
Streptokinase Treatment Reverses Biofilm-Associated Antibiotic Resistance in Staphylococcus aureus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Plasma Extraction
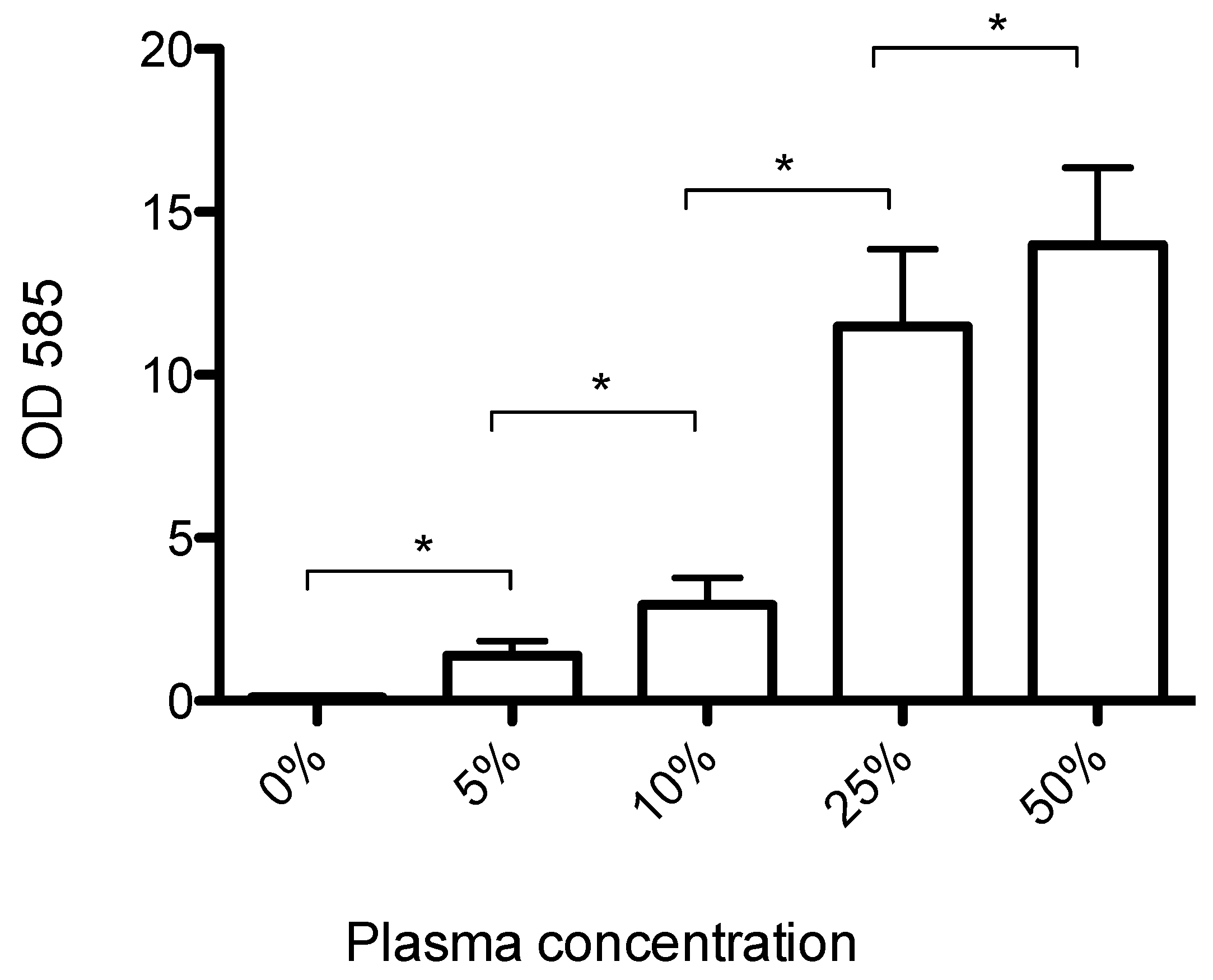
2.2. Quantification of Human Plasma’s Effect on Biofilm Formation
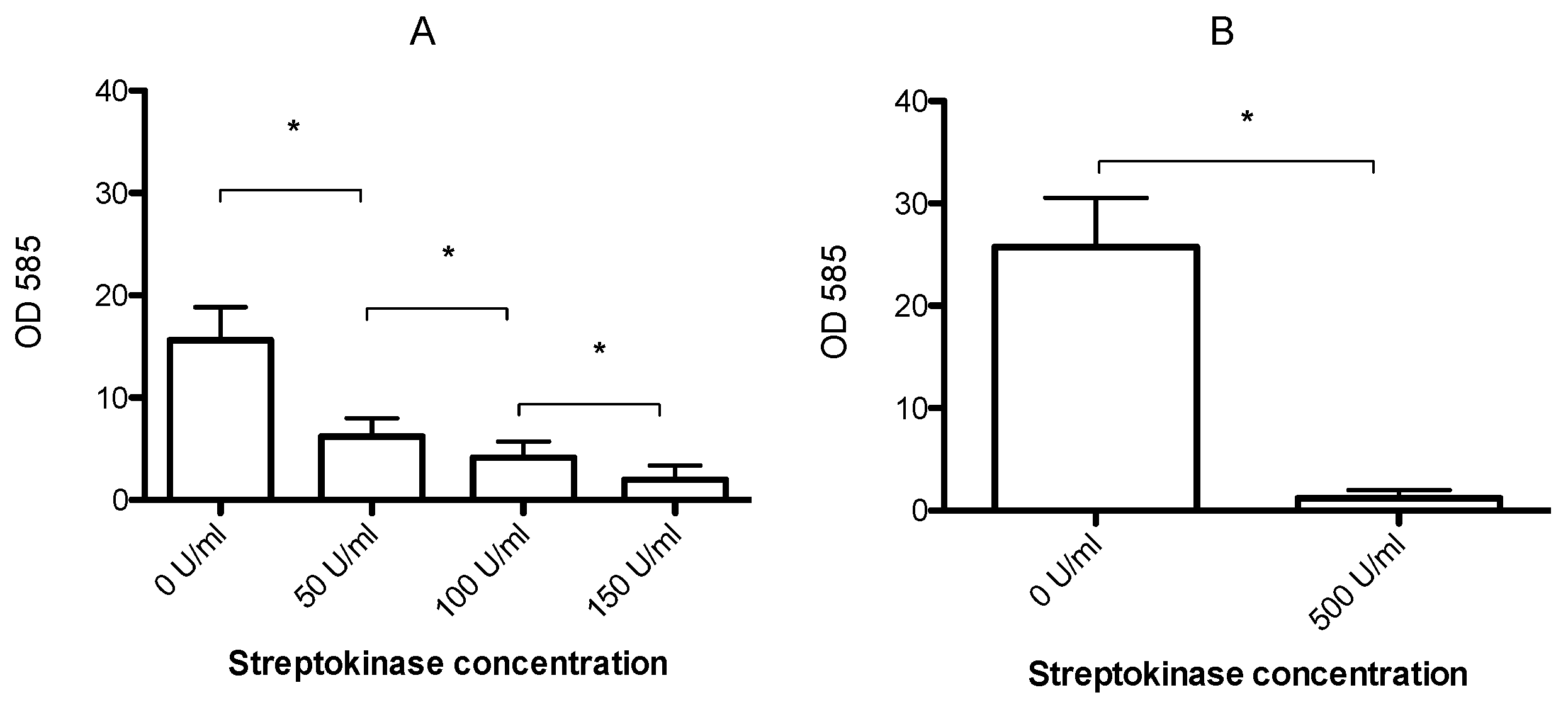
2.3. Biofilm Dispersal with Streptokinase
2.4. MBEC of Antibiotics Combined with Streptokinase
2.5. Confocal Laser Scanning Microscopy (CLSM)
2.6. Statistics
3. Results
3.1. Human Plasma Promotes Formation of Biofilms That Can Be Dispersed by Streptokinase Treatment
3.2. Streptokinase Treatment Lowers the MBEC to Clinically Relevant Levels
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Resch, A.; Leicht, S.; Saric, M.; Pasztor, L.; Jakob, A.; Gotz, F.; Nordheim, A. Comparative proteome analysis of Staphylococcus aureus biofilm and planktonic cells and correlation with transcriptome profiling. Proteomics 2006, 6, 1867–1877. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Hook, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal infections: Mechanisms of biofilm maturation and detachment as critical determinants of pathogenicity. Annu. Rev. Med. 2013, 64, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Deivanayagam, C.C.; Wann, E.R.; Chen, W.; Carson, M.; Rajashankar, K.R.; Hook, M.; Narayana, S.V. A novel variant of the immunoglobulin fold in surface adhesins of Staphylococcus aureus: Crystal structure of the fibrinogen-binding mscramm, clumping factor a. EMBO J. 2002, 21, 6660–6672. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, V.K.; Barbu, E.M.; Deivanayagam, C.C.; Le, B.; Anderson, A.S.; Matsuka, Y.V.; Lin, S.L.; Foster, T.J.; Narayana, S.V.; Hook, M. Structural and biochemical characterization of Staphylococcus aureus clumping factor B/ligand interactions. J. Biol. Chem. 2011, 286, 25963–25972. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, L.; Kerrigan, S.W.; Kaw, G.; Hogan, M.; Penades, J.; Litt, D.; Fitzgerald, D.J.; Foster, T.J.; Cox, D. Multiple mechanisms for the activation of human platelet aggregation by Staphylococcus aureus: Roles for the clumping factors CLFA and CLFB, the serine-aspartate repeat protein sdre and protein A. Mol. Microbiol. 2002, 44, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Keane, F.M.; Loughman, A.; Valtulina, V.; Brennan, M.; Speziale, P.; Foster, T.J. Fibrinogen and elastin bind to the same region within the a domain of fibronectin binding protein A, an mscramm of Staphylococcus aureus. Mol. Microbiol. 