Pantoea spp. Associated with Smooth Crabgrass (Digitaria ischaemum) Seed Inhibit Competitor Plant Species

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection
2.2. Surface Sterilization and Microbe Isolation
2.3. Bacterial Identification and Characterization
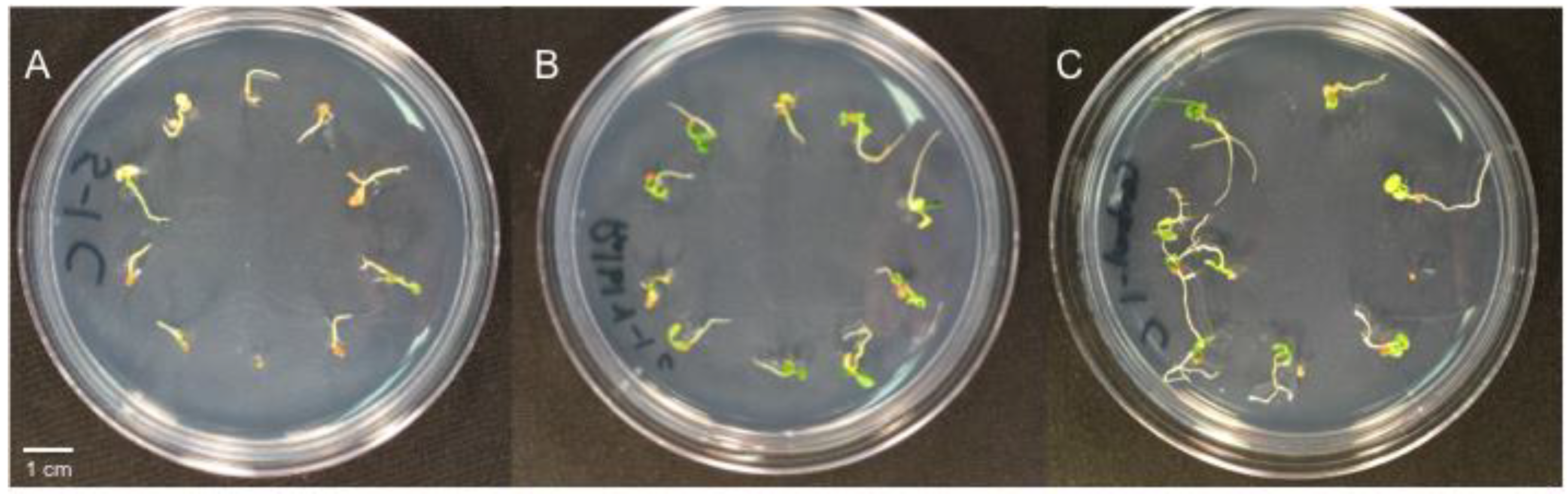
2.4. Experiment 1: Effect of Bacterial Isolates on Taraxacum Officinale Seedling Mortality
2.5. Experiment 2: Effect of Bacterial Isolates on D. ischaemum Germination and Seedling Mortality
2.6. Experiment 3: Effect of Bacterial Isolates Alone and in Combination with Curvularia sp. on D. ischaemum, T. repens and P. annua Seedlings
2.7. Statistical Analysis
3. Results
3.1. Bacterial and Fungal Isolates
3.2. Experiment 1: Effect of Bacterial Isolates on Taraxacum Officinale Seedling Mortality
3.3. Experiment 2: Effect of Bacterial Isolates on D. ischaemum Germination and Seedling Mortality
3.4. Experiment 3: Effect of Bacterial Isolates alone and in Combination with Curvularia sp. on D. ischaemum, T. repens and P. annua Seedling Mortality
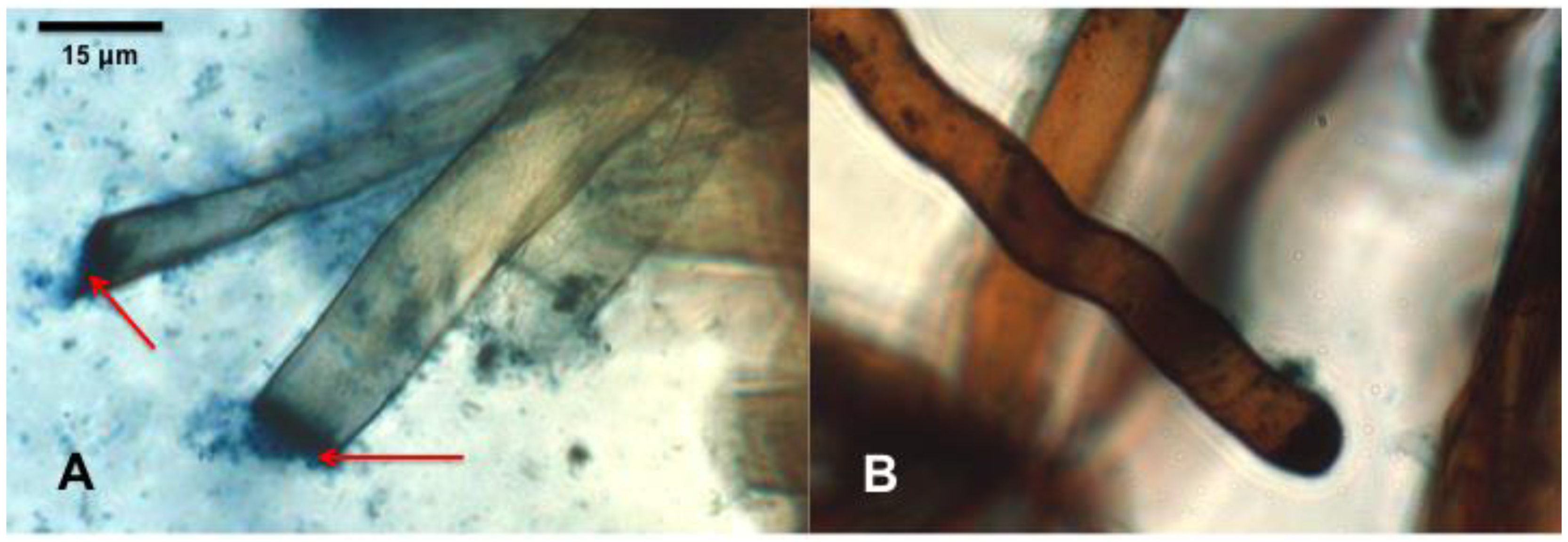
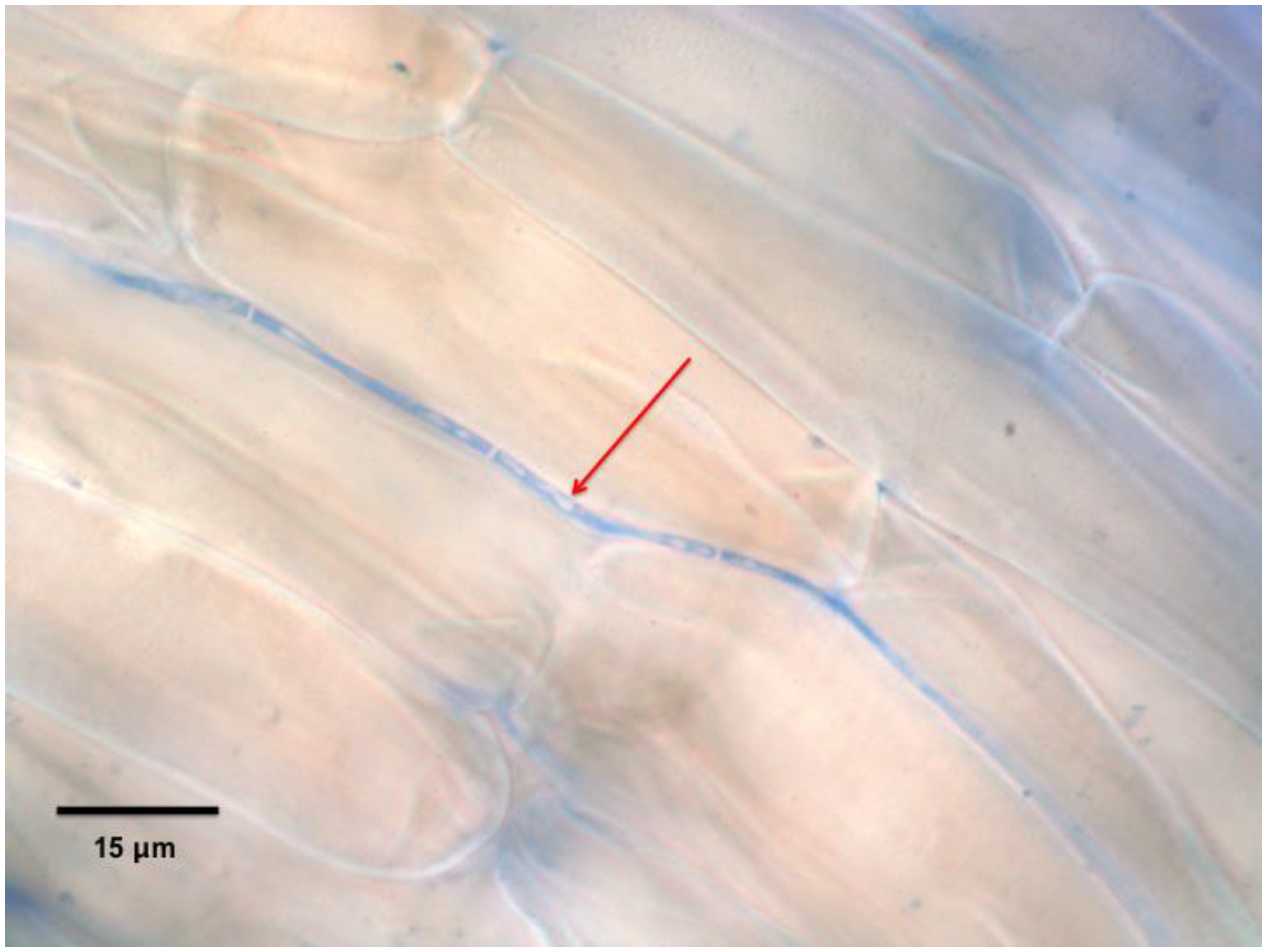
3.5. Observation of Bacteria in Seedling Roots
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kim, T.J.; Neal, J.C.; Ditomaso, J.M.; Rossi, F.S. A survey of weed scientists’ perceptions on the significance of crabgrasses (Digitaria spp.) in the United States. Weed Technol. 2002, 16, 239–242. [Google Scholar] [CrossRef]
- Zhou, B.; Kong, C.H.; Wang, P.; Li, Y.H. Chemical constituents of the essential oils of wild oat and crabgrass and their effects on the growth and allelochemical production of wheat. Weed Biol. Manag. 2013, 13, 62–69. [Google Scholar] [CrossRef]
- Zhou, B.; Kong, C.H.; Wang, P.; Li, Y.H. Crabgrass (Digitaria sanguinalis) allelochemicals that interfere with crop growth and the soil microbial community. J. Agric. Food Chem. 2013, 61, 5310–5317. [Google Scholar] [CrossRef] [PubMed]
- Chwedorzewska, K.J.; Gielwanowska, I.; Olech, M.; Molina-Montenegro, M.A.; Wódkiewicz, M.; Galera, H. Poa annua L. in the maritime Antarctic: An overview. Polar Rec. 2015, 51, 637–643. [Google Scholar] [CrossRef]
- Beard, J.B.; Rieke, P.E.; Turgeon, A.J.; Vargas, J.M. Annual Bluegrass (Poa annua L.) Description, Adaptation, Culture, and Control; Michigan State University: East Lansing, MI, USA, 1978; pp. 1–32. [Google Scholar]
