Mycotoxins in Flanders’ Fields: Occurrence and Correlations with Fusarium Species in Whole-Plant Harvested Maize

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
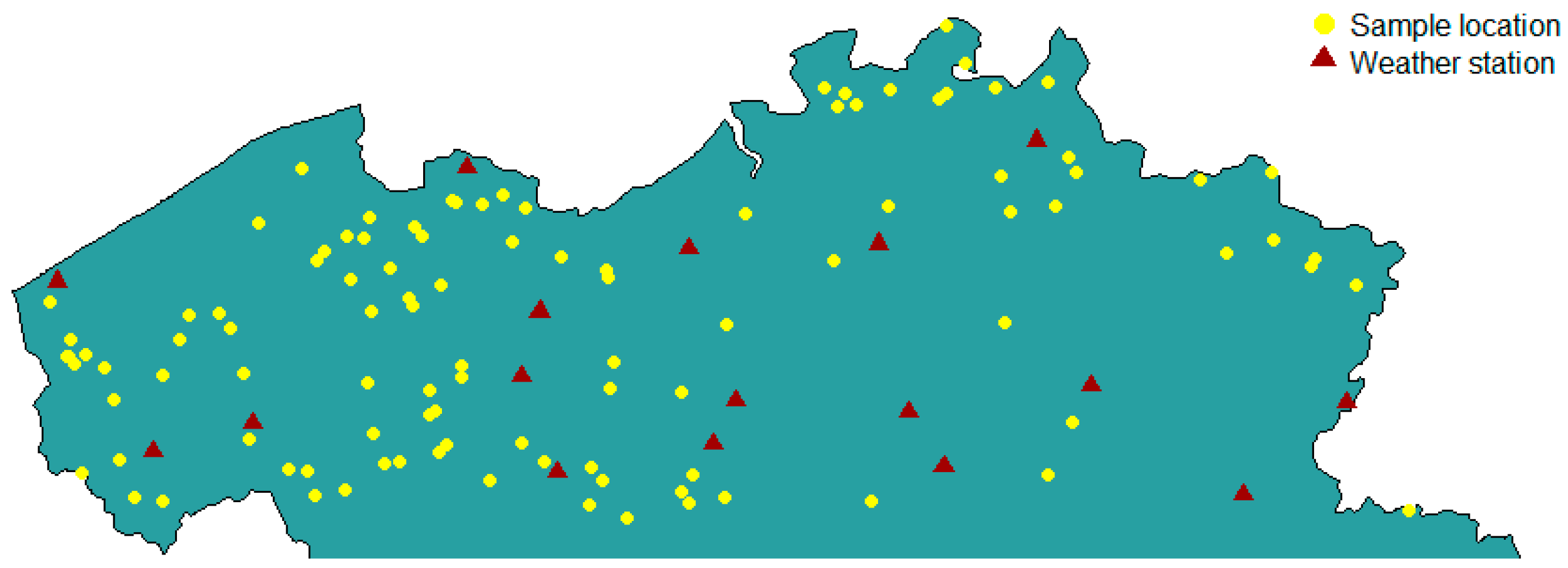
2.1. Maize Sampling
2.2. Reagents and Chemicals for LC-MS/MS
2.3. Sample Preparation for LC-MS/MS
2.4. Mycotoxin Analysis by LC-MS/MS
2.5. LC-MS/MS Quality Control
2.6. qPCR Analysis
2.7. Statistical Analysis
3. Results
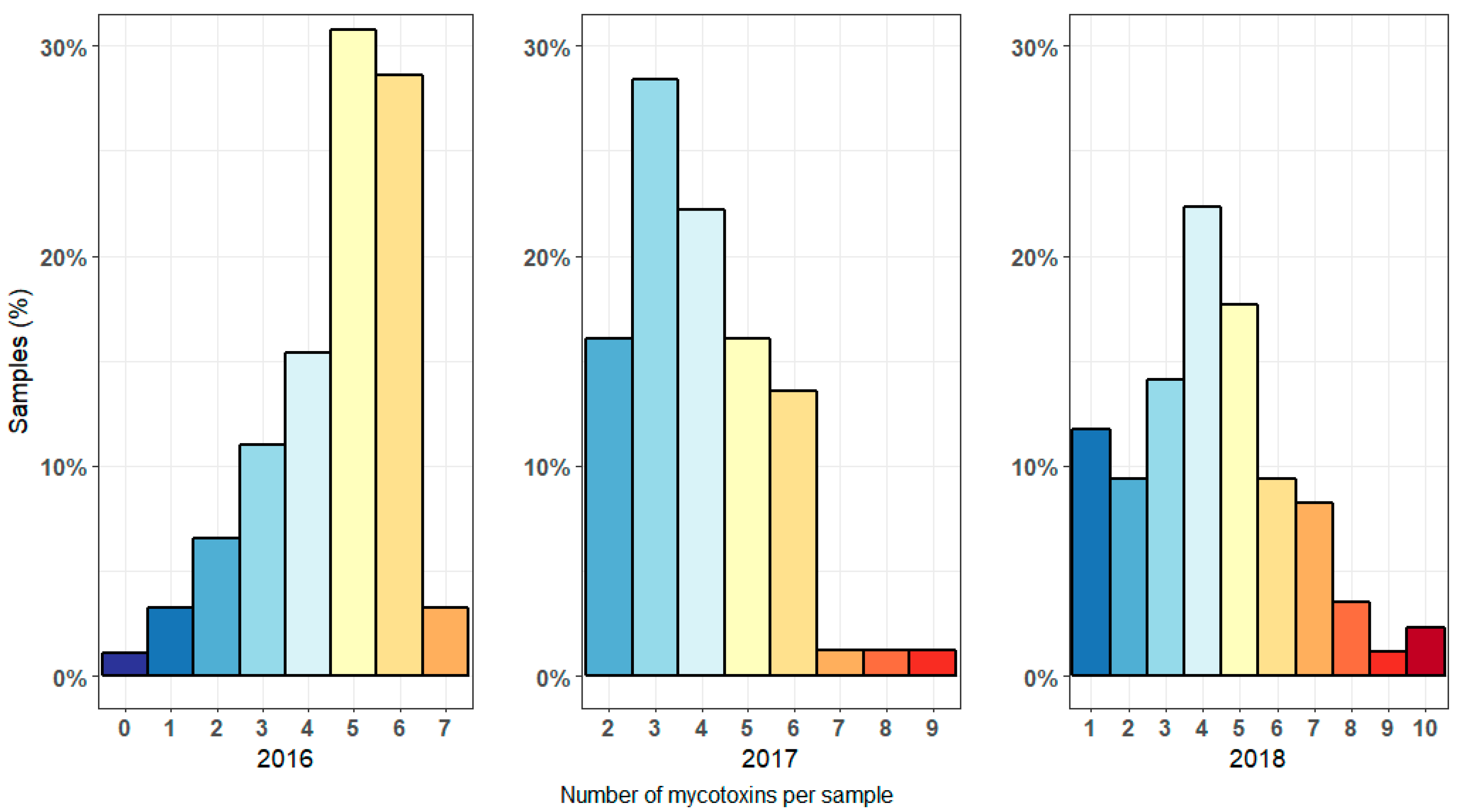
3.1. Mycotoxin Levels in Harvested Maize Samples in 2016–2018
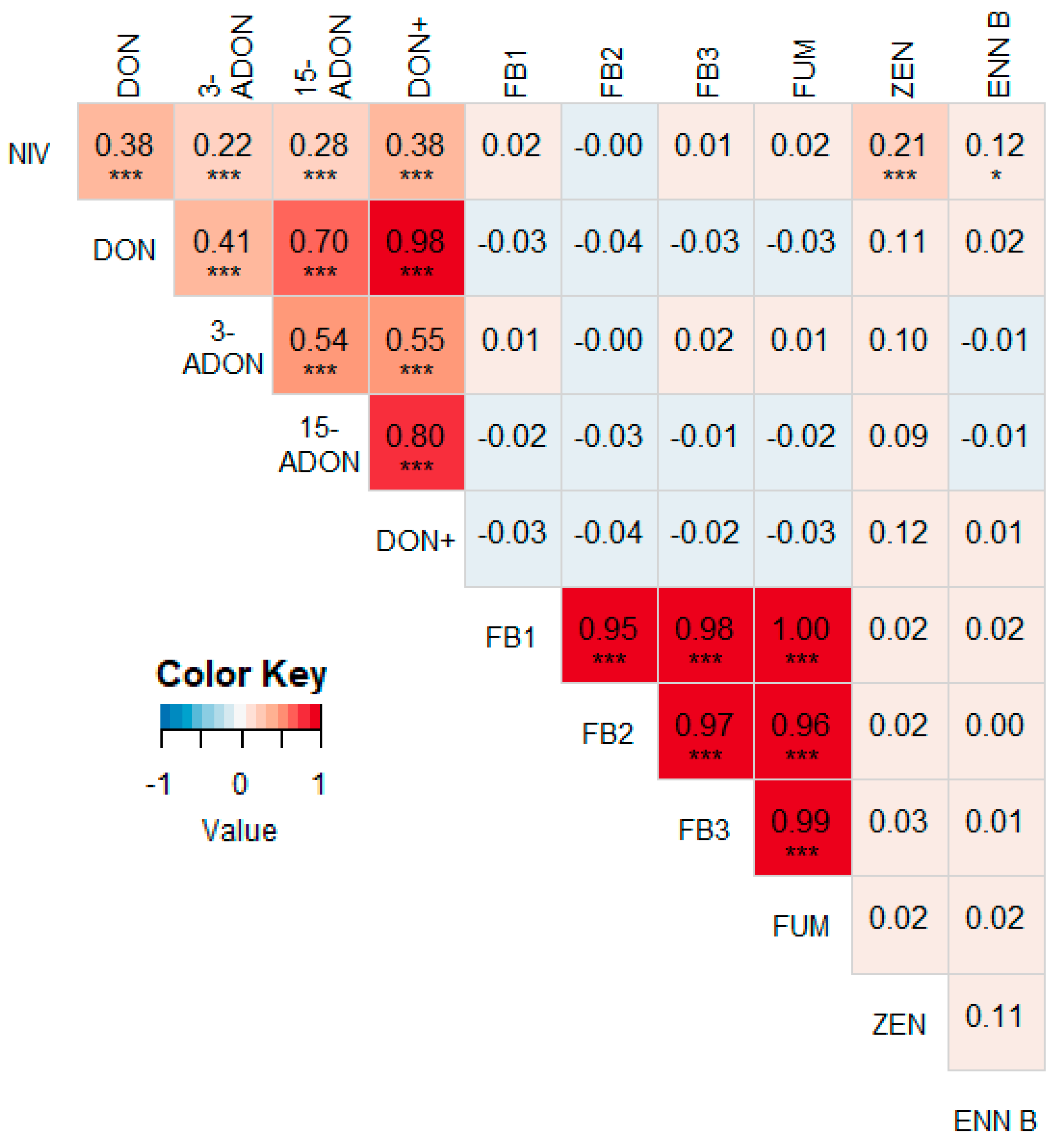
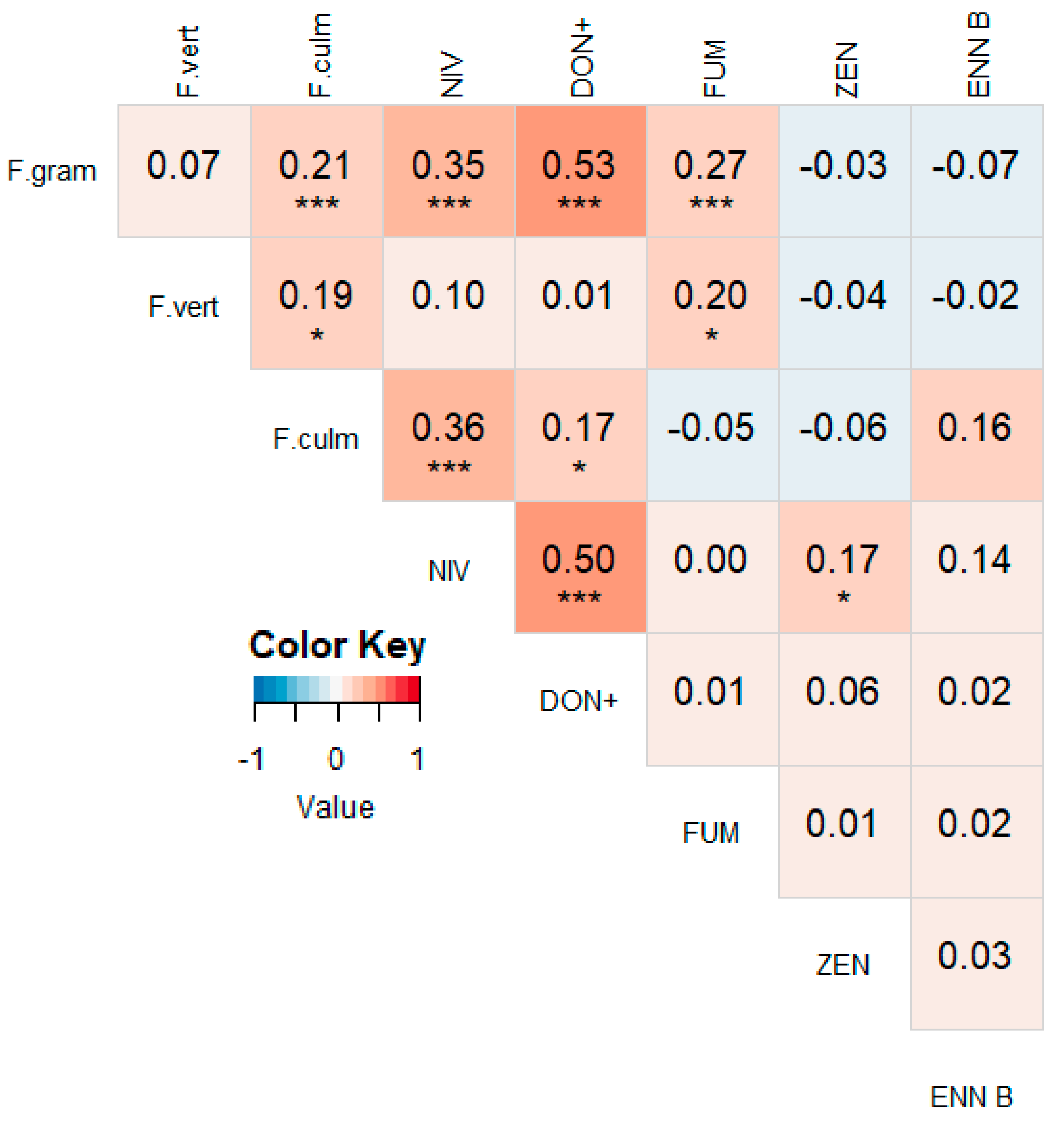
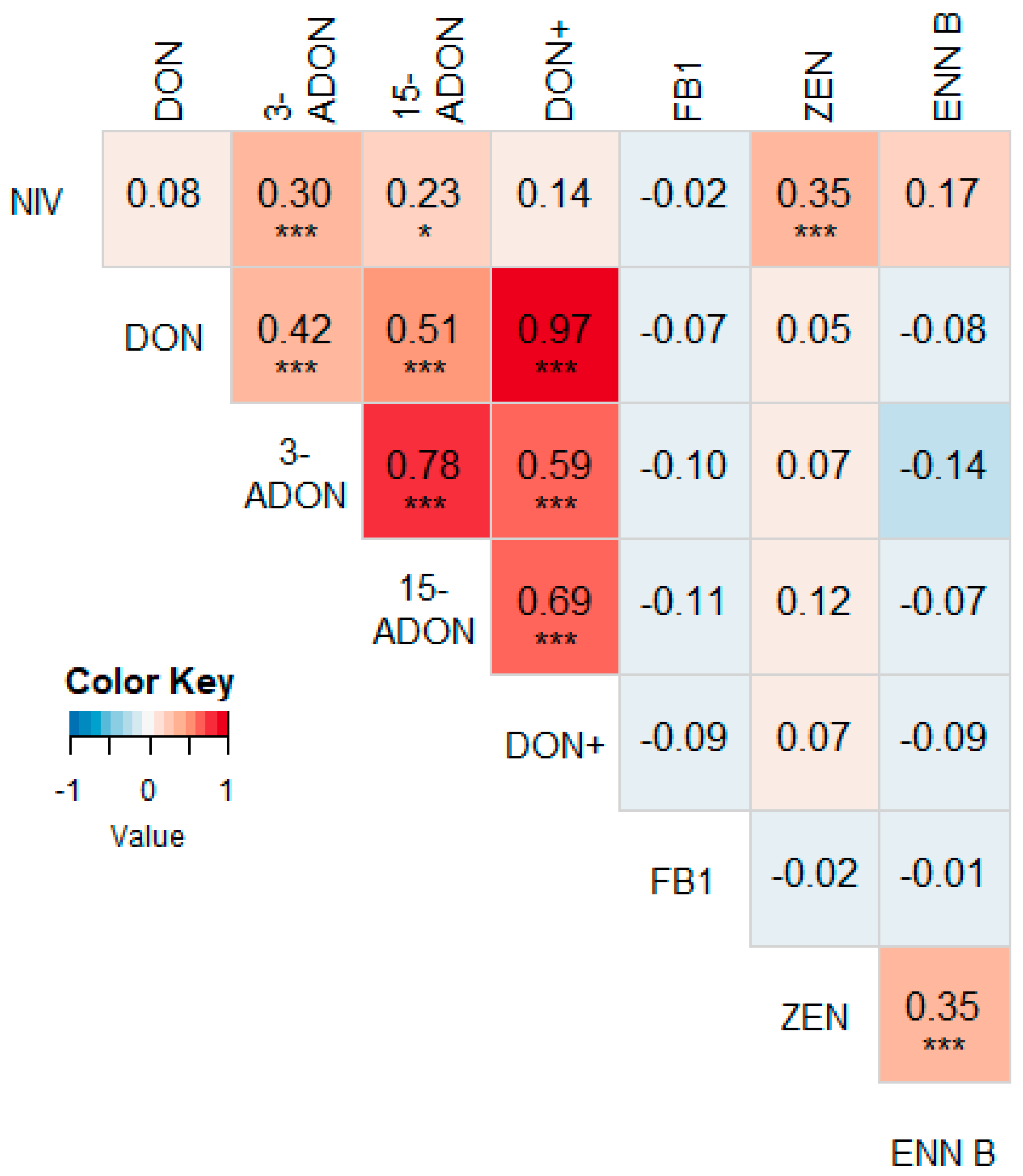
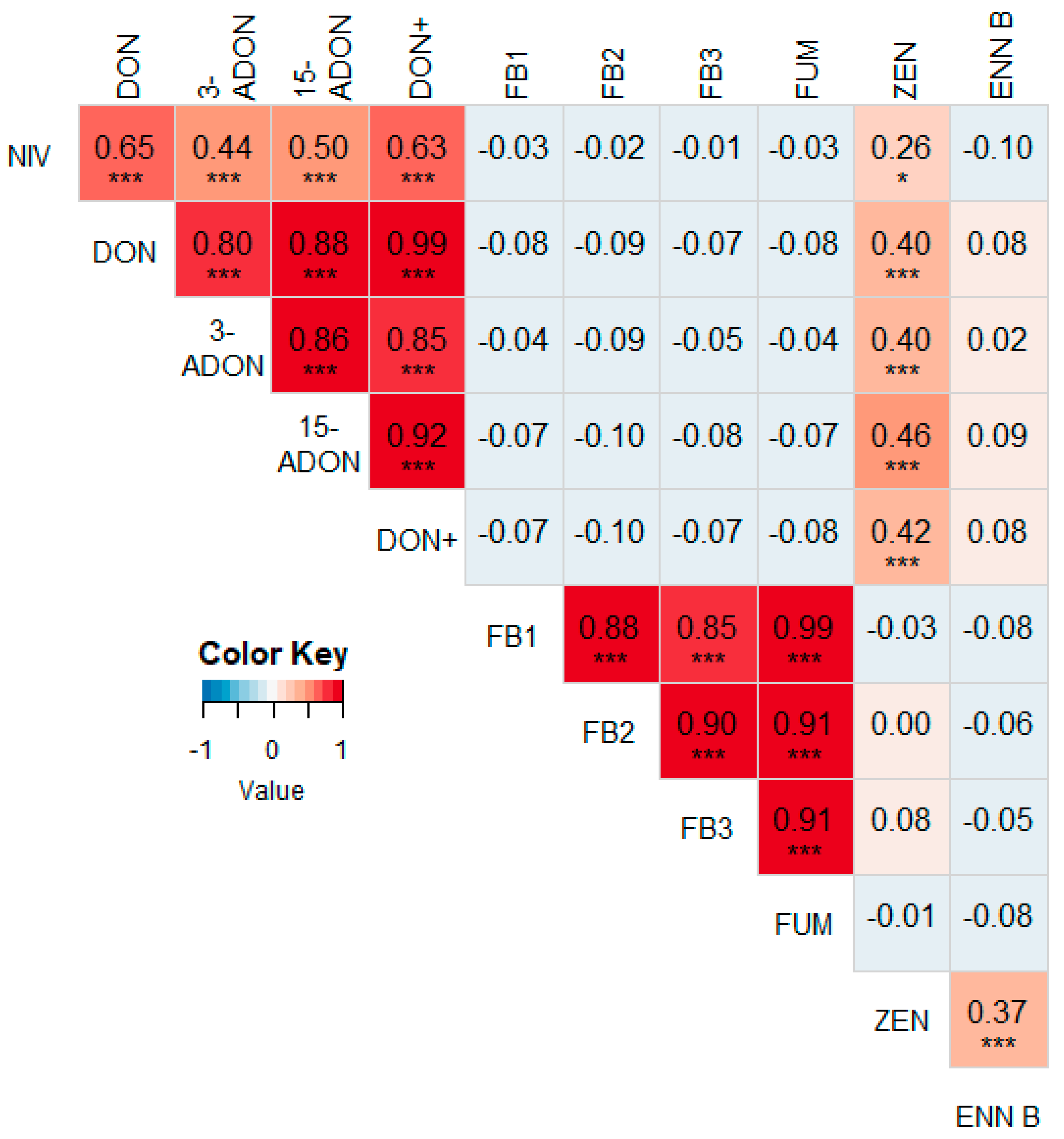
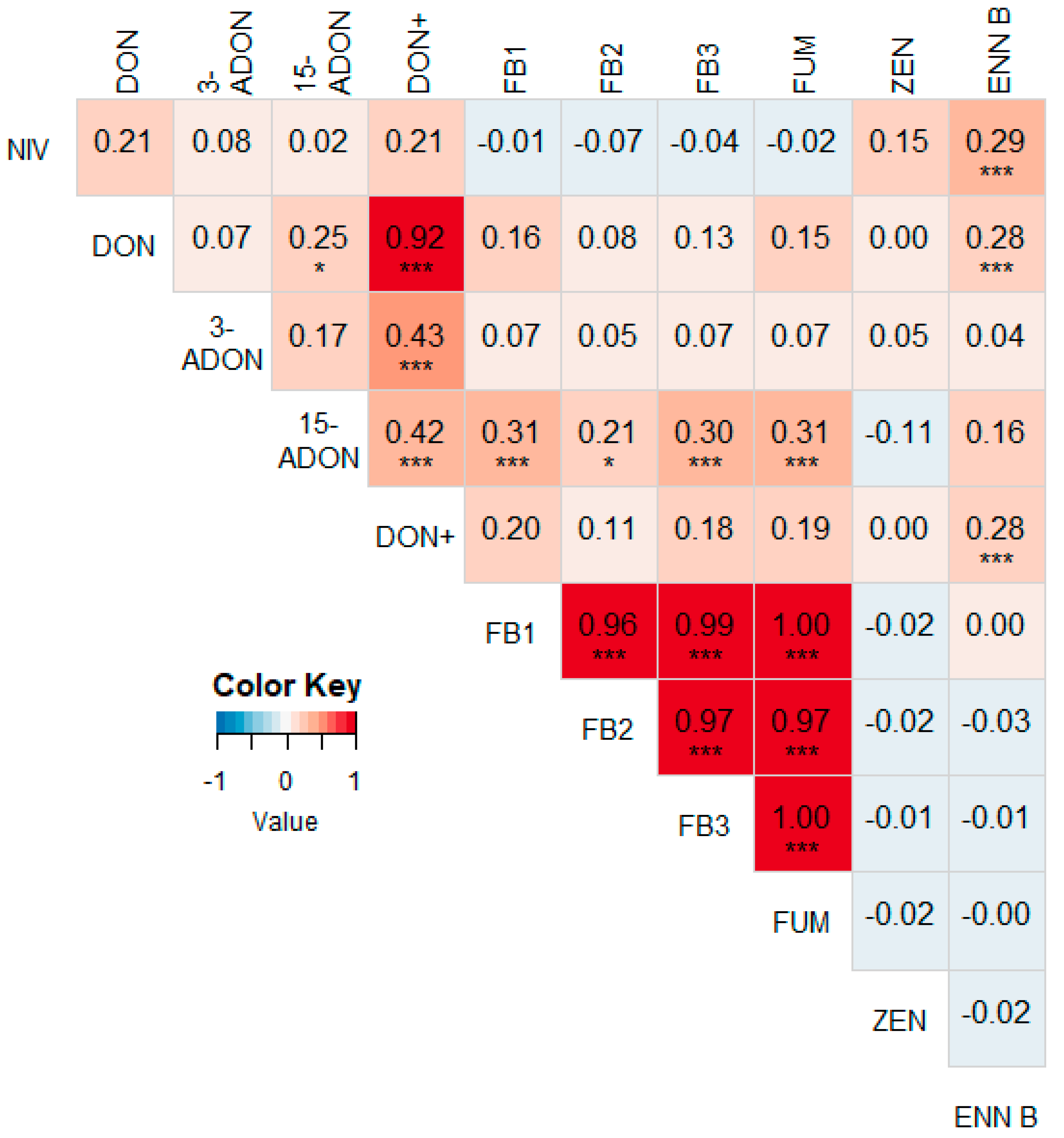
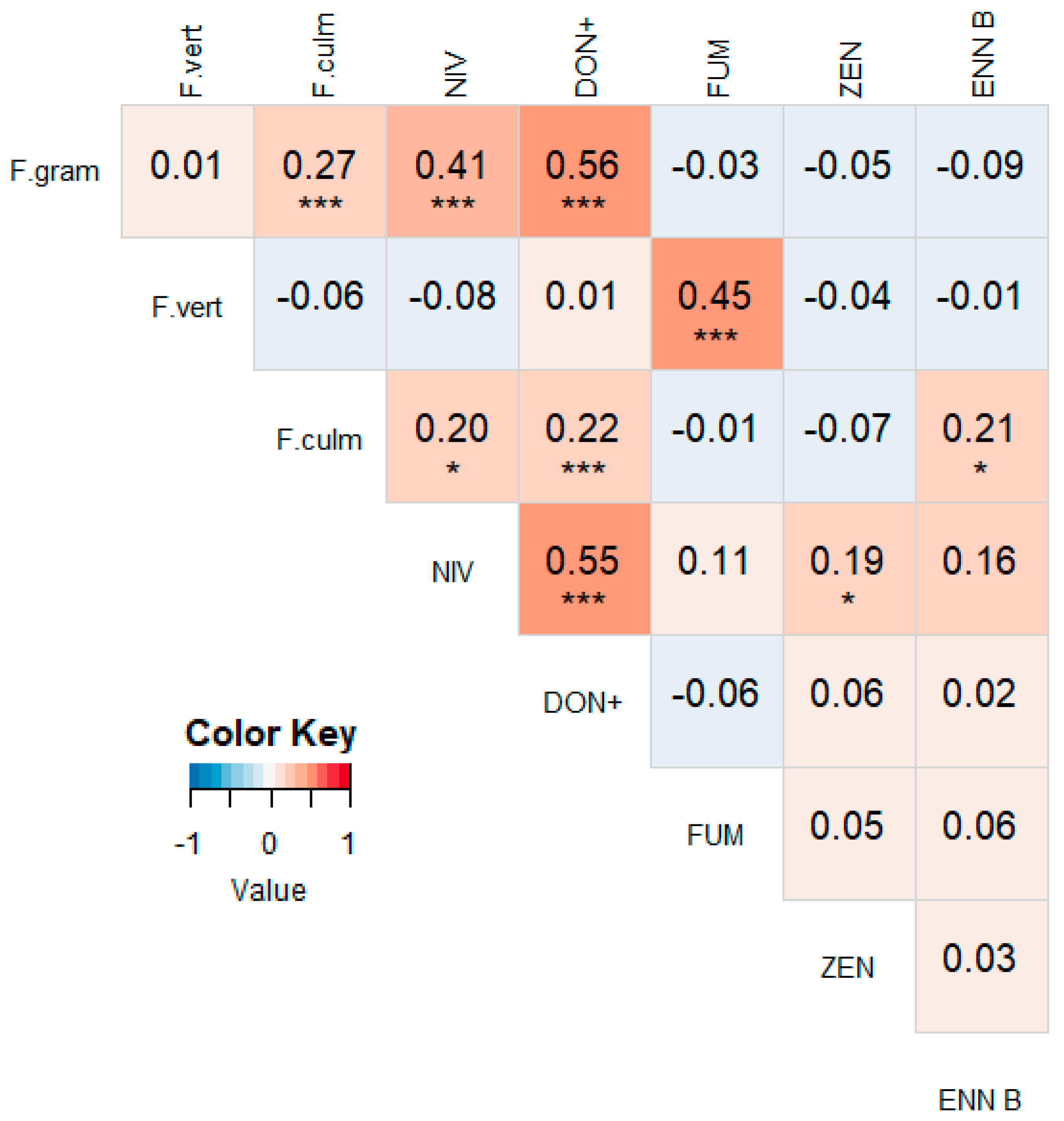
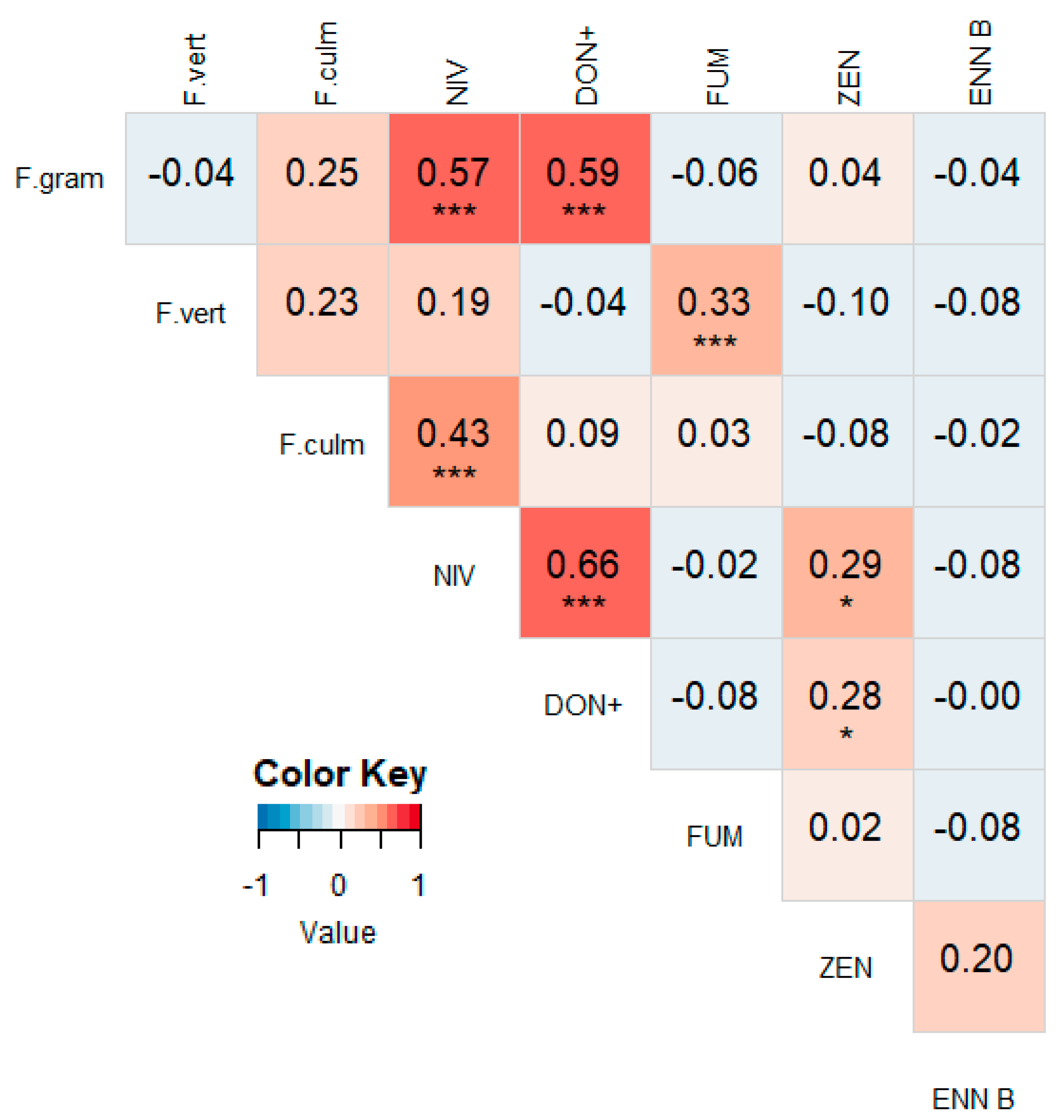
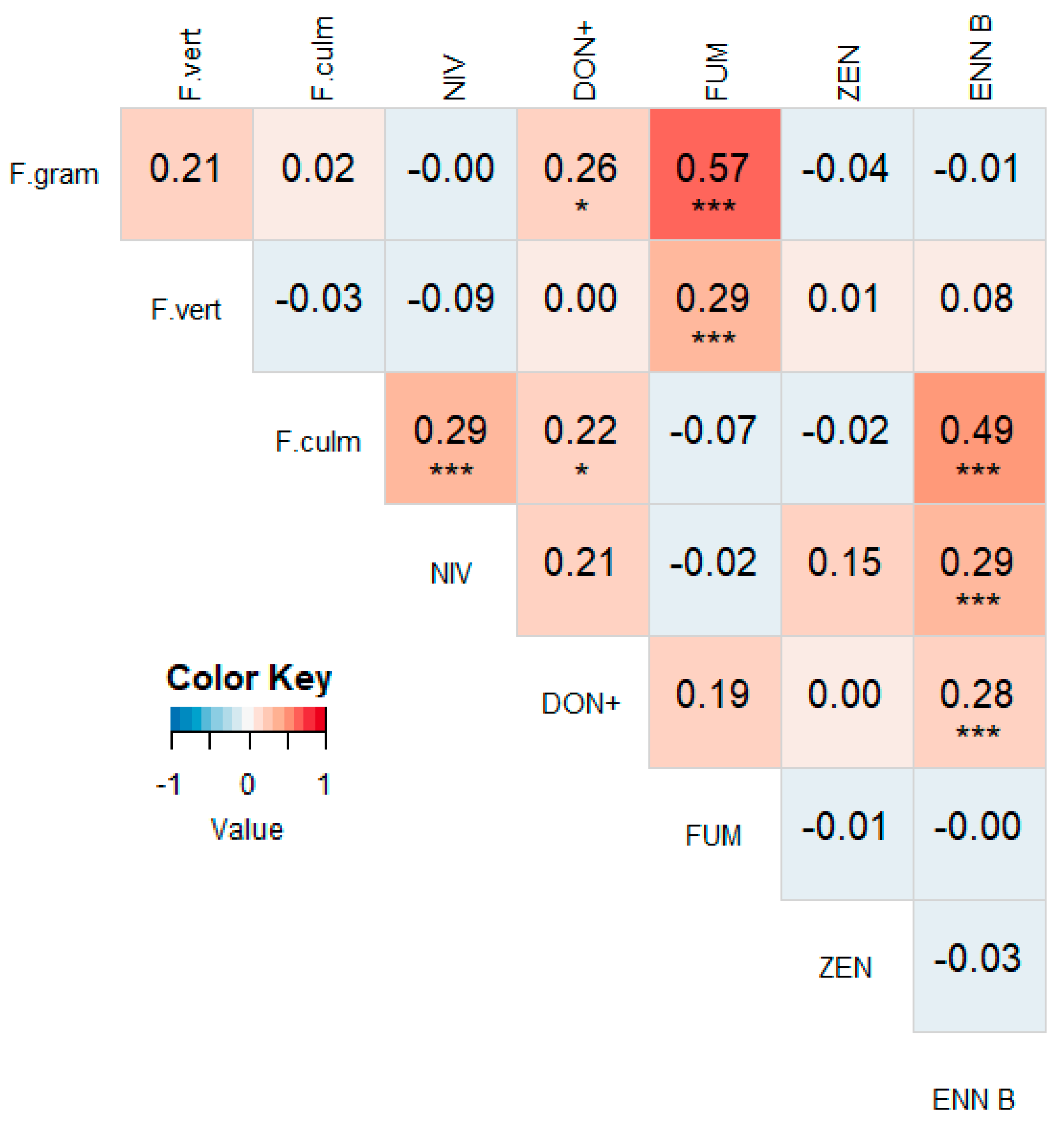
3.2. Correlations between Different Mycotoxins
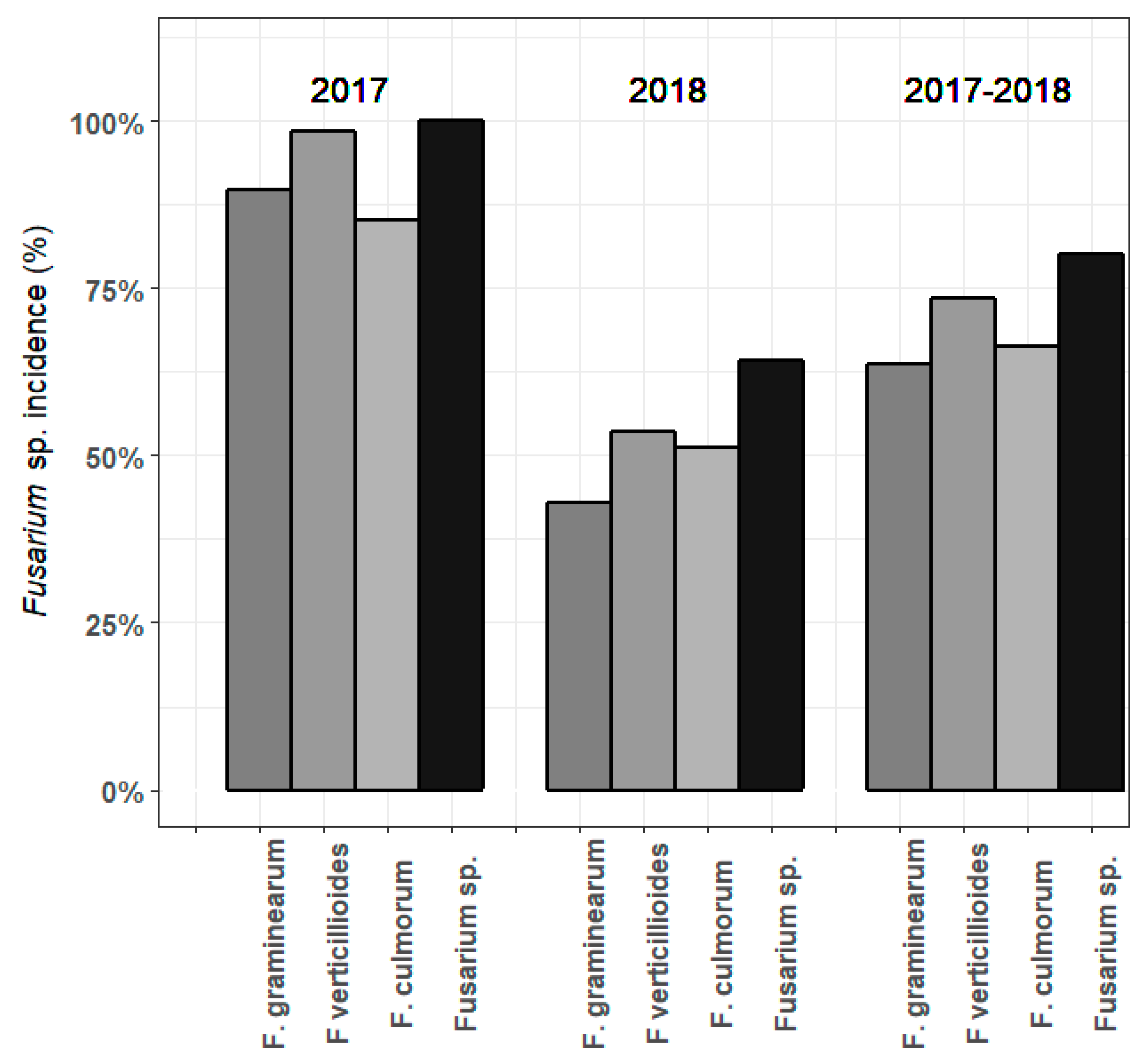
3.3. Fusarium spp. DNA in Maize Samples in 2017-2018
3.4. Correlations between Mycotoxin Concentrations and Fusarium spp. DNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A






References
- Storm, I.M.L.D.; Sørensen, J.L.; Rasmussen, R.R.; Nielsen, K.F.; Thrane, U. Mycotoxins in silage. Stewart Postharvest Rev. 2008, 4, 1–12. [Google Scholar]
- Tangni, E.K.; Pussemier, L.; van Hove, F. Mycotoxin contaminating maize and grass silages for dairy cattle feeding: Current state and challenges. J. Anim. Sci. Adv. 2013, 3, 492–511. [Google Scholar]
- Rüdelsheim, P.L.J.; Smets, G. Baseline Information on Agricultural Practices in the EU: Maize (Zea Mays L.); Perseus BVBA: Sint-Martens-Latem, Belgium, 2011. [Google Scholar]
- Department of Agriculture and Fisheries Voorlopige Arealen Landbouwteelten uit de Verzamelaanvraag 2019. Available online: https://lv.vlaanderen.be/nl/nieuws/voorlopige-arealen-landbouwteelten-uit-de-verzamelaanvraag-2019 (accessed on 20 September 2019).
- Kiessling, K.H.; Pettersson, H.; Sandholm, K.; Olsen, M. Metabolism of aflatoxin, ochratoxin, zearalenone, and three trichothecenes by intact rumen fluid, rumen protozoa, and rumen bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar]
- Fink-Gremmels, J. Mycotoxins in cattle feeds and carry-over to dairy milk: A review. Food Addit. Contam. 2008, 25, 172–180. [Google Scholar] [CrossRef]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on mycotoxin issues in ruminants: Occurrence in forages, effects of mycotoxin ingestion on health status and animal performance and practical strategies to counteract their negative effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed]
- Caloni, F.; Spotti, M.; Auerbach, H.; Op Den Camp, H.; Fink Gremmels, J.; Pompa, G. In vitro metabolism of fumonisin B1 by ruminal microflora. Vet. Res. Commun. 2000, 24, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Seeling, K.; Dänicke, S.; Ueberschär, H.; Lebzien, P.; Flachowsky, G. On the effects of Fusarium toxin-contaminated wheat and the feed intake level on the metabolism and carry over of zearalenone in dairy cows. Food Addit. Contam. 2005, 22, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, I. A review on the effects of mycotoxins in dairy ruminants. Anim. Prod. Sci. 2014, 54, 1155–1165. [Google Scholar] [CrossRef]
