Combating Parasitic Nematode Infections, Newly Discovered Antinematode Compounds from Marine Epiphytic Bacteria


1
Centre for Marine Science and Innovation, School of Biological, Earth and Environmental Sciences, UNSW, Sydney, NSW 2052, Australia
2
School of Industrial Technology, Universiti Sains Malaysia, USM, 11800 Penang, Malaysia
*
Author to whom correspondence should be addressed.
Microorganisms 2020, 8(12), 1963; https://doi.org/10.3390/microorganisms8121963
Submission received: 4 November 2020
/
Revised: 8 December 2020
/
Accepted: 8 December 2020
/
Published: 11 December 2020
(This article belongs to the Special Issue Microbial Secondary Metabolites and Biotechnology)
Abstract
:Parasitic nematode infections cause debilitating diseases and impede economic productivity. Antinematode chemotherapies are fundamental to modern medicine and are also important for industries including agriculture, aquaculture and animal health. However, the lack of suitable treatments for some diseases and the rise of nematode resistance to many available therapies necessitates the discovery and development of new drugs. Here, marine epiphytic bacteria represent a promising repository of newly discovered antinematode compounds. Epiphytic bacteria are ubiquitous on marine surfaces where they are under constant pressure of grazing by bacterivorous predators (e.g., protozoans and nematodes). Studies have shown that these bacteria have developed defense strategies to prevent grazers by producing toxic bioactive compounds. Although several active metabolites against nematodes have been identified from marine bacteria, drug discovery from marine microorganisms remains underexplored. In this review, we aim to provide further insight into the need and potential for marine epiphytic bacteria to become a new source of antinematode drugs. We discuss current and emerging strategies, including culture-independent high throughput screening and the utilization of Caenorhabditis elegans as a model target organism, which will be required to advance antinematode drug discovery and development from marine microbial sources.
1. Introduction
Humans are vulnerable to infectious diseases caused by parasitic helminths (nematodes) resulting in morbidity and mortality within the population [1,2,3]. Approximately 24% of the global human population, corresponding to 1.5 billion people, suffer from parasitic helminth infections [4]. There are almost 300 nematodes associated with zoonotic diseases that are able to infect humans, including some of the most devastating parasites such as Ascaris lumbricoides (roundworm), Ancylostoma duodenale (hookworm), Gnathostoma spinigerum, Halicephalobus gingivalis and Trichinella spiralis (Trichina worm) [1,5,6]. While some parasitic nematodes (e.g., Ancylostoma duodenale, Strongyloides stercoralis and Halicephalobus gingivalis) can penetrate human skin or invade through existing skin lesions [7,8], several parasites infect humans via ingestion of food products contaminated with the embryonated eggs (e.g., Ascaris lumbricoides and Trichuris trichiura) [9,10] or from eating raw or undercooked freshwater fish, birds, frogs or reptiles contaminated with the parasitic nematode larvae (e.g., Gnathostoma spinigerum, Dracunculus medinensis, Eustrongylides sp. and Trichinella spiralis) [11,12,13]. Apart from causing diseases, a high nematode burden also reduces human fecundity [14] and affects children through malnutrition, stunted development and cognitive delay [15,16].
Plant parasitic nematodes (PPNs) also cause diseases [17,18] either by damaging the root system, retarding the plant development or by exposing plants to secondary bacterial, fungal or viral infections [19,20,21,22]. It is estimated that damages caused by PPNs result in a >12% loss in global crop productivity and an average annual loss of ~US$215 billion [23]. Parasitic nematodes also impede the productivity of fisheries and aquaculture industries, resulting in worldwide economic losses and health hazards (i.e., zoonotic diseases or allergies) for consumers [24,25]. For example, the global financial loss in the finfish industry due to parasitic infection is estimated to be as large as US$134 million per annum [24]. Major fish products such as Atlantic mackerel, herring, European hake, Atlantic cod and anchovy are commonly associated with parasitic nematodes e.g., Anisakis sp. and Pseudoterranova sp. [26], which are transmitted to humans via ingestion of undercooked or raw fish [27,28]. Furthermore, parasitic nematodes cause different pathophysiological symptoms in livestock, reduce meat quality and cause animal mortality [29,30]. In Australia, the economic loss due to parasite infection (e.g., Ostertagia sp. and Trichostrongylus sp.) and the cost of control management is estimated to be ~AU$1 billion [31,32,33] whereas in Kenya, South Africa and India, ~US$26, 46 and 103 million is spent just to control the Haemonchus contortus nematode infection of livestock, respectively [34,35]. While diminishing animal fecundity, nematode parasites such as Ostertagia ostertagi (brown stomach worm) and Dictyocaulus viviparus (lungworm) reduce dairy production levels (~1.25 L/day/animal or 1.6 L/day/cow, respectively) [36,37].
Currently, the most promising longer-term control strategy is dependent on pharmaceutically derived chemotherapeutic treatments to kill parasitic nematodes and/or mitigate the spread of infection [38,39]. Unfortunately, the emergence of drug resistance among parasitic nematodes, due to prolonged treatment and incorrect drug dosage with an unpredictable infection trend as a result of climate change, is alarming [40,41,42,43,44]. The current prevalence of drug resistance is not only limited to the older classes of antinematode drugs but also those introduced in recent years. For example, monepantel resistance in T. circumcincta, Trichostrongylus colubriformis and H. contortus has been reported in New Zealand and the Netherlands within just five years of launching the drug [45,46,47].
Drug resistance has also been reported for almost all of the currently available anthelmintic drugs and nematicide classes including piperazine, benzimidazoles, levamisole, (pyrantel and morantel), paraherquamide, ivermectin (macrocylic lactones and milbemycins), emodepside (cyclodepsipeptides, PF1022A) and nitazoxanide [47,48,49].
Given the impacts of resistant parasitic nematodes to human and economic growth, novel antinematode chemotherapeutic agents are urgently needed as a preventive control against parasite infestation [4,50]. For decades, microorganisms, particularly bacteria, have served as a precious source for bioactive compounds, some of which have been developed into novel drugs including those with nematicidal activity [51,52]. This review highlights the potential of marine epiphytic bacteria as a new repository for newly discovered antinematode metabolites and the underlying mechanism of antinematode compound production by marine microbial biofilms. Furthermore, the utilization of C. elegans as a surrogate organism for antinematode drug development will be reviewed.