- Hardoim, P.R.; van Overbeek, L.S.; van Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef]
- Truyens, S.; Weyens, N.; Cuypers, A.; Vangronsveld, J. Bacterial seed endophytes: Genera, vertical transmission and interaction with plants. Environ. Microbiol. Rep. 2015, 7, 40–50. [Google Scholar] [CrossRef]
- Wilson, D. Endophyte―The evolution of a term, and clarification on its use and definition. Oikos 1995, 73, 274–276. [Google Scholar] [CrossRef]
- Compant, S.; Clement, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo- and endosphere of plants: Their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Podolich, O.; Ardanov, P.; Zaets, I.; Pirttila, A.M.; Kozyrovska, N. Reviving of the endophytic bacterial community as a putative mechanism of plant resistance. Plant Soil 2015, 388, 367–377. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Clay, K. Fungal Endophytes of Grasses. Annu. Rev. Ecol. Syst. 1990, 21, 275–297. [Google Scholar] [CrossRef]
- Schardl, C.L. Epichloë festucae and related mutualistic symbionts of grasses. Fungal Genet. Biol. 2001, 33, 69–82. [Google Scholar] [CrossRef]
- Trognitz, F.; Hackl, E.; Widhalm, S.; Sessitsch, A. The role of plant-microbiome interactions in weed establishment and control. FEMS Microbiol. Ecol. 2016, 92, 1–15. [Google Scholar] [CrossRef]
- Shipunov, A.; Newcombe, G.; Raghavendra, A.K.H.; Anderson, C.L. Hidden diversity of endophytic fungi in an invasive plant. Am. J. Bot. 2008, 95, 1096–1108. [Google Scholar] [CrossRef]
- Aschehoug, E.T.; Metlen, K.L.; Callaway, R.M.; Newcombe, G. Fungal endophytes directly increase the competitive effects of an invasive forb. Ecology 2012, 93, 3–8. [Google Scholar] [CrossRef]
- Clay, K.; Shearin, Z.R.C.; Bourke, K.A.; Bickford, W.A.; Kowalski, K.P. Diversity of fungal endophytes in non-native Phragmites australis in the Great Lakes. Biol. Invasions 2016, 18, 2703–2716. [Google Scholar] [CrossRef]
- Ernst, M.; Mendgen, K.W.; Wirsel, S.G.R. Endophytic fungal mutualists: Seed-borne Stagonospora spp. enhance reed biomass production in axenic microcosms. Mol. Plant-Microbe Interact. 2003, 16, 580–587. [Google Scholar] [CrossRef]
