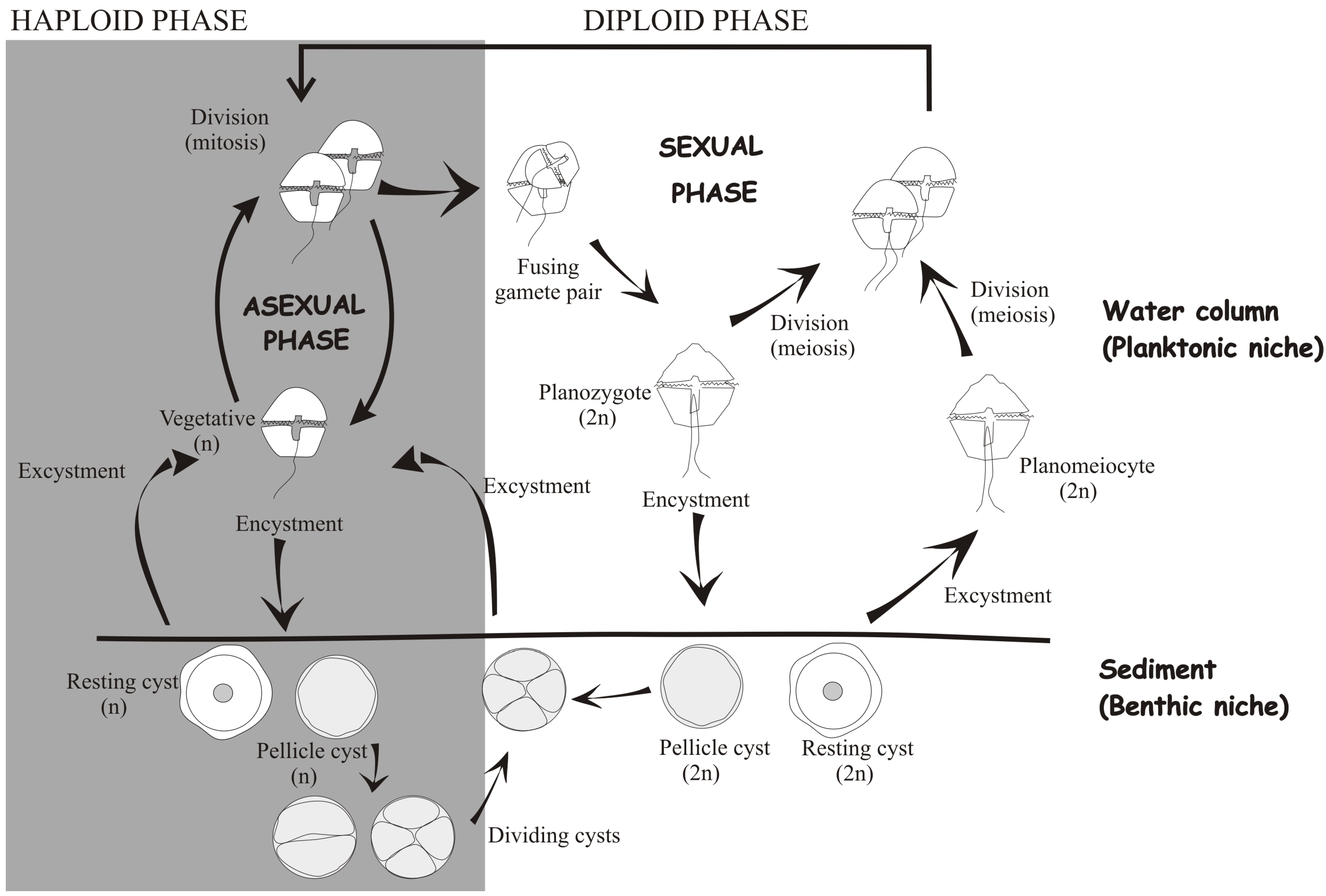
2. Structural Features of Dinoflagellate Cysts
2.1. Cyst Wall Structure
2.2. Cytoplasmic Features
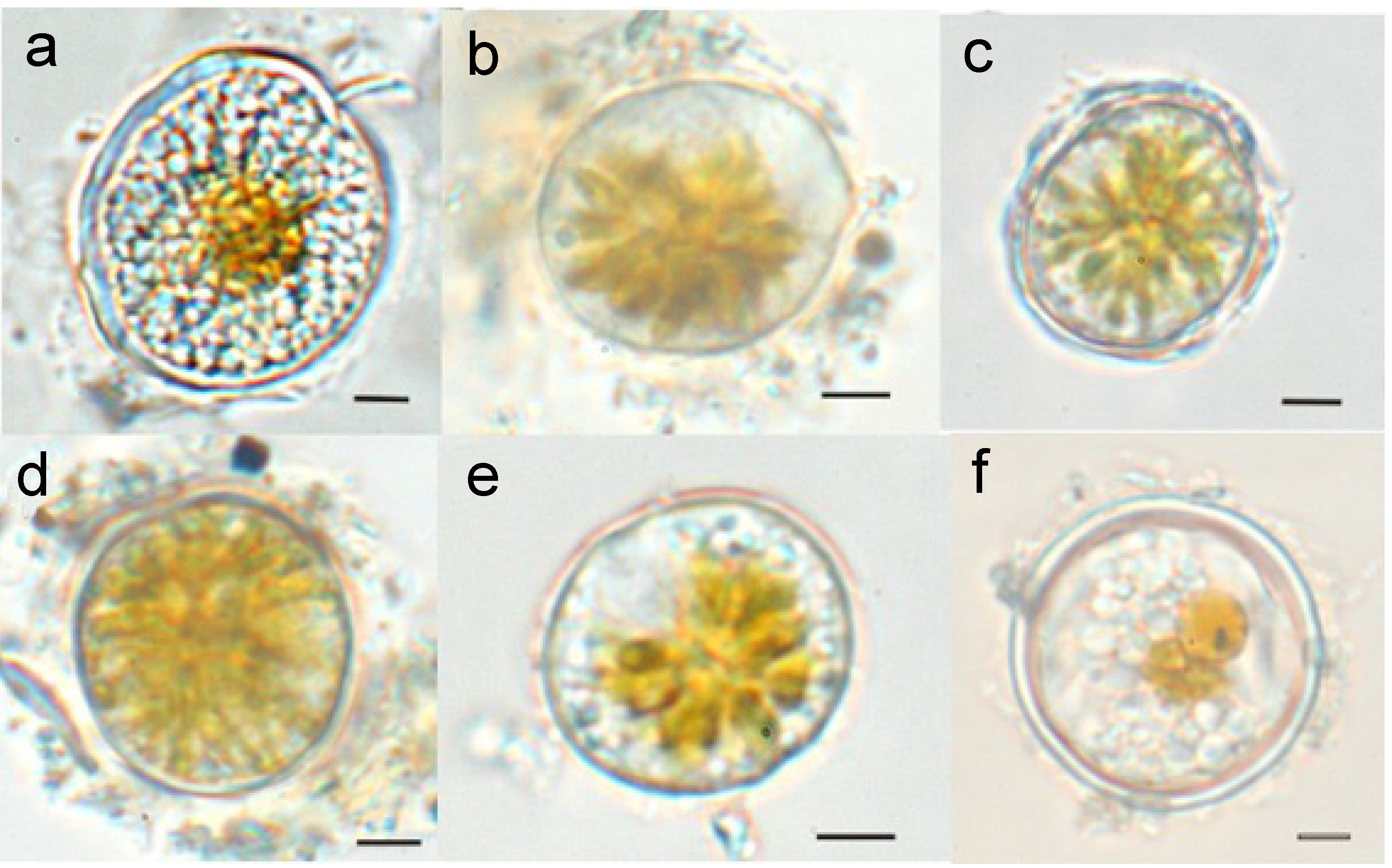
2.3. Environmental Influences
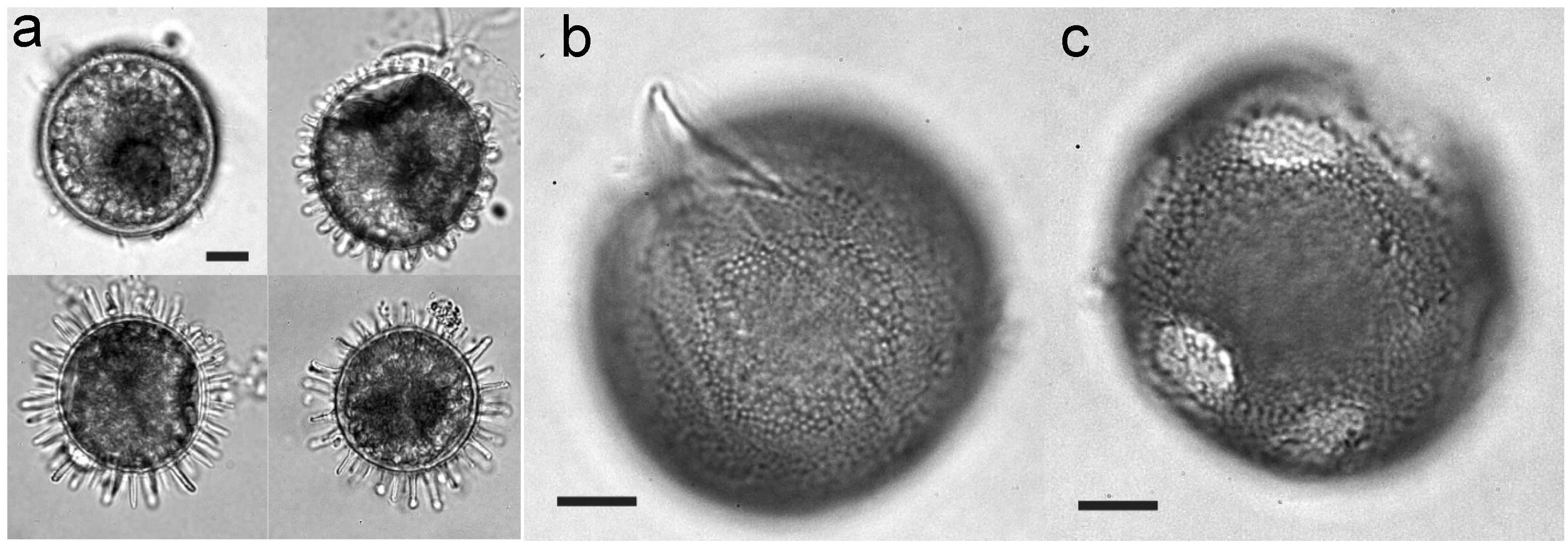
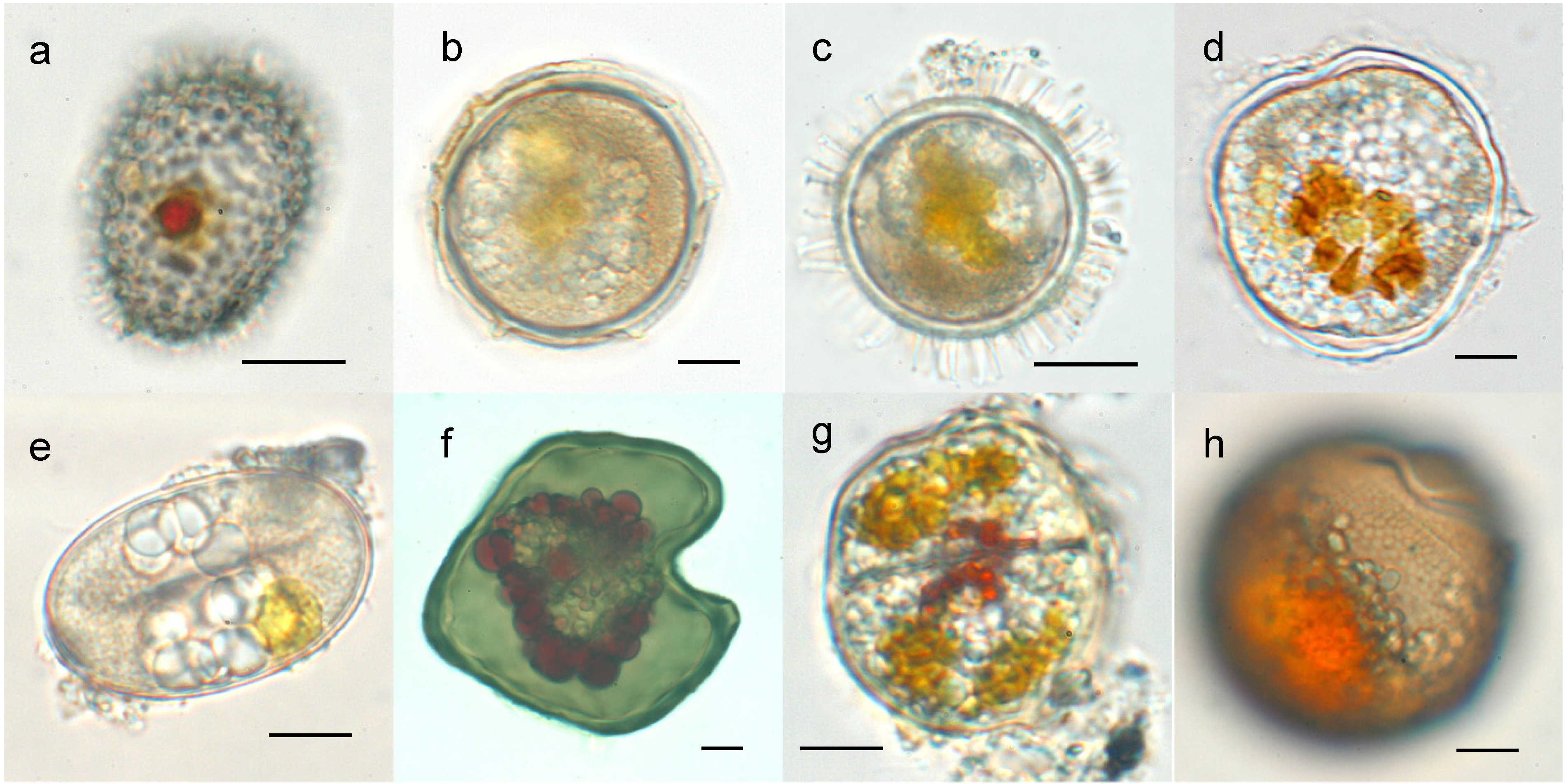{kind=link}
{kind=link}
{kind=link}
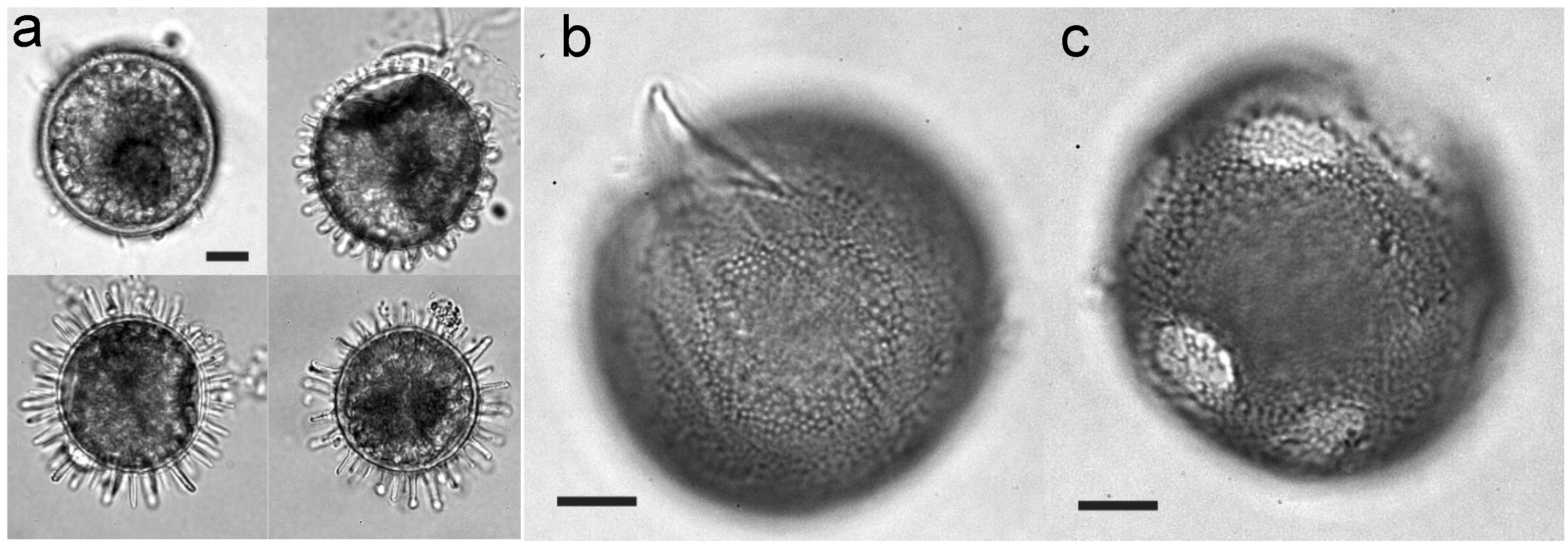{kind=link}
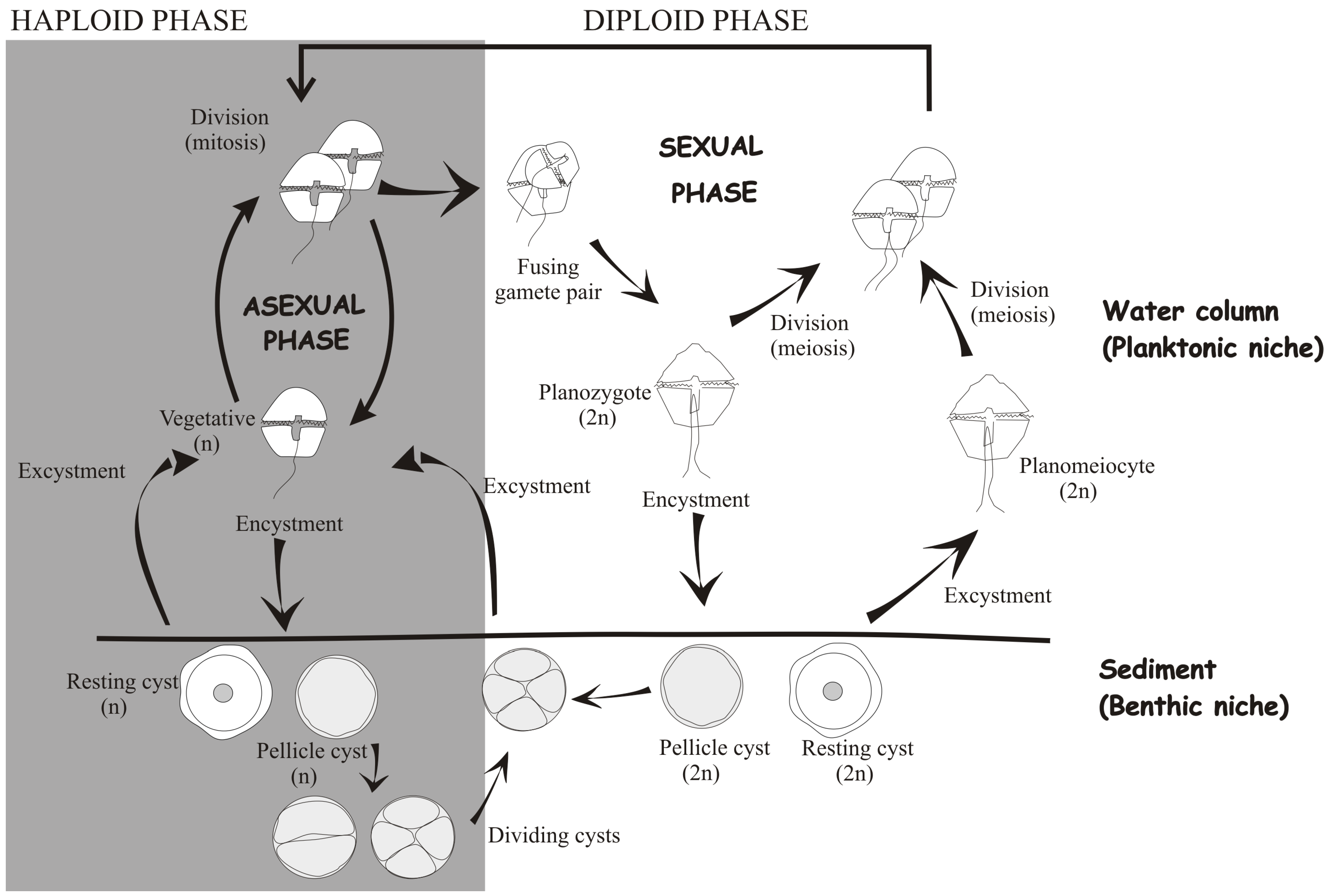
3. Functions of Dinoflagellate Cysts
3.1. Survival during Unfavorable Conditions
3.1.1. Resting Cyst Formation as a Strategy for Withstanding Unfavorable Temperature and Nutrient Conditions
3.1.2. Resting Cyst Formation as a Protective Strategy during Anoxia and Darkness
3.1.3. Resting Cyst Formation as a Strategy to Avoid Grazing
3.1.4. Resting Cyst Formation as a Defense against Parasitic Attack
3.2. Long-Term versus Short-Term Survival
3.3. Life-Cycle Strategy and Habitat
3.4. Dispersion
4. Future Perspectives
Conflicts of Interest
References
- Cavalier-Smith, T. Origins of the machinery of recombination and sex. Heredity 2002, 88, 125–141. [Google Scholar] [CrossRef]
- Wagmann, K.; Hautekeete, N.; Piquot, Y.; Meunier, C.; Schmitt, S.; van Dijk, H. Seed dormancy distribution: Explanatory ecological factors. Ann. Bot. 2012, 110, 1205–1219. [Google Scholar] [CrossRef]
- Debieu, M.; Tang, C.; Stich, B.; Sikosek, T.; Effgen, S.; Josephs, E.; Schmitt, J.; Nordborg, M.; Koornneef, M.; de Meaux, J. Co-variation between seed dormancy, growth rate and flowering time changes with latitude in Arabidopsis thaliana. PLoS One 2013, 8, e61075. [Google Scholar]
- Reinsch, P.F. Die palinosphärien, ein mikroskopischer vegetabile organismus in der mukronatenkreide. Cent. Miner. Geol. Palaeontol. 1905, 402–407. [Google Scholar]
- Erdtman, G. Fynd av Hystrichosphaera furcata i Gullmaren. Geologiska Föreningen I Stokholmen Förhanlingar 1950, 72, 221. [Google Scholar] [CrossRef]
- Erdtman, G. On pollegrains and dinoflagellate cysts in the firth of Gullmaren, S. W. Sweden. Botaniska Notiser 1954, 1954, 103–111. [Google Scholar]
- Braarud, T. Morphological Observations on Marine Dinoflagellate Cultures (Porella Perforata, Goniaulax Tamarensis, Protoceratium Reticulatum); Dybwad: Oslo, Norway, 1945; pp. 1–18. [Google Scholar]
- Nordli, E. Resting spores in Gonyaulax polyedra Stein. Nytt Mag. Naturvidenskapene 1951, 88, 207–212. [Google Scholar]
- Von Stosch, H. Dinophyta; Springer Verlag: Berlin, Germany, 1967; Volume 18, pp. 626–636. [Google Scholar]
- Von Stosch, H.A. La signification cytologique de la “cyclose nucleaire” dans le cycle de vie des dinoflagelles. Mem. Soc. Bot. Fr. 1972, 1972, 201–212. [Google Scholar]
- Von Stosch, H.A. Sexualität bei Ceratium cornutum (dinophyta). Naturwissenschaften 1965, 52, 112–113. [Google Scholar] [CrossRef]
- Von Stosch, H.A. Observations on vegetative reproduction and sexual life cycles of two freshwater dinoflagellates, Gymondinium pseudopalustre Schiller and Woloszynskia apiculata sp. nov. Br. Phycol. 1973, 8, 105–134. [Google Scholar] [CrossRef]
- Loeblich, A.I.; Loeblich, L. Dinoflagellate Cysts; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Lenormand, T.; Otto, S. The evolution of recombination in a heterogeneous environment. Genetics 2000, 156, 423–438. [Google Scholar]
