Moraxella nonliquefaciens and M. osloensis Are Important Moraxella Species That Cause Ocular Infections

 ,
, 
 and
and
Abstract
1. Introduction
2. Methods
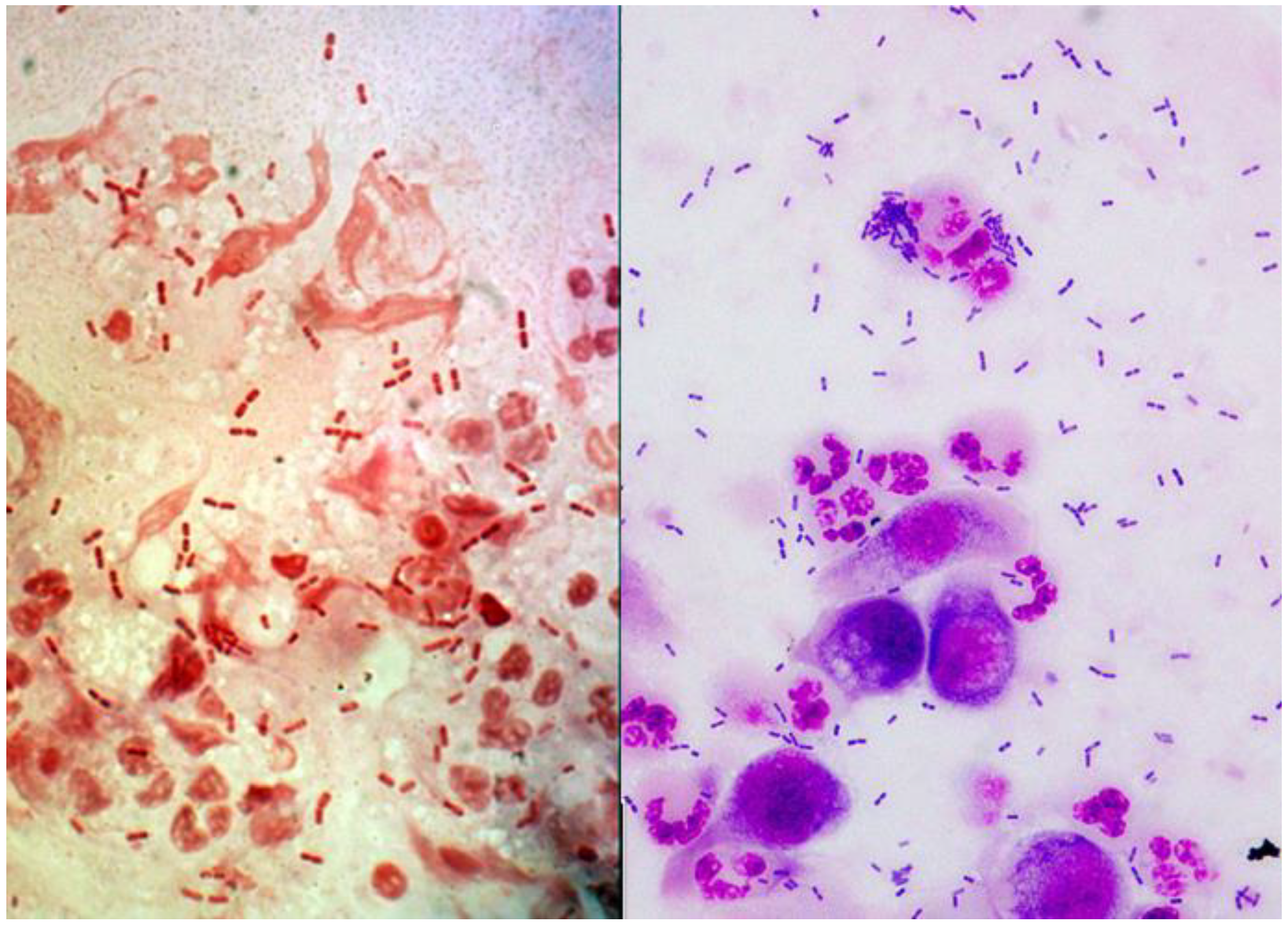
2.1. Laboratory Diagnosis of Ocular Moraxella Infection
2.2. Antibiotic Susceptibility of Ocular Moraxella
2.3. Moraxella Study Isolates
2.4. DNA Sequencing
2.5. MALDI-TOF MS
2.6. Biolog Gen III Plate
2.7. Identification of Moraxella to Species
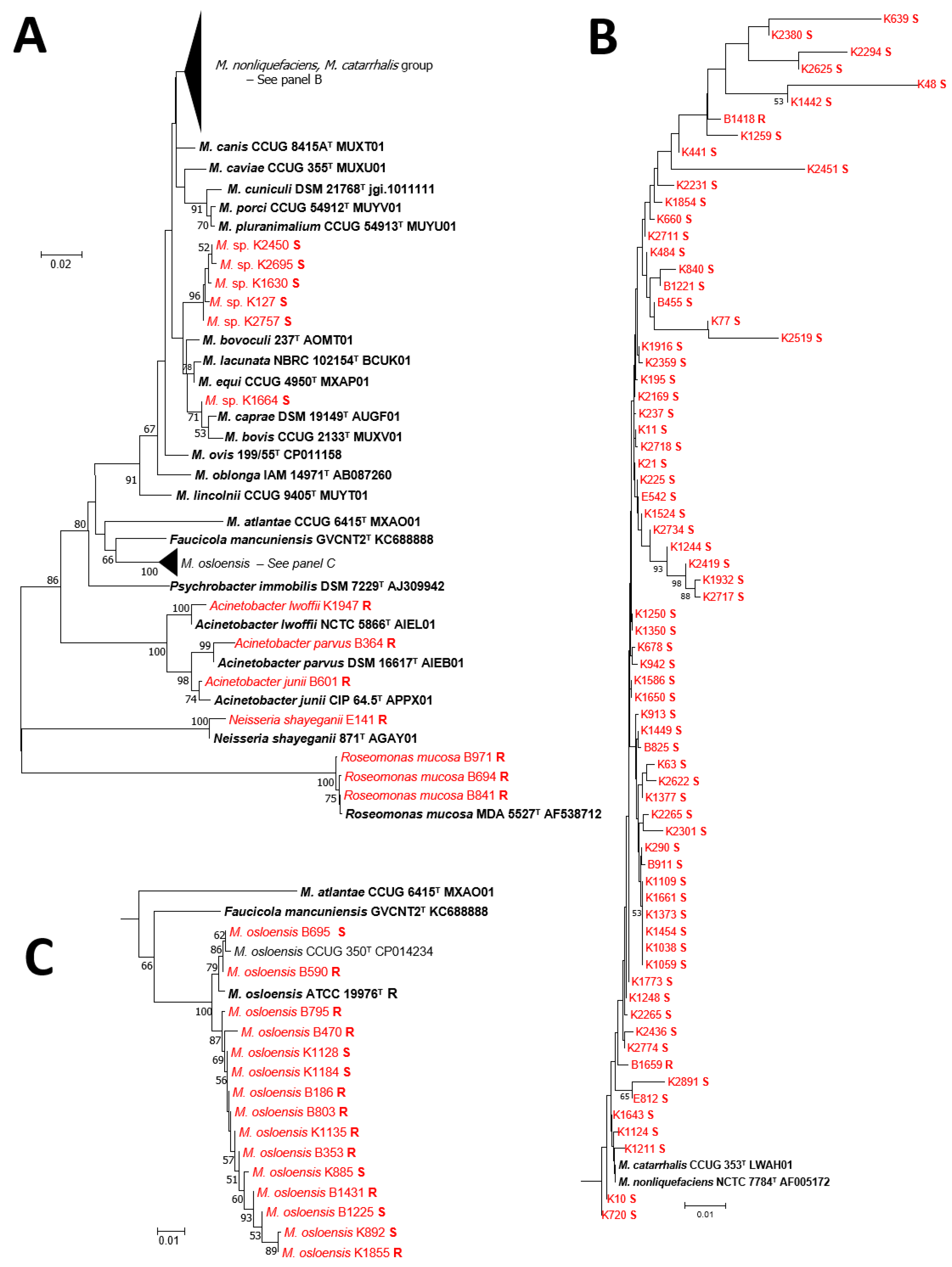
3. Results
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Morax, V. Note sur un diplobacille pathogéne pour la conjunctivite humaine. Ann. Inst. Pasteur. 1896, 10, 337–345. [Google Scholar]
- Tobimatsu, Y.; Inada, N.; Shoji, J.; Yamagami, S. Clinical characteristics of 17 patients with Moraxella keratitis. Sem. Ophthalmol. 2018, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zafar, H.; Tan, S.Z.; Walkden, A.; Fullwood, C.; Au, L.; Brahma, A.; Carley, F. Clinical characteristics and outcomes of Moraxella keratitis. Cornea 2018, 37, 1551–1554. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Suzuki, T.; Inoue, T.; Hattori, T.; Nejima, R.; Todokoro, D.; Hoshi, S.; Eguchi, H.; Miyamoto, H.; Ohashi, Y. Clinical characteristics and bacteriological profile of Moraxella keratitis. Cornea 2015, 34, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Durrani, A.; Faith, S.C.; Kowalski, R.P.; Yu, M.; Romanowski, E.G.; Shanks, R.M.; Dhaliwal, D.K.; Jhanji, V. Moraxella keratitis: Analysis of risk factors, clinical characteristics, management and treatment outcomes. Am. J. Ophthalmol. 2018, 18, 30517–30518. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, R.P.; Harwick, J.C. Incidence of Moraxella conjunctival infection. Am. J. Ophthalmol. 1986, 101, 437–440. [Google Scholar] [CrossRef]
- Ebright, J.R.; Lentino, J.R.; Juni, E. Endophthalmitis caused by Moraxella nonliquefaciens. Am. J. Clin. Pathol. 1982, 7, 362–363. [Google Scholar] [CrossRef]
- Tan, S.Z.; Walkden, A.; Au, L.; Fullwood, C.; Hamilton, A.; Qamruddin, A.; Armstrong, M.; Brahma, A.K.; Carley, F. Twelve-year analysis of microbial keratitis at a UK tertiary hospital. Eye 2017, 31, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Thygeson, P.; Kimura, S. Chronic conjunctivitis. Trans. Am. Acad. Ophthalomol. Otolaryngol. 1963, 63, 494–517. [Google Scholar]
- Dawson, C. Follicular conjunctivitis. Conjunctivitis. In Clinical Ophthalmology External Disease; Duane, T.D., Ed.; Harper and Row: Philadelphia, PA, USA, 1983; pp. 1–19. [Google Scholar]
- Balliart, P.; Tillé, H. Lésions cutanées tenaces des paupiéres et de la face, et lesions des conjonctives dues au diplobacille de Morax. Bull. Soc. Ophtalmol. 1935, 157–159. [Google Scholar]