2. Antinematode Drug Discovery: Transition from Terrestrial to Marine-Derived Microbial Compounds
Terrestrial plants and plant extracts have been documented as an ancient therapeutic treatment against parasitic nematodes [53] and today, extensive studies to isolate plant-derived nematicidal compounds are still ongoing [54,55]. However, the re-discovery rate of bioactive metabolites is high [4], reducing the number of newly discovered compounds in the drug discovery pipeline [56]. Moreover, external factors, e.g., specific planting season, environmental temperature and humidity, may also affect the compounds’ reproducibility by the plant producers [4]. Given those inevitable challenges, microbial-derived anthelmintic compounds offer a promising solution [57].
Extensive exploration of terrestrial microbial compounds for therapeutic drug development was initiated in the 20th century [53]. Since then, more than 50,000 beneficial bioactive metabolites have been successfully identified [58,59] some of which have potent nematotoxicity via distinct modes of action (MOAs) (Table 1). Unfortunately, after almost 50 years of drug screening, few newly discovered compounds have been identified from terrestrial-borne microorganisms [59] hence the requirement for a new source of antinematode drug discovery to combat the rapidly growing nematode resistance. Here, the underexplored marine ecosystem, with highly diverse unidentified macro- and microorganisms, represents a new repository for novel nematicidal drugs [59]. In fact, marine bioactive compounds have been acknowledged as having substantial chemical novelty compared to terrestrial metabolites [60].
The marine environment represents the largest biome on the earth (70% of the earth surface; ~361 million km2 with average depth of 3730 m) and provides a habitat for a wide diversity of life that outnumbers terrestrial environments [61,62,63,64]. Microorganisms are abundant in this ecosystem (105 to 106 of cells per milliliter), reaching an average 1028 to 1029 cells either in the open ocean, deep sea, sediment or on the subsurface [64,65,66]. However, the majority of marine bacteria remain unrecognized or uncultivable [67]. Given the enormous diversity and untapped bioactive potential, marine microorganisms are likely to produce newly discovered bioactive compounds with a well-defined molecular architecture and biological function including nematicidal activities [68,69].

{kind=link}
{kind=link}
Table 1.
Examples of nematicidal compounds produced by bacteria isolated from terrestrial environments and their modes of action (MOAs) against the target nematodes.
Table 1.
Examples of nematicidal compounds produced by bacteria isolated from terrestrial environments and their modes of action (MOAs) against the target nematodes.
| Microbial Producer | Compound | Mode of Action | Target Nematode | Affected Nematode Region | Reference |
|---|---|---|---|---|---|
| Bacillus thuringiensis | Crystal toxin Cry5B, Cry21A | Toxin binds to nematode glycoconjugate receptor and disrupt the intestinal cells membrane integrity. This action causes fall of nematode brood size and mortality | Ancylostoma ceylanicum, Ascaris suum, C. elegans | Gastrointestinal system | [70,71,72] |
| Bacillus simplex, B. subtilis, B. weihenstephanensis, Microbacterium oxydans, Stenotrophomonas maltophilia, Streptomyces lateritius and Serratia marcescens | Volatile organic compound (VOC) i.e., benzaldehyde, benzeneacetaldehyde, decanal, 2-nonanone, 2-undecanone, cyclohexene and dimethyl disulfide | VOCs reduce nematode motility and death | Panagrellus redivivus, Bursaphelenchus xylophilus | Unknown | [73] |
| Streptomyces avermectinius | Avermectin and ivermectin (semi-synthetic) | Compound exposure resulted in pharyngeal paralysis and nematode death | Haemonchus contortus, Brugia malayi, C. elegans | Neuromuscular system | [48,74,75,76] |
| Serratia marcescens | Prodigiosin | Compound is toxic against juveniles larvae and inhibit egg hatching competency | Radopholus similis, Meloidogyne javanica | Unknown | [77] |
| Pseudomonas aeruginosa | Phenazine toxin (phenazine-1-carboxylic, pyocyanin and 1-hydroxyphenazine) | Phenazine-1-carboxylic shows fast killing activity against nematode in acidic environment whilst pyocyanin is toxic in neutral or basic pH. The toxicity of 1-hydroxyphenazine is not dependent on environmental pH. Continuous exposure to phenazine affects protein homeostasis and causes neurodegeneration | C. elegans | Neuromuscular system, cell mitochondria and protein folding | [78,79] |
| Pseudomonas aeruginosa | Exotoxin A and other undetermined effectors | Slow-killing activity against nematode is based on infection-like process thus resulting in accumulation of bacteria in the gut. Continuous exposure leads to ceased pharyngeal pumping, nematode immobility and death | C. elegans | Gastrointestinal system | [80] |
| Pseudomonas aeruginosa | Chitinase enzyme | Chitinase degrades nematode cuticle, intestine and egg shell leading to the animal death | C. elegans | Cuticle, eggs, gastrointestinal system | [81] |
| Pseudomonas plecoglossicida | Glycolipid biosurfactant | Reduction of nematode development, survival and fecundity | C. elegans | Unknown | [82] |
3. Surface Associated Marine Bacteria: A Reservoir for Novel Antimicrobial and Antinematode Drug Discovery
Marine inhabitants, particularly microorganisms, are continuously exposed to multiple detrimental interactions imposed by competitors and predators [83,84] and different physical–chemical variables such as fluctuating temperature, pH, UV exposure, salinity, toxic compounds and desiccation, particularly in the intertidal zone [85,86,87]. As a survival strategy, some marine bacteria adhere to each other and/or surfaces and are embedded in enclosed matrices to form a biofilm (Figure 1) [88,89].
The continuous development of biofilm on marine surfaces leads to epibiosis, which involves multispecies biofilm formation [90,91]. Macroalgal surfaces, for example, are a hot-spot for colonization by opportunistic epibionts such as algal spores, invertebrate larvae, diatoms, fungi and other bacteria [85,92], mostly due to the accumulation of nutrients and macroalgal exudates composed of organic carbon and nitrogen particles [93,94,95]. Consequently, the competition among marine microorganisms to reserve a space within the biofilm community is tremendously intense and bacterial strains that are equipped with broad-spectrum inhibitory phenotypes are likely to be successful epibiotic colonizers (Figure 1) [83,96].