- Shearin, Z.R.C.; Filipek, M.; Desai, R.; Bickford, W.A.; Kowalski, K.P.; Clay, K. Fungal endophytes from seeds of invasive, non-native Phragmites australis and their potential role in germination and seedling growth. Plant Soil 2017, 1–12. [Google Scholar] [CrossRef]
- White, J.F.; Kingsley, K.I.; Kowalski, K.P.; Irizarry, I.; Micci, A.; Soares, M.A.; Bergen, M.S. Disease protection and allelopathic interactions of seed-transmitted endophytic pseudomonads of invasive reed grass (Phragmites australis). Plant Soil 2018, 422, 195–208. [Google Scholar] [CrossRef]
- Anton, A.; Connor, H.E. Floral biology and reproduction in Poa (Poeae: Gramineae). Aust. J. Bot. 1995, 43, 577–599. [Google Scholar] [CrossRef]
- Kardol, P.; Cornips, N.J.; van Kempen, M.M.; Bakx-Schotman, J.M.; van der Putten, W. Microbe-mediated plant–soil feedback causes historical contingency effects in plant community assembly. Ecol. Monogr. 2007, 77, 147–162. [Google Scholar] [CrossRef]
- Zhou, W.-N.; White, J.F.; Soares, M.A.; Torres, M.S.; Zhou, Z.-P.; Li, H.-Y. Diversity of fungi associated with plants growing in geothermal ecosystems and evaluation of their capacities to enhance thermotolerance of host plants. J. Plant Interact. 2015, 10, 305–314. [Google Scholar] [CrossRef] [Green Version]
- Redman, R.S.; Litvintseva, A.; Sheehan, K.B.; Henson, J.M.; Rodriguez, R.J. Fungi from geothermal soils in Yellowstone National Park. Appl. Environ. Microb. 1999, 65, 5193–5197. [Google Scholar]
- Redman, R.S.; Sheehan, K.B.; Stout, R.G.; Rodrigues, R.J.; Henson, J.M. Thermotolerance generated by plant/fungal symbiosis. Science 2002, 298, 1581. [Google Scholar] [CrossRef]
- Marquez, L.; Redman, R.; Rodrigues, R.; Roossinck, M.J. A virus in a fungus in a plant. Science 2007, 315, 513–516. [Google Scholar] [CrossRef]
- Lush, W.M. Biology of Poa annua in a temperate zone golf putting green (Agrostis stolonifera/Poa annua) II. The seed bank. J. Appl. Ecol. 1988, 25, 989–997. [Google Scholar] [CrossRef]
- Rijavec, T.; Lapanje, A.; Dermastia, M.; Rupnik, M. Isolation of bacterial endophytes from germinated maize kernels. Can. J. Microbiol. 2007, 53, 802–808. [Google Scholar] [CrossRef] [PubMed]
- Sheibani-Tezerji, R.; Naveed, M.; Jehl, M.; Sessitsch, A.; Rattei, T.; Mitter, B. The genomes of closely related Pantoea ananatis maize seed endophytes having different effects on the host plant differ in secretion system genes and mobile genetic elements. Front. Microbiol. 2015, 6, 1–16. [Google Scholar] [CrossRef]
