Development and Validation of Shiga Toxin-Producing Escherichia coli Immunodiagnostic Assay



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Bacterial Supernatant Preparation
2.3. Antibodies
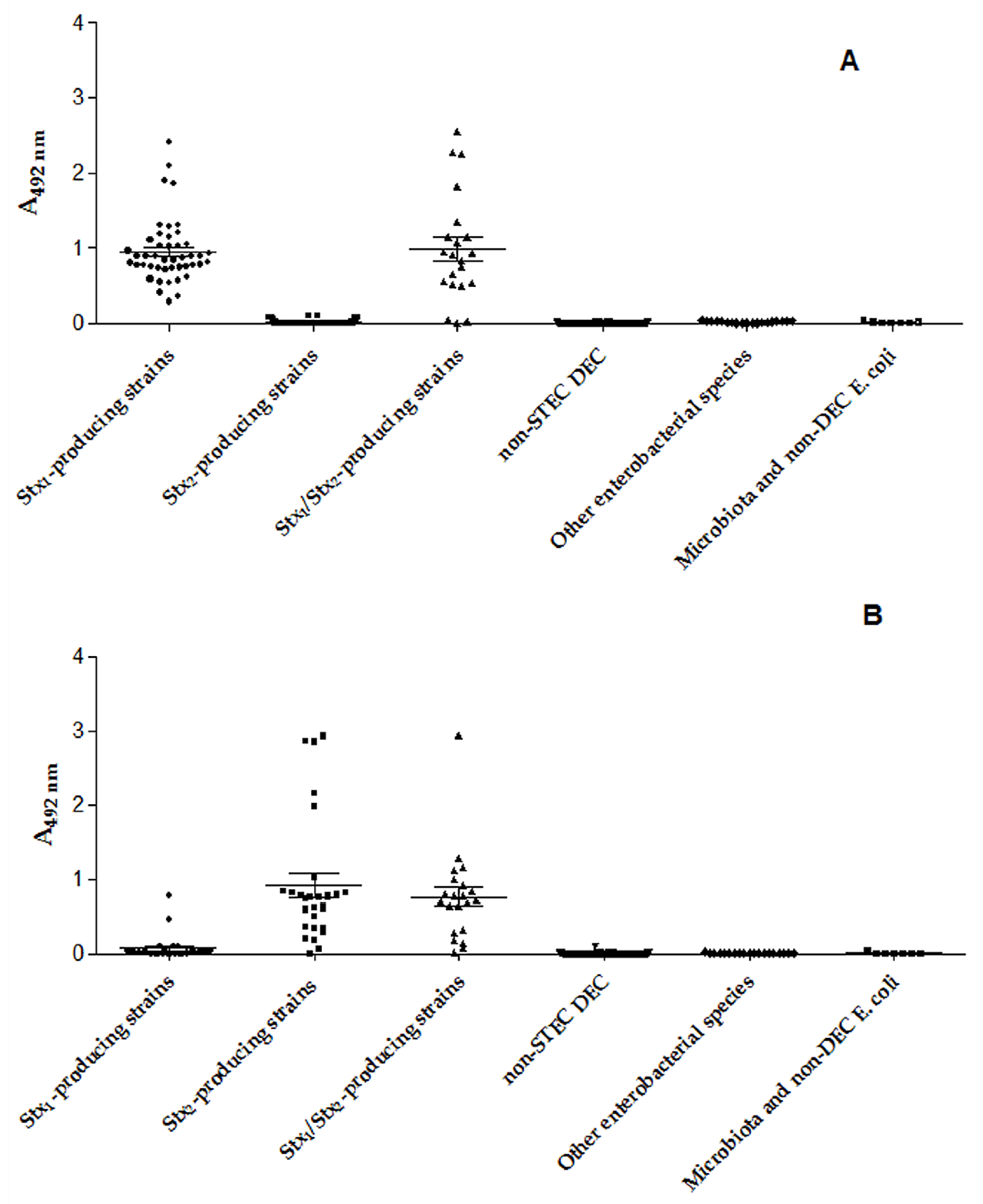
2.4. Capture ELISA Immunoassay (cEIA)
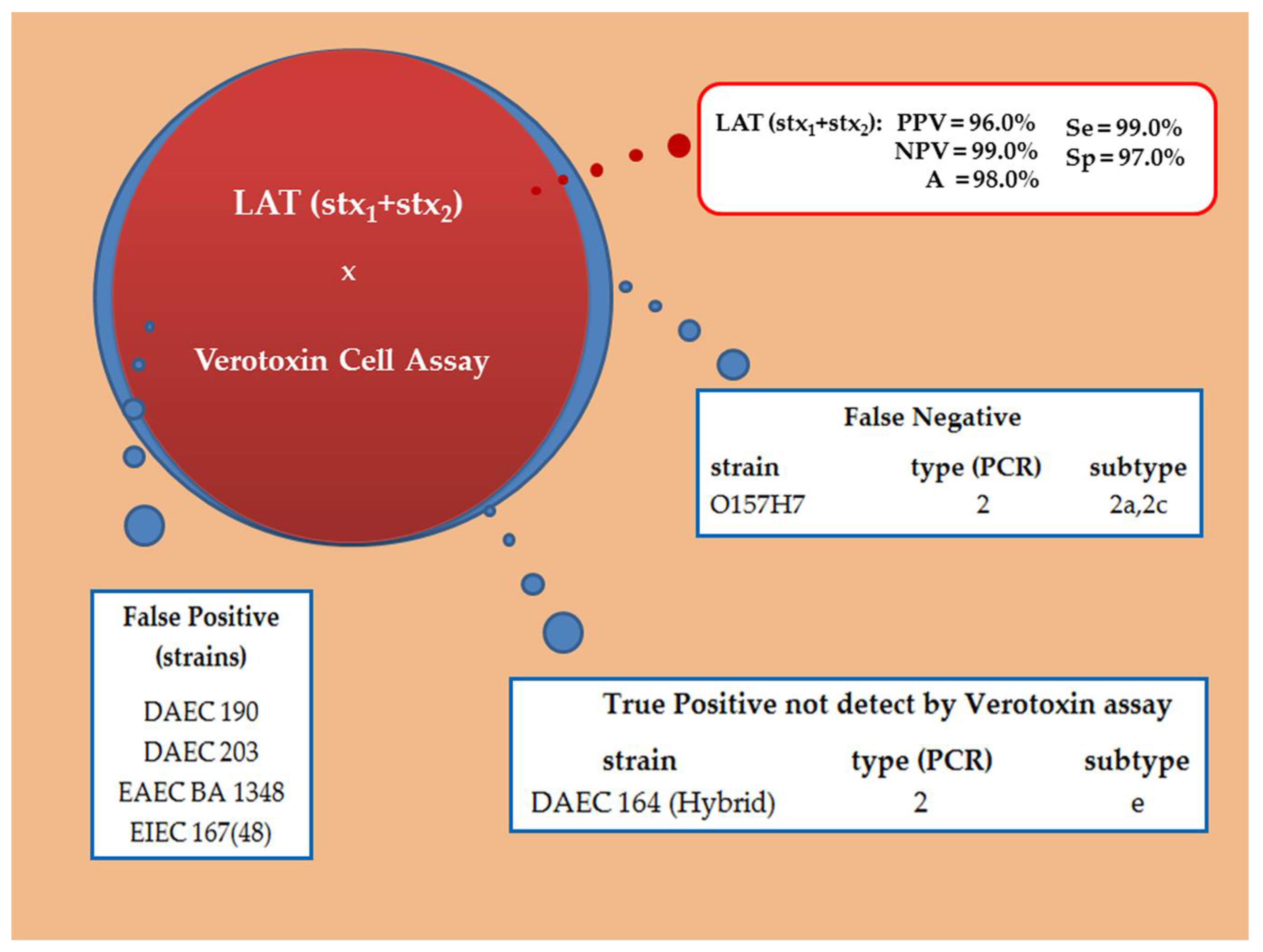
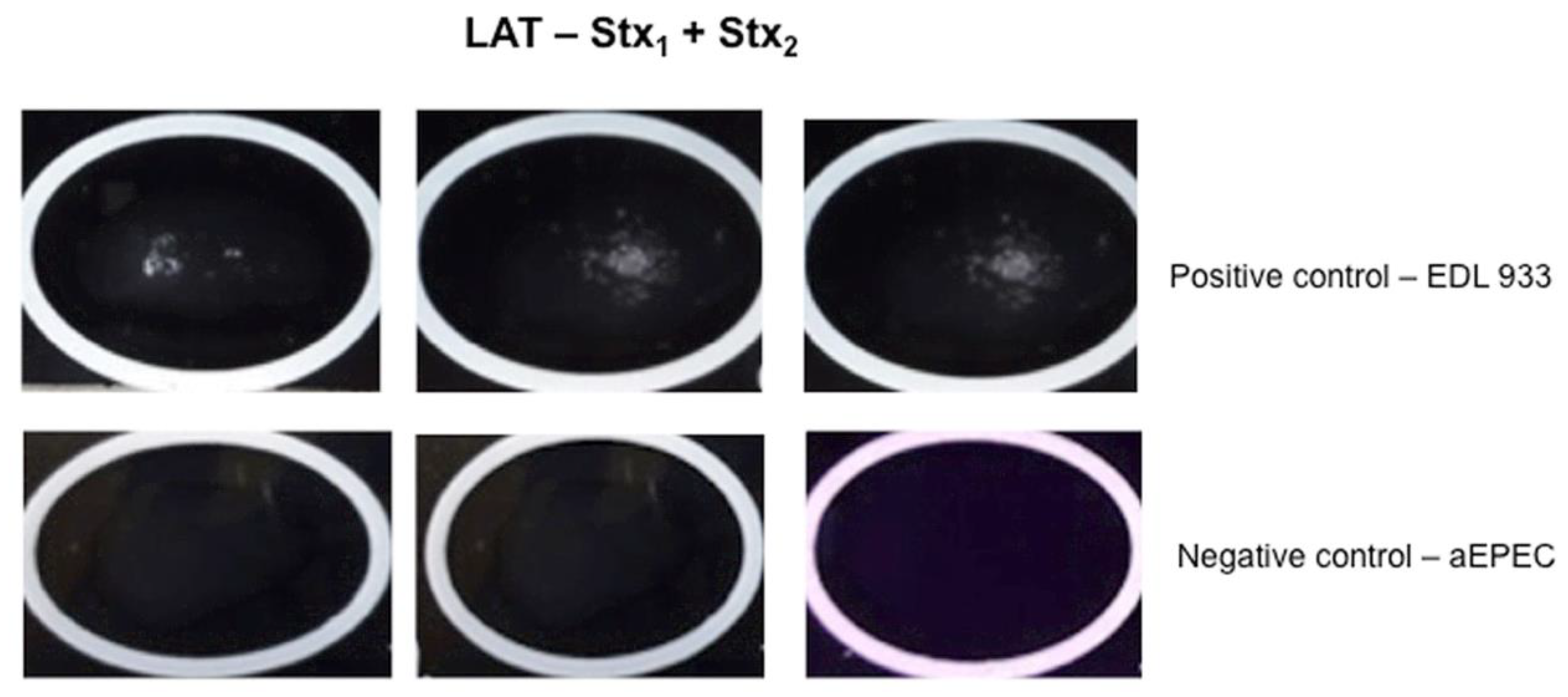
2.5. Latex Agglutination Test (LAT)
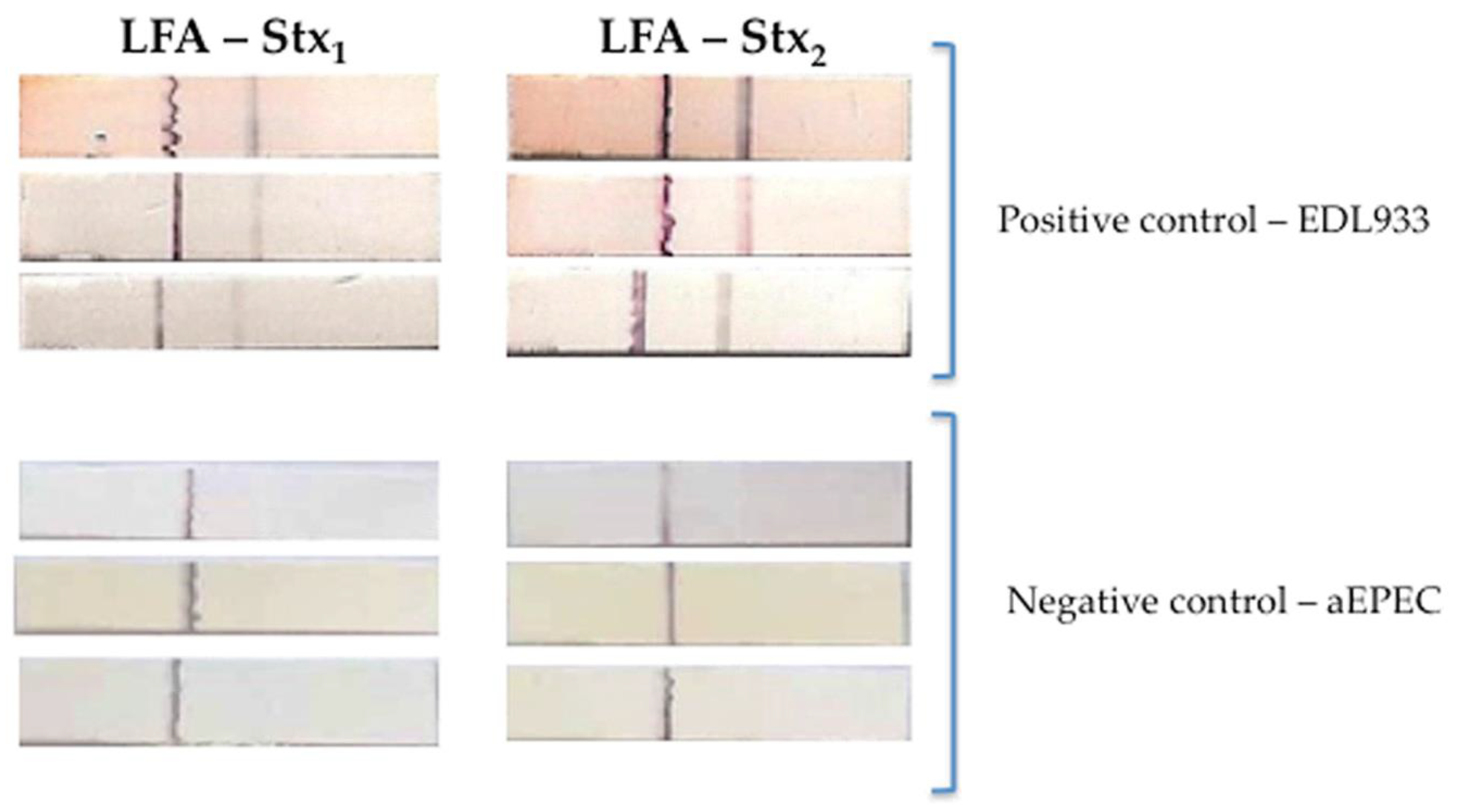
2.6. Lateral Flow Assay (LFA)
2.7. Stx Subtyping
2.8. Statistical Analyses
3. Results
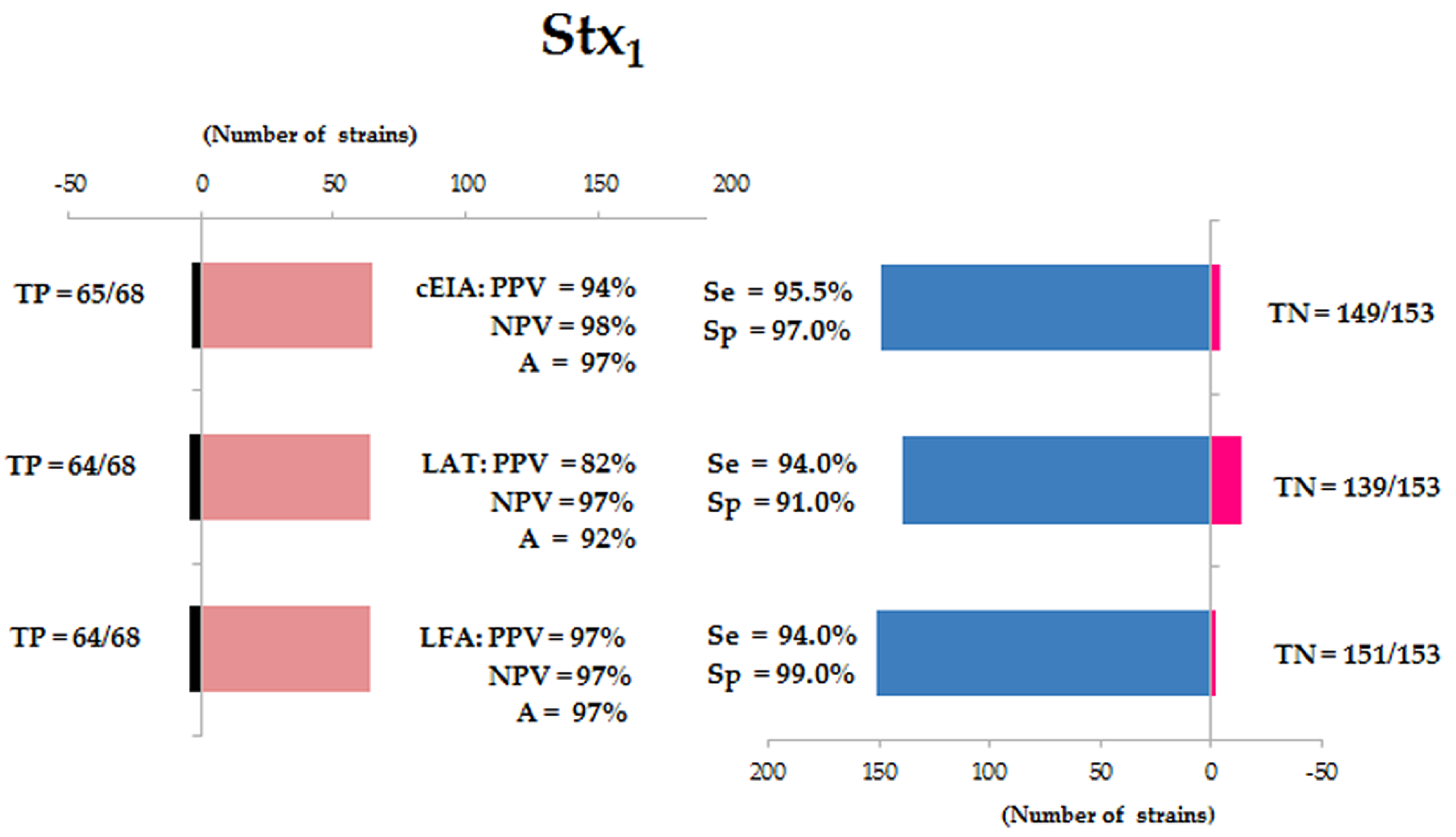
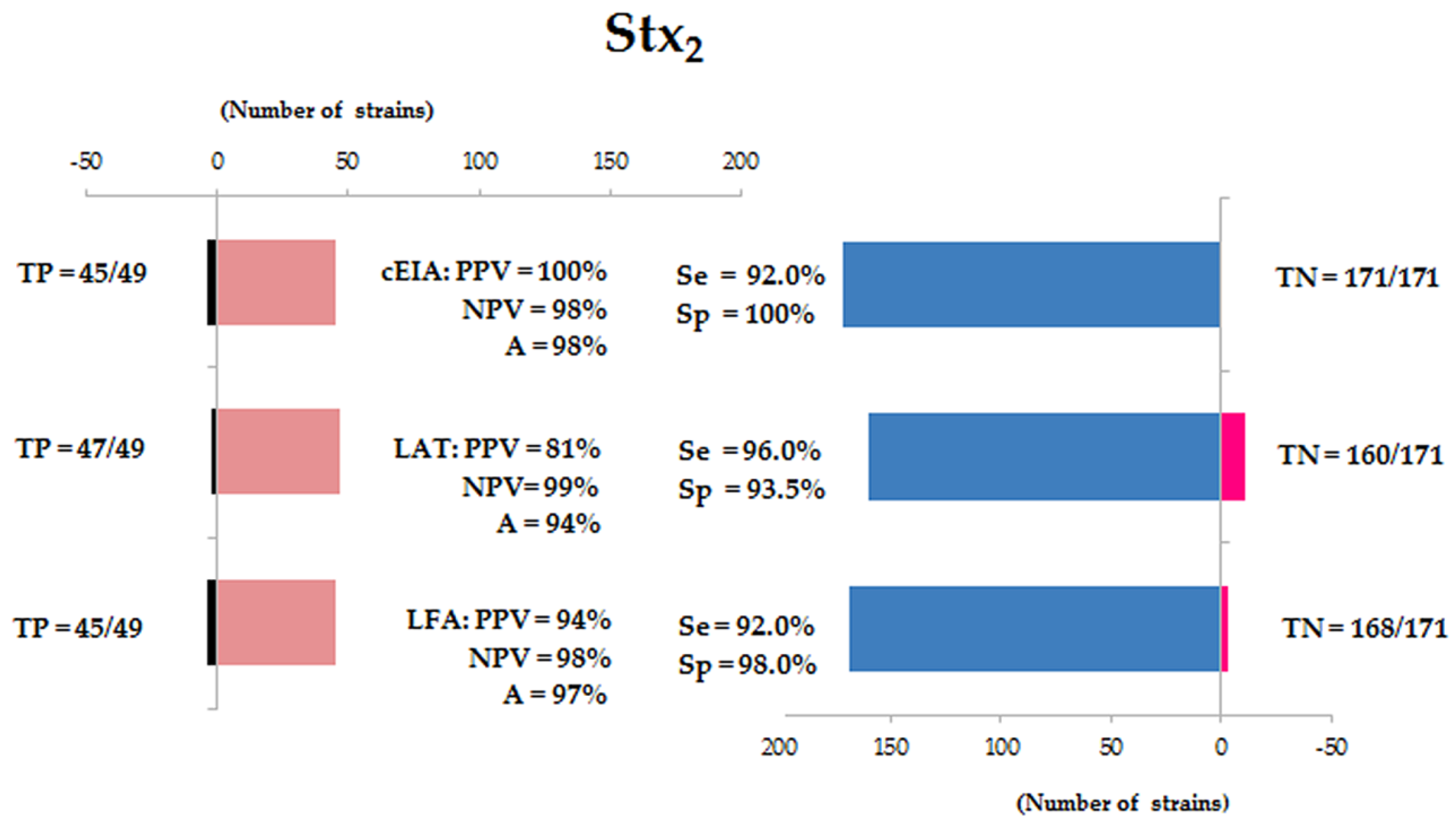
3.1. Presence and Production of Stx1 and Stx2
3.2. Validation of Diagnostic Immunoassays
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Karmali, M.A.; Gannon, V.; Sargeant, J.M. Verocytotoxin-producing Escherichia coli (VTEC). Vet. Microbiol. 2010, 140, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Riley, L.W.; Remis, R.S.; Helgerson, S.D.; McGee, H.B.; Wells, J.G.; Davis, B.R.; Hebert, R.J.; Olcott, E.S.; Johnson, L.M.; Hargrett, N.T.; et al. Hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med. 1983, 308, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Karmali, M.A.; Steele, B.T.; Petric, M.; Lim, C. Sporadic cases of haemolytic-uraemic syndrome associated with faecal cytotoxin and cytotoxin-producing Escherichia coli in stools. Lancet 1983, 1, 619–620. [Google Scholar] [CrossRef]
- Karmali, M.A.; Petric, M.; Lim, C.; Fleming, P.C.; Arbus, G.S.; Lior, H. The association between idiopathic hemolytic uremic syndrome and infection by verotoxin-producing Escherichia coli. J. Infect. Dis. 1985, 151, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Byrne, L.; Jenkins, C.; Launders, N.; Elson, R.; Adak, G.K. The epidemiology, microbiology and clinical impact of Shiga toxin-producing Escherichia coli in England, 2009–2012. Epidemiol. Infect. 2015, 143, 3475–3487. [Google Scholar] [CrossRef] [PubMed]
- Melton-Celsa, A.R. Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Heiman, K.E.; Mody, R.K.; Johnson, S.D.; Griffin, P.M.; Gould, L.H. Escherichia coli O157 outbreaks in the United States, 2003–2012. Emerg. Infect. Dis. 2015, 21, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Gordon, C.A.; Chandler, W.L. Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 2005, 365, 1073–1086. [Google Scholar] [CrossRef]
- Byrne, L.; Vanstone, G.L.; Perry, N.T.; Launders, N.; Adak, G.K.; Godbole, G.; Grant, K.A.; Smith, R.; Jenkins, C. Epidemiology and microbiology of Shiga toxin-producing Escherichia coli other than serogroup O157 in England, 2009–2013. J. Med. Microbiol. 2014, 63, 1181–1188. [Google Scholar] [CrossRef]
- Kuehne, A.; Bouwknegt, M.; Havelaar, A.; Gilsdorf, A.; Hoyer, P.; Stark, K.; Werber, D. HUS active surveillance network Germany. Estimating true incidence of O157 and non-O157 Shiga toxin-producing Escherichia coli illness in Germany based on notification data of haemolytic uraemic syndrome. Epidemiol. Infect. 2016, 114, 3305–3315. [Google Scholar] [CrossRef]