- Mostrom, M.S.; Jacobsen, B.J. Ruminant mycotoxicosis. Vet. Clin. N. Am. Food Anim. Pract. 2011, 27, 315–344. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef]
- Wu, F. Measuring the economic impacts of Fusarium toxins in animal feeds. Anim. Feed Sci. Technol. 2007, 137, 363–374. [Google Scholar] [CrossRef]
- Bhat, R.; Rai, R.V.; Karim, A.A. Mycotoxins in food and feed: Present status and future concerns. Compr. Rev. Food Sci. Food Saf. 2010, 9, 57–81. [Google Scholar] [CrossRef]
- Sweeney, M.J.; Dobson, A.D.W. Mycotoxin production by Aspergillus, Fusarium and Penicillium species. Int. J. Food Microbiol. 1998, 43, 141–158. [Google Scholar] [CrossRef]
- Dorn, B.; Forrer, H.R.; Schürch, S.; Vogelgsang, S. Fusarium species complex on maize in Switzerland: Occurrence, prevalence, impact and mycotoxins in commercial hybrids under natural infection. Eur. J. Plant Pathol. 2009, 125, 51–61. [Google Scholar] [CrossRef]
- Koehler, B. Corn Ear Rots in Illinois; Urbana, Ill., Ed.; Agricultural Experiment Station, University of Illinois: Champaign, IL, USA, 1959. [Google Scholar]
- Logrieco, A.; Mulé, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol. 2002, 108, 579–609. [Google Scholar] [CrossRef]
- Bottalico, A. Fusarium diseases of cereals: Species complex and related mycotoxin profiles, in Europe. J. Plant Pathol. 1998, 80, 85–103. [Google Scholar]
- Oldenburg, E.; Höppner, F.; Ellner, F.; Weinert, J. Fusarium diseases of maize associated with mycotoxin contamination of agricultural products intended to be used for food and feed. Mycotoxin Res. 2017, 33, 167–182. [Google Scholar] [CrossRef]
- Scauflaire, J.; Mahieu, O.; Louvieaux, J.; Foucart, G.; Renard, F.; Munaut, F. Biodiversity of Fusarium species in ears and stalks of maize plants in Belgium. Eur. J. Plant Pathol. 2011, 131, 59–66. [Google Scholar] [CrossRef]
- Goertz, A.; Zuehlke, S.; Spiteller, M.; Steiner, U.; Dehne, H.W.; Waalwijk, C.; de Vries, I.; Oerke, E.C. Fusarium species and mycotoxin profiles on commercial maize hybrids in Germany. Eur. J. Plant Pathol. 2010, 128, 101–111. [Google Scholar] [CrossRef]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar] [CrossRef]
- Eckard, S.; Wettstein, F.E.; Forrer, H.R.; Vogelgsang, S. Incidence of Fusarium species and mycotoxins in silage maize. Toxins 2011, 3, 949–967. [Google Scholar] [CrossRef] [PubMed]
- Picot, A.; Hourcade-Marcolla, D.; Barreau, C.; Pinson-Gadais, L.; Caron, D.; Richard-Forget, F.; Lannou, C. Interactions between Fusarium verticillioides and Fusarium graminearum in maize ears and consequences for fungal development and mycotoxin accumulation. Plant Pathol. 2012, 61, 140–151. [Google Scholar] [CrossRef]
- Velluti, A.; Marín, S.; Gonzalez, R.; Ramos, A.J.; Sanchis, V. Fumonisin B1, zearalenone and deoxynivalenol production by Fusarium moniliforme, F. proliferatum and F. graminearum in mixed cultures on irradiated maize kernels. J. Sci. Food Agric. 2001, 81, 88–94. [Google Scholar] [CrossRef]