- Puente, M.E.; Li, C.Y.; Bashan, Y. Endophytic bacteria in cacti seeds can improve the development of cactus seedlings. Environ. Exp. Bot. 2009, 66, 402–408. [Google Scholar] [CrossRef]
- Ringelberg, D.; Foley, K.; Reynolds, C.M. Bacterial endophyte communities of two wheatgrass varieties following propagation in different growing media. Can. J. Microbiol. 2012, 58, 67–80. [Google Scholar] [CrossRef]
- Hodgson, S.; de Cates, C.; Hodgson, J.; Morley, N.J.; Sutton, B.C.; Gange, A.C. Vertical transmission of fungal endophytes is widespread in forbs. Ecol. Evol. 2014, 4, 1199–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zuo, S.; Xu, L.; Zou, Y.; Song, W. Study on diversity of endophytic bacterial communities in seeds of hybrid maize and their parental lines. Arch. Microbiol. 2012, 194, 1001–1012. [Google Scholar] [CrossRef]
- Videira, S.S.; de Oliveira, D.M.; de Morais, R.F.; Borges, W.L.; Baldani, V.L.D.; Baldani, J.I. Genetic diversity and plant growth promoting traits of diazotrophic bacteria isolated from two Pennisetum purpureum Schum. genotypes grown in the field. Plant Soil 2012, 356, 51–66. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics, 1st ed.; Goodfellow, M., Stackebrandt, E., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- White, J.F.; Torres, M.S.; Somu, M.P.; Johnson, H.; Irizarry, I.; Chen, Q.; Zhang, N.; Walsh, E.; Tadych, M.; Bergen, M. Hydrogen peroxide staining to visualize intracellular bacterial infections of seedling root cells. Microsc. Res. Tech. 2017. [Google Scholar] [CrossRef]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; American Phytopathological Society: St. Paul, MN, USA, 1998; Volume 1, p. 240. [Google Scholar]
- Verma, S.; Kinglsey, K.; Bergen, M.; English, C.; Elmore, M.; Kharwar, R.; White, J. Bacterial endophytes from rice cut grass (Leersia oryzoides L.) increase growth, promote root gravitropic response, stimulate root hair formation, and protect rice seedlings from disease. Plant Soil 2018, 422, 223–238. [Google Scholar] [CrossRef]
