A High Resolution DNA Melting Curve Analysis for the Rapid and Efficient Molecular Diagnostics of Extended Spectrum β-Lactamase Determinants from Foodborne Escherichia coli


Abstract
:1. Introduction
2. Materials and Methods
2.1. E. coli Isolates
2.2. Design of Group-Specific Primers for HRM Assays
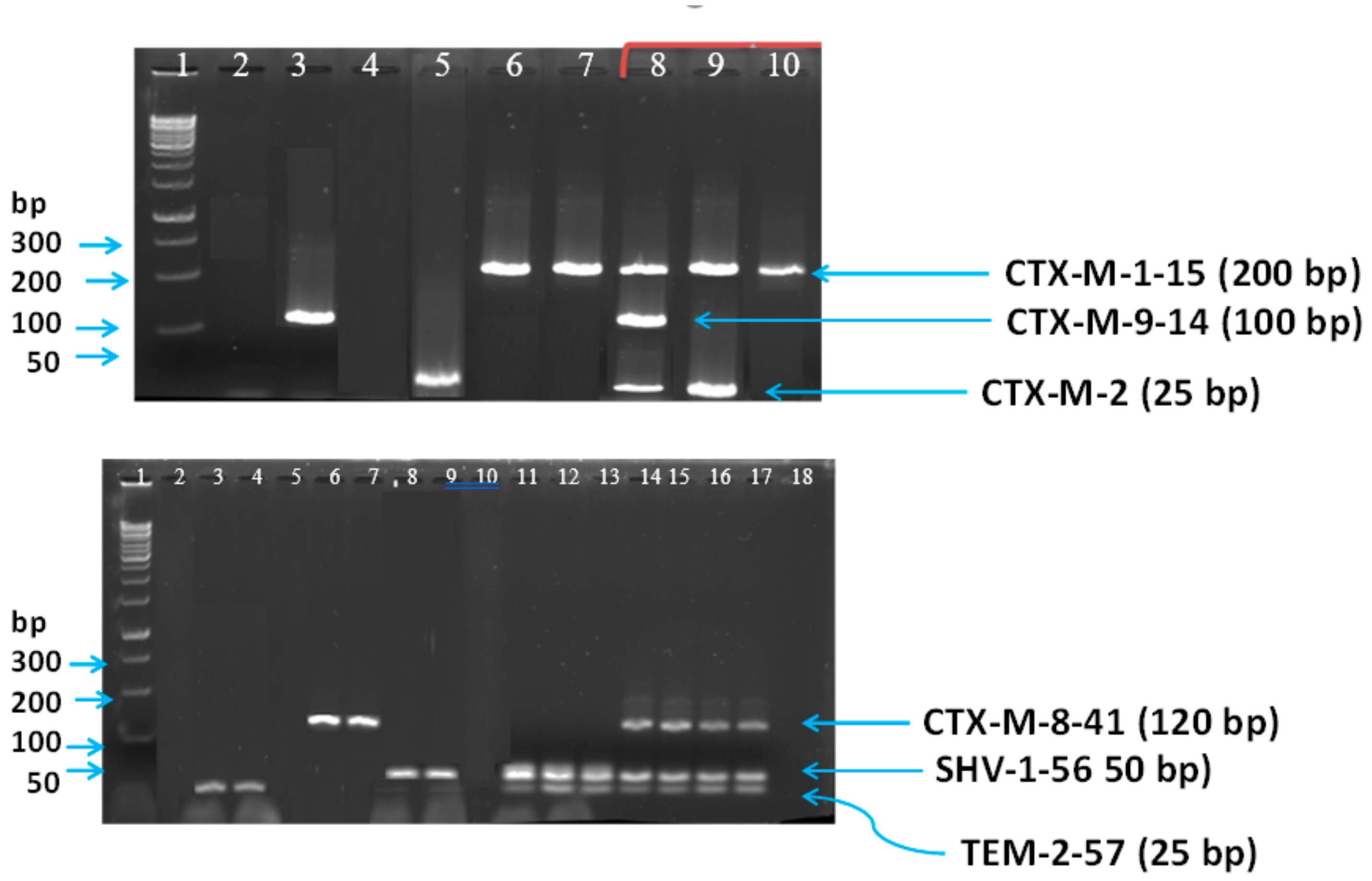
2.3. Multiplex PCR Technique
2.4. Assay Accuracy, Validation, and Sensitivity
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pitout, J.D.; Laupland, K.B. Extended-spectrum β-lactamase-producing Enterobacteriaceae: An emerging public-health concern. Lancet Infect. Dis. 2008, 8, 159–166. [Google Scholar] [CrossRef]
- Moubareck, C.; Lecso, M.; Pinloche, E.; Butel, M.J.; Doucet-Populaire, F. Inhibitory impact of bifidobacteria on the transfer of β-lactam resistance among Enterobacteriaceae in the gnotobiotic mouse digestive tract. Appl. Environ. Microbiol. 2007, 73, 855–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.A.; Hasan, F.; Ahmed, S.; Hameed, A. Extended-spectrum β-lactamases (ESBLs): Characterization, epidemiology and detection. Crit. Rev. Microbiol. 2004, 30, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Geiss, H.K.; Mack, D.; Stürenburg, E.; Seifert, H. Detection of extended-spectrum beta-lactamases among Enterobacteriaceae by use of semiautomated microbiology systems and manual detection procedures. J. Clin. Microbiol. 2007, 45, 1167–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum β-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, G.A. AmpC-Lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO/OIE. Report of the FAO/WHO/OIE Expert Meeting: Joint FAO/WHO/OIE Expert Meeting on Critically Important Antimicrobials; FAO: Rome, Italy, 2007; Available online: http://www.fao.org/3/a-i0204e.pdf (accessed on 16 July 2019).