- Lee, S.; Ni, M.; Li, W.; Shertz, C.; Heitman, J. The evolution of sex: A perspective from the fungal kingdom. Microbiol. Mol. Biol. Rev. 2010, 74, 298–340. [Google Scholar] [CrossRef]
- Wall, D. Modern hystrichospheres and dinoflagellate cysts from the Woods Hole region. Grana Palynol. 1965, 6, 297–314. [Google Scholar] [CrossRef]
- Steidinger, K. Implications of dinoflagellate life cycles on initiation of Gymnodinium breve red tides. Environ. Lett. 1975, 9, 129–139. [Google Scholar] [CrossRef]
- Anderson, D.M.; Wall, D. Potential importance of benthic cysts of Gonyaulax tamarensis and G. excavata in initiating toxic dinoflagellate blooms. J. Phycol. 1978, 14, 224–234. [Google Scholar] [CrossRef]
- Uchida, T. Sexual reproduction of Scrippsiella trochoidea isolated from Muroran Harbor Hokkaido. Bull. Jpn. Soc. Sci. Fish. 1991, 57, 1215. [Google Scholar] [CrossRef]
- Uchida, T.; Matsuyama, Y.; Yamaguchi, M.; Honjo, T. The life cycle of Gyrodinium instriatum (Dinophyceae) in culture. Phycol. Res. 1996, 44, 119–123. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Bravo, I. A study of the sexual reproduction and determination of mating type of Gymnodinium nolleri (Dinophyceae) in culture. J. Phycol. 2005, 41, 74–83. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Bravo, I. Sexual reproduction and two different encystment strategies of Lingulodinium polyedrum (Dinophyceae) in culture. J. Phycol. 2005, 41, 370–379. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Bravo, I.; Garcés, E. Multiples routes of sexuality in Alexandrium taylori (Dinophyceae) in culture. J. Phycol. 2006, 42, 1028–1039. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Bravo, I.; Garcés, E.; Ramilo, I. Nuclear features and effect of nutrients on Gymnodinium catenatum (Dinophyceae) sexual stages. J. Phycol. 2006, 42, 67–77. [Google Scholar] [CrossRef]
- Kremp, A.; Parrow, M. Evidence for asexual resting cysts in the life cycle of the marine peridinoid dinoflagellate, Scrippsiella hangoei. J. Phycol. 2006, 42, 400–409. [Google Scholar] [CrossRef]
- Parrow, M.W.; Burkholder, J.M. The sexual life cycles of Pfiesteria piscicida and cryptoperidiniopsoids (Dinophyceae). J. Phycol. 2004, 40, 664–673. [Google Scholar] [CrossRef]
- Dale, B. Cysts of the toxic red-tide dinoflagellate Gonyaulax excavata (Braarud) Balech from Oslofjorden, Norway. Sarsia 1977, 63, 29–34. [Google Scholar]
- Taylor, F.J.R. On dinoflagellate evolution. BioSystems 1980, 13, 65–108. [Google Scholar] [CrossRef]
- Fritsch, F.E. The Structure and Reproduction of the Algae; Cambridge University Press: London, UK, 1935; Volume I. [Google Scholar]
- Dale, B. Dinoflagellate Resting Cysts. In Survival Strategies of the Algae; Frixell, G.A., Ed.; Cambridge University Press: New York, NY, USA, 1983; pp. 67–136. [Google Scholar]
- Bravo, I.; Figueroa, R.; Garcés, E.; Fraga, S.; Massanet, A. The intricacies of dinoflagellate pellicle cysts: The example of Alexandrium minutum cysts from a bloom-recurrent area (Bay of Baiona, NW Spain). Deep Sea Res. Pt. II Top. Stud. Oceanogr. 2010, 57, 166–174. [Google Scholar] [CrossRef]
- Kita, T.; Fukuyo, Y.; Tokuda, H.; Hirano, R. Life history and ecology of Goniodoma pseudogoniaulax (Pyrrhophyta) in a rockpool. Bull. Mar. Sci. 1985, 37, 643–651. [Google Scholar]