- Cottyn, B.; Regalado, E.; Lanoot, B.; DeCleene, M.; Mew, T.W.; Swings, J. Bacterial populations associated with rice seed in the tropical environment. Phytopathology 2001, 91, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Walterson, A.M.; Stavrinides, J. Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiol. Rev. 2015, 39, 968–984. [Google Scholar] [CrossRef]
- Coutinho, T.A.; Venter, S.N. Pantoea ananatis: An unconventional plant pathogen. Mol. Plant Pathol. 2009, 10, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Gagne-Bourne, F.; Aliferis, K.A.; Seguin, P.; Rani, M.; Samson, R.; Jabaji, S. Isolation and characterization of indigenous endophytic bacteria associated with leaves of switchgrass (Panicum virgatum L.) cultivars. J. Appl. Microbiol. 2013, 114, 836–853. [Google Scholar] [CrossRef] [PubMed]
- Hoon, S.K.; Cho, H.-S.; Cheong, H.; Ryu, C.-M.; Kim, J.F.; Park, S.-H. Two bacterial entophytes eliciting both plant growth promotion and plant defense on pepper (Capsicum annuum L.). J. Microbiol. Biotchnol. 2007, 17, 96–103. [Google Scholar]
- Kim, S.; Cho, W.K.; Kim, W.; Jee, H.J.; Park, C. Growth promotion of pepper plants by Pantoea ananatis B1-9 and its efficient endophytic colonization capacity in plant tissues. Plant Pathol. J. 2012, 28, 270–281. [Google Scholar] [CrossRef]
- Da Silva, J.F.; Barbosa, R.R.; Souza, A.N.; De Motta, O.V.; Teixeira, G.N. Isolation of Pantoea ananatis from sugarcane and characterization of its potential for plant growth promotion. Genet. Mol. Res. 2015, 14, 15301–15311. [Google Scholar] [CrossRef]
- Schmidt, C.S.; Mrnka, L.; Frantík, T.; Lovecká, P.; Vosátka, M. Plant growth promotion of Miscanthus × giganteus by endophytic bacteria and fungi on non-polluted and polluted soils. World J. Microbiol. Biotechnol. 2018, 34, 34–48. [Google Scholar] [CrossRef]
- Kennedy, A.C.; Young, F.L.; Elliott, L.F.; Douglas, C.L. Rhizobacteria suppressive to the weed downy brome. Soil Sci. Soc. Am. J. 1991, 55, 722–727. [Google Scholar] [CrossRef]
- Kremer, R.J.; Kennedy, A.C. Rhizobacteria as biocontrol agents of weeds. Weed Technol. 1996, 10, 601–609. [Google Scholar] [CrossRef]
- Kennedy, A.C. Pseudomonas fluorescens strains selectively suppress annual bluegrass (Poa annua L.). Biol. Control 2016, 103, 210–217. [Google Scholar] [CrossRef]
- Kremer, R.J.; Souissi, T. Cyanide production by rhizobacteria and potential for suppression of weed seedling growth. Curr. Microbiol. 2001, 43, 182–186. [Google Scholar] [CrossRef]