- Reid, L.M.; Nicol, R.W.; Ouellet, T.; Savard, M.; Miller, J.D.; Young, J.C.; Stewart, D.W.; Schaafsma, A.W. Interaction of Fusarium graminearum and F. moniliforme in maize ears: Disease progress, fungal biomass, and mycotoxin accumulation. Phytopathology 1999, 89, 1028–1037. [Google Scholar] [CrossRef]
- Warren, H.L.; Kommedahl, T. Prevalence and pathogenicity to corn of Fusarium species from corn roots, rhizosphere, residues, and soil. Phytopathology 1973, 63, 1288. [Google Scholar] [CrossRef]
- Giorni, P.; Bertuzzi, T.; Battilani, P. Impact of fungi co-occurrence on mycotoxin contamination in maize during the growing season. Front. Microbiol. 2019, 10, 1265. [Google Scholar] [CrossRef]
- Munkvold, G.P. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 2003, 109, 705–713. [Google Scholar] [CrossRef]
- Oldenburg, E.; Ellner, F. Distribution of disease symptoms and mycotoxins in maize ears infected by Fusarium culmorum and Fusarium graminearum. Mycotoxin Res. 2015, 31, 117–126. [Google Scholar] [CrossRef]
- Foley, D.C. Systemic infection of corn by Fusarium moniliforme. Phytopathology 1962, 52, 870–872. [Google Scholar]
- Gilbertson, R.L.; Brown, W.M.J.; Ruppel, E.G.; Capinera, J.L. Association of Corn Stalk Rot Fusarium spp. and Western Corn Rootworm Beetles in Colorado. Phytopathology 1986, 76, 1309–1314. [Google Scholar] [CrossRef]
- Munkvold, G.P.; McGee, D.C.; Carlton, W.M. Importance of different pathways for maize kernel infection by Fusarium moniliforme. Phytopathology 1997, 87, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Thrane, U.; Samson, R.A.; Pitt, J.I. Important mycotoxins and the fungi which produce them. In Advances in Food Mycology. Advances in Experimental Medicine and Biology; Hocking, A.D., Pitt, J.I., Samson, R.A., Thrane, U., Eds.; Springer: Boston, MA, USA, 2006; Volume 571, pp. 3–31. [Google Scholar]
- Miller, J.D. Fungi and mycotoxins in grain: Implication for stored product research. J. Stored Prod. Res. 1995, 31, 1–16. [Google Scholar] [CrossRef]
- Binder, E.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Van Asselt, E.D.; Azambuja, W.; Moretti, A.; Kastelein, P.; de Rijk, T.C.; Stratakou, I.; van der Fels-Klerx, H.J. A Dutch field survey on fungal infection and mycotoxin concentrations in maize. Food Addit. Contam. 2012, 29, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
- Kosicki, R.; Błajet-Kosicka, A.; Grajewski, J.; Twaruzek, M. Multiannual mycotoxin survey in feed materials and feedingstuffs. Anim. Feed Sci. Technol. 2016, 215, 165–180. [Google Scholar] [CrossRef]
- Gruber-Dorninger, C.; Jenkins, T.; Schatzmayr, G. Global mycotoxin occurrence in feed: A ten-year survey. Toxins 2019, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.D. Mycotoxins in small grains and maize: Old problems, new challenges. Food Addit. Contam. Part A 2008, 25, 219–230. [Google Scholar] [CrossRef]
- Paterson, R.R.M.; Lima, N. How will climate change affect mycotoxins in food? Food Res. Int. 2010, 43, 1902–1914. [Google Scholar] [CrossRef] [Green Version]
- Battilani, P.; Toscano, P.; Van Der Fels-Klerx, H.J.; Moretti, A.; Camardo Leggieri, M.; Brera, C.; Rortais, A.; Goumperis, T.; Robinson, T. Aflatoxin B1 contamination in maize in Europe increases due to climate change. Sci. Rep. 2016, 6, 24328. [Google Scholar] [CrossRef]