In addition, predation by heterotrophic protozoa and bacterivorous nematodes represents another biotic stress resulting in major mortality for both planktonic and surface-associated bacteria in the marine habitat (Figure 1) [85,97]. Protozoans, for example Rhynchomonas nasuta and Cafeteria roenbergensis, are among the most abundant ubiquitous species in the ocean and are the major controllers of the food web in the marine environment through their function as bacterial predators [98,99,100]. Nematodes such as Pareudiplogaster pararmatus are among the natural consumers of organic biomass in benthic habitats actively grazing bacterial mats and the biofilms of biotic surfaces [101,102].
The omnipresence of inter- and intra-species interactions supports the evolution of diverse defense strategies by surface-associated marine bacteria. Such defense mechanisms include the production of bioactive compounds showing antibacterial, antifungal, antitumor, antifouling, antiprotozoal and antinematode activities (Table 2) [103,104]. Interestingly, the physical–chemical properties, molecular structure and functional features of those marine microbial compounds are believed to be shaped by the naturally harsh conditions of the marine environment [105]. Moreover, it is speculated that bioactive metabolites originally attributed to marine invertebrates such as sponges, tunicates, bryozoans and molluscs are actually produced by their associated microorganisms [106,107]. For example, the antibiotic peptides andrimid and trisindoline isolated from the sponge Hyatella sp. and Hyrtios altum are believed to be produced by symbiotic Vibrio sp. [108,109]. An antitumor cyclic peptide leucamide A isolated from the sponge Leucetta microraphis is closely related to compounds produced by cyanobacterial symbionts [110]. In addition, a commercialized antitumor drug Didemnin B initially isolated from the tunicate Trididemnum solidum [111] was recently demonstrated to be produced by symbiotic bacteria Tistrella mobilis and T. bauzanensis [112,113]. These observations also hold true for numerous antinematode compounds that have been successfully isolated from marine eukaryotes, with production of many now attributed to host-associated microorganisms [51,114].
The potential for marine surface-associated microorganisms to be repositories of unusual gene functions and bioactivities is further corroborated by global biodiversity studies such as the Tara Oceans [69,115] and the Global Ocean Sampling (GOS) expedition [116,117]. These and other studies continue to reveal unprecedented levels of information on microbial diversity, which has subsequently led to investigations on underexplored bacterial diversity in various marine ecosystems. Furthermore, abundant and newly discovered biosynthetic gene clusters encoding rare non-ribosomal peptides (NRPS), polyketides (PKS) and NRPS–PKS hybrids have been discovered from marine biofilm samples, further strengthening the concept of the marine environment as a rich source of newly discovered bioactive compounds [68,69].
Table 2.
Example of anthelmintic or nematicidal bioactivities isolated from marine bacteria.
| Marine Microbial Producer | Compound | Associated Surface/Host | Mode of Action | Responsible Gene(S) | Reference |
|---|---|---|---|---|---|
| Microbulbifer sp. D250 | Violacein | Algae Delisea pulchra | Facilitate bacterial accumulation accompanied by tissue damage and apoptosis | VioA-VioE | [118] |
| Pseudoalteromonas tunicata D2 | Tambjamine | Algae Ulva australis | Slow-killing activity by a heat-resistant tambjamine and substantial bacterial colonization in the nematode gut | TamA-TamT | [119] |
| Pseudoalteromonas tunicata D2 | Unknown | Algae Ulva australis | Fast-killing activity by a heat sensitive unknown compound through colonization-independent manner | Unknown | [119] |
| Uncultured alpha-proteobacterium, JN874385 (strain U95) | Unknown | Algae Ulva australis | Undetermined | Possibly NRPS gene | [120] |
| Aequorivita sp. | Unknown | Antarctic marine sediment | Undetermined | Unknown | [121] |
| Pseudovibrio sp. Pv348, 1413, HE818384 (strain D323) | Unknown | Algae Delisea pulchra | Undetermined | Unknown | [120] |
| Heterologous clone jj117 (NCBI Accession number SRX4339430) | Unknown | Ulva australis metagenomic library | Undetermined | ATP-grasp protein/alpha-E protein/transglutaminase protein/protease | [122] |
| Vibrio atlanticus strain S-16 | Volatile organic compounds (VOC) | Scallop Argopecten irradians | Undetermined | Unknown | [123] |
| Virgibacillus dokdonensis MCCC 1A00493 | VOC (acetaldehyde, dimethyl disulfide, ethylbenzene and 2-butanone | Polymetallic nodules in the deep sea | Direct contact killing activity, fumigation. Acetaldehyde had a fumigant activity to impede egg hatching | Unknown | [124] |
| Pseudoalteromonas rubra | Unknown | Marine organisms (copepod or fish) or environmental samples | Undetermined | Unknown | [125,126] |
| Pseudoalteromonas piscicida | Unknown | Marine organisms (copepod or fish) or environmental samples | Undetermined | Unknown | [125,126] |
| Arthrobacter davidanieli | Unknown | Marine environmental sample | Undetermined | Unknown | [125,127] |
| Pseudoalteromonas luteoviolacea | Unknown | Marine organisms (copepod or fish) or environmental samples | Undetermined | Unknown | [125,126] |
| Photobacterium halotolerans | Unknown | Marine organisms (copepod or fish) or environmental samples | Undetermined | Unknown | [125,126] |
4. Caenorhabditis elegans as a Model Organism for Antinematode Drug Discovery and Development
Antinematode drug research is impeded by several challenges. These include (i) similarity of biochemical reactions between parasitic nematodes and the infected host, (ii) complex parasite life cycles that involve infections in multiple hosts, (iii) different parasite geographical locations and (iv) rapid development of resistant phenotypes. Therefore, an easily maintained nematode model with the capability for rapid screening of potential antinematode compounds represents a solution to some of these challenges [128,129].