- Roper, M.C. Pantoea stewartii subsp. stewartii: Lessons learned from a xylem-dwelling pathogen of sweet corn. Mol. Plant Pathol. 2011, 12, 628–637. [Google Scholar] [CrossRef]
- De Maayer, P.; Chan, W.-Y.; Blom, J.; Venter, S.N.; Duffy, B.; Smits, T.H.M.; Coutinho, T.A. The large universal Pantoea plasmid LPP-1 plays a major role in biological and ecological diversification. BMC Genom. 2012, 13, 625. [Google Scholar] [CrossRef]
- Kamber, T.; Lansdell, T.A.; Stockwell, V.O.; Ishimaru, C.A.; Smits, T.H.M.; Duffy, B. Characterization of the biosynthetic operon for the antibacterial peptide herbicolin in Pantoea vagans biocontrol strain C9-1 and incidence in Pantoea species. Appl. Environ. Microbiol. 2012, 78, 4412–4419. [Google Scholar] [CrossRef]
- Weinthal, D.M.; Barash, I.; Panijel, M.; Valinski, L.; Gaba, V.; Manulis-Sasson, S. Distribution and replication of the pathogenicity plasmid pPATH in diverse populations of the gall-forming bacterium Pantoea agglomerans. Appl. Environ. Microbiol. 2007, 73, 7552–7561. [Google Scholar] [CrossRef] [PubMed]
- Vivian, A.; Murillo, J.; Jackson, R.W. The roles of plasmids in phytopathogenic bacteria: Mobile arsenals? Microbiology 2001, 147, 763–780. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Isolate No. | Isolated from | Seedling Mortality (%) † | Seedlings Injured (%) |
|---|---|---|---|
| 1 | P. annua | 0 | 21 |
| 2 | P. annua | 0 | 5 |
| 3 | P. annua | 0 | 7 |
| 4 | D. ischaemum | 19 *,§ | 44 * |
| 5 | D. ischaemum | 47 * | 87 * |
| 6 | P. annua | 0 | 6 |
| No bacteria | - | 0 | 18 |
| 7 | P. annua | 0 | 11 |
| 8 | D. ischaemum | 29 | 69 * |
| 9 | D. ischaemum | 13 | 42 * |
| 10 | D. ischaemum | 8 | 67 * |
| 11 | P. annua | 0 | 30 * |
| 12 | D. ischaemum | 10 | 67 * |
| No bacteria | - | 0 | 3 |
| 13 | D. ischaemum | 21 | 54 |
| 14 | P. annua | 0 | 31 |
| 15 | P. annua | 0 | 16 |
| 16 | D. ischaemum | 10 | 27 |
| 17 | D. ischaemum | 14 | 52 |
| 18 | D. ischaemum | 23 | 40 |
| No bacteria | - | 0 | 22 |
| 19 | P. annua | 0 | 45 |
| 20 | D. ischaemum | 0 | 41 |
| 21 | D. ischaemum | 39 * | 80 * |
| 22 | D. ischaemum | 37 * | 79 * |
| 23 | D. ischaemum | 0 | 36 |
| 24 | D. ischaemum | 0 | 46 * |
| RiLB4 | P. australis | 0 | 11 |
| Sandy LB4 | P. australis | 8 | 35 |
| No bacteria | - | 0 | 38 |