- Rivas, M.; Padola, N.L.; Lucchesi, P.M.A.; Masana, M. Diarrheogenic Escherichia coli in Argentina. In Pathogenic Escherichia coli in Latin America; Torres, A.G., Ed.; Bentham Books: Potomac, MD, USA, 2010; Volume 1, pp. 142–161. [Google Scholar]
- Vidal, R.M.; Oñate, A.; Salazar, J.C.; Prado, V. Shiga toxin producing Escherichia coli in Chile. In Pathogenic Escherichia coli in Latin America; Torres, A.G., Ed.; Bentham Books: Potomac, MD, USA, 2010; Volume 1, pp. 179–190. [Google Scholar]
- Guth, B.E.C.; Picheth, C.; Gomes, T.A.T. Escherichia coli situation in Brazil. In Pathogenic Escherichia coli in Latin America; Torres, A.G., Ed.; Bentham Books: Potomac, MD, USA, 2010; Volume 1, pp. 162–178. [Google Scholar]
- Varela, G.; Gómez-Garcia, O.; Ochoa, T. Diarrheogenic Escherichia coli in children from Uruguay, Colombia and Peru. In Pathogenic Escherichia coli in Latin America; Torres, A.G., Ed.; Bentham Books: Potomac, MD, USA, 2010; Volume 1, pp. 209–222. [Google Scholar]
- Navarro, A.; Estrada-Garcia, T. Epidemiology of diarrheogenic Escherichia coli Pathotypes in Mexico, past and present. In Pathogenic Escherichia coli in Latin America; Torres, A.G., Ed.; Bentham Books: Potomac, MD, USA, 2010; Volume 1, pp. 191–208. [Google Scholar]
- Rivas, M.; Chinen, I.; Guth, B.E.C. Enterohemorrhagic (Shiga Toxin-Producing) Escherichia coli. In Escherichia coli in the Americas; Torres, A.G., Ed.; Springer Nature: Basel, Switzerland, 2016; Volume 1, pp. 97–123. [Google Scholar]
- Torres, A.G.; Amaral, M.M.; Betancor, L.; Galli, L.; Goldstein, J.; Krüger, A.; Rojas-López, M. Recent Advances in Shiga Toxin-Producing Escherichia coli Research in Latin America. Microorganisms 2018, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.G.; Cerqueira, A.M.; Guth, B.E.; Coutinho, C.A.; Liberal, M.H.; Souza, R.M.; Andrade, J.R. Serotypes, virulence markers and cell invasion ability of Shiga toxin-producing Escherichia coli (STEC) strains isolated from healthy dairy cattle. J. Appl. Microbiol. 2016, 121, 1130–1143. [Google Scholar] [CrossRef] [PubMed]
- Lascowski, K.M.S.; Guth, B.E.C.; Martins, F.H.; Rocha, S.P.D.; Irino, K.; Pelayo, J.S. Shiga toxin-producing Escherichia coli in drinking water supplies of North Paraná State, Brazil. J. Appl. Microbiol. 2013, 114, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Beraldo, L.G.; Borges, C.A.; Maluta, R.P.; Cardozo, M.V.; Rigobelo, E.C.; Ávila, F.A. Detection of Shiga toxigenic (STEC) and enteropathogenic (EPEC) Escherichia coli in dairy buffalo. Vet. Microbiol. 2014, 170, 162–166. [Google Scholar] [CrossRef] [PubMed]