- Sundsfjord, A.; Simonsen, G.S.; Haldorsen, B.C.; Haaheim, H.; Hjelmevoll, S.O.; Littauer, P.; Dahl, K.H. Genetic methods for detection of antimicrobial resistance. APMIS 2004, 112, 815–837. [Google Scholar] [CrossRef]
- Depoorter, P.; Persoons, D.; Uyttendaele, M.; Butaye, P.; De Zutter, L.; Dierick, K.; Herman, L.; Imberechts, H.; Van Huffel, X.; Dewulf, J. Assessment of human exposure to 3rd generation cephalosporin resistant E. coli (CREC) through consumption of broiler meat in Belgium. Int. J. Food Microbiol. 2012, 159, 30–38. [Google Scholar] [CrossRef]
- Manges, A.R.; Johnson, J.R. Food-borne origins of escherichia coli causing extraintestinal infections. Clin. Infect. Dis. 2012, 55, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Liu, J.H.; Lv, L.; Zong, Z.; Sun, Y.; Zheng, H.; Chen, Z.L.; Zeng, Z.L. Characterization of extended-spectrum β-lactamase genes found among Escherichia coli isolates from duck and environmental samples obtained on a duck farm. Appl. Environ. Microbiol. 2012, 78, 3668–3673. [Google Scholar] [CrossRef] [Green Version]
- Smet, A.; Martel, A.; Persoons, D.; Dewulf, J.; Heyndrickx, M.; Herman, L.; Haesebrouck, F.; Butaye, P. Broad-spectrum β-lactamases among Enterobacteriaceae of animal origin: Molecular aspects, mobility and impact on public health. FEMS Microbiol. Rev. 2010, 34, 295–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witte, W. Ecological impact of antibiotic use in animals on different complex microflora: Environment. Int. J. Antimicrob. Agents 2000, 14, 321–325. [Google Scholar] [CrossRef]
- Njage, P.M.K.; Buys, E.M. Quantitative assessment of human exposure to extended spectrum and AmpC β-lactamases bearing E. coli in lettuce attributable to irrigation water and subsequent horizontal gene transfer. Int. J. Food Microbiol. 2017, 240, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Njage, P.M.K.; Buys, E.M. Pathogenic and commensal Escherichia coli from irrigation water show potential in transmission of extended spectrum and AmpC β-lactamases determinants to isolates from lettuce. Microb. Biotechnol. 2015, 8, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Dallenne, C.; da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important β-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, J.; Wittwer, C.T.; Palais, R.; Zhou, L. Simultaneous mutation scanning and genotyping by high-resolution DNA melting analysis. Nat. Protoc. 2007, 2, 59–66. [Google Scholar] [CrossRef]
- Wittwer, C.T. High-resolution DNA melting analysis: Advancements and limitations. Hum. Mutat. 2009, 30, 857–859. [Google Scholar] [CrossRef]
- Zeinzinger, J.; Pietzka, A.T.; Stöger, A.; Kornschober, C.; Kunert, R.; Allerberger, F.; Mach, R.; Ruppitsch, W. One-step triplex high-resolution melting analysis for rapid identification and simultaneous subtyping of frequently isolated Salmonella serovars. Appl. Environ. Microbiol. 2012, 78, 3352–3360. [Google Scholar] [CrossRef] [Green Version]
- Pietzka, A.T.; Indra, A.; Stöger, A.; Zeinzinger, J.; Konrad, M.; Hasenberger, P.; Allerberger, F.; Ruppitsch, W. Rapid identification of multidrug-resistant Mycobacterium tuberculosis isolates by rpoB gene scanning using high-resolution melting curve PCR analysis. J. Antimicrob. Chemother. 2009, 63, 1121–1127. [Google Scholar] [CrossRef] [Green Version]
