Rotations with Indian Mustard and Wild Rocket Suppressed Cucumber Fusarium Wilt Disease and Changed Rhizosphere Bacterial Communities

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Preparation
2.2. Pot Experiment
2.3. Inoculation of FOC and Fusarium Wilt Disease Severity Measurement
2.4. Plant Dry Biomass Measurement and Soil Sampling
2.5. Soil Chemical Analysis
2.6. Soil DNA Extraction
2.7. Quantitative PCR Analysis
2.8. High-Throughput Amplicon Sequencing and Data Processing
2.9. Statistical Analysis
3. Results
3.1. Cucumber Fusarium Wilt Disease Severity and Seedling Biomass
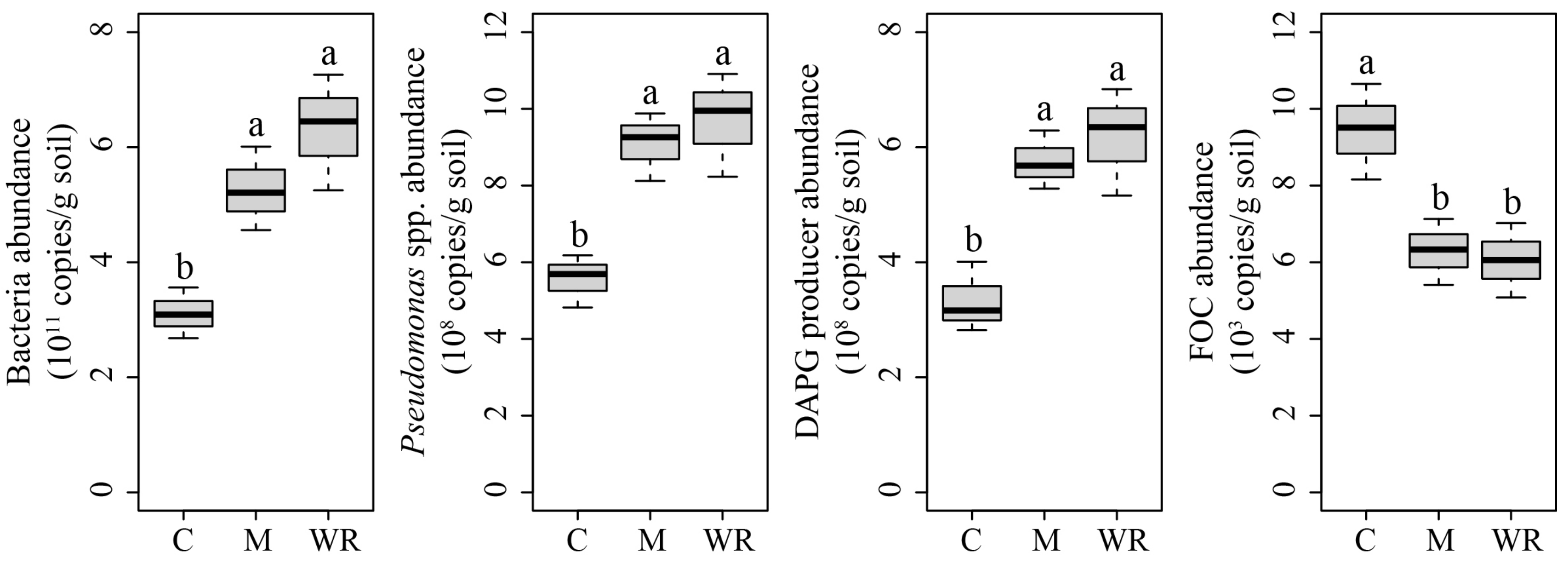
3.2. Soil Chemical Properties and Cucumber Rhizosphere Microbial Abundances
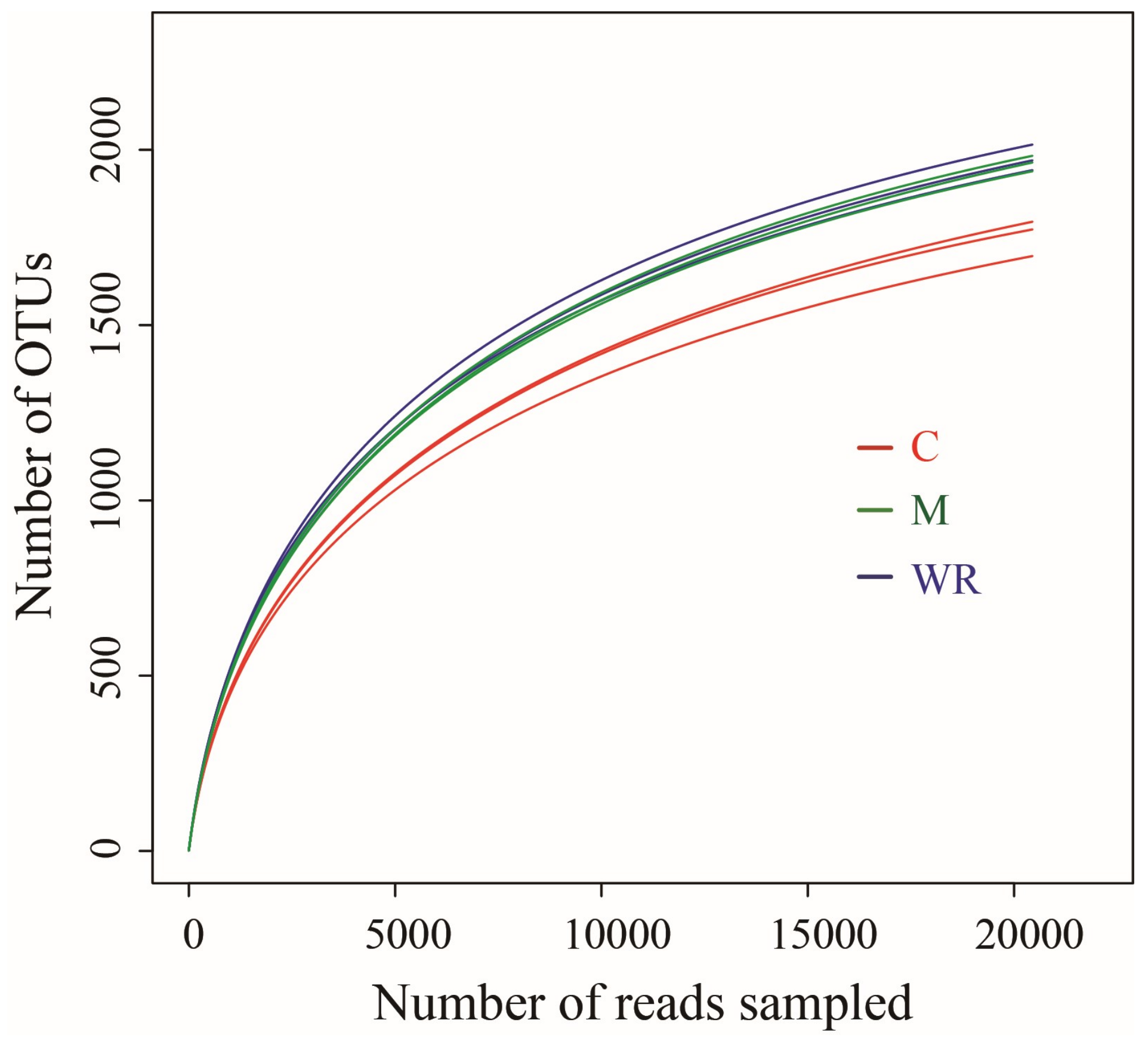
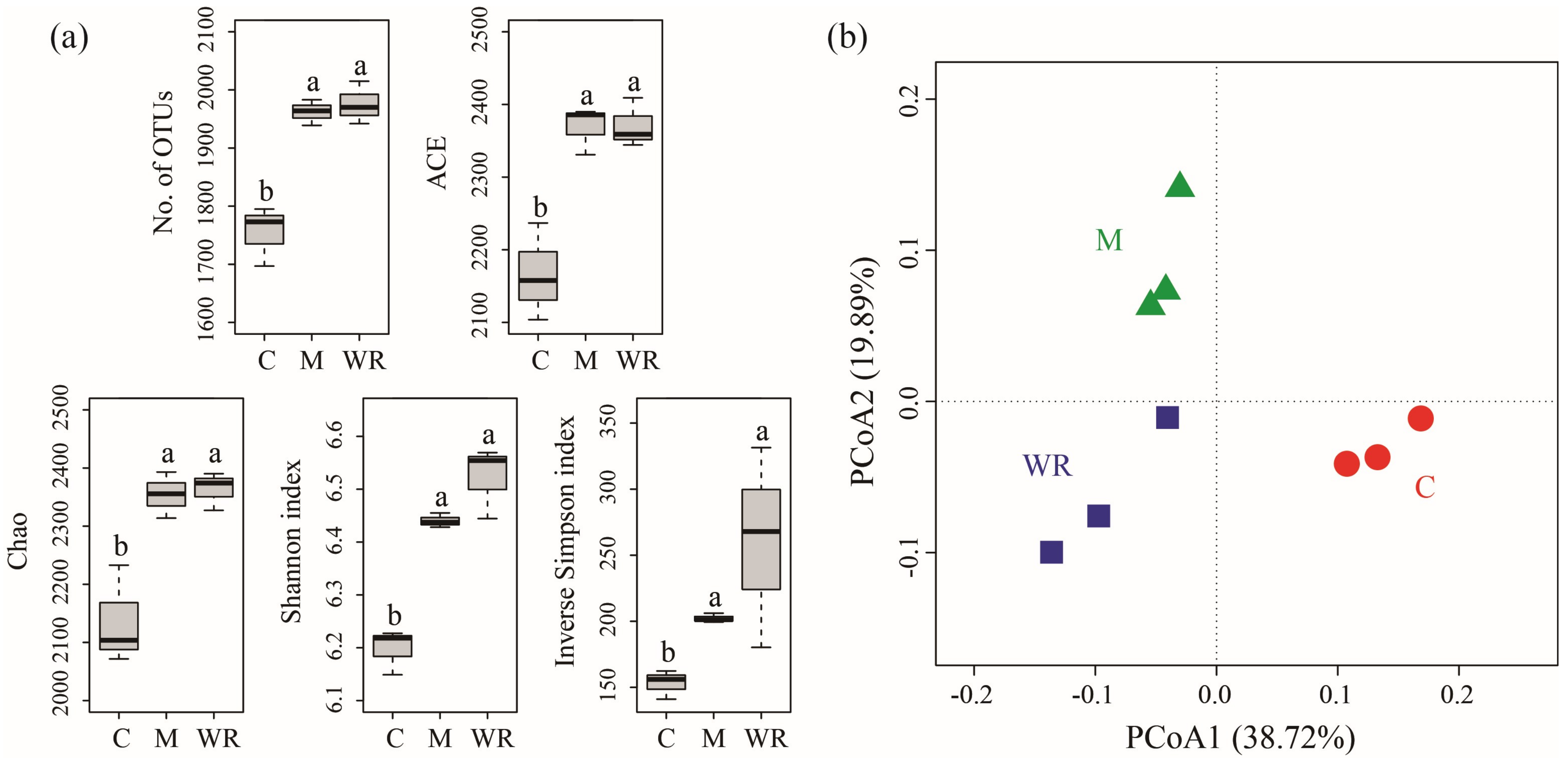
3.3. Cucumber Rhizosphere Bacterial Community Alpha and Beta Diversities
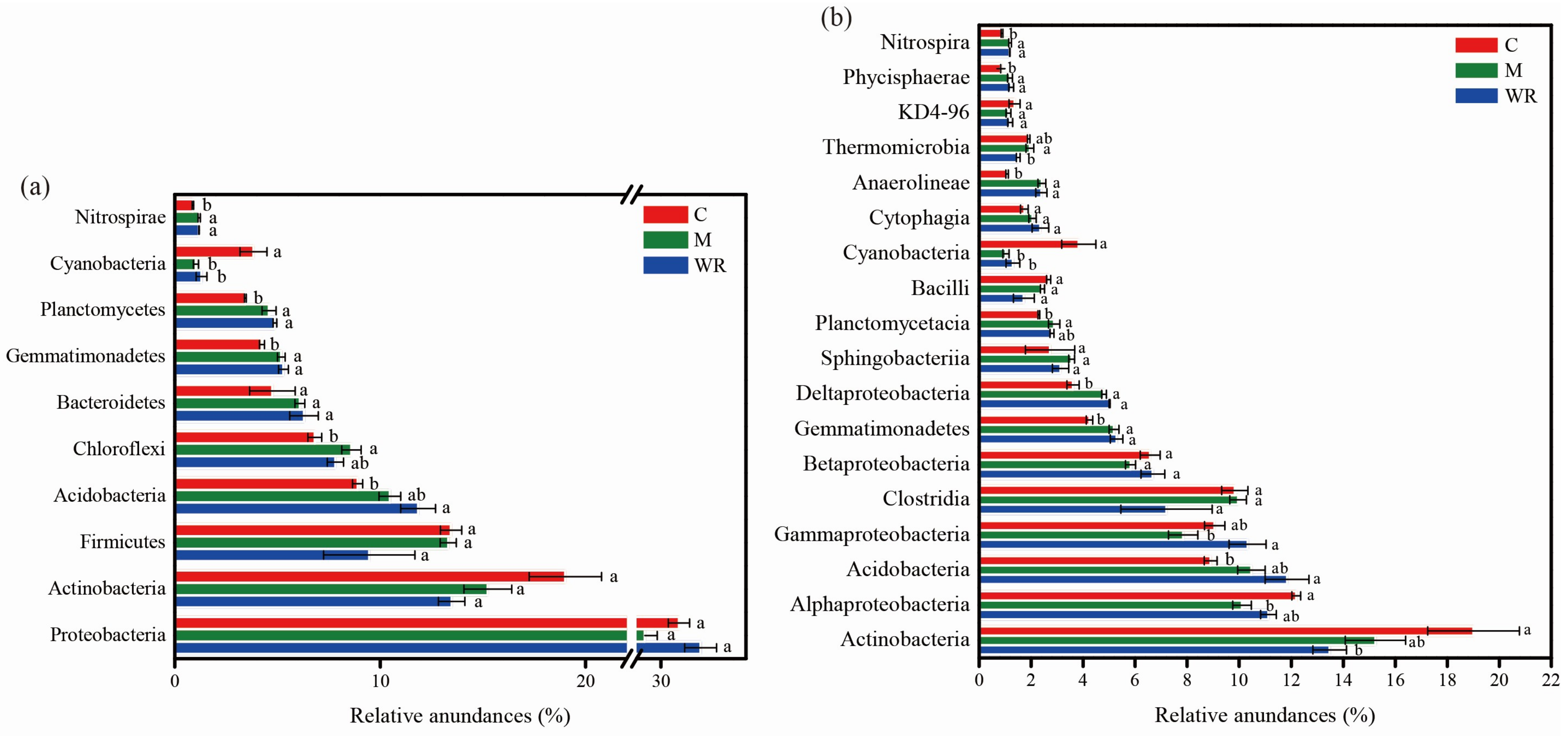
3.4. Cucumber Rhizosphere Bacterial Community Composition
3.5. Correlation between Cucumber Seedling Biomass, Fusarium Wilt Disease Severity and Bacterial Taxa Abundance
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bonanomi, G.; Antignani, V.; Pane, C.; Scala, F. Suppression of soilborne fungal diseases with organic amendments. J. Plant Pathol. 2007, 89, 311–324. [Google Scholar]
- Chakraborty, S.; Newton, A.C. Climate change, plant diseases and food security: An overview. Plant Pathol. 2011, 60, 2–14. [Google Scholar] [CrossRef]
- Bennett, A.J.; Bending, G.D.; Chandler, D.; Hilton, S.; Mills, P. Meeting the demand for crop production: The challenge of yield decline in crops grown in short rotations. Biol. Rev. 2012, 87, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liu, J.; Wu, F. Soil microbial communities in cucumber monoculture and rotation systems and their feedback effects on cucumber seedling growth. Plant Soil 2017, 415, 507–520. [Google Scholar] [CrossRef]
- Peralta, A.L.; Sun, Y.M.; McDaniel, M.D.; Lennon, J.T. Crop rotational diversity increases disease suppressive capacity of soil microbiomes. Ecosphere 2018, 9, e02235. [Google Scholar] [CrossRef]
- Zhou, X.; Yu, G.; Wu, F. Effects of intercropping cucumber with onion or garlic on soil enzyme activities, microbial communities and cucumber yield. Eur. J. Soil Biol. 2011, 47, 279–287. [Google Scholar] [CrossRef]