- Maluta, R.P.; Fairbrother, J.M.; Stella, A.E.; Rigobelo, E.C.; Martinez, R.; Ávila, F.A. Potentially pathogenic Escherichia coli in healthy, pasture-raised sheep on farms and at the abattoir in Brazil. Vet. Microbiol. 2014, 169, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.H.; Guth, B.E.; Piazza, R.M.; Leão, S.C.; Ludovico, A.; Ludovico, M.S.; Dahbi, G.; Marzoa, J.; Mora, A.; Blanco, J.; et al. Diversity of Shiga toxin-producing Escherichia coli in sheep flocks of Paraná State, southern Brazil. Vet. Microbiol. 2015, 175, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Borges, C.A.; Beraldo, L.G.; Maluta, R.P.; Cardozo, M.V.; Guth, B.E.; Rigobelo, E.C.; Ávila, F.A. Shiga toxigenic and atypical enteropathogenic Escherichia coli in the feces and carcasses of slaughtered pigs. Food Path. Dis. 2012, 10, 1–7. [Google Scholar]
- Martins, R.P.; Silva, M.C.; Dutra, V.; Nakazato, L.; Leite, D.S. Preliminary virulence genotyping and phylogeny of Escherichia coli from the gut of pigs at slaughtering stage in Brazil. Meat Sci. 2013, 93, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Gioia-Di Chiacchio, R.M.C.; Cunha, M.P.; Sturn, R.M.; Moreno, L.Z.; Moreno, A.M.; Pereira, C.B.; Martins, F.H.; Franzolin, M.R.; Piazza, R.M.; Knöbl, T. Shiga toxin-producing Escherichia coli (STEC): Zoonotic risks associated with psittacine pet birds in home environments. Vet. Microbiol. 2016, 184, 27–30. [Google Scholar] [CrossRef]
- Gioia-Di Chiacchio, R.M.; Cunha, M.P.V.; Sá, L.R.M.; Davies, Y.M.; Pereira, C.B.P.; Martins, F.H.; Munhoz, D.D.; Abe, C.M.; Franzolin, M.R.; Santos, L.F.; et al. Novel Hybrid of Typical Enteropathogenic Escherichia coli and Shiga-Toxin-3 E. coli (tEPEC/STEC) Emerging From Pet Birds. Front. Microbiol. 2018, 9, 1–10. [Google Scholar] [CrossRef]
- Ribeiro, L.F.; Barbosa, M.M.C.; Pinto, F.R. Shiga toxigenic andenteropathogenic Escherichia coli in water and fishfrom pay-to-fish ponds. Lett. Appl. Microbiol. 2015, 62, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.H.; Guth, B.E.; Piazza, R.M.; Blanco, J.; Pelayo, J.S. First description of a Shiga toxin-producing Escherichia coli O103:H2 strain isolated from sheep in Brazil. J. Infect. Dev. Ctries. 2014, 8, 126–128. [Google Scholar] [CrossRef]
- Clements, A.; Young, J.C.; Constantinou, N.; Frankel, G. Infection strategies of enteric pathogenic Escherichia coli. Gut Microbes 2012, 3, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Bettelheim, K.A.; Beutin, L. Rapid laboratory identification and characterization of verocytotoxigenic (Shiga toxin producing) Escherichia coli (VTEC/STEC). J. Appl. Microbiol. 2003, 95, 205–217. [Google Scholar] [CrossRef]
- Paton, J.C.; Paton, A.W. Pathogenesis and diagnosis of Shiga toxin-producing Escherichia coli infections. Clin. Microbiol. Rev. 1998, 11, 450–479. [Google Scholar] [CrossRef]
- Gould, L.H.; Bopp, C.; Strockbine, N.; Atkinson, R.; Baselski, V.; Body, B.; Carey, R.; Crandall, C.; Hurd, S.; Kaplan, R.; et al. Centers for Disease Control and Prevention. Recommendations for Diagnosis of Shiga Toxin. Morb. Mortal. Wkly. Rep. 2009, 58, 1–14. [Google Scholar]