2007, 63, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Sinha, B.; Francois, P.; Que, Y.A.; Hussain, M.; Heilmann, C.; Moreillon, P.; Lew, D.; Krause, K.H.; Peters, G.; Herrmann, M. Heterologously expressed Staphylococcus aureus fibronectin-binding proteins are sufficient for invasion of host cells. Infect. Immun. 2000, 68, 6871–6878. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.R.; Harris, L.G.; Richards, R.G.; Foster, S.J. Analysis of ebh, a 1.1-megadalton cell wall-associated fibronectin-binding protein of Staphylococcus aureus. Infect. Immun. 2002, 70, 6680–6687. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Xu, Y.; Liang, X.; Keene, D.R.; Hook, A.; Gurusiddappa, S.; Hook, M.; Narayana, S.V. A ‘collagen hug’ model for Staphylococcus aureus cna binding to collagen. EMBO J. 2005, 24, 4224–4236. [Google Scholar] [CrossRef] [PubMed]
- Walden, M.; Edwards, J.M.; Dziewulska, A.M.; Bergmann, R.; Saalbach, G.; Kan, S.Y.; Miller, O.K.; Weckener, M.; Jackson, R.J.; Shirran, S.L.; et al. An internal thioester in a pathogen surface protein mediates covalent host binding. Elife 2015, 4, e06638. [Google Scholar] [CrossRef] [PubMed]
- Brady, R.A.; Leid, J.G.; Calhoun, J.H.; Costerton, J.W.; Shirtliff, M.E. Osteomyelitis and the role of biofilms in chronic infection. FEMS Immunol. Med. Microbiol. 2008, 52, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Archer, N.K.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms: Properties, regulation, and roles in human disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Lister, J.L.; Horswill, A.R. Staphylococcus aureus biofilms: Recent developments in biofilm dispersal. Front. Cell. Infect. Microbiol. 2014, 4, 178. [Google Scholar] [CrossRef] [PubMed]
- Gristina, A.G. Biomaterial-centered infection: Microbial adhesion versus tissue integration. Science 1987, 237, 1588–1595. [Google Scholar] [CrossRef] [PubMed]
- Montanaro, L.; Speziale, P.; Campoccia, D.; Ravaioli, S.; Cangini, I.; Pietrocola, G.; Giannini, S.; Arciola, C.R. Scenery of staphylococcus implant infections in orthopedics. Future Microbiol. 2011, 6, 1329–1349. [Google Scholar] [CrossRef] [PubMed]
