Root-Associated Fungal Communities in Two Populations of the Fully Mycoheterotrophic Plant Arachnitis uniflora Phil. (Corsiaceae) in Southern Chile

 , ,
, ,
Abstract
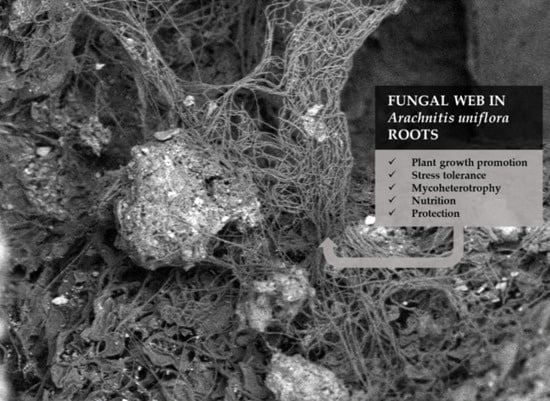
:
1. Introduction
2. Materials and Methods
2.1. Site Description and Sampling
2.2. Isolation and Characterization of Rhizospheric and Endophytic Fungi
2.3. Molecular Identification of Fungi
2.4. Screening of Plant-Growth-Promoting Traits
2.5. Data Analyses
3. Results
3.1. Sampling
3.2. Isolation of Fungi
3.3. Production of Plant-Growth-Promoter Metabolites and Organic Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Leake, J.R. The biology of myco-heterotrophic (‘saprophytic’) plants. New Phytol. 1994, 127, 171–216. [Google Scholar] [CrossRef]
- Tĕšitel, J.; Těšitelová, T.; Minasiewicz, J.; Selosse, M.-A. Mixotrophy in Land Plants: Why To Stay Green? Trends Plant Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Leake, J.R.; Cameron, D.D. Physiological ecology of mycoheterotrophy. New Phytol. 2010, 185, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-I.; Yang, C.-K.; Gebauer, G. The importance of associations with saprotrophic non-Rhizoctonia fungi among fully mycoheterotrophic orchids is currently under-estimated: Novel evidence from sub-tropical Asia. Ann. Bot. 2015, 116, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.H.; Taylor, D.L.; Watson, L.E. Mycorrhizal specificity in the fully mycoheterotrophic Hexalectris Raf.(Orchidaceae: Epidendroideae). Mol. Ecol. 2011, 20, 1303–1316. [Google Scholar] [CrossRef]
- Leake, J.; Johnson, D.; Donnelly, D.; Muckle, G.; Boddy, L.; Read, D. Networks of power and influence: The role of mycorrhizal mycelium in controlling plant communities and agroecosystem functioning. Can. J. Bot. 2004, 82, 1016–1045. [Google Scholar] [CrossRef]
- Selosse, M.-A.; Roy, M. Green plants that feed on fungi: Facts and questions about mixotrophy. Trends Plant Sci. 2009, 14, 64–70. [Google Scholar] [CrossRef]
- Dearnaley, J.; Perotto, S.; Selosse, M.A. Structure and development of orchid mycorrhizas. Mol. Mycorrhizal Symbiosis 2016, 63–86. [Google Scholar] [CrossRef]
- Herrera, H.; Valadares, R.; Contreras, D.; Bashan, Y.; Arriagada, C. Mycorrhizal compatibility and symbiotic seed germination of orchids from the Coastal Range and Andes in south central Chile. Mycorrhiza 2017, 27, 175–188. [Google Scholar] [CrossRef]
- Gilbert, L.; Johnson, D. Plant–Plant Communication Through Common Mycorrhizal Networks. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2017; Volume 82, pp. 83–97. [Google Scholar]
- Hynson, N.A.; Schiebold, J.M.-I.; Gebauer, G. Plant family identity distinguishes patterns of carbon and nitrogen stable isotope abundance and nitrogen concentration in mycoheterotrophic plants associated with ectomycorrhizal fungi. Ann. Bot. 2016, 118, 467–479. [Google Scholar] [CrossRef]
- Ogura-Tsujita, Y.; Gebauer, G.; Xu, H.; Fukasawa, Y.; Umata, H.; Tetsuka, K.; Kubota, M.; Schweiger, J.M.I.; Yamashita, S.; Maekawa, N. The giant mycoheterotrophic orchid Erythrorchis altissima is associated mainly with a divergent set of wood-decaying fungi. Mol. Ecol. 2018, 27, 1324–1337. [Google Scholar] [CrossRef] [PubMed]
- Braun, A.C.; Troeger, D.; Garcia, R.; Aguayo, M.; Barra, R.; Vogt, J. Assessing the impact of plantation forestry on plant biodiversity: A comparison of sites in Central Chile and Chilean Patagonia. Glob. Ecol. Conserv. 2017, 10, 159–172. [Google Scholar] [CrossRef]
- Garces-Voisenat, J.-P.; Mukherjee, Z. Paying for green energy: The case of the Chilean Patagonia. J. Policy Model. 2016, 38, 397–414. [Google Scholar] [CrossRef]
- Novoa, P.; Espejo, J.; Cisternas, M.; Rubio, M.; Dominguez, E. Guía de Campo de las Orquídeas Chilenas, 2nd ed.; Corporación Chilena De La Madera (Corma): Santiago, Chile, 2015; Available online: https://www.corma.cl/wp-content/uploads/2018/10/guia-de-campo-orquideas-2015-web.pdf (accessed on 9 September 2019).
