Zoonotic Fecal Pathogens and Antimicrobial Resistance in Canadian Petting Zoos

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey Data Collection
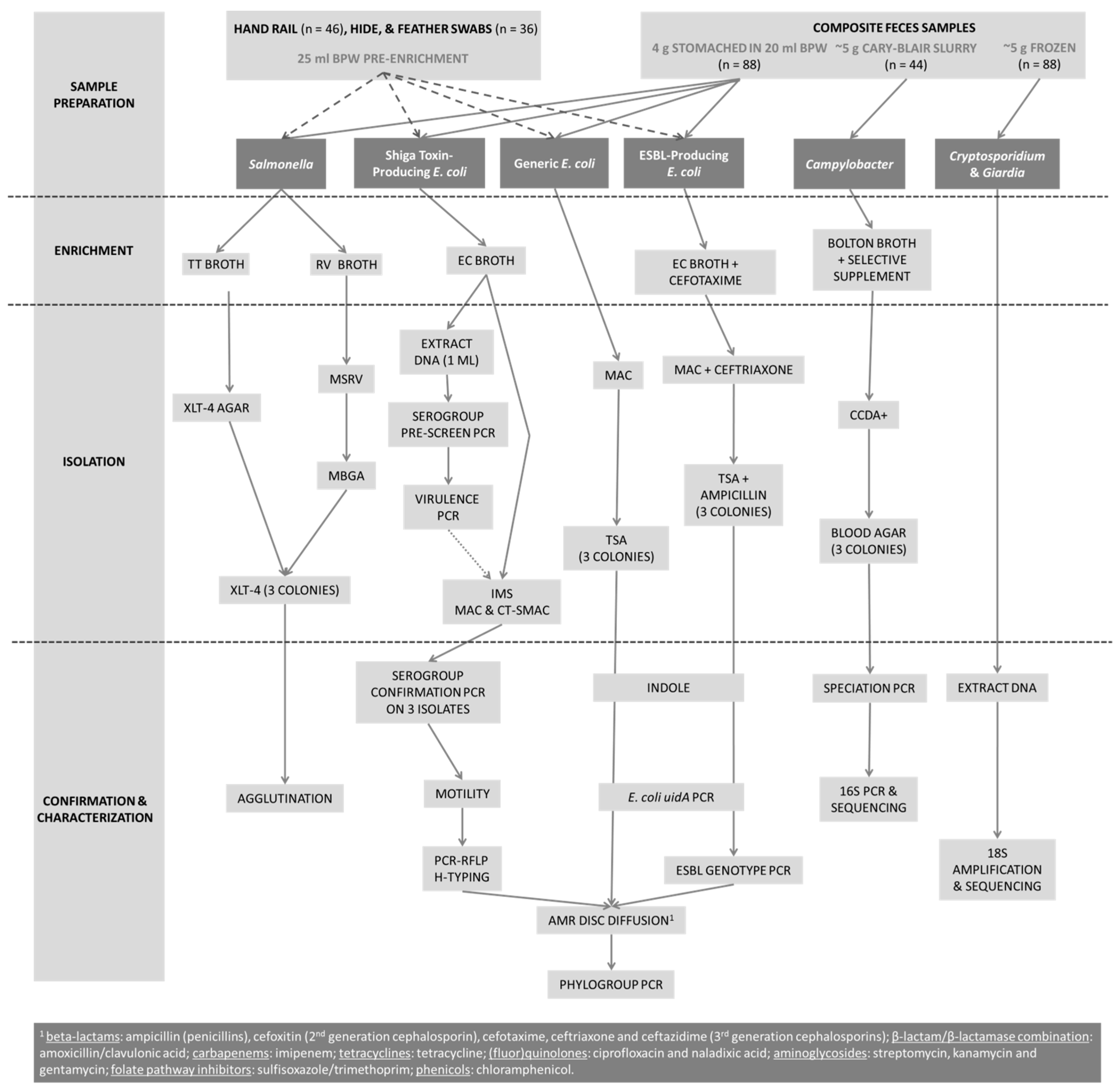
2.2. Sample Collection and Preparation
2.3. Salmonella Screening
2.4. STEC Screening
2.5. Generic E. coli Screening
2.6. Broad- and Extended-Spectrum β-Lactamase E. coli Screening
2.7. Campylobacter Screening
2.8. Cryptosporidium and Giardia Screening
2.9. MRSA Screening
2.10. Antimicrobial Resistance Testing
2.11. E. coli Isolate Phylogroups
2.12. Statistical Analysis
3. Results
3.1. Survey
3.2. Prevalence of O157 and Non-O157 STEC, Salmonella, Campylobacter, MRSA, Cryptosporidium and Giardia.
3.3. Screening for STEC and Related Genes
3.4. Screening for Broad- and Extended-Spectrum β-Lactamase E. coli
3.5. Antimicrobial Resistance among Generic E. coli, Broad- and Spectrum β-Lastamase E. coli, and STEC
3.6. Phylogroups among Generic E. coli and β-Lactamase-Producing E. coli
4. Discussion
4.1. Survey
4.2. Prevalence of O157 and Non-O157 STEC
4.3. Prevalence of Salmonella, MRSA, Campylobacter, Cryptosporidium and Giardia
4.4. AMR, Broad- and Extended-Spectrum β-Lactamase- E. coli
4.5. Phylogroups
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weese, J.S.; McCarthy, L.; Mossop, M.; Martin, H.; Lefebvre, S. Observation of practices at petting zoos and the potential impact on zoonotic disease transmission. Clin. Infect. Dis. 2007, 45, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Conrad, C.C.; Stanford, K.; Narvaez-Bravo, C.; Callaway, T.; McAllister, T. Farm fairs and petting zoos: A review of animal contact as a source of zoonotic enteric disease. Foodborne Pathog. Dis. 2017, 14, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Hale, C.R.; Scallan, E.; Cronquist, A.B.; Dunn, J.; Smith, K.; Robinson, T.; Lathrop, S.; Tobin-D’Angelo, M.; Clogher, P. Estimates of enteric illness attributable to contact with animals and their environments in the United States. Clin. Infect. Dis. 2012, 54, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Ferens, W.A.; Hovde, C.J. Escherichia coli O157:H7: Animal reservoir and sources of human infection. Foodborne Pathog. Dis. 2011, 8, 465–487. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.A.; Rubinelli, P.M.; Park, S.H.; Carbonero, F.; Ricke, S.C. Shiga toxin-producing Escherichia coli in food: Incidence, ecology, and detection strategies. Food Control 2016, 59, 407–419. [Google Scholar] [CrossRef]
- Bolton, D.J. Campylobacter virulence and survival factors. Food Microbiol. 2015, 48, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Erdozain, G.; KuKanich, K.; Chapman, B.; Powell, D. Observation of public health risk behaviours, risk communication and hand hygiene at Kansas and Missouri petting zoos. Zoonoses public health 2013, 60, 304–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.E.; Weese, J.S. Video observation of hand hygiene practices at a petting zoo and the impact of hand hygiene interventions. Epidemiol. Infect. 2012, 140, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Collignon, P.; Powers, J.H.; Chiller, T.M.; Aidara-Kane, A.; Aarestrup, F.M. World health organization ranking of antimicrobials according to their importance in human medicine: A critical step for developing risk management strategies for the use of antimicrobials in food production animals. Clin. Infect. Dis. 2009, 49, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-spectrum beta-lactamase-producing and ampc-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Roug, A.; Byrne, B.A.; Conrad, P.A.; Miller, W.A. Zoonotic fecal pathogens and antimicrobial resistance in county fair animals. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.T.; Sowers, E.G.; Wells, J.G.; Greene, K.D.; Griffin, P.M.; Hoekstra, R.M.; Strockbine, N.A. Non-O157 shiga toxin–producing Escherichia coli infections in the United States, 1983–2002. J. Infect. Dis. 2005, 192, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.; Johnson, R.P.; Alexander, T.W.; McAllister, T.A.; Reuter, T. Influence of season and feedlot location on prevalence and virulence factors of seven serogroups of Escherichia coli in feces of western-Canadian slaughter cattle. PloS ONE 2016, 11, e0159866. [Google Scholar] [CrossRef] [PubMed]