| T. officinale | T. repens | |||
|---|---|---|---|---|
| Isolate No. | Seedling Mortality (%) † | Seedlings Injured (%) | Seedling Mortality (%) | Seedlings Injured (%) |
| 4 | 12 | 49 * | 16 * | 88 * |
| 5 | 39 *,§ | 68 * | 5 | 76 * |
| 8 | 17 * | 42 * | 8 | 83 * |
| 10 | 3 | 19 | 8 | 84 * |
| 12 | 11 | 33 * | 12 | 93 * |
| 13 | 11 | 22 | 8 | 78 * |
| 17 | 4 | 40 * | 7 | 67 * |
| 18 | 0 | 29 * | 49 * | 84 * |
| 21 | 4 | 17 | 7 | 89 * |
| 22 | 4 | 40 * | 0 | 96 * |
| RiLB4 | - | - | 0 | 36 |
| Sandy LB4 | 0 | 22 | 0 | 23 |
| None | 0 | 4 | 0 | 34 |
| Bacterial Isolate | Germination (%) | Seedling Mortality (%) † | |||||
|---|---|---|---|---|---|---|---|
| D. ischaemum | D. ischaemum | T. officinale | |||||
| 14 days | 14 days | 28 days | 21 days | ||||
| 4 | 40 | abc § | 0 | b | 29 | 71 | a |
| 5 | 53 | ab | 6 | ab | 13 | 86 | a |
| 8 | 43 | ab | 18 | a | 43 | 78 | a |
| 18 | 43 | ab | 0 | ab | 7 | 33 | b |
| 22 | 33 | bc | 19 | a | 25 | 79 | a |
| 4 + 5 | 20 | c | - | - | - | 72 | a |
| none | 57 | a | 0 | b | 0 | 17 | b |
| Bacterial Isolate | Curvularia | Seedling Mortality (%) † | |||
|---|---|---|---|---|---|
| T. repens | D. ischaemum | ||||
| 4 | no | 67 | a § | 19 | b |
| 5 | no | 81 | a | 31 | b |
| 4 + 5 | no | 57 | a | 2 | b |
| RiLB4 | no | 0 | b | 0 | b |
| none | no | 4 | b | 3 | b |
| 4 | yes | 73 | a | 77 | a |
| 5 | yes | 87 | a | 63 | a |
| 4 + 5 | yes | 87 | a | 68 | a |
| RiLB4 | yes | 64 | a | 77 | a |
| none | yes | 73 | a | 83 | a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmore, M.T.; White, J.F.; Kingsley, K.L.; Diehl, K.H.; Verma, S.K. Pantoea spp. Associated with Smooth Crabgrass (Digitaria ischaemum) Seed Inhibit Competitor Plant Species. Microorganisms 2019, 7, 143. https://doi.org/10.3390/microorganisms7050143
Elmore MT, White JF, Kingsley KL, Diehl KH, Verma SK. Pantoea spp. Associated with Smooth Crabgrass (Digitaria ischaemum) Seed Inhibit Competitor Plant Species. Microorganisms. 2019; 7(5):143. https://doi.org/10.3390/microorganisms7050143
Chicago/Turabian StyleElmore, Matthew T., James F. White, Kathryn L. Kingsley, Katherine H. Diehl, and Satish K. Verma. 2019. "Pantoea spp. Associated with Smooth Crabgrass (Digitaria ischaemum) Seed Inhibit Competitor Plant Species" Microorganisms 7, no. 5: 143. https://doi.org/10.3390/microorganisms7050143