- Wang, B.B.; Li, R.; Ruan, Y.Z.; Ou, Y.N.; Zhao, Y.; Shen, Q.R. Pineapple-banana rotation reduced the amount of Fusarium oxysporum more than maize-banana rotation mainly through modulating fungal communities. Soil Biol. Biochem. 2015, 86, 77–86. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Wees, S.C.M.V.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Paterson, E.; Sim, A. Effect of nitrogen supply and defoliation on loss of organic compounds from roots of Festuca rubra. J. Exp. Bot. 2000, 51, 1449–1457. [Google Scholar] [CrossRef] [PubMed]
- Weisskopf, L.; Le Bayon, R.C.; Kohler, F.; Page, V.; Jossi, M.; Gobat, J.M.; Martinoia, E.; Aragno, M. Spatio-temporal dynamics of bacterial communities associated with two plant species differing in organic acid secretion: A one-year microcosm study on lupin and wheat. Soil Biol. Biochem. 2008, 40, 1772–1780. [Google Scholar] [CrossRef]
- Mendes, L.W.; Kuramae, E.E.; Navarrete, A.A.; Van Veen, J.A.; Tsai, S.M. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- Birouste, M.; Kazakou, E.; Blanchard, A.; Roumet, C. Plant traits and decomposition: Are the relationships for roots comparable to those for leaves? Ann. Bot. 2012, 109, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Maarastawi, S.A.; Frindte, K.; Linnartz, M.; Knief, C. Crop rotation and straw application impact microbial communities in Italian and Philippine soils and the rhizosphere of Zea mays. Front. Microbiol. 2018, 9, 1295. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhang, X.; Liu, J.; Chen, Q.; Gao, L. Microbial properties of rhizosphere soils as affected by rotation, grafting, and soil sterilization in intensive vegetable production systems. Sci. Hortic. 2009, 123, 139–147. [Google Scholar] [CrossRef]
- Smolinska, U. Survival of Sclerotium cepivorum sclerotia and Fusarium oxysporum chlamydospores in soil amended with cruciferous residues. J. Phytopathol. 2000, 148, 343–349. [Google Scholar] [CrossRef]
- Mowlick, S.; Yasukawa, H.; Inoue, T.; Takehara, T.; Kaku, N.; Ueki, K.; Ueki, A. Suppression of spinach wilt disease by biological soil disinfestation incorporated with Brassica juncea plants in association with changes in soil bacterial communities. Crop Prot. 2013, 54, 185–193. [Google Scholar] [CrossRef]