- Mitsui, Y.; Hinokuma, S.; Tanaka, C. Etiology of angular conjunctivitis. Am. J. Ophthalmol. 1951, 34, 1579–1586. [Google Scholar] [CrossRef]
- Van Bijsterveld, O.P. Acute conjunctivitis and Moraxella. Am. J. Ophthalmol. 1967, 63, 1702–1705. [Google Scholar] [CrossRef]
- Murray, P.R. What is new in clinical microbiology-Microbial identification by MALDI-TOF Mass Spectrometry. J. Mol. Diagn. 2012, 14, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Enright, M.C.; Carter, P.E.; MacLean, I.A.; McKenzie, H. Phylogenetic relationships between some members of the genera Neisseria, Acinetobacter, Moraxella, and Kingella based on partial 16S ribosomal DNA sequence analysis. Int. J. Syst. Bacteriol. 1994, 44, 387–391. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Clinical and Laboratory Standards. Performance Standards for Antimicrobial Disk Susceptibility Tests, 10th ed.; Approved standard; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2009; document M02-A10, vol.29, No.1. [Google Scholar]
- Frank, J.A.; Reich, C.I.; Sharma, S.; Weisbaum, J.S.; Wilson, B.A.; Olsen, G.J. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl. Environ. Microbiol. 2008, 74, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Nucleotide Blast [Internet]. Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. 2004. Available online: https://blast.ncbi.nlm.nih.gov (accessed on 22 January 2018).