- Conrad, C.C.; Stanford, K.; McAllister, T.A.; Thomas, J.; Reuter, T. Further development of sample preparation and detection methods for O157 and the top 6 non-O157 STEC serogroups in cattle feces. J. Microbiol. Methods 2014, 105, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.; Grimont, F.; Grimont, P.A. Identification of Escherichia coli flagellar types by restriction of the amplified flic gene. Res. Microbiol. 2000, 151, 535–546. [Google Scholar] [CrossRef]
- Bej, A.K.; DiCesare, J.L.; Haff, L.; Atlas, R.M. Detection of Escherichia coli and Shigella spp. In water by using the polymerase chain reaction and gene probes for uid. Appl. Environ. Microbiol. 1991, 57, 1013–1017. [Google Scholar] [PubMed]
- Maheux, A.F.; Picard, F.J.; Boissinot, M.; Bissonnette, L.; Paradis, S.; Bergeron, M.G. Analytical comparison of nine PCR primer sets designed to detect the presence of Escherichia coli/Shigella in water samples. Water Res. 2009, 43, 3019–3028. [Google Scholar] [CrossRef] [PubMed]
- Dallenne, C.; Da Costa, A.; Decre, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Jokinen, C.C.; Koot, J.M.; Carrillo, C.D.; Gannon, V.P.; Jardine, C.M.; Mutschall, S.K.; Topp, E.; Taboada, E.N. An enhanced technique combining pre-enrichment and passive filtration increases the isolation efficiency of Campylobacter jejuni and Campylobacter coli from water and animal fecal samples. J. Microbiol. Methods 2012, 91, 506–513. [Google Scholar] [CrossRef] [PubMed]
- LaGier, M.J.; Joseph, L.A.; Passaretti, T.V.; Musser, K.A.; Cirino, N.M. A real-time multiplexed PCR assay for rapid detection and differentiation of Campylobacter jejuni and Campylobacter coli. Mol. Cell Probes 2004, 18, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Tani, K.; Nakamura, K.; Suzuki, Y.; Kitagawa, M.; Eguchi, M.; Nasu, M. Monitoring impact of in situ biostimulation treatment on groundwater bacterial community by DGGE. FEMS Microbiol. Ecol. 2000, 32, 129–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, M.; Matsutera, E.; Kanda, H.; Yamaguchi, N.; Tani, K.; Nasu, M. 16s ribosomal DNA-based analysis of bacterial diversity in purified water used in pharmaceutical manufacturing processes by PCR and denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 2002, 68, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Ruecker, N.J.; Hoffman, R.M.; Chalmers, R.M.; Neumann, N.F. Detection and resolution of Cryptosporidium species and species mixtures by genus-specific nested PCR-restriction fragment length polymorphism analysis, direct sequencing, and cloning. Appl. Environ. Microbiol. 2011, 77, 3998–4007. [Google Scholar] [CrossRef] [PubMed]
- Ruecker, N.J.; Matsune, J.C.; Lapen, D.R.; Topp, E.; Edge, T.A.; Neumann, N.F. The detection of Cryptosporidium and the resolution of mixtures of species and genotypes from water. Infect. Genet. Evol. 2013, 15, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, R.M.; Meloni, B.P.; Groth, D.M.; Wetherall, J.D.; Reynoldson, J.A.; Thompson, R.C.A. Ribosomal RNA sequencing reveals differences between the genotypes of Giardia isolates recovered from humans and dogs living in the same locality. J. Parasitol. 1997, 83, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Neumann, N.; Banting, G.; Klein, C.; Liccioli, S.; Massolo, A. Giardia spp. And cryptosporidium spp. Infecting urban dogs and coyote pose a low risk for public. Zoonoses 2017. Under Review. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disc Susceptibility Tests; Approved Standard—Twelfth Edition M02-A12; CLSI: Wayne, PA, USA, 2015. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing; M100-S26; CLSI: Wayne, PA, USA, 2016. [Google Scholar]