Sydney Brenner and colleagues first introduced the free-living soil nematode C. elegans in 1965 as an animal model for research including anti-infective and antinematode drug studies [130,131,132,133,134,135]. Owing to its small size (1–1.5 mm-adult length, 80 µm-diameter), transparency, rapid life cycle (~3 days) and diet based on a simple bacterial culture of Escherichia coli OP50, C. elegans has emerged as a valuable animal model [134,136,137]. C. elegans possess several features which make the organism an efficient and low-cost surrogate organism for antinematode drug discovery. Unlike parasitic nematodes which require vertebrate hosts for reproduction and maintenance [48,138], synchronized C. elegans can be easily propagated at the desired life stage on Nematode Growth Media (NGM) ready to be used for antinematode drug studies [139]. The earliest antinematode drug testing using synchronized C. elegans individuals was performed by exposing the animals to nematicidal agents incorporated into the agar media [140]. After a few years, the screening protocol evolved rapidly with the development of high throughput screening methods employing micro-fluidic systems or high-content screening (HCS) technologies, allowing for fast and large-scale drug testing (~14,000 to 360,000 of compounds) using C. elegans as the model organism (Figure 2) [141,142]. Today, important drug discoveries, such as benzimidazoles [143], ivermectin and its analogues, moxidectin, milbemycin oxime, doramectin, selamectin, abamectin, eprinomectin [144] and the nematicidal activity of crystal protein insecticide Cry5B, Cry21A from Bacillus thuringiensis [72,145], can be attributed to the use of C. elegans as an effective animal model [48].
C. elegans shares many conserved genes and protein functions with parasitic nematodes. Analysis of the intestinal parasite Strongyloides stercoralis genetic sequences showed 85% of protein homologs to C. elegans genes. The infective stage of S. stercoralis (L3i/dauer) also shows an increased proportion of protein homologs to C. elegans dauer larvae [146]. Genetic manipulation and RNA interference (RNAi) studies have been widely performed on C. elegans to provide a better understanding of the nematode response against the nematicidal drugs at the molecular level (Figure 2). Quantitative polymerase chain reaction (qPCR) and “omic” technologies, e.g., transcriptomic profiling and proteomics, are also being used to provide a global snapshot of the molecular response of C. elegans to drug exposure [147,148]. Given the conserved gene homologs and protein function among the members of phylum Nematoda, these studies provide insight into the possible mechanisms used by parasitic nematodes against similar drug exposure [48].
The majority of the current drugs used to treat parasitic nematode infections target proteins that regulate neuromuscular activity including neurotransmitter receptors and ion channels [48]. Utilization of C. elegans as a model organism confers a better understanding of the mode of action (MOA) of potential nematicidal drugs (Figure 2). Studies have shown that C. elegans’ neuromuscular system, its major neurotransmitters GABA (4-aminobutyric acid) and glutamate and its enzyme choline acetyltransferase, responsible for the synthesis of the neurotransmitter acetylcholine, display strong similarities to parasitic roundworms Ascaris suum and Ascaris lumbricoides [48,149,150,151,152]. Therefore, exposure of C. elegans to antinematode compounds which target the neurotransmitter receptor and ion channels has enabled the discovery of the target binding molecule and the resulting toxicity to parasitic nematodes [153]. For example, observations of body muscle contraction and spastic paralysis in C. elegans exposed to levamisole, led Lewis and colleagues [154] to determine that binding to the muscle acetylcholine receptors was key to its activity. The initiation of amino-acetonitrile derivative (AAD) toxic activity against nematodes by binding to a nicotinic acetylcholine receptor was also revealed via forwards genetic screening using C. elegans mutants [155]. More recently, the MOA of paraherquamide, a broad-spectrum nicotinic nematicidal alkaloid isolated from Penicillium paraherquei [156], and antinematode plant-derived compounds [157] were also elucidated using C. elegans [157,158,159]. While using C. elegans to identify specific targets is promising, assessing side effects related to these newly discovered compounds will be important considering many of the neurotransmitter targets will also be present in the host organism [48]. Nevertheless, in the event that novel drugs negatively impact non-target organisms, derivatives may be developed with reduced side effects [160,161].
5. Conclusions
The increasing prevalence of parasitic nematode infection and the emergence of antinematode drug resistance represent critical global issues, impacting human wellbeing and economic development. Moreover, increasing temperatures and changing weather patterns, moisture and rainfall as a result of global climate change may also escalate the prevalence of parasitic nematode diseases worldwide [40,41,42,43,44]. Although for decades many anthelmintic/nematicidal drugs have been successfully derived from terrestrial microorganisms (see examples in Table 1), the paucity of new antinematode drug classes and the challenges now presented with drug resistance underline the importance of discovering new antinematode bioactive metabolites from natural sources [4,50]. This review highlighed the potential of marine epiphytic bacteria as a new platform for novel antinematode drug development. Marine epiphytic bacteria are highly diverse, harboring unique genes expressing newly discovered bioactive metabolites with commercially or pharmaceutically relevant biological potentials including antinematode activities [118,119,120]. Owing to the extraordinary molecular structure and physical/chemical properties, marine bioactive compounds are regarded as “blue gold from the ocean” and are believed to be a promising source for future novel antinematode drugs. The ability to uncover these novel marine-derived antinematode drugs will be dependent on the successful implementation of innovative cultures, culture-independent techniques and high-throughput bioassays, for which the model nematode C. elegans is well suited.