- Domínguez, L.; Sérsic, A.; Melville, L.; Peterson, R.L. ‘Prepackaged symbioses’: Propagules on roots of the myco-heterotrophic plant Arachnitis uniflora. New Phytol. 2006, 169, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Bidartondo, M.I.; Redecker, D.; Hijri, I.; Wiemken, A.; Bruns, T.D.; Domínguez, L.; Sérsic, A.; Leake, J.R.; Read, D.J. Epiparasitic plants specialized on arbuscular mycorrhizal fungi. Nature 2002, 419, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, L.S.; Sérsic, A. The southernmost myco-heterotrophic plant, Arachnitis uniflora: Root morphology and anatomy. Mycologia 2004, 96, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Renny, M.; Acosta, M.C.; Cofré, N.; Domínguez, L.S.; Bidartondo, M.I.; Sérsic, A.N. Genetic diversity patterns of arbuscular mycorrhizal fungi associated with the mycoheterotroph Arachnitis uniflora Phil.(Corsiaceae). Ann. Bot. 2017, 119, 1279–1294. [Google Scholar] [CrossRef]
- Domínguez, L.S.; Melville, L.; Sersic, A.; Faccio, A.; Peterson, R.L. The mycoheterotroph Arachnitis uniflora has a unique association with arbuscular mycorrhizal fungi. Botany 2009, 87, 1198–1208. [Google Scholar] [CrossRef]
- Khan, A.R.; Ullah, I.; Waqas, M.; Shahzad, R.; Hong, S.-J.; Park, G.-S.; Jung, B.K.; Lee, I.-J.; Shin, J.-H. Plant growth-promoting potential of endophytic fungi isolated from Solanum nigrum leaves. World J. Microbiol. Biotechnol. 2015, 31, 1461–1466. [Google Scholar] [CrossRef]
- Grelet, G.-A.; Ba, R.; Goeke, D.F.; Houliston, G.J.; Taylor, A.F.; Durall, D.M. A plant growth-promoting symbiosis between Mycena galopus and Vaccinium corymbosum seedlings. Mycorrhiza 2017, 27, 831–839. [Google Scholar] [CrossRef]
- Soto, J.; Ortiz, J.; Herrera, H.; Fuentes, A.; Almonacid, L.; Charles, T.C.; Arriagada, C. Enhanced Arsenic Tolerance in Triticum aestivum Inoculated with Arsenic-Resistant and Plant Growth Promoter Microorganisms from a Heavy Metal-Polluted Soil. Microorganisms 2019, 7, 348. [Google Scholar] [CrossRef] [PubMed]
- Blain, N.P.; Helgason, B.L.; Germida, J.J. Endophytic root bacteria associated with the natural vegetation growing at the hydrocarbon-contaminated Bitumount Provincial Historic site. Can. J. Microbiol. 2017, 63, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Faria, D.C.; Dias, A.C.F.; Melo, I.S.; de Carvalho Costa, F.E. Endophytic bacteria isolated from orchid and their potential to promote plant growth. World J. Microbiol. Biotechnol. 2013, 29, 217–221. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Chen, J.; Hu, K.-X.; Hou, X.-Q.; Guo, S.-X. Endophytic fungi assemblages from 10 Dendrobium medicinal plants (Orchidaceae). World J. Microbiol. Biotechnol. 2011, 27, 1009–1016. [Google Scholar] [CrossRef]
- Samson, R.A.; Seifert, K.A.; Kuijpers, A.F.; Houbraken, J.; Frisvad, J.C. Phylogenetic analysis of Penicillium subgenus Penicillium using partial β-tubulin sequences. Stud. Mycol. 2004, 49, 175–200. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Ahemad, M.; Khan, M.S. Plant growth promoting activities of phosphate-solubilizing Enterobacter asburiae as influenced by fungicides. Eurasian J. Biosci. 2010, 4, 88–95. [Google Scholar] [CrossRef]
- Khalid, A.; Arshad, M.; Zahir, Z. Screening plant growth-promoting rhizobacteria for improving growth and yield of wheat. J. Appl. Microbiol. 2004, 96, 473–480. [Google Scholar] [CrossRef]
- Herrera, H.; Valadares, R.; Oliveira, G.; Fuentes, A.; Almonacid, L.; do Nascimento, S.V.; Bashan, Y.; Arriagada, C. Adaptation and tolerance mechanisms developed by mycorrhizal Bipinnula fimbriata plantlets (Orchidaceae) in a heavy metal-polluted ecosystem. Mycorrhiza 2018, 28, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Cawthray, G.R. An improved reversed-phase liquid chromatographic method for the analysis of low-molecular mass organic acids in plant root exudates. J. Chromatogr. A 2003, 1011, 233–240. [Google Scholar] [CrossRef]