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Khot, P.D.; Couturier, M.R.; Wilson, A.; Croft, A.; Fisher, M.A. Optimization of matrix-assisted laser desorpotion ionization-time of flight mass spectrometry analysis for bacterial identification. J. Clin. Microbiol. 2012, 50, 3845–3852. [Google Scholar] [CrossRef] [PubMed]
- Forbes, B.A.; Sahm, D.F.; Weissfeld, A.S. (Eds.) Bailey and Scott’s―Diagnostic Microbiology. In Neisseria and Moraxella catarrhalis, 12th ed.; Mosby Elsevier: St Louis, MO, USA, 2007; pp. 447–454. [Google Scholar]
- Wallace, R.J.; Nash, D.R.; Steingrube, V.A. Antibiotic susceptibilities and drug resistance in Moraxella (Branhamella) catarrhalis. Am. J. Med. 1990, 88, 46S–50S. [Google Scholar] [CrossRef]
- Barbé, G.; Babolat, M.; Boeufgras, J.M.; Monget, D.; Frene, J. Evaluation of API NH, a new 2-hour system for identification of Neisseria and Haemophilus species and Moraxella catarrhalis in routine clinical labotatory. J. Clin. Microbiol. 1994, 32, 187–189. [Google Scholar] [PubMed]
- Goyal, S.; Warner, D.B. Roseomonas keratitis after remote penetrating keratoplasty. Arch. Clin. Exp. Ophthalmol. 2013, 251, 1025–1027. [Google Scholar] [CrossRef] [PubMed]
- Bhende, M.; Karpe, A.; Sukanya, A.; Therese, K.L.; Biswas, J. Endogenous endophthalmitis due to Roseomonas mucosa presenting as a subretinal abscess. J. Ophthalmic Inflamm. Inf. 2017, 7, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.J.; Lai, C.C.; Kuo, Y.H.; Wu, W.C.; Chen, T.L. Chronic postoperative Roseomonas endophthalmitis. J. Clin. Microbiol. 2009, 47, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Graham, D.R.; Band, J.D.; Thornsberry, C.; Hollis, D.G.; Weaver, R.E. Infections caused by Moraxella, Moraxella urethalis, Moraxella-like groups M-5 and M-6, and Kingella kingae in the United Sates, 1953–1980. Rev. Inf. Dis. 1990, 12, 423–431. [Google Scholar] [CrossRef]
- Public Health England. Identification of Moraxella Species and Morphologically Similar Organisms. UK Standards for Microbiology Investigations. ID 11 Issue 3. 2015. Available online: https:www.gov.uk/uk-standards-for-microbiology-investigations-smi-quality-and-consistency-in-clinical laboratories (accessed on 3 June 2019).




{kind=link}
{kind=link}
{kind=link}
| Isolate | Van S/R | Identification Based on: (DNA Sequencing with Vancomycin Susceptibility) | Biolog ID | MALDI-TOF MS |
|---|---|---|---|---|
| ATCC Controls | ||||
| Moraxella bovis 10900 | S | M. bovis | M. bovis | M. bovis |
| M. caviae 14659 | R | M. caviae | M. caviae | No ID |
| M. cuniculi 14688 | R | M. cuniculi | M. cuniculi | M. catarrhalis |
| M. nonliquefaciens 19975 | S | M. nonliquefaciens | No ID | M. nonliquefaciens |
| M. osloensis 19976 | R | M. osloensis | M. osloensis | M. osloensis |
| M. atlantae 29525 | I | M. atlantae | No ID | M. atlantae |
| M. lincolnii 51388 | Q | M. lincolnii | No ID | M. lincolnii |
| M. lacunata 17967 | Q | M. lacunata | No ID | M. lacunata |
| M. catarrhalis 24250 | R | M. catarrhalis | M. catarrhalis | M. catarrhalis |
| Keratitis Isolates | ||||
| 1. K10 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 2. K11 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 3. K21 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 4. K48 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 5. K63 | S | M. nonliquefaciens | M. equi | M. nonliquefaciens |
| 6. K77 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 7. K127 | S | M. equi, M. lacunata, M. bovoculi | M. nonliquefaciens | M. lacunata, M. bovis |
| 8. K195 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 9. K225 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 10. K237 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 11. K290 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 12. K441 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 13. K484 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 14. K639 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| Keratitis Isolate | ||||