- Franz, E.; Veenman, C.; van Hoek, A.; Husman, A.d.R.; Blaak, H. Pathogenic Escherichia coli producing extended-spectrum β-lactamases isolated from surface water and wastewater. Sci. Rep. 2015, 5, 14372. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.B.; Shulman, S.A. Reports of zoonotic disease outbreaks associated with animal exhibits and availability of recommendations for preventing zoonotic disease transmission from animals to people in such settings. J. Am. Vet. Med. Assoc. 2004, 224, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- LeJeune, J.T.; Davis, M.A. Outbreaks of zoonotic enteric disease associated with animal exhibits. J. Am. Vet. Med. Assoc. 2004, 224, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- National Association of State Public Health Veterinarians. Compendium of measures to prevent disease associated with animals in public settings. J. Am. Vet. Med. Assoc. 2013, 243, 1270–1288. [Google Scholar]
- Keen, J.E.; Wittum, T.E.; Dunn, J.R.; Bono, J.L.; Durso, L.M. Shiga-toxigenic Escherichia coli O157 in agricultural fair livestock, United States. Emerg. Infect. Dis. 2006, 12, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Goode, B.; O’Reilly, C.; Dunn, J.; Fullerton, K.; Smith, S.; Ghneim, G.; Keen, J.; Durso, L.; Davies, M.; Montgomery, S. Outbreak of Escherichia coli O157: H7 infections after petting zoo visits, North Carolina state fair, October-November 2004. Arch. Pediatr. Adolesc. Med. 2009, 163, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Varma, J.K.; Greene, K.D.; Reller, M.E.; DeLong, S.M.; Trottier, J.; Nowicki, S.F.; DiOrio, M.; Koch, E.M.; Bannerman, T.L.; York, S.T.; et al. An outbreak of Escherichia coli O157 infection following exposure to a contaminated building. Jama 2003, 290, 2709–2712. [Google Scholar] [CrossRef] [PubMed]
- Steinmuller, N.; Demma, L.; Bender, J.B.; Eidson, M.; Angulo, F.J. Outbreaks of enteric disease associated with animal contact: Not just a foodborne problem anymore. Clin. Infect. Dis. 2006, 43, 1596–1602. [Google Scholar] [CrossRef] [PubMed]
- Karmali, M.A.; Gannon, V.; Sargeant, J.M. Verocytotoxin-producing Escherichia coli (VTEC). Vet. Microbiol. 2010, 140, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Chandran, A.; Mazumder, A. Prevalence of diarrhea-associated virulence genes and genetic diversity in Escherichia coli isolates from fecal material of various animal hosts. Appl. Environ. Microbiol. 2013, 79, 7371–7380. [Google Scholar] [CrossRef] [PubMed]
- Rostagno, M.H. Can stress in farm animals increase food safety risk? Foodborne Pathog. Dis. 2009, 6, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Besser, T.E.; Goldoft, M.; Pritchett, L.C.; Khakhria, R.; Hancock, D.D.; Rice, D.H.; Gay, J.M.; Johnson, W.; Gay, C.C. Multiresistant Salmonella Typhimurium DT104 infections of humans and domestic animals in the pacific northwest of the United States. Epidemiol. Infect. 2000, 124, 193–200. [Google Scholar] [CrossRef]
- Sorensen, O.; Van Donkersgoed, J.; McFall, M.; Manninen, K.; Gensler, G.; Ollis, G. Salmonella spp. Shedding by Alberta beef cattle and the detection of Salmonella spp. in ground beef. J. Food Prot. 2002, 65, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Liljebjelke, K.A.; Hofacre, C.L.; Liu, T.; White, D.G.; Ayers, S.; Young, S.; Maurer, J.J. Vertical and horizontal transmission of Salmonella within integrated broiler production system. Foodborne Pathog. Dis. 2005, 2, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Dressler, A.E.; Scheibel, R.P.; Wardyn, S.; Harper, A.L.; Hanson, B.M.; Kroeger, J.S.; Diekema, D.J.; Bender, J.B.; Gray, G.C.; Smith, T.C. Prevalence, antibiotic resistance and molecular characterisation of Staphylococcus aureus in pigs at agricultural fairs in the USA. Vet. Rec. 2012, 170, 495. [Google Scholar] [CrossRef] [PubMed]