Author Contributions
N.H.S., J.N., M.E.M. and S.E. developed the conceptual ideas. N.H.S. wrote the initial draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
Authors would like to thank CMSI staff and students for helpful discussions. Nor Hawani Salikin was supported by the Ministry of Higher Education Malaysia and Universiti Sains Malaysia under the Academic Staff Training Scheme Fellowship.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cox, F.E.G. History of human parasitic diseases. Infect. Dis. Clin. N. Am. 2004, 18, 171–188. [Google Scholar] [CrossRef] [PubMed]
- Bañuls, A.-L.; Thomas, F.; Renaud, F. Of parasites and men. Infect. Genet. Evol. 2013, 20, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viney, M. How can we understand the genomic basis of nematode parasitism? Trends Parasitol. 2017, 33, 444–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Bustos, J.F.; Sleebs, B.E.; Gasser, R.B. An appraisal of natural products active against parasitic nematodes of animals. Parasites Vectors 2019, 12, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashford, R.; Crewe, W. Parasites of Homo Sapiens: An Annotated Checklist of the Protozoa, Helminths and Arthropods for Which We are Home, 2nd ed.; Taylor & Francis: London, UK; New York, NY, USA, 2003; pp. 87–93. [Google Scholar]
- Stoltzfus, D.J.; Pilgrim, A.A.; Herbert, D.B.R. Perusal of parasitic nematode ‘omics in the post-genomic era. Mol. Biochem. Parasitol. 2017, 215, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Papadi, B.; Eberhard, M.E.; Boudreaux, C.; Bishop, H.; Mathison, B.; Tucker, J.A. Halicephalobus gingivalis: A rare cause of fatal meningoencephalomyelitis in humans. Am. J. Trop. Med. Hyg. 2013, 88, 1062–1064. [Google Scholar] [CrossRef] [Green Version]
- Bryant, A.S.; Ruiz, F.; Gang, S.S.; Castelletto, M.L.; Lopez, J.B.; Hallem, E.A. A critical role for thermosensation in host seeking by skin-penetrating nematodes. Curr. Biol. 2018, 28, 2338–2347.e6. [Google Scholar] [CrossRef] [Green Version]
- Bundy, D.A.; De Silva, N.; Appleby, L.J.; Brooker, S.J. Intestinal nematodes: Ascariasis. In Hunter’s Tropical Medicine and Emerging Infectious Diseases, 10th ed.; Ryan, E.T., Hill, D.R., Solomon, T., Aronson, N.E., Endy, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 840–844. [Google Scholar] [CrossRef]
- Jourdan, P.M.; Lamberton, P.H.L.; Fenwick, A.; Addiss, D.G. Soil-transmitted helminth infections. Lancet 2018, 391, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Frean, J. Gnathostomiasis acquired by visitors to the Okavango Delta, Botswana. Trop. Med. Infect. Dis. 2020, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Eberhard, M.L.; Cleveland, C.A.; Zirimwabagabo, H.; Yabsley, M.J.; Ouakou, P.T.; Ruiz-Tiben, E. Guinea worm (Dracunculus medinensis) infection in a wild-caught frog, Chad. Emerg. Infect. Dis. 2016, 22, 1961–1962. [Google Scholar] [CrossRef]
- Magnino, S.; Colin, P.; Dei-Cas, E.; Madsen, M.; McLauchlin, J.; Nöckler, K.; Maradona, M.P.; Tsigarida, E.; Vanopdenbosch, E.; Van Peteghem, C. Biological risks associated with consumption of reptile products. Int. J. Food Microbiol. 2009, 134, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Persson, G.; Ekmann, J.R.; Hviid, T.V.F. Reflections upon immunological mechanisms involved in fertility, pregnancy and parasite infections. J. Reprod. Immunol. 2019, 136, 102610. [Google Scholar] [CrossRef] [PubMed]
- Ezeamama, A.E.; McGarvey, S.T.; Olveda, R.M.; Acosta, L.P.; Kurtis, J.D.; Bellinger, D.C.; Langdon, G.C.; Manalo, D.L.; Friedman, J.F. Helminth infection and cognitive impairment among Filipino children. Am. J. Trop. Med. Hyg. 2005, 72, 540–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doni, N.Y.; Zeyrek, F.Y.; Simsek, Z.; Gurses, G.; Sahin, I. Risk factors and relationship between intestinal parasites and the growth retardation and psychomotor development delays of children in Şanlıurfa, Turkey. Turk. Parazitolojii Derg. 2015, 39, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Abad, P.; Gouzy, J.; Aury, J.; Castagnone-Sereno, P.; Danchin, E.G.J.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, G.C.; Egnin, M.; Bonsi, C. The impact of plant-parasitic nematodes on agriculture and methods of control. In Nematology-Concepts, Diagnosis and Control; Manjur-Shah, M., Mahamood, M., Eds.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef]
- Kassie, Y.G.; Ebrahim, A.S.; Mohamed, M.Y. Interaction effect between Meloidogyne incognita and Fusarium oxysporum f. sp. lycopersici on selected tomato (Solanum lycopersicum L.) genotypes. Afr. J. Agric. Res. 2020, 15, 330–342. [Google Scholar] [CrossRef]
- Liu, W.; Park, S.-W. Underground mystery: Interactions between plant roots and parasitic nematodes. Curr. Plant Biol. 2018, 15, 25–29. [Google Scholar] [CrossRef]
- Villate, L.; Morin, E.; Demangeat, G.; Van Helden, M.; Esmenjaud, D. Control of Xiphinema index populations by fallow plants under greenhouse and field conditions. Phytopathology 2012, 102, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elgawad, M.; Askary, T.H. Impact of phytonematodes on agriculture economy. In Biocontrol Agents of Phytonematodes; Askary, T.H., Martinelli, P.R.P., Eds.; CAB International Publishing: Oxfordshire, UK; Boston, MA, USA, 2015; pp. 12–17. [Google Scholar]
- Shinn, A.; Pratoomyot, J.; Bron, J.E.; Paladini, G.; Brooker, E.; Brooker, A.J. Economic costs of protistan and metazoan parasites to global mariculture. Parasitology 2015, 142, 196–270. [Google Scholar] [CrossRef]
- Mehrdana, F.; Buchmann, K. Excretory/secretory products of anisakid nematodes: Biological and pathological roles. Acta Vet. Scand. 2017, 59. [Google Scholar] [CrossRef] [Green Version]
- Levsen, A.; Svanevik, C.S.; Cipriani, P.; Mattiucci, S.; Gay, M.; Hastie, L.C.; Bušelić, I.; Mladineo, I.; Karl, H.; Ostermeyer, U.; et al. A survey of zoonotic nematodes of commercial key fish species from major European fishing grounds—Introducing the FP7 PARASITE exposure assessment study. Fish. Res. 2018, 202, 4–21. [Google Scholar] [CrossRef]
- Aibinu, I.E.; Smooker, P.M.; Lopata, A.L. Anisakis nematodes in fish and shellfish- from infection to allergies. Int. J. Parasitol. Parasites Wildl. 2019, 9, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, S. Seafood-borne parasitic diseases: A “one-health” approach is needed. Fishes 2019, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Mushonga, B.; Habumugisha, D.; Kandiwa, E.; Madzingira, O.; Samkange, A.; Segwagwe, B.E.; Jaja, I.F. Prevalence of Haemonchus contortus infections in sheep and goats in Nyagatare District, Rwanda. J. Vet. Med. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elseadawy, R.; Abbas, I.; Al-Araby, M.; Hildreth, M.B.; Abu-Elwafa, S. First evidence of Teladorsagia circumcincta infection in sheep from Egypt. J. Parasitol. 2019, 105, 484–490. [Google Scholar] [CrossRef]
- McLeod, R. Costs of major parasites to the Australian livestock industries. Int. J. Parasitol. 1995, 25, 1363–1367. [Google Scholar] [CrossRef]
- Roeber, F.; Jex, A.R.; Gasser, R.B. Impact of gastrointestinal parasitic nematodes of sheep, and the role of advanced molecular tools for exploring epidemiology and drug resistance—An Australian perspective. Parasites Vectors 2013, 6, 153. [Google Scholar] [CrossRef] [Green Version]
- Sackett, D.; Sackett, H.; Abbott, K.; Barber, M. Assessing the Economic Cost of Endemic Disease on the Profitability of Australian Beef Cattle and Sheep Producers. Meat & Livestock Australia Report AHW. 2006. Available online: https://www.mla.com.au/Research-and-development/Search-RD-reports/RD-report-details/Animal-Health-and-Biosecurity/Assessing-the-economic-cost-of-endemic-disease-on-the-profitability-of-Australian-beef-cattle-and-sheep-producers/120 (accessed on 16 May 2020).
