Large Plasmid Complement Resolved: Complete Genome Sequencing of Lactobacillus plantarum MF1298, a Candidate Probiotic Strain Associated with Unfavorable Effect


Abstract
1. Introduction
2. Materials and Methods
2.1. Growth Conditions and DNA Preparation
2.2. Genome Sequencing and Assembly
2.3. Genomic Analysis
3. Results and Discussion
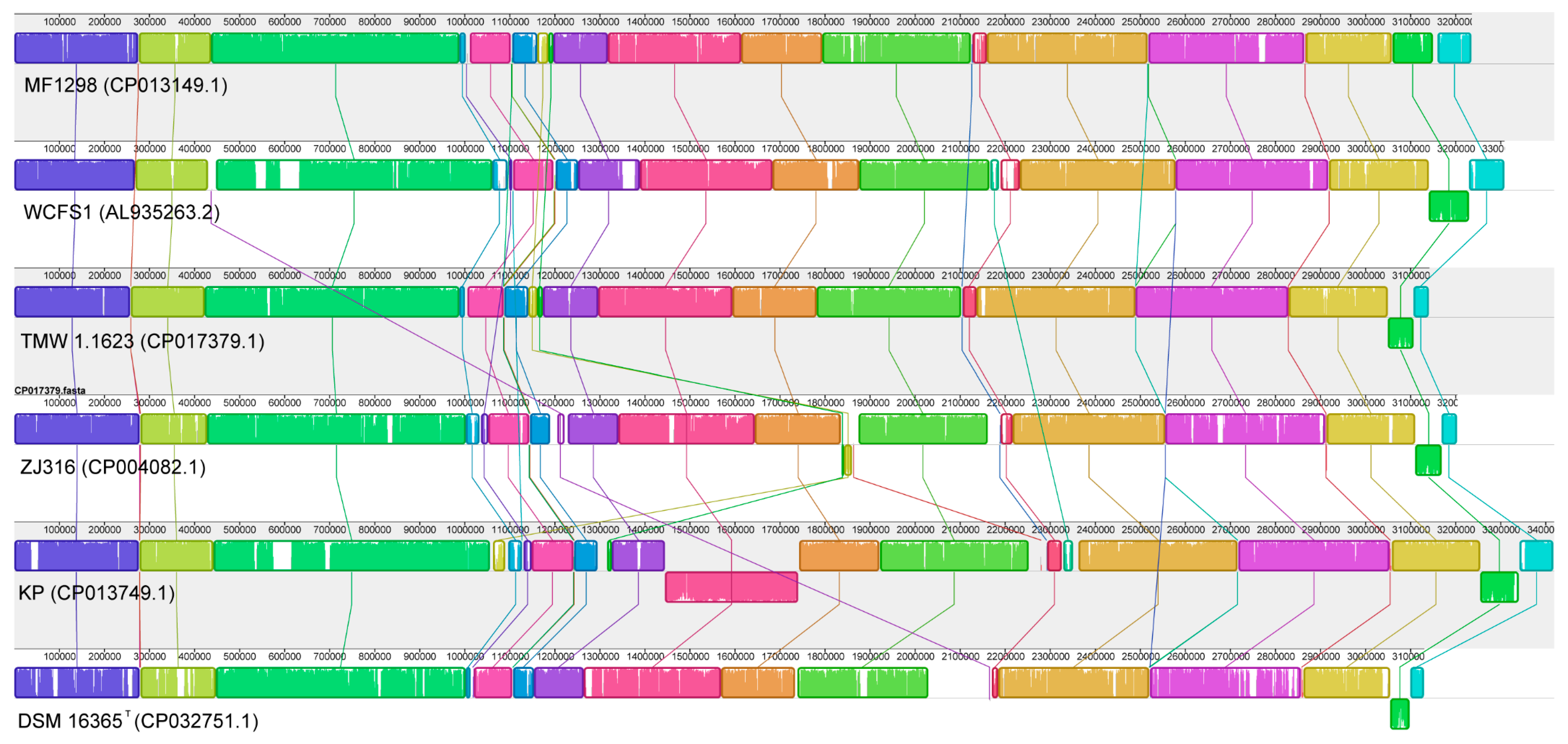
3.1. Genome Characteristics
3.2. The Plasmid Complement
3.3. Probiotic Potential
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siezen, R.J.; Tzeneva, V.A.; Castioni, A.; Wels, M.; Phan, H.T.; Rademaker, J.L.; Starrenburg, M.J.; Kleerebezem, M.; Molenaar, D.; van Hylckama Vlieg, J.E. Phenotypic and genomic diversity of Lactobacillus plantarum strains isolated from various environmental niches. Environ. Microbiol. 2010, 12, 758–773. [Google Scholar] [CrossRef] [PubMed]
- Martino, M.E.; Bayjanov, J.R.; Caffrey, B.E.; Wels, M.; Joncour, P.; Hughes, S.; Gillet, B.; Kleerebezem, M.; van Hijum, S.A.; Leulier, F. Nomadic lifestyle of Lactobacillus plantarum revealed by comparative genomics of 54 strains isolated from different habitats. Environ. Microbiol. 2016, 18, 4974–4989. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.J.; van Hylckama Vlieg, J.E. Genomic diversity and versatility of Lactobacillus plantarum, a natural metabolic engineer. Microb. Cell Fact. 2011, 10, S3. [Google Scholar] [CrossRef] [PubMed]
- Connelly, P. Lactobacillus plantarum—A literature review of therapeutic benefits. J. Aust. Tradit. Med. Soc. 2008, 14, 79–82. [Google Scholar]
- Klarin, B.; Molin, G.; Jeppsson, B.; Larsson, A. Use of the probiotic Lactobacillus plantarum 299 to reduce pathogenic bacteria in the oropharynx of intubated patients: A randomised controlled open pilot study. Crit. Care 2008, 12, R136. [Google Scholar] [CrossRef]
- Cebeci, A.; Gurakan, C. Properties of potential probiotic Lactobacillus plantarum strains. Food Microbiol. 2003, 20, 511–518. [Google Scholar] [CrossRef]
- de Vries, M.C.; Vaughan, E.E.; Kleerebezem, M.; de Vos, W.M. Lactobacillus plantarum—Survival, functional and potential probiotic properties in the human intestinal tract. Int. Dairy J. 2006, 16, 1018–1028. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Panigrahi, P.; Parida, S.; Nanda, N.C.; Satpathy, R.; Pradhan, L.; Chandel, D.S.; Baccaglini, L.; Mohapatra, A.; Mohapatra, S.S.; Misra, P.R.; et al. A randomized synbiotic trial to prevent sepsis among infants in rural India. Nature 2017, 548, 407–412. [Google Scholar] [CrossRef]
- Boyle, R.J.; Robins-Browne, R.M.; Tang, M.L.K. Probiotic use in clinical practice: What are the risks? Am. J. Clin. Nutr. 2006, 83, 1256–1264. [Google Scholar] [CrossRef]