- Koizumi, T.; Hattori, M.; Nara, K. Ectomycorrhizal fungal communities in alpine relict forests of Pinus pumila on Mt. Norikura, Japan. Mycorrhiza 2018, 28, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Guo, S. Fungal endophyte communities in four Rhizophoraceae mangrove species on the south coast of China. Ecol. Res. 2011, 26, 403–409. [Google Scholar] [CrossRef]
- Yuan, Z.-l.; Chen, Y.-C.; Yang, Y. Diverse non-mycorrhizal fungal endophytes inhabiting an epiphytic, medicinal orchid (Dendrobium nobile): Estimation and characterization. World J. Microbiol. Biotechnol. 2009, 25, 295. [Google Scholar] [CrossRef]
- Jia, S.; Nakano, T.; Hattori, M.; Nara, K. Root-associated fungal communities in three Pyroleae species and their mycobiont sharing with surrounding trees in subalpine coniferous forests on Mount Fuji, Japan. Mycorrhiza 2017, 27, 733–745. [Google Scholar] [CrossRef] [Green Version]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterjournalComplex Syst. 2006, 1695, 1–9. [Google Scholar]
- Visagie, C.; Houbraken, J.; Dijksterhuis, J.; Seifert, K.; Jacobs, K.; Samson, R. A taxonomic review of Penicillium species producing conidiophores with solitary phialides, classified in section Torulomyces. Pers. Mol. Phylogeny Evol. Fungi 2016, 36, 134–155. [Google Scholar] [CrossRef] [Green Version]
- Girlanda, M.; Selosse, M.; Cafasso, D.; Brilli, F.; Delfine, S.; Fabbian, R.; Ghignone, S.; Pinelli, P.; Segreto, R.; Loreto, F. Inefficient photosynthesis in the Mediterranean orchid Limodorum abortivum is mirrored by specific association to ectomycorrhizal Russulaceae. Mol. Ecol. 2006, 15, 491–504. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.; Cardinali, G.; Houbraken, J. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Bolaños, A.C.; Bononi, V.L.R.; de Mello Gugliotta, A. New records of Ganoderma multiplicatum (Mont.) Pat.(Polyporales, Basidiomycota) from Colombia and its geographic distribution in South America. Check List 2016, 12, 1948. [Google Scholar]
- You, Y.-H.; Yoon, H.-J.; Woo, J.-R.; Rim, S.-O.; Lee, J.-H.; Kong, W.-S.; Kim, J.-G. Diversity of Endophytic Fungi Isolated from the Rootlet of Pinus densiflora Colonized by Tricholoma matsutake. Korean J. Mycol. 2011, 39, 223–226. [Google Scholar] [CrossRef]
- Visagie, C.M.; Yilmaz, N.; Renaud, J.B.; Sumarah, M.W.; Hubka, V.; Frisvad, J.C.; Chen, A.J.; Meijer, M.; Seifert, K.A. A survey of xerophilic Aspergillus from indoor environment, including descriptions of two new section Aspergillus species producing eurotium-like sexual states. MycoKeys 2017, 19, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Vera, J.; Gutiérrez, M.H.; Palfner, G.; Pantoja, S. Diversity of culturable filamentous Ascomycetes in the eastern South Pacific Ocean off Chile. World J. Microbiol. Biotechnol. 2017, 33, 157. [Google Scholar] [CrossRef]
- Kernaghan, G.; Patriquin, G. Host associations between fungal root endophytes and boreal trees. Microb. Ecol. 2011, 62, 460–473. [Google Scholar] [CrossRef]
- Visagie, C.M.; Seifert, K.A.; Houbraken, J.; Samson, R.A.; Jacobs, K. Diversity of Penicillium section Citrina within the fynbos biome of South Africa, including a new species from a Protea repens infructescence. Mycologia 2014, 106, 537–552. [Google Scholar] [CrossRef]