- McLeod, R.S. Economic impact of worm infestations in small ruminants in South East Asia, India and Austrailia. In Worm Control of Small Ruminants in Tropical Asia; Sani, R.A., Gray, G.D., Baker, R.L., Eds.; ACIAR Monograph: Canberra, Australia, 2004; Volume 113, pp. 23–33. [Google Scholar]
- Maqbool, I.; Wani, Z.A.; Shahardar, R.A.; Allaie, I.M.; Shah, M.M. Integrated parasite management with special reference to gastro-intestinal nematodes. J. Parasit. Dis. 2017, 41, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.; Akbar, H.; Ahmad, L.; Hassan, M.A.; Ashraf, K.; Saeed, K.; Gharbi, M. A systematic review on modelling approaches for economic losses studies caused by parasites and their associated diseases in cattle. Parasitology 2019, 146, 129–141. [Google Scholar] [CrossRef]
- May, K.; Brügemann, K.; König, S.; Strube, C. The effect of patent Dictyocaulus viviparus (re)infections on individual milk yield and milk quality in pastured dairy cows and correlation with clinical signs. Parasites Vectors 2018, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Abongwa, M.; Martin, R.J.; Robertson, A.P. A brief review on the mode of action of antinematodal drugs. Acta Vet. Beogr. 2017, 67, 137–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajíčková, M.; Nguyen, L.T.; Skálová, L.; Stuchlíková, L.R.; Matoušková, P. Anthelmintics in the future: Current trends in the discovery and development of new drugs against gastrointestinal nematodes. Drug Discov. Today 2020, 25, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Fox, N.J.; Marion, G.; Davidson, R.S.; White, P.C.L.; Hutchings, M.R. Climate-driven tipping-points could lead to sudden, high-intensity parasite outbreaks. R. Soc. Open Sci. 2015, 2, 140296. [Google Scholar] [CrossRef] [Green Version]
- Verschave, S.H.; Charlier, J.; Rose, H.; Claerebout, E.; Morgan, E.R. Cattle and nematodes under global change: Transmission models as an ally. Trends Parasitol. 2016, 32, 724–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, E.R.; Aziz, N.A.A.; Blanchard, A.; Charlier, J.; Charvet, C.; Claerebout, E.; Geldhof, P.; Greer, A.W.; Hertzberg, H.; Hodgkinson, J.; et al. 100 questions in livestock helminthology research. Trends Parasitol. 2019, 35, 52–71. [Google Scholar] [CrossRef] [Green Version]
- Genchi, C.; Rinaldi, L.; Mortarino, M.; Genchi, M.; Cringoli, G. Climate and Dirofilaria infection in Europe. Vet. Parasitol. 2009, 163, 286–292. [Google Scholar] [CrossRef]
- McIntyre, K.M.; Setzkorn, C.; Hepworth, P.J.; Morand, S.; Morse, A.P.; Baylis, M. Systematic assessment of the climate sensitivity of important human and domestic animals pathogens in Europe. Sci. Rep. 2017, 7, 7134. [Google Scholar] [CrossRef] [Green Version]
- Scott, I.; Pomroy, W.; Kenyon, P.; Smith, G.; Adlington, B.; Moss, A. Lack of efficacy of monepantel against Teladorsagia circumcincta and Trichostrongylus colubriformis. Vet. Parasitol. 2013, 198, 166–171. [Google Scholar] [CrossRef]
- Brom, R.V.D.; Moll, L.; Kappert, C.; Vellema, P. Haemonchus contortus resistance to monepantel in sheep. Vet. Parasitol. 2015, 209, 278–280. [Google Scholar] [CrossRef]
- Ploeger, H.W.; Everts, R.R. Alarming levels of anthelmintic resistance against gastrointestinal nematodes in sheep in the Netherlands. Vet. Parasitol. 2018, 262, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Holden-Dye, L.; Walker, R.J. Anthelmintic drugs and nematocides: Studies in Caenorhabditis elegans. In WormBook: The Online Review of C. elegans Biology; The C. elegans Research Community, Ed.; WormBook, 2014; Available online: http://www.wormbook.org/chapters/www_anthelminticdrugs.2/anthelminticdrugs.2.html (accessed on 24 June 2020).