- Khanna, T.; Friendship, R.; Dewey, C.; Weese, J.S. Methicillin resistant Staphylococcus aureus colonization in pigs and pig farmers. Vet. Microbiol. 2008, 128, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Oporto, B.; Esteban, J.I.; Aduriz, G.; Juste, R.A.; Hurtado, A. Prevalence and strain diversity of thermophilic campylobacters in cattle, sheep and swine farms. J. Appl. Microbiol. 2007, 103, 977–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriarty, E.M.; Downing, M.; Bellamy, J.; Gilpin, B.J. Concentrations of faecal coliforms, Escherichia coli, Enterococci and Campylobacter spp. In equine faeces. N. Z. Vet. J. 2015, 63, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Pintar, K.D.; Christidis, T.; Thomas, M.K.; Anderson, M.; Nesbitt, A.; Keithlin, J.; Marshall, B.; Pollari, F. A systematic review and meta-analysis of the Campylobacter spp. prevalence and concentration in household pets and petting zoo animals for use in exposure assessments. PloS ONE 2015, 10, e0144976. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Fayer, R.; Xiao, L. Cryptosporidium species in humans and animals: Current understanding and research needs. Parasitology 2014, 141, 1667–1685. [Google Scholar] [CrossRef] [PubMed]
- Gow, S.P.; Waldner, C.L.; Harel, J.; Boerlin, P. Associations between antimicrobial resistance genes in fecal generic Escherichia coli isolates from cow-calf herds in western Canada. Appl. Environ. Microbiol. 2008, 74, 3658–3666. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.; Menzies, P.; Reid-Smith, R.J.; Avery, B.P.; McEwen, S.A.; Moon, C.S.; Berke, O. Antimicrobial resistance in fecal generic Escherichia coli and Salmonella spp. obtained from Ontario sheep flocks and associations between antimicrobial use and resistance. Can. J. Vet. Res. 2012, 76, 109–119. [Google Scholar] [PubMed]
- Aarestrup, F.M.; Oliver Duran, C.; Burch, D.G. Antimicrobial resistance in swine production. Anim. Health Res. Rev. 2008, 9, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Benedict, K.M.; Gow, S.P.; McAllister, T.A.; Booker, C.W.; Hannon, S.J.; Checkley, S.L.; Noyes, N.R.; Morley, P.S. Antimicrobial resistance in Escherichia coli recovered from feedlot cattle and associations with antimicrobial use. PloS ONE 2015, 10, e0143995. [Google Scholar] [CrossRef] [PubMed]
- Sayah, R.S.; Kaneene, J.B.; Johnson, Y.; Miller, R. Patterns of antimicrobial resistance observed in Escherichia coli isolates obtained from domestic- and wild-animal fecal samples, human septage, and surface water. Appl. Environ. Microbiol. 2005, 71, 1394–1404. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, R.R.; Henderson, I.R. The evolution of the Escherichia coli phylogeny. Infect. Genet. Evol. 2012, 12, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Carlos, C.; Pires, M.M.; Stoppe, N.C.; Hachich, E.M.; Sato, M.I.; Gomes, T.A.; Amaral, L.A.; Ottoboni, L.M. Escherichia coli phylogenetic group determination and its application in the identification of the major animal source of fecal contamination. BMC Microbiol. 2010, 10, 161–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Question | Answer | % Fairs | |
|---|---|---|---|
| How many visitors attend the fair each year? (n = 185) | under 5000 visitors | 36 | |
| 5000–19,999 visitors | 36 | ||
| 20,000–99,999 visitors | 20 | ||
| 100,000 and over | 8 | ||
| Where is the fair located? (n = 185) | Alberta | 14 | |
| British Columbia | 11 | ||
| Manitoba | 5 | ||
| Ontario | 49 | ||
| Quebec | 9 | ||
| Other | 12 | ||
| Is there a petting zoo featured at the fair? (n = 171) | Yes | 71 | |
| Is there a livestock pavilion or barn? (n = 184) | Yes | 72 | |
| Has there been an outbreak or problem with a pathogen in the past 10 years? (n = 164) | Yes | 2 | |
| What was the pathogen of concern? (among those that reported a problem or outbreak) (n = 4) | E. coli | 75 | |
| Unknown | 25 | ||
| Question | Answer | % Livestock Pavilions at fairs | % Petting Zoos at fairs |