- Rocha, L.B.; Piazza, R.M.F. Production of Shiga toxin by Shiga toxin-expressing Escherichia coli (STEC) in broth media: From divergence to definition. Lett. Appl. Microbiol. 2007, 45, 411–417. [Google Scholar] [CrossRef]
- Mendes-Ledesma, M.R.; Rocha, L.B.; Bueris, V.; Krause, G.; Beutin, L.; Franzolin, M.R.; Trabulsi, L.R.; Elias, W.P.; Piazza, R.M. Production and characterization of rabbit polyclonal sera against Shiga toxins Stx1 and Stx2 for detection of Shiga toxin-producing Escherichia coli. Microbiol. Immunol. 2008, 52, 484–491. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.B.; Luz, D.E.; Moraes, C.T.; Caravelli, A.; Fernandes, I.; Guth, B.E.; Horton, D.S.; Piazza, R.M. Interaction between Shiga toxin and monoclonal antibodies: Binding characteristics and in vitro neutralizing abilities. Toxins 2012, 4, 729–747. [Google Scholar] [CrossRef]
- Nara, J.M.; Cianciarullo, A.M.; Culler, H.F.; Bueris, V.; Horton, D.S.; Menezes, M.A.; Franzolin, M.R.; Elias, W.P.; Piazza, R.M. Differentiation of typical and atypical enteropathogenic Escherichia coli using colony immunoblot for detection of bundle-forming pilus expression. J. Appl. Microbiol. 2010, 109, 35–43. [Google Scholar]
- Menezes, M.A.; Rocha, L.B.; Koga, P.C.; Fernandes, I.; Nara, J.M.; Magalhães, C.A.; Abe, C.M.; Ayala, C.O.; Burgos, Y.K.; Elias, W.P.; et al. Identification of enteropathogenic and enterohaemorrhagic Escherichia coli strains by immunoserological detection of intimin. J. Appl. Microbiol. 2010, 108, 878–887. [Google Scholar] [CrossRef] [PubMed]
- Abe, C.M.; Trabulsi, L.R.; Blanco, J.; Blanco, M.; Dahbi, G.; Blanco, J.E.; Mora, A.; Franzolin, M.R.; Taddei, C.R.; Martinez, M.B.; et al. Virulence features of atypical enteropathogenic Escherichia coli identified by the eae(+) EAF-negative stx(-) genetic profile. Diagn. Microbiol. Infect. Dis. 2009, 64, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.B.; Ozaki, C.Y.; Horton, D.S.; Menezes, C.A.; Silva, A.; Fernandes, I.; Magnoli, F.C.; Vaz, T.M.; Guth, B.E.; Piazza, R.M. Different assay conditions for detecting the production and release of heat-labile and heat-stable toxins in enterotoxigenic Escherichia coli isolates. Toxins 2013, 5, 2384–2402. [Google Scholar] [CrossRef]
- Ozaki, C.Y.; Silveira, C.R.; Andrade, F.B.; Nepomuceno, R.; Silva, A.; Munhoz, D.D.; Yamamoto, B.B.; Luz, D.; Abreu, P.A.; Horton, D.S.; et al. Single Chain Variable Fragments Produced in Escherichia coli against Heat-Labile and Heat-Stable Toxins from Enterotoxigenic, E. coli. PLoS ONE 2015, 10, 1–14. [Google Scholar] [CrossRef]
- Bueris, V.; Sircili, M.P.; Taddei, C.R.; Santos, M.F.; Franzolin, M.R.; Martinez, M.B.; Ferrer, S.R.; Barreto, M.L.; Trabulsi, L.R. Detection of diarrheagenic Escherichia coli from children with and without diarrhea in Salvador, Bahia, Brazil. Mem Inst. Oswaldo Cruz 2007, 102, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Taddei, C.R.; Moreno, A.C.; Fernandes Filho, A.; Montemor, L.P.; Martinez, M.B. Prevalence of secreted autotransporter toxin gene among diffusely adhering Escherichia coli isolated from stools of children. FEMS Microbiol. Lett. 2003, 227, 249–253. [Google Scholar] [CrossRef]