- Garcés, E.; Masó, M.; Camp, J. Role of temporary cysts in the population dynamics of Alexandrium taylori (Dinophyceae). J. Plankton Res. 2002, 24, 681–686. [Google Scholar] [CrossRef]
- Lindberg, K.; Moestrup, Ø.; Daugbjerg, N. Studies on woloszynskioid dinoflagellates I: Woloszynskia coronata re-examined using light and electron microscopy and partial LSU rDNA sequences, with description of Tovellia gen. nov. and Jadwigia gen. nov. (Tovelliaceae fam. nov.). Phycologia 2005, 44, 416–440. [Google Scholar] [CrossRef]
- Gribble, K.E.; Anderson, D.M.; Coats, D.W. Sexual and asexual processes in Protoperidinium steidingerae Balech (Dinophyceae), with observations on life-history stages of Protoperidinium depressum (Bailey) Balech (Dinophyceae). J. Eukaryot. Microbiol. 2009, 56, 88–103. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Bravo, I.; Fraga, S.; Garces, E.; Llaveria, G. The life history and cell cycle of Kryptoperidinium foliaceum, a dinoflagellate with two eukaryotic nuclei. Protist 2009, 160, 285–300. [Google Scholar] [CrossRef]
- Litaker, R.W.; Vandersea, M.W.; Kibler, S.R.; Madden, V.J.; Noga, E.J.; Tester, P.A. Life cycle of the heterotrophic dinoflagellate Pfiesteria piscicida (Dinophyceae). J. Phycol. 2002, 38, 442–463. [Google Scholar]
- Zeng, N.; Gu, H.; Smith, K.F.; Rhodes, L.L.; Selwood, A.I.; Yang, W. The first report of Vulcanodinium rugosum (Dinophyceae) from the South China Sea with a focus on the life cycle. N. Z. J. Mar. Freshw. Res. 2012, 46, 511–521. [Google Scholar] [CrossRef]
- Evitt, W.R. Observations on the morphology of fossil dinoflagellates. Micropaleontology 1961, 7, 385–420. [Google Scholar] [CrossRef]
- Evitt, W.R. Dinoflagellates and Other Organisms in Palynological Preparations. In Aspects of Palynology; Tschudy, R.H., Scott, R.A., Eds.; Wiley-Interscience: New York, NY, USA, 1969; pp. 439–479. [Google Scholar]
- Harland, R. A summary review of the morphology and classification of the fossil peridiniales (dinoflagellates) with respect to their modern representatives. Geophytology 1971, 1, 135–150. [Google Scholar]
- Evitt, W.R.; Lentin, J.K.; Millioud, M.E.; Stover, L.E.; Williams, G.L. Dinoflagellate Cyst Terminology; Energy, Mines, and Resources Canada: Ottawa, Canada, 1977; pp. 1–11. [Google Scholar]
- Williams, G.L.; Sarjeant, W.A.S.; Kidson, E.J. A Glossary of the Terminology Applied to Dinoflagellate Amphiesmae and Cysts and Acritarchs; American Association of Stratigraphic Palynologists: Houston, TX, USA, 1978; Volume 2, pp. 1–121. [Google Scholar]
- Matsuoka, K.; Fukuyo, Y.; Anderson, D. Methods for Modern Dinoflagellate Cyst Studies. In Red Tides: Biology, Environmental Science and Toxicology; Okaichi, T., Anderson, D.M., Nemoto, K., Eds.; Elsevier: New York, NY, USA, 1989; pp. 461–479. [Google Scholar]
- Matsuoka, K.; Fukuyo, Y. Technical Guide for Modern Dinoflagellate Cyst Study; WESTPAC-HAB/WESTPAC/IOC: Tokio, Japan, 2000; p. 75. [Google Scholar]
- Mertens, K.N.; Ribeiro, S.; Bouimetarhan, I.; Caner, H.; Nebout, N.C.; Dale, B.; De Vernal, A.; Ellegaard, M.; Filipova, M.; Godhe, A.; et al. Process length variation in cysts of a dinoflagellate, Lingulodinium machaerophorum, in surface sediments: Investigating its potential as salinity proxy. Mar. Micropaleontol. 2009, 70, 54–69. [Google Scholar] [CrossRef]