| Which animal species are present at the facility? (L n = 131; P n = 119) | Cattle and Calves | 89 | 44 |
| Horses and Ponies | 77 | 71 | |
| Sheep | 69 | 72 | |
| Goats | 55 | 78 | |
| Rabbits | 62 | 73 | |
| Pigs | 40 | 52 | |
| Donkeys | NA | 52 | |
| Alpacas or Llamas | NA | 36 | |
| Guinea Pigs | NA | 19 | |
| What is the level of contact between humans and animals within petting zoos? (n = 117) | Animals roam freely without barriers and visitors can pet, hold, and feed the animals | NA | 17 |
| Animals roam freely without barriers visitors can pet and hold the animals | NA | 5 | |
| A barrier separates animals and visitors, and visitors can pet and feed the animals | NA | 44 | |
| A barrier separates animals and visitors, and visitors can only pet the animals | NA | 34 | |
| What is the level of contact between humans and animals at the livestock pavilion? (n = 131) | None, only exhibitors have contact with the animals | 11 | NA |
| Public is permitted to walk through the livestock area | 71 | NA | |
| Public is encouraged to touch or interact with the animals | 18 | NA | |
| Is there a children’s play area directly adjacent? (L n = 130; P n = 115) | Yes | 18 | 30 |
| Is there a food or beverage area directly adjacent? (L n = 131; P n = 116) | Yes | 16 | 2 |
| Are there rules regarding visitors bringing food or drink into the animal area? (L n = 125; P n = 117) | Not allowed | 29 | 49 |
| Allowed, but no touching animals | 7 | 5 | |
| Allowed, not for feeding animals | 25 | 25 | |
| No restrictions | 39 | 21 | |
| Is there continuous removal of feces? (L n = 103; P n = 84) | Yes | 78 | 69 |
| What is the frequency of removal of feces from the animal area? (L n = 117; P n = 111) | Immediately | 57 | 60 |
| 2–4 times daily | 32 | 32 | |
| Once daily | 8 | 8 | |
| Less frequently | 3 | 0 | |
| Are there hand-washing stations? (L n = 123; P n = 117) | Yes | 73 | 69 |
| Are there hand sanitizers? (L n = 96; P n = 96) | Yes | 73 | 79 |
| Does the staff remind visitors to wash their hands? (L n = 117; P n = 111) | Yes | 44 | 69 |
| Is there hand-washing signage? (L n = 97; P n = 88) | Yes | 73 | 72 |
| Is there no smoking signage? (L n = 89; P n = 63) | Yes | 67 | 52 |
| Is there no eating signage? (L n = 39; P n = 43) | Yes | 30 | 35 |
| How often are surfaces sanitized? (L n = 111; P n = 107) | Multiple times a day | 18 | 30 |
| Once daily | 19 | 20 | |
| Only when visibly dirty | 32 | 27 | |
| Never | 31 | 23 | |
| Is there a safe area for storage of visitor items? (L n = 117; P n = 113) | Yes | 25 | 30 |
| Sample Type | Host | % STEC | % Salmonella | % Campylobacter 1 | % Cryptosporidium | % Giardia 6 | |||
|---|---|---|---|---|---|---|---|---|---|
| O157 | Non-O157 | jejuni | coli | lanienae | |||||
| Composite Fecal Samples | Cattle | 0 (0/9) | 44 (4/9) | 0 (0/9) | 0 (0/5) | 0 (0/5) | 0 (0/5) | 0 (0/9) | 22 (2/9) |
| Sheep and Goats | 3 (1/36) | 19 (7/36) | 0 (0/36) | 7 (1/15) | 7 (1/15) | 0 (0/15) | 6 (2/36) 5 | 25 (9/36) | |
| Pigs | 0 (0/12) | 33 (4/12) | 0 (0/12) | 0 (0/10) | 20 (2/10) | 30 (3/10) | 0 (0/12) | 0 (0/12) | |
| Horses and Donkeys | 0 (0/18) | 28 (5/18) | 0 (0/18) | 0 (0/8) | 38 (3/8) 2 | 25 (2/8) 2 | 0 (0/18) | 11 (2/18) | |
| Llamas | 0 (0/6) | 100 (6/6) | 0 (0/6) | NT 4 | NT 4 | NT 4 | 0 (0/6) | 0 (0/6) | |
| Birds 3 | 0 (0/7) | 29 (2/7) | 0 (0/7) | 17 (1/6) | 17 (1/6) | 0 (0/6) | 0 (0/7) | 0 (0/7) | |
| Total | 1 (1/88) | 32 (28/88) | 0 (0/88) | 5 (2/44) | 16 (7/44) | 11 (5/44) | 2 (2/88) | 15 (13/88) | |
| Hide/Feather Swabs | Cattle | 0 (0/4) | 0 (0/4) | 0 (0/4) | NT 4 | NT 4 | NT 4 | NT 4 | NT 4 |
| Sheep and Goats | 0 (0/15) | 7 (1/15) | 0 (0/15) | NT 4 | NT 4 | NT 4 | NT 4 | NT 4 | |
| Pigs | 0 (0/5) | 20 (1/5) | 0 (0/5) | NT 4 | NT 4 | NT 4 | NT 4 | NT 4 | |
| Horses and Donkeys | 0 (0/5) | 0 (0/5) | 0 (0/5) | NT 4 | NT 4 | NT 4 | NT 4 | NT 4 | |
| Llamas | 0 (0/3) | 0 (0/3) | 0 (0/3) | NT 4 | NT 4 | NT 4 | NT 4 | NT 4 | |