- Nethercott, C.; Mabbett, A.N.; Totsika, M.; Peters, P.; Ortiz, J.C.; Nimmo, G.R.; Coombs, G.W.; Walker, M.J.; Schembri, M.A. Molecular characterization of endocarditis-associated Staphylococcus aureus. J. Clin. Microbiol. 2013, 51, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Cardile, A.P.; Sanchez, C.J., Jr.; Samberg, M.E.; Romano, D.R.; Hardy, S.K.; Wenke, J.C.; Murray, C.K.; Akers, K.S. Human plasma enhances the expression of staphylococcal microbial surface components recognizing adhesive matrix molecules promoting biofilm formation and increases antimicrobial tolerance in vitro. BMC Res. Notes 2014, 7, 457. [Google Scholar] [CrossRef] [PubMed]
- Zapotoczna, M.; McCarthy, H.; Rudkin, J.K.; O’Gara, J.P.; O’Neill, E. An essential role for coagulase in Staphylococcus aureus biofilm development reveals new therapeutic possibilities for device-related infections. J. Infect. Dis. 2015, 212, 1883–1893. [Google Scholar] [CrossRef] [PubMed]
- Kwiecinski, J.; Peetermans, M.; Liesenborghs, L.; Na, M.; Bjornsdottir, H.; Zhu, X.; Jacobsson, G.; Johansson, B.R.; Geoghegan, J.A.; Foster, T.J.; et al. Staphylokinase control of Staphylococcus aureus biofilm formation and detachment through host plasminogen activation. J. Infect. Dis. 2016, 213, 139–148. [Google Scholar] [CrossRef] [PubMed]
- McCourt, J.; O’Halloran, D.P.; McCarthy, H.; O’Gara, J.P.; Geoghegan, J.A. Fibronectin-binding proteins are required for biofilm formation by community-associated methicillin-resistant Staphylococcus aureus strain lac. FEMS Microbiol. Lett. 2014, 353, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.G.; DeDent, A.C.; Schneewind, O.; Missiakas, D. A play in four acts: Staphylococcus aureus abscess formation. Trends Microbiol. 2011, 19, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Rammelkamp, C.H.; Hezebicks, M.M.; Dingle, J.H. Specific coagulases of Staphylococcus aureus. J. Exp. Med. 1950, 91, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Coraca-Huber, D.C.; Fille, M.; Hausdorfer, J.; Pfaller, K.; Nogler, M. Evaluation of MBEC-HTP biofilm model for studies of implant associated infections. J. Orthop. Res. 2012, 30, 1176–1180. [Google Scholar] [CrossRef] [PubMed]
- Molina-Manso, D.; del Prado, G.; Ortiz-Perez, A.; Manrubia-Cobo, M.; Gomez-Barrena, E.; Cordero-Ampuero, J.; Esteban, J. In vitro susceptibility to antibiotics of staphylococci in biofilms isolated from orthopaedic infections. Int. J. Antimicrob. Agents 2013, 41, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Girard, L.P.; Ceri, H.; Gibb, A.P.; Olson, M.; Sepandj, F. MIC versus mbec to determine the antibiotic sensitivity of Staphylococcus aureus in peritoneal dialysis peritonitis. Perit. Dial. Int. 2010, 30, 652–656. [Google Scholar] [CrossRef] [PubMed]