- Kokinos, J.P.; Anderson, D.M. Morphological development of resting cysts in cultures of the marine dinoflagellate Lingulodinium polyedrum (=L. machaerophorum). Palynology 1995, 19, 143–166. [Google Scholar]
- Bibby, B.T.; Dodge, J.D. The encystment of a freshwater dinoflagellate: A light and electron-microscopical study. Br. Phycol. J. 1972, 7, 85–100. [Google Scholar]
- Dürr, V.G. Electron microscope studies on the theca of dinoflagellates. 3. The cysts of Peridinium cinctum. Arch. Protistenkd. 1979, 122, 121–139. [Google Scholar] [CrossRef]
- De Leeuw, J.; Versteegh, G.; van Bergen, P. Biomacromolecules of algae and plants and their fossil analogues. Plant Ecol. 2006, 182, 209–233. [Google Scholar]
- Kokinos, J.P.; Eglinton, T.I.; Goñi, M.A.; Boon, J.J.; Martoglio, P.A.; Anderson, D.M. Characterization of a highly resistant biomacromolecular material in the cell wall of a marine dinoflagellate resting cyst. Org. Geochem. 1998, 28, 265–288. [Google Scholar] [CrossRef]
- Versteegh, G.; Blokker, P.; Wood, G.; Collinson, M.; Damste, J.; de Leeuw, J. An example of oxidative polymerization of unsaturated fatty acids as a preservation pathway for dinoflagellate organic matter. Org. Geochem. 2004, 35, 1129–1139. [Google Scholar] [CrossRef]
- Versteegh, G.; Blokker, P.; Bogus, K.; Harding, I.; Lewis, J.; Oltmanns, S.; Rochon, A.; Zonneveld, K. Infra red spectroscopy, flash pyrolysis, thermally assisted hydrolysis and methylation (THM) in the presence of tetramethylammonium hydroxide (TMAH) of cultured and sediment-derived Lingulodinium polyedrum (Dinoflagellata) cyst walls. Org. Geochem. 2012, 43, 92–102. [Google Scholar] [CrossRef]
- Taylor, F.J.R. Dinoflagellate Morphology. In The Biology of Dinoflagellates; Taylor, F.J.R., Ed.; Blackwell Scientific Publications: Oxford, UK, 1987; Volume 21, pp. 24–91. [Google Scholar]
- Loeblich, A.R.I. The Amphiesma or Dinoflagellate Cell Covering. In Proceedings of the North American Paleontological Convention: Field Museum of Natural History, Chicago, IL, USA, 5–7 September, 1970; Yochelson, E.L., Ed.; Allen Press: Lawrence, KS, USA, 1971; Volume 2, pp. 867–929. [Google Scholar]
- Morrill, L.C.; Loeblich, A.R. The dinoflagellate pellicular wall layer and its occurrence in the Division Pyrrhophyta. J. Phycol. 1981, 17, 315–323. [Google Scholar] [CrossRef]
- Höhfeld, I.; Melkonian, M. Amphiesmal ultrastructure of dinoflagellates: A reevaluation of pellicle formation. J. Phycol. 1992, 28, 82–89. [Google Scholar]
- Gao, X.P.; Dodge, J.D.; Lewis, J. An ultrastructural-study of planozygotes and encystment of a marine dinoflagellate, Scrippsiella sp. Br. Phycol. J. 1989, 24, 153–165. [Google Scholar]
- Chapman, D.V.; Dodge, J.D.; Heaney, S.I. Cyst formation in the fresh-water dinoflagellate Ceratium hirundinella (Dinophyceae). J. Phycol. 1982, 18, 121–129. [Google Scholar] [CrossRef]
- Fritz, L.; Anderson, D.M.; Triemer, R.E. Ultrastructural aspects of sexual reproduction in the red tide dinoflagellate Gonyaulax tamarensis. J. Phycol. 1989, 25, 95–107. [Google Scholar] [CrossRef]