| 15. K660 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 16. K678 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 17. K720 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 18. K840 | S | M. nonliquefaciens | M. equi | M. nonliquefaciens |
| 19. K885 | S | M. osloensis | M. canis | M. osloensis |
| 20. K892 | S | M. osloensis | M. osloensis | M. osloensis |
| 21. K913 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 22. K942 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 23. K1038 | S | M. nonliquefaciens | M. equi | M. nonliquefaciens |
| 24. K1059 | S | M. nonliquefaciens | M. bovis | M. nonliquefaciens |
| 25. K1109 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 26. K1124 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 27. K1128 | S | M. osloensis | M. osloensis | M. osloensis |
| 28. K1135 | R | M. osloensis | M. osloensis | M. osloensis |
| 29. K1184 | S | M. osloensis | M. nonliquefaciens | M. nonliquefaciens |
| 30. K1211 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 31. K1219 | S | M. nonliquefaciens | No ID | No ID |
| 32. K1244 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 33. K1248 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 34. K1250 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 35. K1259 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 36. K1350 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 37. K1361B | R | Acinetobacter lwoffii | M. osloensis | Acinetobacter lwoffii |
| 38. K1373 | S | M. nonliquefaciens | M. catarrhalis | M. nonliquefaciens |
| 39. K1377 | S | M. nonliquefaciens | No ID | No ID |
| 40. K1442 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 41. K1449 | S | M. nonliquefaciens | No ID | No ID |
| 42. K1454 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 43. K1524 | S | M. nonliquefaciens | M. equi | M. nonliquefaciens |
| 44. K1586 | S | M. nonliquefaciens | M. equi | M. nonliquefaciens |
| 45. K1630 | S | M. equi, M. bovoculi, M. lacunata | M. equi | M. lacunata |
| 46. K1643 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 47. K1650 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 48. K1661 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 49. K1664 | S | M. equi, M. bovoculi, M. lacunata | No ID | No ID |
| 50. K1773 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 51. K1784 | S | M. equi, M. bovoculi, M. lacunata | M. ovis | No ID |
| 52. K1661 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 53. K1854 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 54. K1855 | R | M. osloensis | M. osloensis | M. osloensis |
| 55. K1916 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 56. K1932 | S | M. nonliquefaciens | M. caprae | |
| 57. K1947 | R | Acinetobacter lwoffii | Acinetobacter lwoffi | Acinetobacter lwoffii |
| 58. K2169 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 59. K2231 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 60. K2265 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 61. K2275 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 62. K2294 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 63. K2301 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 64. K2359 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 65. K2380 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 66. K2419 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 67. K2436 | S | M. nonliquefaciens | M. equi | |
| 68. K2450 | S | M. equi, M. bovoculi, M. lacunata | M. nonliquefaciens | M. lacunata |
| 69. K2451 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 70. K2519 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 71. K2565 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 72. K2622 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 73. K2625 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 74. K2695 | S | M. equi, M. bovoculi, M. lacunata | M. nonliquefaciens | M. lacunata |
| 75. K2711 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 76. K2717 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 77. K2718 | S | M. nonliquefaciens | M. bovis | M. bovis |