- Klein, E.; Ofek, M.; Katan, J.; Minz, D.; Gamliel, A. Soil suppressiveness to Fusarium disease: Shifts in root microbiome associated with reduction of pathogen root colonization. Phytopathology 2013, 103, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Inderbitzin, P.; Ward, J.; Barbella, A.; Solares, N.; Izyumin, D.; Burman, P.; Chellemi, D.O.; Subbarao, K.V. Soil microbiomes associated with Verticillium wilt-suppressive broccoli and chitin amendments are enriched with potential biocontrol agents. Phytopathology 2018, 108, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Larkin, R.P.; Griffin, T.S. Control of soilborne potato diseases using Brassica green manures. Crop Prot. 2007, 26, 1067–1077. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, T.S.; Honeycutt, C.W. Rotation and cover crop effects on soilborne potato diseases, tuber yield, and soil microbial communities. Plant Dis. 2010, 94, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Larkin, R.P.; Halloran, J.M. Management effects of disease-suppressive rotation crops on potato yield and soilborne disease and their economic implications in potato production. Am. J. Potato Res. 2014, 91, 429–439. [Google Scholar] [CrossRef]
- Michielse, C.B.; Rep, M. Pathogen profile update: Fusarium oxysporum. Mol. Plant Pathol. 2009, 10, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Latz, E.; Eisenhauer, N.; Rall, B.C.; Allan, E.; Roscher, C.; Scheu, S.; Jousset, A. Plant diversity improves protection against soil-borne pathogens by fostering antagonistic bacterial communities. J. Ecol. 2012, 100, 597–604. [Google Scholar] [CrossRef]
- Liu, L.; Kloepper, J.W.; Tuzun, S. Induction of systemic resistance in cucumber against Fusarium wilt by plant growth-promoting rhizobacteria. Phytopathology 1995, 85, 695–698. [Google Scholar] [CrossRef]
- Zhou, X.; Shen, Y.; Fu, X.; Wu, F. Application of sodium silicate enhances cucumber resistance to Fusarium wilt and alters soil microbial communities. Front. Plant Sci. 2018, 9, 624. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, J.; Pan, D.; Ge, X.; Jin, X.; Chen, S.; Wu, F. p-Coumaric can alter the composition of cucumber rhizosphere microbial communities and induce negative plant-microbial interactions. Biol. Fert. Soils 2018, 54, 363–372. [Google Scholar] [CrossRef]