- Visagie, C.; Renaud, J.; Burgess, K.; Malloch, D.; Clark, D.; Ketch, L.; Urb, M.; Louis-Seize, G.; Assabgui, R.; Sumarah, M. Fifteen new species of Penicillium. Pers. Mol. Phylogeny Evol. Fungi 2016, 36, 247–280. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.-J.; Kwon, O.-C.; Son, E.-S.; Yoon, D.-E.; Han, W.; Nam, J.-Y.; Yoo, Y.-B.; Lee, C.-S. Genetic diversity analysis of Ganoderma species and development of a specific marker for identification of medicinal mushroom Ganoderma lucidum. Afr. J. Microbiol. Res. 2012, 6, 5417–5425. [Google Scholar]
- Zhou, Y.; He, S.; Gong, G.; Zhang, S.; Chang, X.; Liu, N.; Sun, X.; Qi, X.; Ye, K.; Wang, Y. Soil fungal diversity in three nature reserves of Jiuzhaigou County, Sichuan Province, China. Ann. Microbiol. 2014, 64, 1275–1290. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Figura, T.; Tylová, E.; Šoch, J.; Selosse, M.-A.; Ponert, J. In vitro axenic germination and cultivation of mixotrophic Pyroloideae (Ericaceae) and their post-germination ontogenetic development. Ann. Bot. 2018. [Google Scholar] [CrossRef]
- Gebauer, G.; Preiss, K.; Gebauer, A.C. Partial mycoheterotrophy is more widespread among orchids than previously assumed. New Phytol. 2016, 211, 11–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Li, C.-M.; Han, Y.-L.; Chiang, T.-Y.; Chiang, Y.-C.; Sung, H.-M. Highly diversified fungi are associated with the achlorophyllous orchid Gastrodia flavilabella. BMC Genomics 2015, 16, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, M.K.; Whigham, D.F.; Sloan, D.; O’Malley, K.; Hodkinson, B. Orchid–fungus fidelity: A marriage meant to last? Ecology 2006, 87, 903–911. [Google Scholar] [CrossRef]
- Herrera, H.; García-Romera, I.; Meneses, C.; Pereira, G.; Arriagada, C. Orchid Mycorrhizal Interactions on the Pacific Side of the Andes from Chile. A Review. J. Soil Sci. Plant Nutr. 2019, 19, 187–202. [Google Scholar] [CrossRef]
- Waud, M.; Busschaert, P.; Lievens, B.; Jacquemyn, H. Specificity and localised distribution of mycorrhizal fungi in the soil may contribute to co-existence of orchid species. Fungal Ecol. 2016, 20, 155–165. [Google Scholar] [CrossRef]
- Pecoraro, L.; Caruso, T.; Cai, L.; Gupta, V.K.; Liu, Z.-J. Fungal networks and orchid distribution: New insights from above-and below-ground analyses of fungal communities. IMA Fungus 2018. [Google Scholar] [CrossRef] [Green Version]
- Villarreal-Ruiz, L.; Neri-Luna, C.; Anderson, I.C.; Alexander, I.J. In vitro interactions between ectomycorrhizal fungi and ericaceous plants. Symbiosis 2012, 56, 67–75. [Google Scholar] [CrossRef]
- Bidartondo, M.; Bruns, T. On the origins of extreme mycorrhizal specificity in the Monotropoideae (Ericaceae): Performance trade-offs during seed germination and seedling development. Mol. Ecol. 2005, 14, 1549–1560. [Google Scholar] [CrossRef]
- Valadares, R.; Perotto, S.; Santos, E.; Lambais, M. Proteome changes in Oncidium sphacelatum (Orchidaceae) at different trophic stages of symbiotic germination. Mycorrhiza 2014, 24, 349–360. [Google Scholar] [CrossRef]
- Kuga, Y.; Sakamoto, N.; Yurimoto, H. Stable isotope cellular imaging reveals that both live and degenerating fungal pelotons transfer carbon and nitrogen to orchid protocorms. New Phytol. 2014, 202, 594–605. [Google Scholar] [CrossRef]
- Cosme, M.; Fernández, I.; Van der Heijden, M.G.; Pieterse, C.M. Non-mycorrhizal plants: The exceptions that prove the rule. Trends Plant Sci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fracchia, S.; Aranda-Rickert, A.; Rothen, C.; Sede, S. Associated fungi, symbiotic germination and in vitro seedling development of the rare Andean terrestrial orchid Chloraea riojana. Flora 2016, 224, 106–111. [Google Scholar] [CrossRef]
- Steinfort, U.; Verdugo, G.; Besoain, X.; Cisternas, M.A. Mycorrhizal association and symbiotic germination of the terrestrial orchid Bipinnula fimbriata (Poepp.) Johnst (Orchidaceae). Flora-Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 811–817. [Google Scholar] [CrossRef]