- Basler, R. Diversity of Fusarium species isolated from UK forage maize and the population structure of F. graminearum from maize and wheat. PeerJ 2016, 4, e2143. [Google Scholar] [CrossRef] [Green Version]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Moretti, A.; Pascale, M.; Logrieco, A.F. Mycotoxin risks under a climate change scenario in Europe. Trends Food Sci. Technol. 2019, 84, 38–40. [Google Scholar] [CrossRef]
- Garon, D.; Richard, E.; Sage, L.; Bouchart, V.; Pottier, D.; Lebailly, P. Mycoflora and multimycotoxin detection in corn silage: Experimental study. J. Agric. Food Chem. 2006, 54, 3479–3484. [Google Scholar] [CrossRef] [PubMed]
- Drejer Storm, I.M.L.; Rasmussen, R.R.; Rasmussen, P.H. Occurrence of pre- and post-harvest mycotoxins and other secondary metabolites in Danish maize silage. Toxins 2014, 6, 2256–2269. [Google Scholar] [CrossRef] [Green Version]
- Lepom, P. Occurrence of Fusarium species and their mycotoxins in maize: 1. Method of determining zearalenone in maize and maize silage by means of high performance liquid chomatography (HPLC) using fluorescence detection. Arch. Anim. Nutr. 1988, 38, 799–806. [Google Scholar]
- Boudra, H.; Morgavi, D.P. Reduction in Fusarium toxin levels in corn silage with low dry matter and storage time. J. Agric. Food Chem. 2008, 56, 4523–4528. [Google Scholar] [CrossRef]
- Richard, E.; Heutte, N.; Sage, L.; Pottier, D.; Bouchart, V.; Lebailly, P.; Garon, D. Toxigenic fungi and mycotoxins in mature corn silage. Food Chem. Toxicol. 2007, 45, 2420–2425. [Google Scholar] [CrossRef]
- Pelhate, J. Maize silage: Incidence of moulds during conservation. Folia Vet. Lat. 1977, 7, 1–16. [Google Scholar]
- Latorre, A.; Dagnac, T.; Lorenzo, B.F.; Llompart, M. Occurrence and stability of masked fumonisins in corn silage samples. Food Chem. 2015, 189, 38–44. [Google Scholar] [CrossRef]
- Zachariasova, M.; Dzuman, Z.; Veprikova, Z.; Hajkova, K.; Jiru, M.; Vaclavikova, M.; Zachariasova, A.; Pospichalova, M.; Florian, M.; Hajslova, J. Occurrence of multiple mycotoxins in European feedingstuffs, assessment of dietary intake by farm animals. Anim. Feed Sci. Technol. 2014, 193, 124–140. [Google Scholar] [CrossRef]
- Keller, L.A.M.; González Pereyra, M.L.; Keller, K.M.; Alonso, V.A.; Oliveira, A.A.; Almeida, T.X.; Barbosa, T.S.; Nunes, L.M.T.; Cavaglieri, L.R.; Rosa, C.A.R. Fungal and mycotoxins contamination in corn silage: Monitoring risk before and after fermentation. J. Stored Prod. Res. 2013, 52, 42–47. [Google Scholar] [CrossRef]
- Mansfield, M.A.; Jones, A.D.; Kuldau, G.A. Contamination of fresh and ensiled maize by multiple Penicillium mycotoxins. Phytopathology 2008, 98, 330–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheli, F.; Campagnoli, A.; Dell’Orto, V. Fungal populations and mycotoxins in silages: From occurrence to analysis. Anim. Feed Sci. Technol. 2013, 183, 1–16. [Google Scholar] [CrossRef]
- Schatzmayr, G.; Streit, E. Global occurrence of mycotoxins in the food and feed chain: Facts and figures. World Mycotoxin J. 2013, 6, 213–222. [Google Scholar] [CrossRef]