- Perna, N.T.; Plunkett, G., 3rd; Burland, V.; Mau, B.; Glasner, J.D.; Rose, D.J.; Mayhew, G.F.; Evans, P.S.; Gregor, J.; Kirkpatrick, H.A.; et al. Genome sequence of enterohaemorrhagic Escherichia coli O157:H7. Nature 2001, 409, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, R.K.; Indrayan, A. A simple nomogram for sample size for estimating sensitivity and specificity of medical tests. Indian J. Ophthalmol. 2010, 58, 519–522. [Google Scholar] [CrossRef]
- Hajian-Tilaki, K. Sample size estimation in diagnostic test studies of biomedical informatics. J. Biomed. Inform. 2014, 49, 193–204. [Google Scholar] [CrossRef]
- Lanata, C.F.; Fischer-Walker, C.L.; Olascoaga, A.C.; Torres, C.X.; Aryee, M.J.; Black, R.E.; Child Health Epidemiology Reference Group of the World Health Organization and UNICEF. Global causes of diarrheal disease mortality in children <5 years of age: A systematic review. PLoS ONE 2013, 8, e72788. [Google Scholar] [CrossRef]
- Ori, E.L.; Takagi, E.H.; Andrade, T.S.; Miguel, B.T.; Cergole-Novella, M.C.; Guth, B.E.C.; Hernandes, R.T.; Dias, R.C.B.; Pinheiro, S.R.S.; Camargo, C.H.; et al. Diarrhoeagenic Escherichia coli and Escherichia albertii in Brazil: Pathotypes and serotypes over a 6-year period of surveillance. Epidemiol. Infect. 2018, 19, 1–9. [Google Scholar] [CrossRef]
- Rocha, L.B.; Santos, A.R.; Munhoz, D.D.; Cardoso, L.T.; Luz, D.E.; Andrade, F.B.; Horton, D.S.; Elias, W.P.; Piazza, R.M. Development of a rapid agglutination latex test for diagnosis of enteropathogenic and enterohemorrhagic Escherichia coli infection in developing world: Defining the biomarker, antibody and method. PLoS Negl. Trop. Dis. 2014, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Oliver, C. Immunocytochemical methods and protocols. In Methods in Molecular Biology, 3rd ed.; Walker, J.M., Ed.; Humana Press: New York, NY, USA, 2010; Volume 588, pp. 363–387. [Google Scholar]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [PubMed]
- Konowalchuk, J.; Speirs, J.I.; Stravic, S. Vero response to a cytotoxin of Escherichia coli. Infect. Immun. 1977, 18, 775–779. [Google Scholar] [PubMed]
- Cohen, J.A. Coefficient of agreement for nominal scales. Ed. Psych. Meas 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Piazza, R.M.F.; Abe, C.M.; Horton, D.S.P.Q.; Miliwebsky, E.; Chinen, I.; Vaz, T.M.I.; Irino, K. Detection and subtyping methods of diarrheogenic Escherichia coli strains. In Pathogenic Escherichia coli in Latin America; Torres, A.G., Ed.; Bentham Books: Potomac, MD, USA, 2010; Volume 1, pp. 95–115. [Google Scholar]