- Bidartondo, M.I.; Burghardt, B.; Gebauer, G.; Bruns, T.D.; Read, D.J. Changing partners in the dark: Isotopic and molecular evidence of ectomycorrhizal liaisons between forest orchids and trees. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2004, 271, 1799–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waud, M.; Wiegand, T.; Brys, R.; Lievens, B.; Jacquemyn, H. Nonrandom seedling establishment corresponds with distance-dependent decline in mycorrhizal abundance in two terrestrial orchids. New Phytol. 2016, 211, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hynson, N.A.; Bruns, T.D. Fungal hosts for mycoheterotrophic plants: A nonexclusive, but highly selective club. New Phytol. 2010, 185, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Watthana, S.; Stier, A.; Richard, F.; Vessabutr, S.; Selosse, M.-A. Two mycoheterotrophic orchids from Thailand tropical dipterocarpacean forests associate with a broad diversity of ectomycorrhizal fungi. BMC Biol. 2009, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Martos, F.; Dulormne, M.; Pailler, T.; Bonfante, P.; Faccio, A.; Fournel, J.; Dubois, M.P.; Selosse, M.A. Independent recruitment of saprotrophic fungi as mycorrhizal partners by tropical achlorophyllous orchids. New Phytol. 2009, 184, 668–681. [Google Scholar] [CrossRef]
- Gorzelak, M.A.; Asay, A.K.; Pickles, B.J.; Simard, S.W. Inter-plant communication through mycorrhizal networks mediates complex adaptive behaviour in plant communities. AoB Plants 2015, 7, plv050. [Google Scholar] [CrossRef] [Green Version]
- Babikova, Z.; Gilbert, L.; Bruce, T.J.; Birkett, M.; Caulfield, J.C.; Woodcock, C.; Pickett, J.A.; Johnson, D. Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack. Ecol. Lett. 2013, 16, 835–843. [Google Scholar] [CrossRef]
- Bingham, M.A.; Simard, S.W. Do mycorrhizal network benefits to survival and growth of interior Douglas-fir seedlings increase with soil moisture stress? Ecol. Evol. 2011, 1, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.Y.; Zeng, R.S.; Xu, J.F.; Li, J.; Shen, X.; Yihdego, W.G. Interplant communication of tomato plants through underground common mycorrhizal networks. PLoS ONE 2010, 5, e13324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selosse, M.A.; Bocayuva, M.F.; Kasuya, M.C.M.; Courty, P.E. Mixotrophy in mycorrhizal plants: Extracting carbon from mycorrhizal networks. Mol. Mycorrhizal Symbiosis 2016, 451–471. [Google Scholar] [CrossRef]
- Valadares, R.B.; Pereira, M.C.; Otero, J.T.; Cardoso, E.J. Narrow fungal mycorrhizal diversity in a population of the orchid Coppensia doniana. Biotropica 2012, 44, 114–122. [Google Scholar] [CrossRef]
- Yagame, T.; Ogura-Tsujita, Y.; Kinoshita, A.; Iwase, K.; Yukawa, T. Fungal partner shifts during the evolution of mycoheterotrophy in Neottia. Am. J. Bot. 2016, 103, 1630–1641. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, A.; Ogura-Tsujita, Y.; Umata, H.; Sato, H.; Hashimoto, T.; Yukawa, T. How do fungal partners affect the evolution and habitat preferences of mycoheterotrophic plants? A case study in Gastrodia. Am. J. Bot. 2016, 103, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.; Khan, A.L.; Hamayun, M.; Shahzad, R.; Kang, S.-M.; Kim, J.-G.; Lee, I.-J. Endophytic fungi promote plant growth and mitigate the adverse effects of stem rot: An example of Penicillium citrinum and Aspergillus terreus. J. Plant Interact. 2015, 10, 280–287. [Google Scholar] [CrossRef]
- Koike, N.; Hyakumachi, M.; Kageyama, K.; Tsuyumu, S.; Doke, N. Induction of systemic resistance in cucumber against several diseases by plant growth-promoting fungi: Lignification and superoxide generation. Eur. J. Plant Pathol. 2001, 107, 523–533. [Google Scholar] [CrossRef]