- Kennaway, G.M.; Lewis, J.M. An ultrastructural study of hypnozygotes of Alexandrium species (Dinophyceae). Phycologia 2004, 43, 353–363. [Google Scholar]
- Bravo, I.; Garces, E.; Diogene, J.; Fraga, S.; Sampedro, N.; Figueroa, R.I. Resting cysts of the toxigenic dinoflagellate genus Alexandrium in recent sediments from the Western Mediterranean Coast, including the first description of cysts of A. kutnerae and A. peruvianum. Eur. J. Phycol. 2006, 41, 293–302. [Google Scholar] [CrossRef]
- Mertens, K.N.; Bringue, M.; van Nieuwenhove, N.; Takano, Y.; Pospelova, V.; Rochon, A.; de Vernal, A.; Radi, T.; Dale, B.; Patterson, R.T.; et al. Process length variation of the cyst of the dinoflagellate Protoceratium reticulatum in the North Pacific and Baltic-Skagerrak region: Calibration as an annual density proxy and first evidence of pseudo-cryptic speciation. J. Quat. Sci. 2012, 27, 734–744. [Google Scholar] [CrossRef]
- Ellegaard, M.; Lewis, J.; Harding, I. Cyst-theca relationship, life cycle, and effects of temperature and salinity on the cyst morphology of Gonyaulax baltica sp nov (Dinophyceae) from the Baltic Sea area. J. Phycol. 2002, 38, 775–789. [Google Scholar] [CrossRef]
- Zonneveld, K.A.F.; Susek, E. Effects of temperature, light and salinity on cyst production and morphology of Tuberculodinium vancampoae (the resting cyst of Pyrophacus steinii). Rev. Palaeobot. Palynol. 2007, 145, 77–88. [Google Scholar] [CrossRef]
- Hallett, R.I. Consequences of Environmental Change on the Growth and Morphology of Lingulodinium polyedrum (Dinophyceae) in Culture. Ph.D. Thesis, University of Westminster, London, UK, 1999. [Google Scholar]
- Wall, D. Biological problems concerning fossilizable dinoflagellates. Geosci. Man 1971, 3, 1–15. [Google Scholar] [CrossRef]
- Anderson, D.M. The Roles of Dormant Cysts in Toxic Dinoflagellate Blooms and Shellfish Toxicity. In Seafood Toxins; Ragelis, E.P., Ed.; American Chemical Society: Washington, DC, USA, 1984; Volume 262, pp. 125–138. [Google Scholar]
- Pouchet, G. Contribution à l’histoire des ciclioflagellés. J. Anat. Physiol. 1883, 19, 399–455. [Google Scholar]
- Biecheler, B. Recherches sur les péridiniens. Bull. Biol. Fr. Belg. 1952, 36 (Suppl.), 1–149. [Google Scholar]
- Barlow, S.B.; Triemer, R.E. Alternate life-history stages in Amphidinium klebsii (Dinophyceae, Pyrrophyta). Phycologia 1988, 27, 413–420. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Garces, E.; Bravo, I. Comparative study of the life cycles of Alexandrium tamutum and Alexandrium minutum (Gonyaulacales, Dinophyceae) in culture. J. Phycol. 2007, 43, 1039–1053. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Rengefors, K.; Bravo, I. Effects of parental factors and meiosis on sexual offspring of Gymnodinium nolleri (Dinophyceae). J. Phycol. 2006, 42, 350–362. [Google Scholar] [CrossRef]
- Kühn, S.F.; Medlin, L.K. The systematic position of the parasitoid marine dinoflagellate Paulsenella vonstoschii (Dinophyceae) inferred from nuclear-encoded small subunit ribosomal DNA. Protist 2005, 156, 393–398. [Google Scholar] [CrossRef]
- Alpermann, T.J.; Beszteri, B.; John, U.; Tillmann, U.; Cembella, A.D. Implications of life-history transitions on the population genetic structure of the toxigenic marine dinoflagellate Alexandrium tamarense. Mol. Ecol. 2009, 18, 2122–2133. [Google Scholar]