| Birds 3 | 0 (0/3) | 100 (3/3) | 0 (0/3) | NT 4 | NT 4 | NT 4 | NT 4 | NT 4 | |
| Rabbits | 0 (0/1) | 0 (0/1) | 0 (0/1) | NT 4 | NT 4 | NT 4 | NT 4 | NT 4 | |
| Total | 0 (0/36) | 14 (5/36) | 0 (0/36) | ||||||
| Hand Rail Swabs | Wood | 0 (0/28) | 14 (4/28) | 0 (0/28) | NT 4 | NT 4 | NT 4 | NT 4 | NT 4 |
| Metal | 0 (0/18) | 0 (0/18) | 0 (0/18) | NT 4 | NT 4 | NT 4 | NT 4 | NT 4 | |
| Total | 0 (0/46) | 9 (4/46) | 0 (0/46) | ||||||
| Grand Total | 0.6 (1/170) | 22 (37/170) | 0 (0/170) | 5 (2/44) | 16 (7/44) | 11 (5/44) | 2 (2/88) 5 | 15 (13/88) 6 | |
| Serogroup | H-Type | Motility 1 | Petting Zoo ID 2 | Sample Type | Host | Virulence Markers | Phylogenetic Group 3 | # of Isolates | Antibiotic Resistance Pattern (# Antibiotic Classes) 4,5 |
|---|---|---|---|---|---|---|---|---|---|
| O26 | H34 | + | A2 | feces | cattle | ehxA, eae, stx1 | U | 1 | NA |
| O26 | H46 | + | A2 | feces | cattle | stx1 | U | 1 | KAN-STREP (1) |
| O26 | H46 | + | A2 | feces | cattle | ehxA, stx1 | U | 1 | KAN-STREP (1) |
| O26 | H34 | + | A2 | feces | cattle | stx1 | Multiple 6 | 2 | NA |
| O26 | H46 | + | F | feces | donkey | ehxA, eae, stx1, stx2 | A | 1 | TET (1) |
| O26 | H46 | + | A1 | feces | donkey | eae, stx1 | U | 1 | KAN-STREP (1) |
| O26 | H46 | + | A1 | feces | donkey | ehxA, stx1 | U | 1 | KAN-STREP (1) |
| O26 | H46 | + | A1, F | feces | donkey | stx1 | U | 47 | KAN-STREP (1) |
| O26 | H46 | + | A2 | feces | donkey | stx1 | A | 2 | TET (1) |
| O26 | H11 | + | C1 | feces | goat | ehxA, eae, stx1 | U | 27 | NA |
| O26 | H46 | + | A2 | feces | goat | eae, stx1 | U | 1 | KAN-STREP (1) |
| O26 | H46 | + | F | feces | goat | stx1, stx2 | A | 1 | TET (1) |
| O26 | H46 | + | F | feces | horse | eae, stx1, stx2 | A | 1 | TET (1) |
| O26 | H34 | + | A2 | feces | llama | stx1 | B2 | 27 | NA |
| O26 | H46 | + | F | feces | pig | stx1, stx2 | A | 37 | TET (1) |
| O26 | H46 | + | F | feather swab | chicken | stx1 | A | 1 | TET (1) |
| O26 | H46 | + | F | hand rail | wood | stx1 | A | 1 | NA |
| O45 | H34 | + | A2 | feces | llama | eae, stx1 | U | 1 | NA |
| O45 | H21 | NM | F | hand rail | wood | ehxA, eae, stx1 | U | 1 | CHL-STREP-TET-TMS (4) |
| O45 | H19 | NM | F | feather swab | chicken | stx1 | E | 1 | AMC-AMP-CAZ-CFX-CHL-CRO-CTX-GEN-STREP-TET (5) |
| O103 | H43 | + | B | feces | cattle | ehxA, eae, stx2 | B1 | 1 | NA |
| O103 | H43 | + | B | feces | cattle | stx2 | U | 1 | NA |
| O103 | H43 | + | B | feces | cattle | stx1 | B1 | 1 | NA |
| O103 | H38 | + | F | feces | donkey | ehxA, eae, stx1 | B1 | 2 | NA |
| O103 | H38 | + | F | feces | duck | ehxA, eae, stx1 | Multiple 8 | 3 7 | NA |
| O103 | H38 | + | F | feces | goat | ehxA, eae, stx1 | B1 | 2 | NA |
| O103 | H43 | + | B | feces | sheep or goat | stx2 | B1 | 1 | NA |
| O103 | H43 | + | B | feces | sheep or goat | stx1 | E | 1 | NA |
| O103 | H19 | + | B | feces | sheep or goat | stx2 | U | 1 | NA |
| O103 | H38 | + | F | feces | horse | ehxA, eae, stx1 | B1 | 1 | NA |
| O103 | H21 | + | F | feces | horse | stx1 | B1 | 1 | NA |
| O103 | H38 | + | F | feces | pig | ehxA, eae, stx1 | B1 | 1 | NA |
| O103 | H19 | + | C2 | feces | pig | stx2 | B1 | 1 | NA |
| O103 | H2 | + | A2 | hand rail | wood | eae, stx1 | Multiple 9 | 3 | STREP-TET-TMS (3) |
| O103 | H4 | + | F, C2 | hand rail | wood | eae, stx1 | B1 | 3 7 | NA |
| O103 | H42 | + | C2 | hand rail | wood | eae, stx1 | B1 | 1 | NA |
| O103 | H38 | + | F | feather swab | duck | ehxA, eae, stx1 | B1 | 1 | NA |
| O103 | H38 | + | C1 | hide swab | goat | ehxA, eae, stx1 | E | 1 | TET (1) |
| O121 | H19 | + | A1 | feces | llama | eae, stx2 | B1 | 1 | NA |
| O121 | H7 | + | C2 | feces | pig | stx1 | B1 | 1 | TET (1) |
| O145 | H25 | + | A1 | feces | llama | eae, stx1 | Multiple 10 | 3 7 | NA |
| O145 | H28 | NM | F | feces | pig | eae, stx1 | E | 1 | NA |
| O145 | H25 | + | E | hide swab | pig | eae, stx1 | B2 | 2 | NA |
| O157 | H7 | + | C1 | feces | goat | ehxA, eae, stx1, stx2 | E | 3 | NA |
| ESBL Genotype | Animal Group | Phylogenetic Group 1 | # Isolates |