- Spiller, R. Review article: Probiotics and prebiotics in irritable bowel syndrome. Aliment. Pharmacol. Ther. 2008, 28, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Shoenfeld, Y.; Matthias, T. Probiotics: If it does not help it does not do any harm. Really? Microorganisms 2019, 7, 104. [Google Scholar] [CrossRef] [PubMed]
- Klingberg, T.D.; Axelsson, L.; Naterstad, K.; Elsser, D.; Budde, B.B. Identification of potential probiotic starter cultures for Scandinavian-type fermented sausages. Int. J. Food Microbiol. 2005, 105, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Klingberg, T.D.; Pedersen, M.H.; Cencic, A.; Budde, B.B. Application of measurements of transepithelial electrical resistance of intestinal epithelial cell monolayers to evaluate probiotic activity. Appl. Environ. Microbiol. 2005, 71, 7528–7530. [Google Scholar] [CrossRef] [PubMed]
- Klingberg, T.D.; Budde, B.B. The survival and persistence in the human gastrointestinal tract of five potential probiotic lactobacilli consumed as freeze-dried cultures or as probiotic sausage. Int. J. Food Microbiol. 2006, 109, 157–159. [Google Scholar] [CrossRef]
- Farup, P.G.; Jacobsen, M.; Ligaarden, S.C.; Rudi, K. Probiotics, symptoms, and gut microbiota: What are the relations? A randomized controlled trial in subjects with irritable bowel syndrome. Gastroenterol. Res. Pract. 2012, 2012, 214102. [Google Scholar] [CrossRef]
- Ligaarden, S.; Axelsson, L.; Naterstad, K.; Lydersen, S.; Farup, P. A candidate probiotic with unfavourable effects in subjects with irritable bowel syndrome: A randomised controlled trial. BMC Gastroenterol. 2010, 10, 16. [Google Scholar] [CrossRef]
- Cui, Y.; Hu, T.; Qu, X.; Zhang, L.; Ding, Z.; Dong, A. Plasmids from food lactic acid bacteria: Diversity, similarity, and new developments. Int. J. Mol. Sci. 2015, 16, 13172–13202. [Google Scholar] [CrossRef]
- Arredondo-Alonso, S.; Willems, R.J.; van Schaik, W.; Schürch, A.C. On the (im)possibility of reconstructing plasmids from whole-genome short-read sequencing data. Microb. Genom. 2017, 3, e000128. [Google Scholar] [CrossRef]
- Koren, S.; Phillippy, A.M. One chromosome, one contig: Complete microbial genomes from long-read sequencing and assembly. Curr. Opin. Microbiol. 2015, 23, 110–120. [Google Scholar] [CrossRef]