- Dalton, B.R.; Weed, S.B.; Blum, U. Plant phenolic acids in soils: A comparison of extraction procedures. Soil Sci. Soc. Am. J. 1987, 51, 1515–1521. [Google Scholar] [CrossRef]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes encoding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Garbeva, P.; van Veen, J.A.; van Elsas, J.D. Assessment of the diversity, and antagonism towards Rhizoctonia solani AG3, of Pseudomonas species in soil from different agricultural regimes. FEMS Microbiol. Ecol. 2004, 47, 51–64. [Google Scholar] [CrossRef]
- Almario, J.; Moenne-Loccoz, Y.; Muller, D. Monitoring of the relation between 2,4-diacetylphloroglucinol-producing Pseudomonas and Thielaviopsis basicola populations by real-time PCR in tobacco black root-rot suppressive and conducive soils. Soil Biol. Biochem. 2013, 57, 144–155. [Google Scholar] [CrossRef]
- Scarlett, K.; Tesoriero, L.; Daniel, R.; Guest, D. Detection and quantification of Fusarium oxysporum f. sp. cucumerinum in environmental samples using a specific quantitative PCR assay. Eur. J. Plant Pathol. 2013, 137, 315–324. [Google Scholar]
- Jia, H.T.; Chen, S.C.; Yang, S.Y.; Shen, Y.H.; Qiao, P.L.; Wu, F.Z.; Zhou, X.G. Effects of vanillin on cucumber rhizosphere bacterial community. Allelopathy J. 2018, 44, 191–200. [Google Scholar]
- Schöler, A.; Jacquiod, S.; Vestergaard, G.; Schulz, S.; Schloter, M. Analysis of soil microbial communities based on amplicon sequencing of marker genes. Biol. Fert. Soils 2017, 53, 485–489. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Motisi, N.; Montfort, F.; Doré, T.; Romillac, N.; Lucas, P. Duration of control of two soilborne pathogens following incorporation of above- and below-ground residues of Brassica juncea into soil. Plant Pathol. 2009, 58, 470–478. [Google Scholar] [CrossRef]
- Hanschen, F.S.; Yim, B.; Winkelmann, T.; Smalla, K.; Schreiner, M. Degradation of biofumigant isothiocyanates and allyl glucosinolate in soil and their effects on the microbial community composition. PLoS ONE 2015, 10, e0132931. [Google Scholar] [CrossRef] [PubMed]