- Miyazono, A.; Nagai, S.; Kudo, I.; Tanizawa, K. Viability of Alexandrium tamarense cysts in the sediment of Funka Bay, Hokkaido, Japan: Over a hundred year survival times for cysts. Harmful Algae 2012, 16, 81–88. [Google Scholar] [CrossRef]
- Pfiester, L.A.; Anderson, D.M. Dinoflagellate Reproduction. In The Biology of Dinoflagellates; Taylor, F.J.R, Ed.; Blackwell Scientific: Oxford, UK, 1987; pp. 611–648. [Google Scholar]
- Anderson, D.M.; Lindquist, N.L. Time-course measurements of phosphorus depletion and cyst formation in the dinoflagellate Gonyaulax tamarensis Lebour. J. Exp. Mar. Biol. Ecol. 1985, 86, 1–13. [Google Scholar] [CrossRef]
- Anderson, D.M.; Coats, D.W.; Tyler, M.A. Encystment of the dinoflagellate Gyrodinium uncatenum temperatura and nutrient effect. J. Phycol. 1985, 21, 200–206. [Google Scholar] [CrossRef]
- Rengefors, K.; Anderson, D.M. Environmental and endogenous regulation of cyst germination in two freshwater dinoflagellates. J. Phycol. 1998, 34, 568–577. [Google Scholar]
- Grzebyk, D.; Berland, B. Influences of temperature, salinity and irradiance on growth of Prorocentrum minimum (Dinophyceae) from the Mediterranean Sea. J. Plankton Res. 1996, 18, 1837–1849. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Bravo, I.; Garces, E. Effects of nutritional factors and different parental crosses on the encystment and excystment of Alexandrium catenella (Dinophyceae) in culture. Phycologia 2005, 44, 658–670. [Google Scholar]
- Anderson, D.M.; Taylor, C.D.; Armbrust, E.V. The effect of darkness and anaerobiosis on dinoflagellate cyst germination. Limnol. Oceanogr. 1987, 32, 340–351. [Google Scholar] [CrossRef]
- Bravo, I.; Anderson, D.M. The effects of temperatura, growth-medium and darkness on excystment and growth of the toxic dinoflagellate Gymnodinium catenatum from northwest Spain. J. Plankton Res. 1994, 16, 513–525. [Google Scholar] [CrossRef]
- Kremp, A.; Anderson, D.M. Factors regulating germination of resting cysts of the spring bloom dinoflagellate Scrippsiella hangoei from the Northern Baltic Sea. J. Plankton Res. 2000, 22, 1311–1327. [Google Scholar] [CrossRef]
- Lundgren, V.; Graneli, E. Influence of altered light conditions and grazers on Scrippsiella trochoidea (Dinophyceae) cyst formation. Aquat. Microb. Ecol. 2011, 63, 231–243. [Google Scholar] [CrossRef]
- Rintala, J.M.; Spilling, K.; Blomster, J. Temporary cyst enables long-term dark survival of Scrippsiella hangoei (Dinophyceae). Mar. Biol. 2007, 152, 57–62. [Google Scholar] [CrossRef]
- Fistarol, G.O.; Legrand, C.; Rengefors, K.; Graneli, E. Temporary cyst formation in phytoplankton: A response to allelopathic competitors? Environ. Microbiol. 2004, 6, 791–798. [Google Scholar] [CrossRef]
- Tillmann, U.; John, U.; Cembella, A. On the allelochemical potency of the marine dinoflagellate Alexandrium ostenfeldii against heterotrophic and autotrophic protists. J. Plankton Res. 2007, 29, 527–543. [Google Scholar] [CrossRef]
- Montresor, M.; Nuzzo, L.; Mazzocchi, M.G. Viability of dinoflagellate cysts after the passage through the copepod gut. J. Exp. Mar. Biol. Ecol. 2003, 287, 209–221. [Google Scholar] [CrossRef]