- Kamala, A.; Ortiz, J.; Kimanya, M.; Haesaert, G.; Donoso, S.; Tiisekwa, B.; De Meulenaer, B. Multiple mycotoxin co-occurrence in maize grown in three agro-ecological zones of Tanzania. Food Control 2015, 54, 208–215. [Google Scholar] [CrossRef]
- Kovalsky, P.; Kos, G.; Nährer, K.; Schwab, C.; Jenkins, T.; Schatzmayr, G.; Sulyok, M.; Krska, R. Co-occurrence of regulated, masked and emerging mycotoxins and secondary metabolites in finished feed and maize – An extensive survey. Toxins 2016, 8, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorn, B.; Forrer, H.R.; Jenny, E.; Wettstein, F.E.; Bucheli, T.D.; Vogelgsang, S. Fusarium species complex and mycotoxins in grain maize from maize hybrid trials and from grower’s fields. J. Appl. Microbiol. 2011, 111, 693–706. [Google Scholar] [CrossRef]
- Schollenberger, M.; Müller, H.M.; Ernst, K.; Sondermann, S.; Liebscher, M.; Schlecker, C.; Wischer, G.; Drochner, W.; Hartung, K.; Piepho, H.P. Occurrence and distribution of 13 trichothecene toxins in naturally contaminated maize plants in Germany. Toxins 2012, 4, 778–787. [Google Scholar] [CrossRef]
- European Commission. Commission regulation (EU) No 574/2011 of 16 June 2011 amending Annex I to directive 2002/32/EC of the European Parliament and of the Council as regards maximum levels for nitrite, melamine, Ambrosia spp. and carry-over of certain coccidiostats and histomonostats and consolidating Annexes I and II thereto. Off. J. Eur. Union 2011, 159, 7–24. [Google Scholar]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- European Commission. Commission recommendation of 27 March 2013 on the presence of T-2 and HT-2 in cereals and cereal products (2013/165/EU). Off. J. Eur. Union 2013, 91, 12–15. [Google Scholar]
- De Boevre, M.; Landschoot, S.; Audenaert, K.; Maene, P.; Di Mavungu, J.D.; Eeckhout, M.; Haesaert, G.; De Saeger, S. Occurrence and within field variability of Fusarium mycotoxins and their masked forms in maize crops in Belgium. World Mycotoxin J. 2014, 7, 91–102. [Google Scholar] [CrossRef]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [Green Version]
- Speijers, G.J.A.; Speijers, M.H.M. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Oswald, I. Mycotoxin co-contamination of food and feed: Meta-analysis of publications describing toxicological interactions. World Mycotoxin J. 2011, 4, 285–313. [Google Scholar] [CrossRef]
- Smith, T.K.; Seddon, I.R. Toxicological synergism between Fusarium mycotoxins in feeds. In Biotechnology in the Feed Industry; Lyons, T.P., Jacques, K.A., Eds.; Nottingham University Press: Loughborough, UK, 1998; pp. 257–269. [Google Scholar]
- Thuvander, A.; Wikman, C.; Gadhasson, I. In vitro exposure of human lymphocytes to trichothecenes: Individual variation in sensitivity and effects of combined exposure on lymphocyte function. Food Chem. Toxicol. 1999, 37, 639–648. [Google Scholar] [CrossRef]
- Fremy, J.M.; Alassane-Kpembi, I.; Oswald, I.P.; Cottrill, B.; Van Egmond, H.P. A review on combined effects of moniliformin and co-occurring Fusarium toxins in farm animals. World Mycotoxin J. 2019, 12, 281–291. [Google Scholar] [CrossRef]