- Van Elsas, J.D.; Chiurazzi, M.; Mallon, C.A.; Elhottovā, D.; Krištůfek, V.; Salles, J.F. Microbial diversity determines the invasion of soil by a bacterial pathogen. Proc. Nat. Acad. Sci. USA 2012, 109, 1159–1164. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef]
- Li, R.; Shen, Z.; Sun, L.; Zhang, R.; Fu, L.; Deng, X.; Shen, Q. Novel soil fumigation method for suppressing cucumber Fusarium wilt disease associated with soil microflora alterations. Appl. Soil Ecol. 2016, 101, 28–36. [Google Scholar] [CrossRef]
- Shang, Q.; Yang, G.; Wang, Y.; Wu, X.; Zhao, X.; Hao, H.; Li, Y.; Xie, Z.; Zhang, Y.; Wang, R. Illumina-based analysis of the rhizosphere microbial communities associated with healthy and wilted Lanzhou lily (Lilium davidii var. unicolor) plants grown in the field. World J. Microbiol. Biotechnol. 2016, 32, 95. [Google Scholar] [CrossRef] [PubMed]
- Ciancio, A.; Pieterse, C.M.J.; Mercado-Blanco, J. Editorial: Harnessing useful rhizosphere microorganisms for pathogen and pest biocontrol. Front. in Microbiol. 2016, 7, 1620. [Google Scholar] [CrossRef] [PubMed]
- Hollister, E.B.; Hu, P.; Wang, A.S.; Hons, F.M.; Gentry, T.J. Differential impacts of brassicaceous and nonbrassicaceous oilseed meals on soil bacterial and fungal communities. FEMS Microbiol. Ecol. 2013, 83, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Postma, J.; Schilder, M.T. Enhancement of soil suppressiveness against Rhizoctonia solani in sugar beet by organic amendments. Appl. Soil Ecol. 2015, 94, 72–79. [Google Scholar] [CrossRef]
- Jang, M.; Hong, E.; Kim, G.H. Evaluation of antibacterial activity of 3-butenyl, 4-pentenyl, 2-phenylethyl, and benzyl Isothiocyanate in Brassica vegetables. J. Food Sci. 2010, 75, M412–M416. [Google Scholar] [CrossRef] [PubMed]
- Njoroge, S.M.C.; Riley, M.B.; Keinath, A.P. Effect of incorporation of Brassica spp. residues on population densities of soilborne microorganisms and on damping-off and Fusarium wilt of watermelon. Plant Dis. 2008, 92, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.A.; Myrold, D.D.; Bottomley, P.J. Carbon flow from 13C-labeled straw and root residues into the phospholipid fatty acids of a soil microbial community under field conditions. Soil Biol. Biochem. 