- Laabir, M.; Amzil, Z.; Lassus, P.; Masseret, E.; Tapilatu, Y.; de Vargas, R.; Grzebyk, D. Viability, growth and toxicity of Alexandrium catenella and Alexandrium minutum (Dinophyceae) following ingestion and gut passage in the oyster Crassostrea gigas. Aquat. Living Resour. 2007, 20, 51–57. [Google Scholar] [CrossRef]
- Toth, G.B.; Noren, F.; Selander, E.; Pavia, H. Marine dinoflagellates show induced life-history shifts to escape parasite infection in response to water-borne signals. Proc. R. Soc. B Biol. Sci. 2004, 271, 733–738. [Google Scholar] [CrossRef]
- Figueroa, R.I.; Garces, E.; Massana, R.; Camp, J. Description, host-specificity, and strain selectivity of the dinoflagellate parasite Parvilucifera sinerae sp nov (Perkinsozoa). Protist 2008, 159, 563–578. [Google Scholar] [CrossRef]
- Chambouvet, A.; Alves-de-Souza, C.; Cueff, V.; Marie, D.; Karpov, S.; Guillou, L. Interplay between the parasite Amoebophrya sp. (Alveolata) and the cyst formation of the red tide dinoflagellate Scrippsiella trochoidea. Protist 2011, 162, 637–649. [Google Scholar] [CrossRef]
- Wall, D. Taxonomy and Cysts of Red-Tide Dinoflagellates. In Proceedings of the First International Conference on Toxic Dinoflagellate Blooms, Boston, MA, USA, November 1974; Lo Cicero, V.R., Ed.; Massachusetts Science and Technology Foundation: Wakefield, MA, USA, 1975; pp. 249–255. [Google Scholar]
- Anderson, D.M. Effects of temperature conditioning on development and germination of Gonyaulax tamarensis (Dinophyceae) hypnozygotes. J. Phycol. 1980, 16, 166–172. [Google Scholar]
- Reynolds, C.S. The Ecology of Freshwater Phytoplankton; Cambridge University Press: London, UK, 1984. [Google Scholar]
- Anderson, D.M.; Keafer, B.A. An endogenous annual clock in the toxic marine dinoflagellate Gonyaulax tamarensis. Nature 1987, 325, 616–617. [Google Scholar] [CrossRef]
- Anderson, D.M. Physiology and Bloom Dynamics of Toxic Alexandrium Species, with Emphasis on Life Cycle Transitions. In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer-Verlag: Heidelberg, Germany, 1998; Volume 41, pp. 29–47. [Google Scholar]
- Steidinger, K.; Haddad, K. Biologic and hydrographic aspects of red tides. BioScience 1981, 31, 814–819. [Google Scholar] [CrossRef]
- Heiskanen, A.S. Mass encystment and sinking of dinoflagellates during a spring bloom. Mar. Biol. 1993, 116, 161–167. [Google Scholar] [CrossRef]
- Kremp, A. Effects of cyst resuspension on germination and seeding of two bloom-forming dinoflagellates in the Baltic Sea. Mar. Ecol. Prog. Ser. 2001, 216, 57–66. [Google Scholar] [CrossRef]
- Spilling, K.; Kremp, A.; Tamelander, T. Vertical distribution and cyst production of Peridiniella catenata (Dinophyceae) during a spring bloom in the Baltic Sea. J. Plankton Res. 2006, 28, 659–665. [Google Scholar] [CrossRef]
- Lundholm, N.; Ribeiro, S.; Andersen, T.; Koch, T.; Godhe, A.; Ekelund, F.; Ellegaard, M. Buried alive-germination of up to a century-old marine protist resting stages. Phycologia 2011, 50, 629–640. [Google Scholar] [CrossRef]
- Ribeiro, S.; Berge, T.; Lundholm, N.; Andersen, T.; Abrantes, F.; Ellegaard, M. Phytoplankton growth after a century of dormancy illuminates past resilience to catastrophic darkness. Nat. Commun. 2011, 2. [Google Scholar] [CrossRef]