|---|---|---|---|
| blaCTX-M-1 | cattle | U | 3 |
| blaCTX-M-15 | birds | A | 1 |
| birds | B1 | 1 | |
| birds | U | 1 | |
| goats | B1 | 3 | |
| pigs | B1 | 2 | |
| blaTEM-1 | cattle | B1 | 4 |
| cattle | E | 1 | |
| cattle | U | 1 | |
| donkeys | F | 1 | |
| pigs | E | 3 |
| Composite Fecal Samples | Hide or Feather Swabs | Hand Rail Swabs | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMR Pattern (# Antibiotic Classes) 1 | Cattle | Sheep/Goats | Pigs | Horses/Donkeys | Llamas | Birds | Cattle | Sheep/Goats | Pigs | Horses/Donkeys | Llamas | Birds | Wood | Metal |
| Generic E. coli (n= # isolates) | n = 25 | n = 108 | n = 35 | n = 52 | n = 18 | n = 18 | n = 12 | n = 33 | n = 8 | n = 14 | n = 9 | n = 7 | n = 66 | n = 20 |
| AMC-AMP-CAZ-CFX-CHL-CRO-CTX-GEN-KAN-STREP-TET-TMS (6) | 1 2 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| AMC-AMP-CAZ-CFX-CHL-CRO-CTX-KAN-STREP-TET-TMS (6) | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| AMC-AMP-CAZ-CFX-CHL-CRO-CTX-GEN-KAN-STREP-TET (5) | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| AMC-AMP-KAN-STREP-TET (4) | - | 3 | - | - | - | - | - | - | - | - | - | - | - | - |
| AMP-STREP-TET-TMS (4) | - | - | - | - | 1 | - | - | - | - | - | - | - | 4 | - |
| CHL-STREP-TET-TMS (4) | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| CHL-STREP-TET (3) | - | - | 1 3 | - | - | - | - | - | - | - | - | - | 2 | - |
| CHL-STREP-TMS (3) | - | - | - | 1 4 | - | - | - | - | - | - | - | - | - | - |
| CTX-CRO (1) | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - |
| KAN-STREP (1) | - | 1 | - | 3 | - | - | - | - | - | - | - | - | - | - |
| STREP-TET (2) | 1 | 3 | 6 | - | - | - | - | 1 | - | 2 | - | 2 | - | - |
| STREP-TMS (2) | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - |
| TET-TMS (2) | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| STREP (1) | - | 1 | 5 | - | - | - | - | - | - | - | - | - | - | - |
| TET (1) | 2 | 10 | 2 | 4 | - | 5 | - | 9 | 1 | 1 | - | - | 7 | 3 |
| AMP (1) | 1 | - | 1 | - | 4 | - | - | 1 | - | - | - | - | - | - |
| Susceptible isolates | 16 | 90 | 19 | 43 | 13 | 13 | 0 | 22 | 7 | 11 | 8 | 5 | 52 | 17 |
| General resistance prevalence per sample (%) 5 | 5/9 (56) | 9/36 (25) | 10/12 (83) | 4/18 (22) | 3/6 (50) | 3/7 (43) | 0/4 (0) | 5/15 (33) | 1/5 (20) | 2/5 (40) | 1/3 (33) | 2/3 (66) | 8/28 (29) | 1/18 (6) |
| AMC-AMP-CAZ-CFX-CHL-CIP-CRO-CTX-NAL-STREP-TET-TMS (7) | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| AMC-AMP-CAZ-CFX-CHL-CRO-CTX-GEN-KAN-STREP-TET (5) | 6 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| AMP-CAZ-CFX-CHL-CIP-CRO-CTX-NAL-STREP-TET-TMS (6) | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - |
| AMC-AMP-CAZ-CFX-CHL-CRO-CTX-NAL-STREP-TET (6) | - | 6 | - | - | - | - | - | - | - | - | - | - | 3 | - |
| AMC-AMP-CAZ-CFX-CHL-CRO-CTX-STREP-TET-TMS (6) | - | 3 | 4 | - | - | - | - | - | - | - | - | - | - | - |
| AMP-CAZ-CHL-CIP-CRO-CTX-NAL-STREP-TET-TMS (6) | - | - | 1 6 | - | - | - | - | - | - | - | - | - | - | - |
| AMC-AMP-CAZ-CFX-CHL-CRO-CTX-STREP-TET (5) | 3 | 3 | 2 | 3 | - | - | - | - | - | - | - | - | - | - |
| AMP-CHL-CIP-CRO-CTX-NAL-STREP-TET-TMS (6) | - | 1 | 2 | - | - | 2 | - | - | - | - | - | - | - | - |
| AMC-AMP-CAZ-CFX-CRO-CTX-STREP-TET (4) | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - |
| AMP-CAZ-CHL-CRO-CTX-STREP-TET-TMS (5) | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| AMC-AMP-CAZ-CFX-CRO-CTX-TET (3) | - | - | 4 | - | - | - | - | - | - | - | - | - | - | - |
| AMC-AMP-CFX-CHL-CTX-STREP-TET (5) | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - |
| AMC-AMP-CAZ-CFX-CRO-CTX (2) | - | 6 | - | - | - | - | - | - | - | - | - | - | - | - |
| AMP-CRO-CTX-TET (2) | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - |
| AMP-CRO-CTX (1) | 3 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| General resistance prevalence per sample (%) 5 | 4/9 (44) | 7/36 (19) | 7/12 (58) | 1/18 (6) | 0/0 (0) | 1/7 (14) | 0/0 (0) | 0/0 (0) | 0/0 (0) | 0/0 (0) | 0/0 (0) | 0/0 (0) | 1/28 (4) | 0/0 (0) |