- Elsharkawy, M.M.; Suga, H.; Shimizu, M. Systemic resistance induced by Phoma sp. GS8-3 and nanosilica against Cucumber mosaic virus. Environ. Sci. Pollut. Res. 2018. [Google Scholar] [CrossRef]
- Muslim, A.; Hyakumachi, M.; Kageyama, K.; Suwandi, S. Induction of Systemic Resistance in Cucumber by Hypovirulent Binucleate Rhizoctonia against Anthracnose Caused by Colletotrichum orbiculare. Trop. Life Sci. Res. 2019, 30, 109–122. [Google Scholar] [CrossRef]
- Yadav, A.N.; Verma, P.; Kumar, V.; Sangwan, P.; Mishra, S.; Panjiar, N.; Gupta, V.K.; Saxena, A.K. Biodiversity of the genus Penicillium in different habitats. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 3–18. [Google Scholar]
- Soltani, J.; Moghaddam, M.S.H. Fungal endophyte diversity and bioactivity in the mediterranean cypress Cupressus sempervirens. Curr. Microbiol. 2015, 70, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.L.; Bruns, T.D.; Szaro, T.M.; Hodges, S.A. Divergence in mycorrhizal specialization within Hexalectris spicata (Orchidaceae), a nonphotosynthetic desert orchid. Am. J. Bot. 2003, 90, 1168–1179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selosse, M.-A.; Faccio, A.; Scappaticci, G.; Bonfante, P. Chlorophyllous and achlorophyllous specimens of Epipactis microphylla (Neottieae, Orchidaceae) are associated with ectomycorrhizal septomycetes, including truffles. Microb. Ecol. 2004, 47, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wu, Y.-Y.; Zhang, T.-Y.; Zhang, M.-Y.; Peng, F.; Lin, B.; Zhang, Y.-X. New antimicrobial compounds produced by endophytic Penicillium janthinellum isolated from Panax notoginseng as potential inhibitors of FtsZ. Fitoterapia 2018, 131, 35–43. [Google Scholar] [CrossRef] [PubMed]
- McKendrick, S.; Leake, J.; Taylor, D.L.; Read, D. Symbiotic germination and development of the myco-heterotrophic orchid Neottia nidus-avis in nature and its requirement for locally distributed Sebacina spp. New Phytol. 2002, 154, 233–247. [Google Scholar] [CrossRef]
- Zeng, X.; Li, Y.; Ling, H.; Liu, S.; Liu, M.; Chen, J.; Guo, S. Transcriptomic analyses reveal clathrin-mediated endocytosis involved in symbiotic seed germination of Gastrodia elata. Bot. Stud. 2017, 58, 31. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Sampling Point 1 | Sampling Point 2 | |
|---|---|---|
| Soil chemical Characterization | ||
| N a | 52 | 28 |
| P a | 19 | 11 |
| K a | 489 | 293 |
| pH b | 6.18 | 5.51 |
| Organic matter c | 41 | 29 |
| K d | 1.25 | 0.75 |
| Na d | 0.15 | 0.15 |
| Ca d | 27.70 | 9.23 |
| Mg d | 9.50 | 3.63 |
| Al d | 0.01 | 0.27 |
| CEC d | 38.61 | 14.03 |
| Plant speciese | Eucryphia cordifolia (Ulmo) Nothofagus obliqua (Roble) Cryptocarya alba (Peumo) Peumus boldus (Boldo) Lapageria rosea (Copihue) Aristotelia chilensis (Maqui) Sophora cassioides (Pelu) Lomatia hirsuta (Radal) Chusquea quila (Quila) Persea lingue (Lingue) Drimys winteri (Canelo) Luma apiculata (Arrayan) Gevuina avellana (Avellano) Chloraea galeata Chloraea philippi | Cupressus sempervirens (Cypress) Pinus radiata (Pino) Peumus boldus (Boldo) Luma apiculata (Arrayan) Gavilea araucana |
| Fungal sporocarps | Morchella esculenta Bondarzewia guaitecasensis Cortinarius magellanicus Ramaria flaccida Ramaria stricta Russula nothofaginea | Lactarius deliciosus Suillus luteus Aleuria aurantia Amanita muscaria Amanita rubescens Mycena alcalina Russula sardonia Mycena leptocephala Mycena chusqueophila |