2006, 38, 759–768. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, J.; Wu, F.; Zhou, X. Changes in rhizosphere microbial communities in potted cucumber seedlings treated with syringic acid. PLoS ONE 2018, 13, e0200007. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, Z.; Jia, H.; Li, L.; Wu, F. Continuously monocropped Jerusalem artichoke changed soil bacterial community composition and ammonia-oxidizing and denitrifying bacteria abundances. Front. Microbiol. 2018, 9, 705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Pan, D.D.; Ge, X.; Shen, Y.H.; Qiao, P.L.; Yang, S.Y.; Wu, F.Z.; Zhou, X.G. Effects of syringic acid on Fusarium and Trichoderma communities in cucumber (Cucumis sativus L.) seedling rhizosphere. Allelopathy J. 2018, 44, 181–190. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Bal, H.B.; Das, S.; Dangar, T.K.; Adhya, T.K. ACC deaminase and IAA producing growth promoting bacteria from the rhizosphere soil of tropical rice plants. J. Basic Microbiol. 2013, 53, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Hayward, A.C.; Fegan, N.; Fegan, M.; Stirling, G.R. Stenotrophomonas and Lysobacter: Ubiquitous plant-associated gamma-proteobacteria of developing significance in applied microbiology. J. Appl. Microbiol. 2010, 108, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.I.A.; Castro, P.M.L. Diversity and characterization of culturable bacterial endophytes from Zea mays and their potential as plant growth-promoting agents in metal-degraded soils. Environ. Sci. Pollut. Res. 2014, 21, 14110–14123. [Google Scholar] [CrossRef] [PubMed]
- Jog, R.; Nareshkumar, G.; Rajkumar, S. Plant growth promoting potential and soil enzyme production of the most abundant Streptomyces spp. from wheat rhizosphere. J. Appl. Microbiol. 2012, 113, 1154–1164. [Google Scholar] [CrossRef] [PubMed]
- Bakker, M.G.; Manter, D.K.; Sheflin, A.M.; Weir, T.L.; Vivanco, J.M. Harnessing the rhizosphere microbiome through plant breeding and agricultural management. Plant Soil 2012, 360, 1–13. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cucumber Dry Biomass (g/plant) | Fusarium Wilt Disease Index (%) | Soil pH | Soil Olsen P (mg/kg) | Soil Inorganic N (mg/kg) | Soil Phenolic Compounds (μg Ferulic Acid/g Soil) | |