| AMC-AMP-CAZ-CFX-CHL-CRO-CTX-GEN-STREP-TET (5) | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - |
| CHL-STREP-TET-TMS (4) | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - |
| STREP-TET-TMS (3) | - | - | - | - | - | - | - | - | - | - | - | - | 3 | - |
| KAN-STREP (1) | 2 | 1 | - | 6 | - | - | - | - | - | - | - | - | - | - |
| TET (1) | - | 1 | 4 | 4 | - | - | - | 1 | - | - | - | 1 | - | - |
| Susceptible isolates | 6 | 10 | 3 | 4 | 7 | 3 | 0 | 0 | 2 | 0 | 0 | 1 | 5 | 0 |
| General resistance prevalence per sample (%) 5 | 1/9 (11) | 2/36 (6) | 3/12 (25) | 5/18 (28) | 0/6 (0) | 0/7 (0) | 0/4 (0) | 1/15 (7) | 0/5 (0) | 0/5 (0) | 0/3 (0) | 2/3 (66) | 2/28 (7) | 0/18 (0) |
| Phylogroup | Cattle | Sheep/Goats | Pigs | Horses/Donkeys | Llamas | Birds | Wood | Metal | % Isolates | |
|---|---|---|---|---|---|---|---|---|---|---|
| Generic E. coli isolates from animal feces (n = 256) | A | 5 | 15 | 5 | 1 | 1 | 2 | NA | NA | 11 |
| B1 | 17 | 67 | 25 | 33 | 13 | 13 | NA | NA | 66 | |
| B2 | 0 | 0 | 0 | 0 | 0 | 0 | NA | NA | 0 | |
| C | 0 | 7 | 0 | 7 | 0 | 1 | NA | NA | 6 | |
| D | 0 | 1 | 1 | 3 | 0 | 0 | NA | NA | 2 | |
| E | 2 | 7 | 2 | 3 | 4 | 1 | NA | NA | 7 | |
| F | 0 | 0 | 0 | 0 | 0 | 0 | NA | NA | 0 | |
| Clade I or II | 0 | 3 | 1 | 0 | 0 | 0 | NA | NA | 2 | |
| Unknown | 1 | 8 | 1 | 5 | 0 | 1 | NA | NA | 6 | |
| Generic E. coli isolates from animal hide swabs (n = 83) | A | 1 | 1 | 1 | 0 | 0 | 7 | NA | NA | 12 |
| B1 | 8 | 23 | 6 | 12 | 5 | 0 | NA | NA | 65 | |
| B2 | 0 | 0 | 0 | 0 | 2 | 0 | NA | NA | 2 | |
| C | 1 | 3 | 1 | 0 | 0 | 0 | NA | NA | 6 | |
| D | 0 | 0 | 0 | 1 | 0 | 0 | NA | NA | 1 | |
| E | 1 | 3 | 0 | 1 | 2 | 0 | NA | NA | 8 | |
| F | 0 | 0 | 0 | 0 | 0 | 0 | NA | NA | 0 | |
| Clade I or II | 0 | 1 | 0 | 0 | 0 | 0 | NA | NA | 1 | |
| Unknown | 1 | 2 | 0 | 0 | 0 | 0 | NA | NA | 4 | |
| Generic E. coli isolates from hand rail swabs (n = 86) | A | NA | NA | NA | NA | NA | NA | 8 | 3 | 13 |
| B1 | NA | NA | NA | NA | NA | NA | 39 | 12 | 59 | |
| B2 | NA | NA | NA | NA | NA | NA | 0 | 0 | 0 | |
| C | NA | NA | NA | NA | NA | NA | 2 | 0 | 2 | |
| D | NA | NA | NA | NA | NA | NA | 0 | 0 | 0 | |
| E | NA | NA | NA | NA | NA | NA | 14 | 5 | 22 | |
| F | NA | NA | NA | NA | NA | NA | 0 | 0 | 0 | |
| Clade I or II | NA | NA | NA | NA | NA | NA | 3 | 0 | 3 | |
| Unknown | NA | NA | NA | NA | NA | NA | 0 | 0 | 0 | |
| B-lactamase -producing E. coli isolates from feces and hand rail swabs (n = 59) | A | 0 | 0 | 3 | 0 | 0 | 1 | 0 | 0 | 7 |
| B1 | 4 | 12 | 7 | 0 | 0 | 1 | 0 | 0 | 41 | |
| B2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| C | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| D | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1.5 | |
| E | 4 | 9 | 3 | 3 | 0 | 0 | 3 | 0 | 37 | |
| F | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1.5 | |
| Clade I or II | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Unknown | 4 | 0 | 2 | 0 | 0 | 1 | 0 | 0 | 12 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conrad, C.C.; Stanford, K.; Narvaez-Bravo, C.; Neumann, N.F.; Munns, K.; Tymensen, L.; Jokinen, C.; McAllister, T.A. Zoonotic Fecal Pathogens and Antimicrobial Resistance in Canadian Petting Zoos. Microorganisms 2018, 6, 70. https://doi.org/10.3390/microorganisms6030070
Conrad CC, Stanford K, Narvaez-Bravo C, Neumann NF, Munns K, Tymensen L, Jokinen C, McAllister TA. Zoonotic Fecal Pathogens and Antimicrobial Resistance in Canadian Petting Zoos. Microorganisms. 2018; 6(3):70. https://doi.org/10.3390/microorganisms6030070
Chicago/Turabian StyleConrad, Cheyenne C., Kim Stanford, Claudia Narvaez-Bravo, Norman F. Neumann, Krysty Munns, Lisa Tymensen, Cassandra Jokinen, and Tim A. McAllister. 2018. "Zoonotic Fecal Pathogens and Antimicrobial Resistance in Canadian Petting Zoos" Microorganisms 6, no. 3: 70. https://doi.org/10.3390/microorganisms6030070