- George, S.; Pankhurst, L.; Hubbard, A.; Votintseva, A.; Stoesser, N.; Sheppard, A.E.; Mathers, A.; Norris, R.; Navickaite, I.; Eaton, C.; et al. Resolving plasmid structures in Enterobacteriaceae using the MinION nanopore sequencer: Assessment of MinION and MinION/Illumina hybrid data assembly approaches. Microb. Genom. 2017, 3, e000118. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with multiplex MinION sequencing. Microb. Genom. 2017, 3, e000132. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Olsen, H.E.; Paten, B.; Akeson, M. The Oxford Nanopore MinION: Delivery of nanopore sequencing to the genomics community. Genome Biol. 2016, 17, 239. [Google Scholar] [CrossRef] [PubMed]
- Aakra, A.; Nyquist, O.L.; Snipen, L.; Reiersen, T.S.; Nes, I.F. Survey of genomic diversity among Enterococcus faecalis strains by microarray-based comparative genomic hybridization. Appl. Environ. Microbiol. 2007, 73, 2207–2217. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comp. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucl. Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Darling, A.E.; Mau, B.; Perna, N.T. progressiveMauve: Multiple Genome Alignment with Gene Gain, Loss and Rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9. [Google Scholar] [CrossRef]
- Conlan, S.; Thomas, P.J.; Deming, C.; Park, M.; Lau, A.F.; Dekker, J.P.; Snitkin, E.S.; Clark, T.A.; Luong, K.; Song, Y.; et al. Single-molecule sequencing to track plasmid diversity of hospital-associated carbapenemase-producing Enterobacteriaceae. Sci. Transl. Med. 2014, 6, 254ra126. [Google Scholar] [CrossRef]
- Crowley, S.; Bottacini, F.; Mahony, J.; van Sinderen, D. Complete genome sequence of Lactobacillus plantarum strain 16, a broad-spectrum antifungal-producing lactic acid bacterium. Genome Announc. 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Ravenhall, M.; Škunca, N.; Lassalle, F.; Dessimoz, C. Inferring horizontal gene transfer. PLoS Comp. Biol. 2015, 11, e1004095. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, M.; Boekhorst, J.; van Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M.; Fiers, M.W.E.J.; et al. Complete genome sequence of Lactobacillus plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar] [CrossRef] [PubMed]
- Siezen, R.J.; Francke, C.; Renckens, B.; Boekhorst, J.; Wels, M.; Kleerebezem, M.; van Hijum, S.A. Complete resequencing and reannotation of the Lactobacillus plantarum WCFS1 genome. J. Bacteriol. 2012, 194, 195–196. [Google Scholar] [CrossRef] [PubMed]
- Petkau, K.; Fast, D.; Duggal, A.; Foley, E. Comparative evaluation of the genomes of three common Drosophila-associated bacteria. Biol. Open 2016, 5, 1305. [Google Scholar] [CrossRef]
- NCBI. Organism Overview; Lactobacillus plantarum—Common Gastrointestinal Bacterium Used in Food Production. Available online: https://www.ncbi.nlm.nih.gov/genome/?term=lactobacillus+plantarum+wcfs1 (accessed on 25 May 2019).