- Vanheule, A.; Audenaert, K.; De Boevre, M.; Landschoot, S.; Bekaert, B.; Munaut, F.; Eeckhout, M.; Höfte, M.; De Saeger, S.; Haesaert, G. The compositional mosaic of Fusarium species and their mycotoxins in unprocessed cereals, food and feed products in Belgium. Int. J. Food Microbiol. 2014, 181, 28–36. [Google Scholar] [CrossRef]
- Landschoot, S.; Audenaert, K.; Waegeman, W.; Pycke, B.; Bekaert, B.; De Baets, B.; Haesaert, G. Connection between primary Fusarium inoculum on gramineous weeds, crop residues and soil samples and the final population on wheat ears in Flanders, Belgium. Crop Prot. 2011, 30, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Monbaliu, S.; Van Poucke, C.; Detavernier, C.T.L.; Dumoultn, F.; Van Velde, M.D.E.; Schoeters, E.; Van Dyck, S.; Averkieva, O.; Van Peteghem, C.; De Saeger, S. Occurrence of mycotoxins in feed as analyzed by a multi-mycotoxin LC-MS/MS method. J. Agric. Food Chem. 2010, 58, 66–71. [Google Scholar] [CrossRef]
- European Commission. Commission Directive 2003/100/EC of 31 October 2003 amending Annex I to Directive 2002/32/EC of the European Parliament an of the Council on undesirable substances in animal feed. Off. J. Eur. Union 2003, 285, 33–37. [Google Scholar]
- Quarta, A.; Mita, G.; Haidukowski, M.; Santino, A.; Mulé, G.; Visconti, A. Assessment of trichothecene chemotypes of Fusarium culmorum occurring in Europe. Food Addit. Contam. 2005, 22, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA. 1984, 81, 8014–8018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolaisen, M.; Suproniene, S.; Nielsen, L.K.; Lazzaro, I.; Spliid, N.H.; Justesen, A.F. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Methods 2009, 76, 234–240. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing 2017; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Pasquali, M.; Giraud, F.; Brochot, C.; Cocco, E.; Hoffmann, L.; Bohn, T. Genetic Fusarium chemotyping as a useful tool for predicting nivalenol contamination in winter wheat. Int. J. Food Microbiol. 2010, 137, 246–253. [Google Scholar] [CrossRef]
- Sampietro, D.A.; Díaz, C.G.; Gonzalez, V.; Vattuone, M.A.; Ploper, L.D.; Catalan, C.A.N.; Ward, T.J. Species diversity and toxigenic potential of Fusarium graminearum complex isolates from maize fields in northwest Argentina. Int. J. Food Microbiol. 2011, 145, 359–364. [Google Scholar] [CrossRef]
- Hellin, P.; Dedeurwaerder, G.; Duvivier, M.; Scauflaire, J.; Huybrechts, B.; Callebaut, A.; Munaut, F.; Legrève, A. Relationship between Fusarium spp. diversity and mycotoxin contents of mature grains in southern Belgium. Food Addit. Contam. 2016, 33, 1228–1240. [Google Scholar] [CrossRef] [Green Version]
- Ueno, Y. Trichothecenes: Chemical, Biological and Toxicological Aspects; Kodansha: Tokyo, Japan, 1983. [Google Scholar]
- Wu, W.; Flannery, B.M.; Sugita-Konishi, Y.; Watanabe, M.; Zhang, H.; Pestka, J.J. Comparison of murine anorectic responses to the 8-ketotrichothecenes 3-acetyldeoxynivalenol, 15-acetyldeoxynivalenol, fusarenon X and nivalenol. Food Chem. Toxicol. 2012, 50, 2056–2061. [Google Scholar] [CrossRef]
- Abbas, H.K.; Yoshizawa, T.; Shier, W.T. Cytotoxicity and phytotoxicity of trichothecene mycotoxins produced by Fusarium spp. Toxicon 2013, 74, 68–75. [Google Scholar] [CrossRef]