|---|---|---|---|---|---|---|
| C 1 | 8.06 ± 0.32 a 2 | 66.49 ± 1.41 a | 7.64 ± 0.05 a | 284.20 ± 7.91 a | 146.59 ± 7.10 a | 217.01 ± 7.75 a |
| M | 5.06 ± 0.48 b | 49.50 ± 1.87 b | 7.45 ± 0.08 a | 266.91 ± 6.62 a | 114.09 ± 3.85 b | 169.05 ± 11.31 b |
| WR | 5.46 ± 0.46 b | 48.46 ± 4.51 b | 7.43 ± 0.07 a | 265.03 ± 5.25 a | 119.98 ± 4.15 b | 172.76 ± 5.34 b |
| C 1 | M | WR | C | M | WR | ||
|---|---|---|---|---|---|---|---|
| Clostridium sensu stricto1 | 5.84 ± 0.24 a 2 | 5.68 ± 0.07 a | 4.49 ± 1.26 a | H16 | 0.57 ± 0.06 a | 0.64 ± 0.03 a | 0.83 ± 0.08 a |
| Gaiella | 2.21 ± 0.35 a | 1.82 ± 0.17 a | 1.69 ± 0.16 a | RB41 | 0.42 ± 0.05 b | 0.74 ± 0.05 a | 0.70 ± 0.10 ab |
| Terrisporobacter | 1.65 ± 0.13 a | 1.74 ± 0.12 a | 1.18 ± 0.29 a | Microlunatus | 0.63 ± 0.02 a | 0.64 ± 0.05 a | 0.49 ± 0.09 a |
| Steroidobacter | 1.39 ± 0.15 a | 1.63 ± 0.02 a | 1.45 ± 0.10 a | Pedomicrobium | 0.53 ± 0.03 a | 0.61 ± 0.04 a | 0.50 ± 0.01 a |
| Acidibacter | 1.49 ± 0.19 a | 1.44 ± 0.09 a | 1.47 ± 0.11 a | Rhodanobacter | 0.72 ± 0.02 a | 0.30 ± 0.05 b | 0.59 ± 0.04 a |
| Nitrospira | 0.88 ± 0.03 b | 1.19 ± 0.05 a | 1.18 ± 0.01 a | Opitutus | 0.31 ± 0.03 b | 0.72 ± 0.07 a | 0.57 ± 0.05 a |
| Lysobacter | 1.40 ± 0.09 a | 0.74 ± 0.06 b | 1.06 ± 0.07 b | Pir4 lineage | 0.47 ± 0.03 a | 0.59 ± 0.08 a | 0.52 ± 0.05 a |
| Streptomyces | 1.58 ± 0.27 a | 0.64 ± 0.07 b | 0.66 ± 0.13 b | Pirellula | 0.39 ± 0.01 b | 0.57 ± 0.05 a | 0.54 ± 0.03 a |
| Actinoplanes | 0.98 ± 0.08 a | 1.02 ± 0.22 a | 0.78 ± 0.09 a | Sporosarcina | 0.55 ± 0.02 a | 0.54 ± 0.03 a | 0.38 ± 0.09 a |
| Gemmatimonas | 0.98 ± 0.08 a | 0.91 ± 0.07 a | 0.89 ± 0.03 a | Phenylobacterium | 0.54 ± 0.02 a | 0.38 ± 0.05 a | 0.47 ± 0.06 a |
| Haliangium | 0.69 ± 0.03 a | 0.91 ± 0.11 a | 0.85 ± 0.03 a | Roseiflexus | 0.41 ± 0.04 a | 0.54 ± 0.07 a | 0.42 ± 0.03 a |
| Pseudoduganella | 1.67 ± 0.15 a | 0.37 ± 0.02 b | 0.32 ± 0.02 b | Novosphingobium | 0.59 ± 0.08 a | 0.23 ± 0.04 b | 0.46 ± 0.04 ab |
| Nocardioides | 1.02 ± 0.10 a | 0.57 ± 0.02 b | 0.68 ± 0.03 b | Agromyces | 0.58 ± 0.05 a | 0.34 ± 0.03 b | 0.36 ± 0.03 b |