- Cho, G.S.; Huch, M.; Mathara, J.M.; van Belkum, M.J.; Franz, C.M.A.P. Characterization of pMRI 5.2, a rolling-circle-type plasmid from Lactobacillus plantarum BFE 5092 which harbours two different replication initiation genes. Plasmid 2013, 69, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Danielsen, M. Characterization of the tetracycline resistance plasmid pMD5057 from Lactobacillus plantarum 5057 reveals a composite structure. Plasmid 2002, 48, 98–103. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Banerjei, L.; Myers, G.S.A.; Nelson, K.E.; Seshadri, R.; Read, T.D.; Fouts, D.E.; Eisen, J.A.; Gill, S.R.; Heidelberg, J.F.; et al. Role of mobile DNA in the evolution of vancomycin-resistant Enterococcus faecalis. Science 2003, 299, 2071–2074. [Google Scholar] [CrossRef]
- Gray, M.J.; Escalante-Semerena, J.C. A new pathway for the synthesis of α-ribazole-phosphate in Listeria innocua. Mol. Microbiol. 2010, 77, 1429–1438. [Google Scholar] [CrossRef]
- Rodionov, D.A.; Hebbeln, P.; Eudes, A.; ter Beek, J.; Rodionova, I.A.; Erkens, G.B.; Slotboom, D.J.; Gelfand, M.S.; Osterman, A.L.; Hanson, A.D.; et al. A novel class of modular transporters for vitamins in prokaryotes. J. Bacteriol. 2009, 191, 42–51. [Google Scholar] [CrossRef]
- Toro, N.; Nisa-Martínez, R. Comprehensive phylogenetic analysis of bacterial reverse transcriptases. PLoS ONE 2014, 9, e114083. [Google Scholar] [CrossRef] [PubMed]
- van Kranenburg, R.; Golic, N.; Bongers, R.; Leer, R.J.; de Vos, W.M.; Siezen, R.J.; Kleerebezem, M. Functional analysis of three plasmids from Lactobacillus plantarum. Appl. Environ. Microbiol. 2005, 71, 1223–1230. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| GenBank Assembly * | GenBank No. | Type | Name | Topology | Size (bp) | GC% | Protein | rRNA | tRNA | Other RNA | Gene | Pseudo Gene |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GCA_001880185.1 and GCA_001880185.2 | CP013149.1 | Chromosome | MF1298 | Circular | 3235952 | 44.6 | 2957 | 16 | 69 | 4 | 3118 | 72 |
| CP013150.1 | Plasmid | pMF1298-1 | Circular | 63114 | 41.1 | 60 | - | - | - | 71 | 11 | |
| CP013151.1 | Plasmid | pMF1298-2 | Circular | 55699 | 40.1 | 50 | - | - | - | 58 | 8 | |
| GCA_001880185.2 | CP013153.2 | Plasmid | pMF1298-3 | Circular | 47476 | 40.7 | 40 | - | - | - | 47 | 7 |
| CP013152.2 | Plasmid | pMF1298-4 | Circular | 45064 | 40.8 | 35 | - | - | - | 49 | 14 | |
| CP013155.2 | Plasmid | pMF1298-5 | Circular | 37426 | 40.7 | 33 | - | - | - | 38 | 5 | |
| CP013156.2 | Plasmid | pMF1298-6 | Circular | 30526 | 45.8 | 29 | - | - | - | 35 | 6 | |
| CP013154.2 | Plasmid | pMF1298-7 | Circular | 23493 | 39.6 | 21 | - | - | - | 29 | 8 | |
| CP013158.2 | Plasmid | pMF1298-8 | Circular | 10848 | 36.5 | 14 | - | - | - | 17 | 3 | |
| CP013160.2 | Plasmid | pMF1298-9 | Circular | 8511 | 34.2 | 10 | - | - | - | 10 | - | |
| CP013166.2 | Plasmid | pMF1298-10 | Circular | 5636 | 36.3 | 9 | - | - | - | 11 | 2 | |
| CP013162.2 | Plasmid | pMF1298-11 | Circular | 4130 | 37.7 | 4 | - | - | - | 4 | - | |
| CP013167.2 | Plasmid | pMF1298-12 | Circular | 3350 | 38.4 | 4 | - | - | - | 4 | - | |
| CP013168.2 | Plasmid | pMF1298-13 | Circular | 2942 | 35.8 | 3 | - | - | - | 5 | 2 | |
| CP013170.2 | Plasmid | pMF1298-14 | Circular | 2273 | 36.6 | 3 | - | - | - | 3 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McLeod, A.; Fagerlund, A.; Rud, I.; Axelsson, L. Large Plasmid Complement Resolved: Complete Genome Sequencing of Lactobacillus plantarum MF1298, a Candidate Probiotic Strain Associated with Unfavorable Effect. Microorganisms 2019, 7, 262. https://doi.org/10.3390/microorganisms7080262
McLeod A, Fagerlund A, Rud I, Axelsson L. Large Plasmid Complement Resolved: Complete Genome Sequencing of Lactobacillus plantarum MF1298, a Candidate Probiotic Strain Associated with Unfavorable Effect. Microorganisms. 2019; 7(8):262. https://doi.org/10.3390/microorganisms7080262
Chicago/Turabian StyleMcLeod, Anette, Annette Fagerlund, Ida Rud, and Lars Axelsson. 2019. "Large Plasmid Complement Resolved: Complete Genome Sequencing of Lactobacillus plantarum MF1298, a Candidate Probiotic Strain Associated with Unfavorable Effect" Microorganisms 7, no. 8: 262. https://doi.org/10.3390/microorganisms7080262
APA StyleMcLeod, A., Fagerlund, A., Rud, I., & Axelsson, L. (2019). Large Plasmid Complement Resolved: Complete Genome Sequencing of Lactobacillus plantarum MF1298, a Candidate Probiotic Strain Associated with Unfavorable Effect. Microorganisms, 7(8), 262. https://doi.org/10.3390/microorganisms7080262