- Minervini, F.; Fornelli, F.; Flynn, K.M. Toxicity and apoptosis induced by the mycotoxins nivalenol, deoxynivalenol and fumonisin B1 in a human erythroleukemia cell line. Toxicol. Vitr. 2004, 18, 21–28. [Google Scholar] [CrossRef]
- Ferreira Lopes, S.; Vacher, G.; Ciarlo, E.; Savova-Bianchi, D.; Roger, T.; Niculita-Hirzel, H. Primary and Immortalized Human Respiratory Cells Display Different Patterns of Cytotoxicity and Cytokine Release upon Exposure to Deoxynivalenol, Nivalenol and Fusarenon-X. Toxins 2017, 9, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audenaert, K.; van Broeck, R.; Bekaert, B.; de Witte, F.; Heremans, B.; Messens, K.; Höfte, M.; Haesaert, G. Fusarium head blight (FHB) in Flanders: Population diversity, inter-species associations and DON contamination in commercial winter wheat varieties. Eur. J. Plant Pathol. 2009, 125, 445–458. [Google Scholar] [CrossRef]
- Waalwijk, C.; Kastelein, P.; De Vries, I.; Kerényi, Z.; Van Der Lee, T.; Hesselink, T.; Köhl, J.; Kema, G. Major changes in Fusarium spp. in wheat in the Netherlands. Eur. J. Plant Pathol. 2003, 109, 743–754. [Google Scholar] [CrossRef]
- Borutova, R.; Aragon, Y.A.; Nährer, K.; Berthiller, F. Co-occurrence and statistical correlations between mycotoxins in feedstuffs collected in the Asia-Oceania in 2010. Anim. Feed Sci. Technol. 2012, 178, 190–197. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [Green Version]
- Zorzete, P.; Castro, R.S.; Pozzi, C.R.; Israel, A.L.M.; Fonseca, H.; Yanaguibashi, G.; Correa, B. Relative populations and toxin production by Aspergillus flavus and Fusarium verticillioides in artificially inoculated corn at various stages of development under field conditions. J. Sci. Food Agric. 2008, 88, 48–55. [Google Scholar] [CrossRef]
- Cooney, J.M.; Lauren, D.R.; Di Menna, M.E. Impact of competitive fungi on trichothecene production by Fusarium graminearum. J. Agric. Food Chem. 2001, 49, 522–526. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Venturi, V. Synergisms between microbial pathogens in plant disease complexes: A growing trend. Front. Plant Sci. 2015, 6, 385. [Google Scholar] [CrossRef] [Green Version]
- Marín, S.; Sanchis, V.; Ramos, A.J.; Vinas, I.; Magan, N. Environmental factors, in vitro interactions, and niche overlap between Fusarium moniliforme, F. proliferatum, and F. graminearum, Aspergillus and Penicillium species from maize grain. Mycol. Res. 1998, 102, 831–837. [Google Scholar] [CrossRef]
- Etcheverry, M.; Magnoli, C.; Dalcero, A.; Chulze, S.; Lecumberry, S. Aflatoxin B1, zearalenone and deoxynivalenol production by Aspergillus parasiticus and Fusarium graminearum in interactive cultures on irradiated corn kernels. Mycopathologia 1998, 142, 37–42. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Plant stress and mycotoxin accumulation in maize. Agrochimica 2014, 58, 116–127. [Google Scholar]
- Brennan, J.M.; Fagan, B.; Van Maanen, A.; Cooke, B.M.; Doohan, F.M. Studies on in vitro growth and pathogenicity of European Fusarium fungi. Eur. J. Plant Pathol. 2003, 109, 577–587. [Google Scholar] [CrossRef]
- KMI. Juli 2019: Absolute Warmterecord Gebroken. Available online: https://www.meteo.be/nl/over-het-kmi/contact/gegevens-kmi (accessed on 10 October 2019).