- Miliwebsky, E.; Schelotto, F.; Varela, G.; Luz, D.; Chinen, I.; Piazza, R.M.F. Human diarrheal infections: Diarrheagenic Escherichia coli pathotypes diagnosis. In Escherichia coli in the Americas; Torres, A.G., Ed.; Springer Nature: Basel, Switzerland, 2016; Volume 1, pp. 343–369. [Google Scholar]
- Gomes, T.A.; Elias, W.P.; Scaletsky, I.C.; Guth, B.E.; Rodrigues, J.F.; Piazza, R.M.; Ferreira, L.C.; Martinez, M.B. Diarrheagenic Escherichia coli. Braz. J. Microbiol. 2016, 47, 3–30. [Google Scholar]
- De Mena, M.F.; Tous, M.; Miliwebsky, E.; Chillemi, G.; Rivas, M. Differential kinetic patterns for Shiga toxin production by Escherichia coli. Rev. Argent. Microbiol. 1997, 29, 167–175. [Google Scholar]
- Kolling, G.L.; Matthews, K.R. Export of virulence genes and Shiga toxin by membrane vesicles of Escherichia coli O157:H7. Appl. Environ. Microbiol. 1999, 65, 1843–1848. [Google Scholar]
- Ching, K.H.; He, X.; Stanker, L.H.; Lin, A.V.; McGarvey, J.A.; Hnasko, R. Detection of Shiga Toxins by Lateral Flow Assay. Toxins 2015, 7, 1163–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Katani, R.; Li, L.; Hegde, N.; Roberts, E.L.; Kapur, V.; DebRoy, C. Rapid Detection of Escherichia coli O157 and Shiga Toxins by Lateral Flow Immunoassays. Toxins 2016, 8, 92. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Serotype | Gene Presence stx1/stx2 | stx Subtype | cEIA (Stx1) | cEIA (Stx2) | LAT (Stx1 + Stx2) | LFA (Stx1) | LFA (Stx2) |
|---|---|---|---|---|---|---|---|---|
| IAL6189 | O24:H4 | 1 | 1a | 1.149 | 0.000 | + | + | − |
| IAL6206 | O24:H4 | 1 | 1a | 0.717 | 0.000 | + | + | + |
| IAL6163 | O26:H11 | 1 | 1a | 0.810 | 0.000 | + | + | − |
| IAL6162 | O26:H11 | 1 | 1a | 0.777 | 0.000 | + | + | − |
| H30 | O26:H11 | 1 | 1a | 1.207 | 0.003 | + | + | − |
| H19 | O26:H11 | 1 | 1a | 1.109 | 0.000 | + | + | − |
| 199 | O26:H11 | 1 | 1a | 0.892 | 0.000 | + | + | − |
| 3529 | O26:H11 | 1 | 1a | 0.772 | 0.000 | + | + | − |
| EPM16 | O26:H11 | 1 | 1a | 1.019 | 0.006 | + | + | − |
| BA4123 | O26:H11 | 1 | 1a | 0.853 | 0.000 | + | + | − |
| D360-4-1 | O26:H11 | 1 | 1a | 0.832 | 0.000 | + | + | − |
| 1557-77 | O26:H11 | 1 | 1a | 2.416 | 0.061 | + | + | − |
| CL5 | O26:H12 | 1 | 1a | 0.760 | 0.000 | + | + | − |
| EPM5 | O55:H19 | 1 | 1a | 0.896 | 0.008 | + | + | − |
| IAL6174 | O71:H8 | 1 | 1a | 0.767 | 0.000 | + | + | − |
| IAL6290 | O76:H19 | 1 | 1c | 0.545 | 0.000 | + | + | − |
| IAL6173 | O91:H14 | 1 | 1a | 0.288 | 0.013 | + | + | − |
| IAL6186 | O103:H− | 1 | 1a | 1.027 | 0.016 | + | + | − |
| IAL6175 | O111:H+ | 1 | 1a | 1.900 | 0.100 | + | + | − |
| IAL6191 | O111:H+ | 1 | 1a | 0.739 | 0.000 | + | + | − |
| IAL6184 | O111:H8 | 1 | 1a | 0.857 | 0.007 | + | + | − |
| IAL6177 | O111:H8 | 1 | 1a | 2.093 | 0.088 | + | + | − |
| IAL6200 | O111:H8 | 1 | 1a | 0.741 | 0.000 | + | + | − |
| EPM20 | O111:H8 | 1 | 1a | 0.539 | 0.000 | + | + | − |
| IAL6183 | O111:H8 | 1 | 1a | 0.893 | 0.000 | + | + | − |
| EPM23 | O111:H8 | 1 | 1a | 0.831 | 0.000 | + | + | − |