- LaPlante, K.L.; Mermel, L.A. In vitro activities of telavancin and vancomycin against biofilm-producing Staphylococcus aureus, S. epidermidis, and enterococcus faecalis strains. Antimicrob. Agents Chemother. 2009, 53, 3166–3169. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Ray, P.; Das, A.; Sharma, M. Penetration of antibiotics through Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Antimicrob. Chemother. 2010, 65, 1955–1958. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, K.K.; Goldmann, D.A.; Pier, G.B. Use of confocal microscopy to analyze the rate of vancomycin penetration through Staphylococcus aureus biofilms. Antimicrob. Agents Chemother. 2005, 49, 2467–2473. [Google Scholar] [CrossRef] [PubMed]
- Chaignon, P.; Sadovskaya, I.; Ragunah, C.; Ramasubbu, N.; Kaplan, J.B.; Jabbouri, S. Susceptibility of staphylococcal biofilms to enzymatic treatments depends on their chemical composition. Appl. Microbiol. Biotechnol. 2007, 75, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.B.; LoVetri, K.; Cardona, S.T.; Madhyastha, S.; Sadovskaya, I.; Jabbouri, S.; Izano, E.A. Recombinant human dnase I decreases biofilm and increases antimicrobial susceptibility in staphylococci. J. Antibiot. 2012, 65, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.B. Biofilm matrix-degrading enzymes. Methods Mol. Biol. 2014, 1147, 203–213. [Google Scholar] [PubMed]
- Nemoto, K.; Hirota, K.; Ono, T.; Murakami, K.; Murakami, K.; Nagao, D.; Miyake, Y. Effect of varidase (streptokinase) on biofilm formed by Staphylococcus aureus. Chemotherapy 2000, 46, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Vanassche, T.; Peetermans, M.; Van Aelst, L.N.; Peetermans, W.E.; Verhaegen, J.; Missiakas, D.M.; Schneewind, O.; Hoylaerts, M.F.; Verhamme, P. The role of staphylothrombin-mediated fibrin deposition in catheter-related Staphylococcus aureus infections. J. Infect. Dis. 2013, 208, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Kwiecinski, J.; Na, M.; Jarneborn, A.; Jacobsson, G.; Peetermans, M.; Verhamme, P.; Jin, T. Tissue plasminogen activator coating on implant surfaces reduces Staphylococcus aureus biofilm formation. Appl. Environ. Microbiol. 2016, 82, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The calgary biofilm device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [PubMed]