- Idris, A.O.; Wintola, O.A.; Afolayan, A.J. Helminthiases; prevalence, transmission, host-parasite interactions, resistance to common synthetic drugs and treatment. Heliyon 2019, 5, e01161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elfawal, M.A.; Savinov, S.N.; Aroian, R.V. Drug screening for discovery of broad-spectrum agents for soil-transmitted nematodes. Sci. Rep. 2019, 9, 12347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekurova, O.N.; Schneider, O.; Zotchev, S.B. Novel bioactive natural products from bacteria via bioprospecting, genome mining and metabolic engineering. Microb. Biotechnol. 2019, 12, 828–844. [Google Scholar] [CrossRef] [Green Version]
- Deng, Q.; Zhou, L.; Luo, M.; Deng, Z.; Zhao, C. Heterologous expression of Avermectins biosynthetic gene cluster by construction of a Bacterial Artificial Chromosome library of the producers. Synth. Syst. Biotechnol. 2017, 2, 59–64. [Google Scholar] [CrossRef]
- Monaghan, R.L.; Tkacz, J.S. Bioactive microbial products: Focus upon mechanism of action. Annu. Rev. Microbiol. 1990, 44, 271–331. [Google Scholar] [CrossRef]
- Giovanelli, F.; Mattellini, M.; Fichi, G.; Flamini, G.; Perrucci, S. In vitro anthelmintic activity of four plant-derived compounds against sheep gastrointestinal nematodes. Vet. Sci. 2018, 5, 78. [Google Scholar] [CrossRef] [Green Version]
- Spiegler, V.; Liebau, E.; Hensel, A. Medicinal plant extracts and plant-derived polyphenols with anthelmintic activity against intestinal nematodes. Nat. Prod. Rep. 2017, 34, 627–643. [Google Scholar] [CrossRef]
- Gaudêncio, S.P.; Pereira, F. Dereplication: Racing to speed up the natural products discovery process. Nat. Prod. Rep. 2015, 32, 779–810. [Google Scholar] [CrossRef]
- Shalaby, H.; Ashry, H.; Saad, M.; Farag, T. In vitro effects of Streptomyces tyrosinase on the egg and dult worm of Toxocara vitulorum. Iran. J. Parasitol. 2020, 15, 67–75. [Google Scholar]
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.-Q.; Wang, J.-F.; Hao, Y.-Y.; Wang, Y. Recent advances in the discovery and development of marine microbial natural products. Mar. Drugs 2013, 11, 700–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, D.-X.; Jiang, Y.-Y.; Zhang, H.-Y. Marine natural products as sources of novel scaffolds: Achievement and concern. Dug Discov. Today 2010, 15, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Costello, M.J.; Cheung, A.; De Hauwere, N. Surface area and the seabed area, volume, depth, slope, and topographic variation for the world’s seas, oceans, and countries. Environ. Sci. Technol. 2010, 44, 8821–8828. [Google Scholar] [CrossRef] [PubMed]
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar] [CrossRef]
- Kanase, H.; Singh, K.N. Marine pharmacology: Potential, challenges, and future in India. J. Med. Sci. 2018, 38, 49–53. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wuertz, S. Bacteria and archaea on earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [Green Version]
- Magnabosco, C.; Lin, L.-H.; Dong, H.; Bomberg, M.; Ghiorse, W.; Stan-Lotter, H.; Pedersen, K.; Kieft, T.L.; Van Heerden, E.; Onstott, T.C. The biomass and biodiversity of the continental subsurface. Nat. Geosci. 2018, 11, 707–717. [Google Scholar] [CrossRef]
- Pascoal, F.; Magalhães, C.; Costa, R. The link between the ecology of the prokaryotic rare biosphere and its biotechnological potential. Front. Microbiol. 2020, 11, 231. [Google Scholar] [CrossRef] [Green Version]
- Blockley, A.; Elliott, D.R.; Roberts, A.P.; Sweet, M.J. Symbiotic microbes from marine invertebrates: Driving a new era of natural product drug discovery. Diversity 2017, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ding, W.; Li, Y.-X.; Tam, C.; Bougouffa, S.; Wang, R.; Pei, B.; Chiang, H.; Leung, P.; Lu, Y.; et al. Marine biofilms constitute a bank of hidden microbial diversity and functional potential. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Conlan, J.V.; Thompson, R.C.A.; Khamlome, B.; Pallant, L.; Fenwick, S.; Elliot, A.; Sripa, B.; Blacksell, S.D.; Vongxay, K. Soil-transmitted helminthiasis in Laos: A community-wide cross-sectional study of humans and dogs in a mass drug administration environment. Am. J. Trop. Med. Hyg. 2012, 86, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.F., Jr.; Hu, Y.; Miller, M.M.; Scheib, U.; Yiu, Y.Y.; Aroian, R.V. Bacillus thuringiensis-derived Cry5B has potent anthelmintic activity against Ascaris suum. PLoS Negl. Trop. Dis. 2013, 7, e2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.-Z.; Hale, K.; Carta, L.; Platzer, E.; Wong, C.; Fang, S.-C.; Aroian, R.V. Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. USA 2003, 100, 2760–2765. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.-Q.; Mo, M.-H.; Zhou, J.-P.; Zou, C.-S.; Zhang, K.-Q. Evaluation and identification of potential organic nematicidal volatiles from soil bacteria. Soil Biol. Biochem. 2007, 39, 2567–2575. [Google Scholar] [CrossRef]
- Burg, R.W.; Miller, B.M.; Baker, E.E.; Birnbaum, J.; Currie, S.A.; Hartman, R.; Kong, Y.-L.; Monaghan, R.L.; Olson, G.; Putter, I.; et al. Avermectins, new family of potent anthelmintic agents: Producing organism and fermentation. Antimicrob. Agents Chemother. 1979, 15, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Omura, S. Ivermectin: 25 years and still going strong. Int. J. Antimicrob. Agents 2008, 31, 91–98. [Google Scholar] [CrossRef]
- Campbell, C.W. History of avermectin and ivermectin, with notes on the history of other macrocyclic lactone antiparasitic agents. Curr. Pharm. Biotechnol. 2012, 13, 853–865. [Google Scholar] [CrossRef]
- Rahul, S.; Chandrashekhar, P.; Hemant, B.; Chandrakant, N.; Laxmikant, S.; Satish, P. Nematicidal activity of microbial pigment from Serratia marcescens. Nat. Prod. Res. 2014, 28, 1399–1404. [Google Scholar] [CrossRef]