| Fungal Isolate | Sampling Point 1 (frequency) | Sampling Point 2 (frequency) | IAA (µg mL−1) | Phosphate Solubilization | Siderophore Production | Citric Acid (µg mL−1) | Lactic Acid (µg mL−1) | Succinic Acid (µg mL−1) | Malic Acid (µg mL−1) | Oxalic Acid (µg mL−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| AF01 | 5 (0.03) | - | 0.14 cd | - | - | 336.9 ± 45.8 b | 412.2 ± 152.2 efgh | 151.6 ± 20.4 e | 476.7 ± 87.4 d | 47.1 ± 8.5 c |
| AF02 | 2 (0.01) | - | 0.02 g | - | - | 53.9 ± 15.6 fg | 407.2 ± 99.3 fgh | 459.2 ± 48.9 c | 918.1 ± 263.4 c | - |
| AF03 | 5 (0.04) | - | 0.19 c | - | - | - | - | - | - | 4.0 ± 1.9 e |
| AF04 | 12 (0.08) | - | 0.12 de | - | - | 187.9 ± 10.2 cde | 99.5 ± 13.1 ij | 68.4 ± 22.3 e | 105.1 ± 13.6 de | - |
| AF05 | 15 (0.09) | 3 (0.01) | 0.05 fg | + | ++ | 190.8 ± 8.5 cde | 378.2 ± 51.4 ghi | 575.8 ± 68.3 c | 39.8 ± 10.2 e | 44.8 ± 7.3 cd |
| AF06 | 3 (0.02) | - | - | - | - | 130.3 ± 17.1 defg | 705.6 ± 43.9 def | 1393.7 ± 150.9 a | 1779.2 ± 68.1 a | 20.6 ± 4.5 de |
| AF07 | 11 (0.08) | - | 0.04 fg | - | - | 244.8 ± 63.4 bcd | 124.0 ± 21.9 hij | 198.5 ± 33.5 de | - | - |
| AF08 | 8 (0.06) | - | 0.03 fg | - | - | 126.8 ± 20.8 efg | 142.6 ± 17.6 hij | 96.3 ± 6.1 e | 1258.8 ± 123.4 bc | - |
| AF09 | 10 (0.08) | - | - | - | ++ | 35.4 ± 6.7 g | 1005.7 ± 95.9 cd | 21.8 ± 1.6 e | - | - |
| AF10 | 7 (0.05) | - | 0.08 efg | - | - | 160.2 ± 17.0 cdef | - | 122.1 ± 4.2 e | 23.9 ± 2.8 e | - |
| AF11 | 16 (0.11) | - | - | - | - | - | 49.0 ± 4.8 j | - | - | 41.0 ± 5.7 cd |
| AF12 | - | 32 (0.12) | 0.06 fg | - | ++ | - | 189.2 ± 12.9 hij | 31.9 ± 3.4 e | - | 14.1 ± 6.6 e |
| AF13 | - | 24 (0.09) | - | - | - | - | 6.0 ± 0.9 j | - | 1565.7 ± 113.6 ab | - |
| AF14 | - | 22 (0.08) | 0.04 fg | - | - | 320.2 ± 12.5 b | 1203.9 ± 43.1 bc | 384.3 ± 12.5 cd | 383.6 ± 34.5 de | 137.2 ± 10.0 a |
| AF15 | - | 39 (0.14) | 0.04 fg | - | + | - | 10.2 ± 1.4 j | - | - | - |
| AF16 | - | 51 (0.19) | 0.02 g | + | - | 138.8 ± 10.8 defg | 494.4 ± 25.9 efg | 207.4 ± 11.8 de | 10.8 ± 1.7 e | 3.4 ± 0.3 e |
| AF17 | 18 (0.13) | - | - | + | - | - | - | 9.2 ± 0.5 e | - | 3.4 ± 0.5 e |
| AF18 | - | 16 (0.06) | 0.44 a | + | + | 103.2 ± 4.3 efg | 711.1 ± 40.0 de | 47.8 ± 1.7 e | 386.9 ± 17.6 de | 103.8 ± 6.2 b |
| AF19 | - | 9 (0.03) | 0.13 d | - | - | 511.9 ± 11.1 a | 1452.2 ± 79.0 ab | - | 262.5 ± 7.4 de | 4.9 ± 0.3 e |
| AF20 | 7 (0.05) | 15 (0.06) | 0.14 d | + | - | 266.1 ± 13.9 bc | 713.9 ± 23.2 de | 147.7 ± 15.8 e | 22.8 ± 1.6 e | - |
| AF21 | 7 (0.05) | 21 (0.08) | 0.06 fg | + | - | 128.7 ± 15.8 efg | 1698.3 ± 70.3a | 1084.9 ± 52.8b | 22.3 ± 0.3 e | 2.6 ± 0.4 e |
| AF22 | 14 (0.10) | 26 (0.10) | 0.25 b | - | - | 180.9 ± 9.2 cde | - | 160.5 ± 19.4de | 9.3 ± 0.4 e | - |
| AF23 | 2 (0.02) | 7 (0.02) | 0.08 ef | - | - | 40.2 ± 2.1 g | 516.3 ± 13.1efg | 1016.8 ± 24.7b | 1293.8 ± 53.0 bc | 3.0 ± 0.5 e |
| AF24 | - | 8 (0.03) | - | - | - | - | - | - | - | - |
| Total strains | 142 | 273 | ||||||||
| Margalef’s richness index | 2.845 | 2.136 | ||||||||
| Shannon’s species diversity | 3.721 | 3.413 | ||||||||
| Simpson’s species diversity | 0.076 | 0.107 |
| Fungal Isolate Final Identification | GenBank Accession Numbers | Isolation Source | Close Relatives (Accession Number) | % Identity | Reference |
|---|---|---|---|---|---|
| AF01 Penicillium wollemiicola | MK826009 MN603790 | Endophyte (Sampling point 1) | Penicillium wollemiicola (KJ174314) | 99 | Visagie et al. [40] |
| AF02 Rhizoctonia sp. | MK826027 | Endophyte (Sampling point 1) | Rhizoctonia sp. (DQ061931) | 99 | Girlanda et al. [41] |
| AF03 Penicillium spinulosum | MK826010 MN603791 | Endophyte (Sampling point 1) | Penicillium spinulosum (KT316692) | 99 | Vu et al. [42] |
| AF04 Fusarium sp. | MK826011 | Endophyte (Sampling point 1) | Fusarium oxysporum (GQ121287) | 100 | GenBank |
| AF05 Ganoderma australe | MK826012 | Endophyte (Sampling point 1 and 2) | Ganoderma australe (KU569541) | 100 | Bolaños et al. [43] |