- Pietrocola, G.; Nobile, G.; Gianotti, V.; Zapotoczna, M.; Foster, T.J.; Geoghegan, J.A.; Speziale, P. Molecular interactions of human plasminogen with fnbpb, a fibrinogen/fibronectin-binding protein from Staphylococcus aureus. J. Biol. Chem. 2016, 291, 18148–18162. [Google Scholar] [CrossRef] [PubMed]
- Salazar, N.; Castiblanco-Valencia, M.M.; da Silva, L.B.; de Castro, I.; Monaris, D.; Masuda, H.P.; Barbosa, A.S.; Areas, A.P. Staphylococcus aureus manganese transport protein c (MNTC) is an extracellular matrix- and plasminogen-binding protein. PLoS ONE 2014, 9, e112730. [Google Scholar] [CrossRef] [PubMed]
- Shanks, R.M.; Donegan, N.P.; Graber, M.L.; Buckingham, S.E.; Zegans, M.E.; Cheung, A.L.; O’Toole, G.A. Heparin stimulates Staphylococcus aureus biofilm formation. Infect. Immun. 2005, 73, 4596–4606. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Lebeaux, D.; Ghigo, J.M.; Beloin, C. Full and broad-spectrum in vivo eradication of catheter-associated biofilms using gentamicin-edta antibiotic lock therapy. Antimicrob. Agents Chemother. 2012, 56, 6310–6318. [Google Scholar] [CrossRef] [PubMed]
- Estes, R.; Theusch, J.; Beck, A.; Pitrak, D.; Mullane, K.M. Activity of daptomycin with or without 25 percent ethanol compared to combinations of minocycline, edta, and 25 percent ethanol against methicillin-resistant Staphylococcus aureus isolates embedded in biofilm. Antimicrob. Agents Chemother. 2013, 57, 1998–2000. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Alhede, M.; Alhede, M.; Eickhardt-Sorensen, S.R.; Moser, C.; Kuhl, M.; Jensen, P.O.; Hoiby, N. The in vivo biofilm. Trends Microbiol. 2013, 21, 466–474. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, H.; Rudkin, J.K.; Black, N.S.; Gallagher, L.; O’Neill, E.; O’Gara, J.P. Methicillin resistance and the biofilm phenotype in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2015, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Stein, G.E.; LeBel, M.; Flor, S.C.; Zinny, M. Bioavailability and pharmacokinetics of oral ofloxacin formulations in normal subjects. Curr. Med. Res. Opin. 1991, 12, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, M.; Benziger, D.P.; Yankelev, S.; Vigliani, G. Pharmacokinetics and tolerability of daptomycin at doses up to 12 milligrams per kilogram of body weight once daily in healthy volunteers. Antimicrob. Agents Chemother. 2006, 50, 3245–3249. [Google Scholar] [CrossRef] [PubMed]