| Bacillus | 0.91 ± 0.04 a | 0.79 ± 0.04 ab | 0.49 ± 0.11 b | Chryseolinea | 0.42 ± 0.02 a | 0.30 ± 0.07 a | 0.53 ± 0.06 a |
| Turicibacter | 0.86 ± 0.13 a | 0.84 ± 0.02 ab | 0.48 ± 0.06 b | Solirubrobacter | 0.51 ± 0.11 a | 0.36 ± 0.05 a | 0.36 ± 0.02 a |
| Bryobacter | 0.65 ± 0.04 a | 0.79 ± 0.05 a | 0.67 ± 0.01 a | Archangium | 0.28 ± 0.05 b | 0.62 ± 0.12 a | 0.32 ± 0.04 ab |
| CSB | FWDS | CSB | FWDS | ||
|---|---|---|---|---|---|
| Clostridium sensu stricto 1 | 0.17 | 0.68 1 | H16 | −0.61 | −0.28 |
| Gaiella | 0.58 | 0.22 | RB41 | −0.58 | −0.83 |
| Terrisporobacter | −0.08 | 0.36 | Microlunatus | −0.10 | 0.43 |
| Steroidobacter | −0.52 | −0.55 | Pedomicrobium | −0.32 | −0.05 |
| Acidibacter | −0.08 | −0.13 | Rhodanobacter | 0.68 | 0.66 |
| Nitrospira | −0.67 | −0.69 | Opitutus | −0.78 | −0.56 |
| Lysobacter | 0.78 | 0.54 | Pir4 lineage | −0.60 | 0.13 |
| Streptomyces | 0.65 | 0.82 | Pirellula | −0.78 | −0.76 |
| Actinoplanes | 0.28 | 0.27 | Sporosarcina | 0.35 | 0.43 |
| Gemmatimonas | 0.43 | −0.15 | Phenylobacterium | 0.75 | 0.13 |
| Haliangium | −0.47 | −0.63 | Roseiflexus | −0.40 | −0.30 |
| Pseudoduganella | 0.57 | 0.71 | Novosphingobium | 0.60 | 0.41 |
| Nocardioides | 0.83 | 0.59 | Agromyces | 0.88 | 0.53 |
| Bacillus | 0.46 | 0.75 | Chryseolinea | 0.21 | 0.02 |
| Turicibacter | 0.08 | 0.27 | Solirubrobacter | 0.54 | 0.10 |
| Bryobacter | −0.43 | −0.17 | Archangium | −0.69 | −0.37 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, X.; Wang, J.; Li, D.; Wu, F.; Zhou, X. Rotations with Indian Mustard and Wild Rocket Suppressed Cucumber Fusarium Wilt Disease and Changed Rhizosphere Bacterial Communities. Microorganisms 2019, 7, 57. https://doi.org/10.3390/microorganisms7020057
Jin X, Wang J, Li D, Wu F, Zhou X. Rotations with Indian Mustard and Wild Rocket Suppressed Cucumber Fusarium Wilt Disease and Changed Rhizosphere Bacterial Communities. Microorganisms. 2019; 7(2):57. https://doi.org/10.3390/microorganisms7020057
Chicago/Turabian StyleJin, Xue, Jian Wang, Dalong Li, Fengzhi Wu, and Xingang Zhou. 2019. "Rotations with Indian Mustard and Wild Rocket Suppressed Cucumber Fusarium Wilt Disease and Changed Rhizosphere Bacterial Communities" Microorganisms 7, no. 2: 57. https://doi.org/10.3390/microorganisms7020057