- Bullerman, L.B.; Schroeder, L.L.; Park, K.Y. Formation and control of mycotoxins in food. J. Food Prot. 1984, 47, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Rodriguez, A.; Magan, N. Effect of climate change on Aspergillus flavus and aflatoxin B 1 production. Front. Microbiol. 2014, 5, 348. [Google Scholar] [CrossRef]
- Miller, J.D. Factors that affect the occurrence of fumonisin. Environ. Health Perspect. 2001, 109, 321–324. [Google Scholar]
- Glenn, A.E. Mycotoxigenic Fusarium species in animal feed. Anim. Feed Sci. Technol. 2007, 137, 213–240. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Positive Samples (%) | Mean Concentration a (µg/kg) | Median Concentration (µg/kg) | Max. Concentration (µg/kg) | Samples Exceeding EU Recommendation (%)b | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | 2016–2018 | 2016 | 2017 | 2018 | 2016–2018 | 2016 | 2017 | 2018 | 2016–2018 | 2016 | 2017 | 2018 | 2016-2018 | 2016 | 2017 | 2018 | 2016–2018 | |
| n samples | 91 | 81 | 85 | 257 | 91 | 81 | 85 | 257 | 91 | 81 | 85 | 257 | 91 | 81 | 95 | 257 | 91 | 81 | 95 | 257 |
| NIV | 98.9 | 100 | 98.8 | 99.2 | 650.7 | 719.0 | 881.9 | 748.7 | 527.5 | 460.6 | 782.1 | 587.1 | 2368.2 | 6776.3 | 2409.5 | 6776.3 | ||||
| DON | 92.3 | 100 | 64.7 | 85.6 | 449.0 | 557.5 | 186.5 | 396.4 | 263.1 | 337.4 | 121.3 | 215.3 | 2777.4 | 5322.4 | 2110.5 | 5322.4 | 2.2 | 3.7 | 1.0 | 2.3 |
| 3-ADON | 78.0 | 29.6 | 15.3 | 42.0 | 53.5 | 36.3 | 23.3 | 38.1 | 43.2 | 0 | 0 | 0 | 297.0 | 380.3 | 1046.8 | 1046.8 | ||||
| 15-ADON | 64.8 | 51.3 | 12.9 | 43.4 | 95.0 | 81.2 | 15.2 | 64.5 | 71.3 | 17.7 | 0 | 0 | 819.3 | 769.2 | 248.6 | 819.3 | ||||
| DON+c | 95.6 | 100 | 64.7 | 86.8 | 597.5 | 675.0 | 225.0 | 498.7 | 376.9 | 406.7 | 130.3 | 261.5 | 3050.1 | 6471.9 | 2110.5 | 6471.9 | ||||
| ZEN | 64.8 | 40.7 | 42.4 | 49.8 | 100.5 | 158.5 | 175.5 | 159.7 | 71.2 | 0 | 0 | 0 | 1085.6 | 1411.9 | 2791.6 | 2791.6 | 1.1 | 8.6 | 12.6 | 7.8 |
| ENN B | 42.9 | 18.5 | 45.9 | 36.2 | 133.1 | 77.7 | 180.3 | 149.5 | 56.2 | 27.5 | 70.7 | 46.2 | 1375.1 | 1041.9 | 1984.9 | 1984.9 | ||||
| AOH | n.d. | 3.7 | 9.4 | 4.3 | n.d. | 1.4 | 6.5 | 2.6 | n.d. | 0 | 0 | 0 | n.d. | 49.1 | 208.6 | 208.6 | ||||
| AME | 2.2 | 3.7 | 10.6 | 5.4 | 0.8 | 11.5 | 19.8 | 10.5 | 0 | 0 | 0 | 0 | 49.1 | 370.6 | 452.8 | 452.8 | ||||
| FB1d | 2.5 | 19.8 | 61.2 | 28.6 | 1.5 | 61.1 | 247.4 | 106.5 | 0 | 0 | 54.0 | 0 | 70.2 | 1362.9 | 4414.9 | 4414.9 | 0 | 0 | 0 | 0 |
| FB2d | n.d. | 4.9 | 24.7 | 10.2 | n.d. | 9.0 | 61.6 | 24.4 | n.d. | 0 | 0 | 0 | n.d. | 412.6 | 1427.4 | 1427.4 | 0 | 0 | 0 | 0 |
| FB3d | n.d. | 7.4 | 18.8 | 9.0 | n.d. | 3.4 | 18.0 | 7.4 | n.d. | 0 | 0 | 0 | n.d. | 90.5 | 451.2 | 451.2 | ||||
| FUMc | 2.5 | 19.8 | 61.2 | 28.6 | 1.3 | 73.6 | 327.0 | 131.8 | 0 | 0 | 58.7 | 0 | 70.2 | 1782.8 | 6293.5 | 6293.5 | ||||
| DAS | 11.0 | 8.6 | 5.9 | 8.6 | 0.3 | 0.3 | 0.4 | 0.4 | 0 | 0 | 0 | 0 | 6.1 | 10.3 | 14.9 | 14.9 | ||||
| FX | n.d. | 7.4 | 2.4 | 3.1 | n.d. | 14.2 | 2.7 | 5.4 | n.d. | 0 | 0 | 0 | n.d. | 223.6 | 161.6 | 223.6 | ||||
| T2 | 1.1 | n.d. | 8.2 | 3.1 | 0.2 | n.d. | 6.2 | 2.1 | 0 | n.d. | 0 | 0 | 16.8 | n.d. | 121.6 | 121.6 | 0 | 0 | 0 | 0 |
| STERIG | 1.1 | n.d. | 1.2 | 0.8 | 0.2 | n.d. | 2.6 | 0.9 | 0 | n.d. | 0 | 0 | 15.1 | n.d. | 73.3 | 204.8 | ||||
| ROQ-Cd | n.d. | 2.5 | 2.9 | 1.7 | n.d. | 0.6 | 0.6 | 0.4 | n.d. | 0 | 0 | 0 | n.d. | 30.4 | 24.6 | 30.4 | ||||
| TOTALc | 98.9 | 100 | 100 | 100 | 1485.1 | 1729.9 | 1877.4 | 1692.0 | 1309.6 | 1088.2 | 1596.1 | 1309.6 | 4153.4 | 13747.6 | 8309.0 | 13747.6 | ||||
| Year | Growing Season | Rainfall (mm) | Relative Humidity (%) | Average Temperature (°C) | Total Daily Radiation (W/m²) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | Range (Min. - Max.) | Mean | Range (Min. - Max.) | Mean | Range (Min. - Max.) | Mean | Range (Min. - Max.) | ||
| 2016 | 20.04–26.10 (189 days) | 423 | 283–610 | 80.5 | 72.8–86.3 | 15.7 | 15.1–16.7 | 3839 | 3697–4024 |
| 2017 | 10.04–28.10 (201 days) | 344 | 186–541 | 78.6 | 70.3–85.3 | 15.7 | 15.1–16.2 | 3389 | 2600–3902 |
| 2018 | 19.04–14.10 (178 days) | 241 | 155–335 | 74.2 | 69.6–84.7 | 17.5 | 16.0–18.3 | 4294 | 4109–4710 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vandicke, J.; De Visschere, K.; Croubels, S.; De Saeger, S.; Audenaert, K.; Haesaert, G. Mycotoxins in Flanders’ Fields: Occurrence and Correlations with Fusarium Species in Whole-Plant Harvested Maize. Microorganisms 2019, 7, 571. https://doi.org/10.3390/microorganisms7110571
Vandicke J, De Visschere K, Croubels S, De Saeger S, Audenaert K, Haesaert G. Mycotoxins in Flanders’ Fields: Occurrence and Correlations with Fusarium Species in Whole-Plant Harvested Maize. Microorganisms. 2019; 7(11):571. https://doi.org/10.3390/microorganisms7110571
Chicago/Turabian StyleVandicke, Jonas, Katrien De Visschere, Siska Croubels, Sarah De Saeger, Kris Audenaert, and Geert Haesaert. 2019. "Mycotoxins in Flanders’ Fields: Occurrence and Correlations with Fusarium Species in Whole-Plant Harvested Maize" Microorganisms 7, no. 11: 571. https://doi.org/10.3390/microorganisms7110571