| IAL6187 | O111:H11 | 1 | 1a | 0.783 | 0.011 | + | + | − |
| IAL6178 | O111:HNM | 1 | 1a | 0.354 | 0.000 | + | + | − |
| EPM26 | O111:HNM | 1 | 1a | 0.931 | 0.000 | + | + | − |
| EPM27 | O111:HNM | 1 | 1a | 1.053 | 0.000 | + | + | − |
| EPM017 | O112:H2 | 1 | 1c | 0.610 | 0.000 | + | + | − |
| EPM11 | O118:H16 | 1 | 1a | 0.869 | 0.000 | + | + | − |
| IAL6196 | O118:H16 | 1 | 1a | 0.801 | 0.000 | + | + | − |
| IAL6188 | O118:H16 | 1 | 1a | 0.566 | 0.009 | + | + | − |
| IAL6171 | O123:H− | 1 | 1a | 0.897 | 0.000 | + | + | − |
| IAL6181 | O123:H2 | 1 | 1a | 1.857 | 0.098 | + | + | − |
| IAL6180 | O123:H2 | 1 | 1a | 0.888 | 0.010 | + | + | − |
| IAL6197 | O123:HNM | 1 | 1a | 0.749 | 0.000 | + | + | − |
| IAL6192 | O153:H21 | 1 | 1a | 0.733 | 0.000 | + | + | − |
| 82 | O157:H7 | 1 | 1a | 0.963 | 0.000 | + | + | − |
| 3299-85 | O157:H7 | 1 | 1a | 1.032 | 0.078 | + | + | − |
| 46240 | O157:H7 | 1 | 1NT | 1.188 | 0.000 | + | + | − |
| 3077-88 | O157:H7 | 1 | 1a | 1.281 | 0.076 | + | + | − |
| C7-88 | O157:H7 | 1 | 1a | 1.312 | 0.006 | + | + | − |
| EPM01 | ONT:H8 | 1 | 1c | 0.584 | 0.000 | + | − | − |
| 184332 | OR:H19 | 1 | 1a | 0.408 | 0.000 | + | + | − |
| BA597 | OR:NM | 1 | 1a | 1.312 | 0.000 | + | + | − |
| IAL6176 | O8:H19 | 2 | 2a + 2d | 0.017 | 0.181 | + | − | − |
| 01-9582-01 | O39:HR | 2 | 2f | 0.018 | 2.929 | + | − | + |
| IALEc1054/05 | O91:H21 | 2 | 2a + 2c | 0.025 | 0.829 | + | − | + |
| IAL6201 | O100:H− | 2 | 2e | 0.028 | 0.602 | + | − | + |
| EPM82 | O112:H21 | 2 | 2c | 0.020 | 0.747 | + | − | + |
| IALEc226/04 | O113:H21 | 2 | 2a | 0.100 | 0.585 | + | − | + |
| IALEc678/04 | O113:H21 | 2 | 2a | 0.024 | 2.858 | + | − | + |
| IALEc603/04 | O141:H49 | 2 | 2a | 0.013 | 2.165 | + | − | + |
| IAL6182 | O153:H28 | 2 | 2NT | 0.000 | 0.202 | + | − | + |
| IALEc1167/05 | O157:H− | 2 | 2a + 2c + 2e | 0.768 | 0.339 | + | + | + |
| IALEc703/04 | O157:H− | 2 | 2a + 2d | 0.017 | 0.737 | + | − | + |
| IAL6193 | O157:H7 | 2 | 2a + 2c | 0.000 | 1.022 | + | − | + |
| IAL6207 | O157:H7 | 2 | 2a + 2c | 0.000 | 0.621 | + | − | + |
| IAL6179 | O157:H7 | 2 | 2a + 2c | 0.000 | 0.268 | − | − | + |
| IAL6202 | O157:H7 | 2 | 2a + 2c | 0.054 | 0.764 | + | − | + |
| EPM1 | O157:H7 | 2 | 2a + 2c | 0.092 | 2.847 | + | − | + |
| EPM2 | O157:H7 | 2 | 2a + 2c | 0.014 | 0.776 | + | − | + |
| EPM03 | O172:NM | 2 | 2a | 0.000 | 0.637 | + | − | + |
| IAL6199 | O177:H− | 2 | 2c | 0.000 | 0.752 | + | − | + |
| IAL6172 | O178:H19 | 2 | 2c | 0.102 | 0.810 | + | + | + |
| IALEc170/04 | ONT:H7 | 2 | 2a + 2f | 0.463 | 0.000 | + | + | − |
| EPM59 | ONT:H16 | 2 | 2d | 0.000 | 0.349 | + | − | + |
| EPM022 | ONT:H16 | 2 | 2b | 0.012 | 0.058 | + | − | − |
| IALEc157/05 | ONT:H23 | 2 | 2c + 2d | 0.027 | 0.332 | + | − | + |
| IAL6195 | ONT:H46 | 2 | 2a + 2d | 0.000 | 0.489 | + | − | + |
| BA1132 | ONT:H49 | 2 | 2a + 2c + 2d | 0.010 | 0.809 | + | − | + |
| BA1189 | ONT:H49 | 2 | 2a + 2d | 0.016 | 0.792 | + | − | + |