| AF06 Penicillium asperosporum | MK826026 MN603792 | Rhizosphere (Sampling point 1) | Penicillium asperosporum (JN376151) | 99 | You et al. [44] |
| AF07 Penicillium pancosmium | MK826013 MN603789 | Rhizosphere (Sampling point 1) | Penicillium pancosmium (MF803943) | 100 | Visagie et al. [45] |
| AF08 Penicillium miczynskii | MK826014 MN603786 | Rhizosphere (Sampling point 1) | Penicillium miczynskii (MH865287) | 99 | Vu et al. [42] |
| AF09 Penicillium exsudans | MK826015 MN603793 | Rhizosphere (Sampling point 1) | Penicillium exsudans (MH864309) | 99 | Vu et al. [42] |
| AF10 Penicillium sanguifluum | MK826016 MN603787 | Rhizosphere (Sampling point 1) | Penicillium sanguifluum (MH858377) | 99 | Vu et al. [42] |
| AF11 Trametes versicolor | MK826025 | Rhizosphere (Sampling point 1) | Trametes versicolor (KY824790) | 99 | GenBank |
| AF12 Penicillium sanguifluum | MK826017 MN603788 | Rhizosphere (Sampling point 2) | Penicillium sanguifluum (MH858377) | 99 | Vu et al. [42] |
| AF13 Penicillium sp. | MK826018 MN603794 | Rhizosphere (Sampling point 2) | Penicillium sp. (KY401069) | 99 | Vera et al. [46] |
| AF14 Penicillium roseopurpureum | MK826019 MN603779 | Rhizosphere (Sampling point 2) | Penicillium roseopurpureum (MH865745) | 99 | Vu et al. [42] |
| AF15 Penicillium simplicissimum | MK826020 MN603780 | Rhizosphere (Sampling point 2) | Penicillium simplicissimum (KM458844) | 99 | GenBank |
| AF16 Penicillium montanense | MK826021 MN603781 | Rhizosphere (Sampling point 2) | Penicillium montanense (HQ157959) | 99 | Kernaghan and Patriquin [47] |
| AF17 Penicillium montanense | MK826022 MN603783 | Endophyte (Sampling point 1) | Penicillium montanense (HQ157959) | 99 | Kernaghan and Patriquin [47] |
| AF18 Penicillium brunneoconidiatum | MK826007 MN603784 | Rhizosphere (Sampling point 2) | Penicillium brunneoconidiatum (JX140766) | 100 | Visagie et al. [48] |
| AF19 Penicilliumpanissanguineum | MK826008 MN603785 | Rhizosphere (Sampling point 2) | Penicillium panissanguineum (KT887862) | 99 | Visagie et al. [49] |
| AF20 Ganoderma annulare | MK826006 | Endophyte (Sampling point 1 and 2) | Ganoderma annulare (JQ520160) | 100 | Park et al. [50] |
| AF21 Penicillium montanense | MK826005 MN603782 | Endophyte (Sampling point 1 and 2) | Penicillium montanense (HQ637347) | 100 | Zhou et al. [51] |
| AF22 Phoma sp. | MK826024 | Endophyte (Sampling point 1 and 2) | Phoma herbarum (KY979198) | 100 | GenBank |
| AF23 Paraboeremia sp. | MK826004 | Endophyte (Sampling point 1 and 2) | Paraboeremia putaminum (MH858878) | 100 | Vu et al. [42] |
| AF24 Podosphaera sp. | MK826023 | Endophyte (Sampling point 2) | Podosphaera sp. (EU273519) | 100 | GenBank |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera, H.; Soto, J.; de Bashan, L.E.; Sampedro, I.; Arriagada, C. Root-Associated Fungal Communities in Two Populations of the Fully Mycoheterotrophic Plant Arachnitis uniflora Phil. (Corsiaceae) in Southern Chile. Microorganisms 2019, 7, 586. https://doi.org/10.3390/microorganisms7120586
Herrera H, Soto J, de Bashan LE, Sampedro I, Arriagada C. Root-Associated Fungal Communities in Two Populations of the Fully Mycoheterotrophic Plant Arachnitis uniflora Phil. (Corsiaceae) in Southern Chile. Microorganisms. 2019; 7(12):586. https://doi.org/10.3390/microorganisms7120586
Chicago/Turabian StyleHerrera, Hector, Javiera Soto, Luz E. de Bashan, Inmaculada Sampedro, and Cesar Arriagada. 2019. "Root-Associated Fungal Communities in Two Populations of the Fully Mycoheterotrophic Plant Arachnitis uniflora Phil. (Corsiaceae) in Southern Chile" Microorganisms 7, no. 12: 586. https://doi.org/10.3390/microorganisms7120586