- Rybak, M.J.; Lomaestro, B.M.; Rotschafer, J.C.; Moellering, R.C.; Craig, W.A.; Billeter, M.; Dalovisio, J.R.; Levine, D.P. Vancomycin therapeutic guidelines: A summary of consensus recommendations from the infectious diseases society of america, the american society of health-system pharmacists, and the society of infectious diseases pharmacists. Clin. Infect. Dis. 2009, 49, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, R.; Lopez Cortes, L.E.; Molina, J.; Cisneros, J.M.; Pachon, J. Optimizing the clinical use of vancomycin. Antimicrob. Agents Chemother. 2016, 60, 2601–2609. [Google Scholar] [CrossRef] [PubMed]
- Chirouze, C.; Alla, F.; Fowler, V.G., Jr.; Sexton, D.J.; Corey, G.R.; Chu, V.H.; Wang, A.; Erpelding, M.L.; Durante-Mangoni, E.; Fernandez-Hidalgo, N.; et al. Impact of early valve surgery on outcome of Staphylococcus aureus prosthetic valve infective endocarditis: Analysis in the international collaboration of endocarditis-prospective cohort study. Clin. Infect. Dis. 2015, 60, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, B.D.; Tornos, P. Surgery for infective endocarditis: Who and when? Circulation 2010, 121, 1141–1152. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antibiotic | MBEC (−Streptokinase) (mg/L) | MBEC (+Streptokinase) (mg/L) |
|---|---|---|
| Daptomycin | 1024 | 1024 |
| Daptomycin + rifampicin | 64 | <4 |
| Vancomycin | 128 | 64 |
| Vancomycin + rifampicin | 64 | 4 |
| Antibiotic | MBEC (+Streptokinase) (mg/L) | MBEC (−Streptokinase) (mg/L) |
|---|---|---|
| Daptomycin + rifampicin | >1024 | >1024 |
| Vancomycin + rifampicin | >1024 | >1024 |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jørgensen, N.P.; Zobek, N.; Dreier, C.; Haaber, J.; Ingmer, H.; Larsen, O.H.; Meyer, R.L. Streptokinase Treatment Reverses Biofilm-Associated Antibiotic Resistance in Staphylococcus aureus. Microorganisms 2016, 4, 36. https://doi.org/10.3390/microorganisms4030036
Jørgensen NP, Zobek N, Dreier C, Haaber J, Ingmer H, Larsen OH, Meyer RL. Streptokinase Treatment Reverses Biofilm-Associated Antibiotic Resistance in Staphylococcus aureus. Microorganisms. 2016; 4(3):36. https://doi.org/10.3390/microorganisms4030036
Chicago/Turabian StyleJørgensen, Nis Pedersen, Natalia Zobek, Cindy Dreier, Jakob Haaber, Hanne Ingmer, Ole Halfdan Larsen, and Rikke L. Meyer. 2016. "Streptokinase Treatment Reverses Biofilm-Associated Antibiotic Resistance in Staphylococcus aureus" Microorganisms 4, no. 3: 36. https://doi.org/10.3390/microorganisms4030036