- Tajiri-Utagawa, E.; Hara, M.; Takahashi, K.; Watanabe, M.; Wakita, T. Development of a rapid high-throughput method for high-resolution melting analysis for routine detection and genotyping of noroviruses. J. Clin. Microbiol. 2009, 47, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Chia, J.H.; Chu, C.; Su, L.H.; Chiu, C.H.; Kuo, A.J.; Sun, C.F.; Wu, T.L. Development of a multiplex PCR and SHV melting-curve mutation detection system for detection of some SHV and CTX-M β-lactamases of Escherichia coli, Klebsiella pneumoniae, and Enterobacter cloacae in Taiwan. J. Clin. Microbiol. 2005, 43, 4486–4491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chromá, M.; Hricová, K.; Kolár, M.; Sauer, P.; Koukalová, D. Using newly developed multiplex polymerase chain reaction and melting curve analysis for detection and discrimination of β-lactamases in Escherichia coli isolates from intensive care patients. Diagn. Microbiol. Infect. Dis. 2011, 71, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Geyer, C.N.; Hanson, N.D. Multiplex high-resolution melting analysis as a diagnostic tool for detection of plasmid-mediated AmpC β-lactamase genes. J. Clin. Microbiol. 2014, 52, 1262–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendes, R.E.; Kiyota, K.A.; Monteiro, J.; Castanheira, M.; Andrade, S.S.; Gales, A.C.; Pignatari, A.C.C.; Tufik, S. Rapid detection and identification of metallo-β-lactamase-encoding genes by multiplex real-time PCR assay and melt curve analysis. J. Clin. Microbiol. 2007, 45, 544–547. [Google Scholar] [CrossRef] [Green Version]
- Aijuka, M.; Charimba, G.; Hugo, C.J.; Buys, E.M. Characterization of bacterial pathogens in rural and urban irrigation water. J. Water Health 2014, 13, 103–117. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Saitou, N. The neighbor-joining method: A new method for reco reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Jukes, T.H.; Cantor, C.R. Evolution of Protein Molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protocol | Target Name | Gene Targets | Name | Sequence (5′–3′) | Expected Amplicon Length (bp) | GenBank Accession No. |
|---|---|---|---|---|---|---|
| Multiplex 1 | CTX-M-1-15 | CTX-M group 1: CTX-M-1, CTX-M-3 and CTX-M-15 | CTX-M-1-15-F CTX-M-1-15-R | CGCAAATACTTTATCGTGCTGAT GATTCGGTTCGCTTTCACTTT | 102 | X92506 Y10278 AY044436 |
| CTX-M-2 | CTX-M group 2: CTX-M-2 | CTX-M-2-F CTX-M-2-R | CTGGTTCTGGTGACCTACTTTAC GCGATACCTCGCTCCATTTAT | 124 | X92507 | |
| CTX-M-9-14 | CTX-M group 9: CTX-M-9 and CTX-M-14 | CTX-M-9-14-F CTX-M-9-14-R | GCTCATCGATACCGCAGATAAT CCGCCATAACTTTACTGGTACT | 86 | AF174129 AF252622 | |
| Multiplex 2 | TEM-2-57 | TEM variants: TEM-1 and TEM-2 | TEM-2-57-F TEM-2-57-R | CGGATGGCATGACAGTAAGA TCCGATCGTTGTCAGAAGTAAG | 89 | X54606 |
| SHV-1-56 | SHV-1, SHV-2 and SHV-56 | SHV-1-56-F SHV-1-56-R | CTGGAGCGAAAGATCCACTATC CGCTGTTATCGCTCATGGTAA | 130 | AF148850 AF148851 EU586041 | |
| CTX-M-8-41 | CTX-M-8, CTX-M-25, CTX-M-26 and CTX-M-39 to CTX-M-41 | CTX-M-8-41-F CTX-M-8-41-R | GAGCCGACGCTCAACAC CACYGCCCAACGTCAGATT | 97 | AF189721 AF518567 AY157676 AY954516 DQ023162 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Njage, P.M.K.; Buys, E. A High Resolution DNA Melting Curve Analysis for the Rapid and Efficient Molecular Diagnostics of Extended Spectrum β-Lactamase Determinants from Foodborne Escherichia coli. Microorganisms 2020, 8, 90. https://doi.org/10.3390/microorganisms8010090
Njage PMK, Buys E. A High Resolution DNA Melting Curve Analysis for the Rapid and Efficient Molecular Diagnostics of Extended Spectrum β-Lactamase Determinants from Foodborne Escherichia coli. Microorganisms. 2020; 8(1):90. https://doi.org/10.3390/microorganisms8010090
Chicago/Turabian StyleNjage, Patrick Murigu Kamau, and Elna Buys. 2020. "A High Resolution DNA Melting Curve Analysis for the Rapid and Efficient Molecular Diagnostics of Extended Spectrum β-Lactamase Determinants from Foodborne Escherichia coli" Microorganisms 8, no. 1: 90. https://doi.org/10.3390/microorganisms8010090