| 78. K2734 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 79. K2757 | S | M. equi, M. bovoculi, M. lacunata | No ID | M. lacunata |
| 80. K2774 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 81. K2880 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 82. K2891 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| Conjunctivitis Isolates | ||||
| 1. B186 | R | M. osloensis | M. osloensis | M. osloensis |
| 2. B353 | R | M. osloensis | M. osloensis | M. osloensis |
| 3. B364 | R | Acinetobacter parvus | No ID | Acinetobacter parvus |
| 4. B455 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 5. B470 | R | M. osloensis | M. osloensis | M. osloensis |
| 6. B590 | R | M. osloensis | M. osloensis | No ID |
| 7. B601 | R | Acinetobacter junii | Acinetobacter junii | Acinetobacter junii |
| 8. B662 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 9. B694 | R | Roseomonas mucosa | No ID | Roseomonas mucosa |
| 10. B695 | S | M. osloensis | M. osloensis | M. osloensis |
| 11. B795 | R | M. osloensis | M. canis | M. osloensis |
| 12. B803 | R | M. osloensis | No ID | M. osloensis |
| 13. B825 | S | M. nonliquefaciens | No ID | No ID |
| 14. B841 | R | Roseomonas mucosa | No ID | Roseomonas mucosa |
| 15. B911 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 16. B971 | R | Roseomonas mucosa | No ID | Roseomonas mucosa |
| 17. B1221 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 18. B1225 | S | M. osloensis | M. bovis | M. osloensis |
| 19. B1418 | R | M. nonliquefaciens | M. equi | M. nonliquefaciens |
| 20. B1431 | R | M. osloensis | M. osloensis | M. osloensis |
| 21. B1659 | R | M. nonliquefaciens | M. catarrhalis | M. catarrhalis |
| Endophthalmitis Isolates | ||||
| 1. E141 | R | Neisseria shayeganii | No ID | No ID |
| 2. E542 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| 3. E614 | S | M. nonliquefaciens | M. bovis | M. nonliquefaciens |
| 4. E812 | S | M. nonliquefaciens | M. nonliquefaciens | M. nonliquefaciens |
| Keratitis (n = 82) | Incidence (percent) |
| Moraxella nonliquefaciens | 66 (80.5%) |
| Moraxella lacunata | 7 (9.0%) |
| Moraxella osloensis | 5 (6.0%) |
| Acinetobacter lwoffii | 2 (2.5%) |
| Moraxella bovis/nonliquefaciens | 1 (1.0%) |
| Moraxella osloensis/nonliquefaciens | 1 (1.0%) |
| Conjunctivitis (n = 21) | |
| Moraxella osloensis | 9 (43.0%) |
| Moraxella nonliquefaciens | 6 (29.0%) |
| Roseomonas mucosa | 3 (14.0%) |
| Acinetobacter (parvus, junii) | 2 (9.5%) |
| Moraxella catarrhalis/nonliquefaciens | 1 (4.5%) |
| Endophthalmitis (n = 4) | |
| Moraxella nonliquefaciens | 3 (75%) |
| Neisseria shayeganii | 1 (25%) |
| All Infections | |
| Moraxella nonliquefaciens | 70% (75 of 107) |
| Moraxella osloensis | 13% (14 of 107) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
LaCroce, S.J.; Wilson, M.N.; Romanowski, J.E.; Newman, J.D.; Jhanji, V.; Shanks, R.M.Q.; Kowalski, R.P. Moraxella nonliquefaciens and M. osloensis Are Important Moraxella Species That Cause Ocular Infections. Microorganisms 2019, 7, 163. https://doi.org/10.3390/microorganisms7060163
LaCroce SJ, Wilson MN, Romanowski JE, Newman JD, Jhanji V, Shanks RMQ, Kowalski RP. Moraxella nonliquefaciens and M. osloensis Are Important Moraxella Species That Cause Ocular Infections. Microorganisms. 2019; 7(6):163. https://doi.org/10.3390/microorganisms7060163
Chicago/Turabian StyleLaCroce, Samantha J., Mollie N. Wilson, John E. Romanowski, Jeffrey D. Newman, Vishal Jhanji, Robert M. Q. Shanks, and Regis P. Kowalski. 2019. "Moraxella nonliquefaciens and M. osloensis Are Important Moraxella Species That Cause Ocular Infections" Microorganisms 7, no. 6: 163. https://doi.org/10.3390/microorganisms7060163
APA StyleLaCroce, S. J., Wilson, M. N., Romanowski, J. E., Newman, J. D., Jhanji, V., Shanks, R. M. Q., & Kowalski, R. P. (2019). Moraxella nonliquefaciens and M. osloensis Are Important Moraxella Species That Cause Ocular Infections. Microorganisms, 7(6), 163. https://doi.org/10.3390/microorganisms7060163