- Smayda, T.; Trainer, V. Dinoflagellate blooms in upwelling systems: Seeding, variability, and contrasts with diatom bloom behaviour. Prog. Oceanogr. 2010, 85, 92–107. [Google Scholar] [CrossRef]
- Garcés, E.; Masó, M.; Camp, J. A recurrent and localized dinoflagellate bloom in Mediterranean beach. J. Plankton Res. 1999, 21, 2373–2391. [Google Scholar] [CrossRef]
- Basterretxea, G.; Garcés, E.; Masó, M.; Jordi, A.; Masó, M.; Tintoré, J. Breeze conditions as a favoring mechanism of Alexandrium taylori blooms at a Mediterranean beach. Estuar. Coast. Mar. Sci. 2005, 62, 1–12. [Google Scholar]
- Bravo, I.; Fraga, S.; Figueroa, R.I.; Pazos, Y.; Massanet, A.; Ramilo, I. Bloom dynamics and life cycle strategies of two toxic dinoflagellates in a coastal upwelling system (NW Iberian Peninsula). Deep Sea Res. Pt. II Top. Stud. Oceanogr. 2010, 57, 222–234. [Google Scholar]
- Figueroa, R.I.; Bravo, I.; Ramilo, I.; Pazos, Y.; Angeles, M. New life-cycle stages of Gymnodinium catenatum (Dinophyceae): Laboratory and field observations. Aquat. Microb. Ecol. 2008, 52, 13–23. [Google Scholar] [CrossRef]
- Kim, C.H.; Cho, H.J.; Shin, J.B.; Moon, C.H.; Matsuoka, K. Regeneration from hyaline cysts of Cochlodinium polykrikoides (Gymnodiniales, Dinophyceae). A red tide organism along the Korean Coast. Phycologia 2002, 41, 667–669. [Google Scholar] [CrossRef]
- Bravo, I.; Vila, M.; Casabianca, S.; Rodriguez, F.; Rial, P.; Riobó, P.; Penna, A. Life cycle stages of the benthic palytoxin-producing dinoflagellate Ostreopsis cf. ovata (Dinophyceae). Harmful Algae 2012, 18, 24–34. [Google Scholar] [CrossRef] [Green Version]
- Salgado, P.; Pizarro, G.; Guzmán, L.; Bravo, I. Primer Registro de Células de Resistencia del Dinoflagelado Prorocentrum lima en Sedimentos de la Costa Austral de Chile. In Proceedings of the XXX Congreso de Ciencias del Mar, Concepción, Chile, 19–22 October 2010.
- Southwood, T.R.E. Tactics, strategies and templets. Oikos 1988, 52, 3–18. [Google Scholar] [CrossRef]
- Kita, T.; Fukuyo, Y.; Tokuda, H.; Hirano, R. Sexual reproduction of Alexandrium hiranoi (Dinophyceae). Bull. Plankon Soc. Jpn. 1993, 39, 79–85. [Google Scholar]
- Anderson, D.M.; Lively, J.J.; Reardon, E.M.; Price, C.A. Sinking characteristics of dinoflagellate cysts. Limnol. Oceanogr. 1985, 30, 1000–1009. [Google Scholar] [CrossRef]
- McNown, J.S.; Malaika, J. Effects of particle shape on settling velocity at low reynolds numbers. Trans. Am. Geophys. Union 1950, 31, 74–82. [Google Scholar] [CrossRef]
- Belmonte, G.; Miglietta, A.; Rubino, F.; Boero, F. Morphological convergence of resting stages of planktonic organisms: A review. Hydrobiologia 1997, 355, 159–165. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bravo, I.; Figueroa, R.I. Towards an Ecological Understanding of Dinoflagellate Cyst Functions. Microorganisms 2014, 2, 11-32. https://doi.org/10.3390/microorganisms2010011
Bravo I, Figueroa RI. Towards an Ecological Understanding of Dinoflagellate Cyst Functions. Microorganisms. 2014; 2(1):11-32. https://doi.org/10.3390/microorganisms2010011
Chicago/Turabian StyleBravo, Isabel, and Rosa Isabel Figueroa. 2014. "Towards an Ecological Understanding of Dinoflagellate Cyst Functions" Microorganisms 2, no. 1: 11-32. https://doi.org/10.3390/microorganisms2010011