| IAL6198 | OR:H− | 2 | 2c | 0.005 | 1.985 | + | − | + |
| EPM79 | O22:H16 | 1/2 | 1a + 2c + 2d | 0.022 | 0.633 | + | − | + |
| IALEc515/05 | O43:H2 | 1/2 | 1NT + 2NT | 0.558 | 0.176 | + | + | + |
| BA3003 | O48:H7 | 1/2 | 1a + 2a | 0.651 | 0.638 | + | + | + |
| IALEc169/04 | O74:H25 | 1/2 | 1a + 2c | 2.267 | 1.149 | + | + | + |
| EPM036 | O75:H8 | 1/2 | 1c + 2b | 1.153 | 0.005 | + | + | − |
| IAL6208 | O75:H14 | 1/2 | 1c + 2NT | 1.819 | 0.141 | + | + | + |
| IALEc617/04 | O84:HNM | 1/2 | 1NT + 2NT | 0.049 | 0.794 | + | − | + |
| EPM50 | O87:H16 | 1/2 | 1NT + 2b | 0.008 | 0.071 | + | − | + |
| EPM4 | O93:H19 | 1/2 | 1a + 2d | 2.538 | 0.703 | + | + | + |
| EPM44 | O98:H4 | 1/2 | 1a + 2NT | 0.910 | 0.778 | + | + | + |
| EPM53 | O98:H17 | 1/2 | 1a + 2a + 2c | 0.747 | 0.781 | + | + | + |
| EPM55 | O98:H17 | 1/2 | 1a + 2a + 2c | 0.839 | 0.712 | + | + | + |
| EPM9 | O103:H2 | 1/2 | 1a + 2c | 0.940 | 0.912 | + | + | + |
| EPM66 | O105:H18 | 1/2 | 1a + 2a + 2b | 1.069 | 0.839 | + | + | + |
| EPM055 | O146:H21 | 1/2 | 1a + 2a + 2b | 1.144 | 0.674 | + | + | + |
| 3104-88 | O157:H7 | 1/2 | 1a + 2a | 0.502 | 1.111 | + | + | + |
| EDL933 | O157:H7 | 1/2 | 1a + 2a | 1.348 | 1.277 | + | + | + |
| EPM45 | O181:H4 | 1/2 | 1a + 2a | 0.941 | 0.274 | + | + | + |
| IALEc161/04 | ONT:H18 | 1/2 | 1a + 2a + 2c | 2.252 | 2.927 | + | + | + |
| EPM81 | ONT:H38 | 1/2 | 1NT + 2a | 0.517 | 0.990 | + | + | + |
| IALEc630/04 | ONT:H46 | 1/2 | 1a + 2f | 0.532 | 0.323 | + | + | + |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, M.A.; Santos, A.R.R.; Rocha, L.B.; Caetano, B.A.; Mitsunari, T.; Santos, L.I.; Polatto, J.M.; Horton, D.S.P.Q.; Guth, B.E.C.; dos Santos, L.F.; et al. Development and Validation of Shiga Toxin-Producing Escherichia coli Immunodiagnostic Assay. Microorganisms 2019, 7, 276. https://doi.org/10.3390/microorganisms7090276
Silva MA, Santos ARR, Rocha LB, Caetano BA, Mitsunari T, Santos LI, Polatto JM, Horton DSPQ, Guth BEC, dos Santos LF, et al. Development and Validation of Shiga Toxin-Producing Escherichia coli Immunodiagnostic Assay. Microorganisms. 2019; 7(9):276. https://doi.org/10.3390/microorganisms7090276
Chicago/Turabian StyleSilva, Miriam A., Anna Raquel R. Santos, Leticia B. Rocha, Bruna A. Caetano, Thais Mitsunari, Luanda I. Santos, Juliana M. Polatto, Denise S. P. Q. Horton, Beatriz E. C. Guth, Luís Fernando dos Santos, and et al. 2019. "Development and Validation of Shiga Toxin-Producing Escherichia coli Immunodiagnostic Assay" Microorganisms 7, no. 9: 276. https://doi.org/10.3390/microorganisms7090276
APA StyleSilva, M. A., Santos, A. R. R., Rocha, L. B., Caetano, B. A., Mitsunari, T., Santos, L. I., Polatto, J. M., Horton, D. S. P. Q., Guth, B. E. C., dos Santos, L. F., & Piazza, R. M. F. (2019). Development and Validation of Shiga Toxin-Producing Escherichia coli Immunodiagnostic Assay. Microorganisms, 7(9), 276. https://doi.org/10.3390/microorganisms7090276