- Cezairliyan, B.; Vinayavekhin, N.; Grenfell-Lee, D.; Yuen, G.J.; Saghatelian, A.; Ausubel, F.M. Identification of Pseudomonas aeruginosa Phenazines that kill Caenorhabditis elegans. PLoS Pathog. 2013, 9, e1003101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, A.; Rentas, C.; Caldwell, G.A.; Caldwell, K.A. Phenazine derivatives cause proteotoxicity and stress in C. elegans. Neurosci. Lett. 2015, 584, 23–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.-W.; Mahajan-Miklos, S.; Ausubel, F.M. Killing of Caenorhabditis elegans by Pseudomonas aeruginosa used to model mammalian bacterial pathogenesis. Proc. Natl. Acad. Sci. USA 1999, 96, 715–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Jiang, H.; Cheng, Q.; Chen, J.; Wu, G.; Kumar, A.; Sun, M.; Liu, Z. Enhanced nematicidal potential of the chitinase pachi from Pseudomonas aeruginosa in association with Cry21Aa. Sci. Rep. 2015, 5, 14395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabarinathan, D.; Amirthalingam, M.; Sabapathy, P.C.; Govindan, S.; Palanisamy, S.; Kathirvel, P. Anthelmintic efficacy of glycolipid biosurfactant produced by Pseudomonas plecoglossicida: An insight from mutant and transgenic forms of Caenorhabditis elegans. Biodegradation 2019, 30, 203–214. [Google Scholar] [CrossRef]
- Rao, D.; Webb, J.S.; Kjelleberg, S. Competitive interactions in mixed-species biofilms containing the marine bacterium Pseudoalteromonas tunicata. Appl. Environ. Microbiol. 2005, 71, 1729–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, R.M.; Zaneveld, J.R.; Rosales, S.M.; Payet, J.P.; Burkepile, D.E.; Thurber, R.V. Bacterial predation in a marine host-associated microbiome. ISME J. 2016, 10, 1540–1544. [Google Scholar] [CrossRef]
- de Carvalho, C.C. Marine biofilms: A successful microbial strategy with economic implications. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Chiu, J.M.Y.; Thiyagarajan, V.; Tsoi, M.M.Y.; Qian, P.Y. Qualitative and quantitative changes in marine biofilms as a function of temperature and salinity in summer and winter. Biofilms 2005, 2, 183–195. [Google Scholar] [CrossRef]
- Ortega-Morales, B.O.; Chan-Bacab, M.J.; Rosa-García, S.D.C.D.L.; Camacho-Chab, J.C. Valuable processes and products from marine intertidal microbial communities. Curr. Opin. Biotechnol. 2010, 21, 346–352. [Google Scholar] [CrossRef]
- Antunes, J.; Leão, P.; Vasconcelos, V. Marine biofilms: Diversity of communities and of chemical cues. Environ. Microbiol. Rep. 2019, 11, 287–305. [Google Scholar] [CrossRef]
- Vlamakis, H.; Chai, Y.; Beauregard, P.B.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, S.; Harder, T.; Burke, C.; Steinberg, P.; Kjelleberg, S.; Thomas, T. The seaweed holobiont: Understanding seaweed–bacteria interactions. FEMS Microbiol. Rev. 2013, 37, 462–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katharios-Lanwermeyer, S.; Xi, C.; Jakubovics, N.; Rickard, A.H. Mini-review: Microbial coaggregation: Ubiquity and implications for biofilm development. Biofouling 2014, 30, 1235–1251. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, P.D.; De Nys, R. Chemical mediation of colonisation of seaweed surfaces. J. Phycol. 2002, 38, 621–629. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, E.; Yan, L.; Boyd, K.G.; Wright, P.C.; Burgess, J.G. The symbiotic role of marine microbes on living surfaces. Hydrobiologia 2001, 461, 37–40. [Google Scholar] [CrossRef]
- Haas, A.F.; Wild, C. Composition analysis of organic matter released by cosmopolitan coral reef-associated green algae. Aquat. Biol. 2010, 10, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Thomas, T.; Evans, F.F.; Schleheck, D.; Mai-Prochnow, A.; Burke, C.; Penesyan, A.; Dalisay, D.S.; Stelzer-Braid, S.; Saunders, N.F.W.; Johnson, J.; et al. Analysis of the Pseudoalteromonas tunicata genome reveals properties of a surface-associated life style in the marine environment. PLoS ONE 2008, 3, e3252. [Google Scholar] [CrossRef]
- Matz, C.; Webb, J.S.; Schupp, P.J.; Phang, S.Y.; Penesyan, A.; Egan, S.; Steinberg, P.; Kjelleberg, S. Marine biofilm bacteria evade eukaryotic predation by targeted chemical defense. PLoS ONE 2008, 3, e2744. [Google Scholar] [CrossRef] [Green Version]
- Patterson, D.J.; Lee, W.J. Geographic distribution and diversity of free-living heterotrophic flagellates. In The Flagellates: Unity, Diversity and Evolution; Leadbeater, B.S.C., Green, J.C., Eds.; Taylor & Francis: London, UK, 2000; pp. 277–283. [Google Scholar]
- Hisatugo, K.F.; Mansano, A.S.; Seleghim, M.H. Protozoans bacterivory in a subtropical environment during a dry/cold and a rainy/warm season. Braz. J. Microbiol. 2014, 45, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Corte, D.; Paredes, G.; Yokokawa, T.; Sintes, E.; Herndl, G.J. Differential response of Cafeteria roenbergensis to different bacterial and archaeal prey characteristics. Microb. Ecol. 2019, 78, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Weitere, M.; Erken, M.; Majdi, N.; Arndt, H.; Norf, H.; Reinshagen, M.; Traunspurger, W.; Walterscheid, A.; Wey, J.K. The food web perspective on aquatic biofilms. Ecol. Monogr. 2018, 88, 543–559. [Google Scholar] [CrossRef]
- Moens, T.; Traunspurger, W.; Bergtold, M. Feeding ecology of free-living benthic nematodes. In Freshwater Nematodes. Ecology and Taxonomy; Abebe, E., Traunspurger, W., Andrássy, I., Eds.; CAB International Publishing: Oxfordshire, UK, 2006; pp. 105–131. [Google Scholar]
- Adnan, M.; Alshammari, E.M.; Patel, M.; Ashraf, S.A.; Khan, S.; Hadi, S. Significance and potential of marine microbial natural bioactive compounds against biofilms/biofouling: Necessity for green chemistry. PeerJ 2018, 6, e5049. [Google Scholar] [CrossRef] [PubMed]
- Penesyan, A.; Kjelleberg, S.; Egan, S. Development of novel drugs from marine surface associated microorganisms. Mar. Drugs 2010, 8, 438–459. [Google Scholar] [CrossRef] [Green Version]
- Rocha-Martin, J.; Harrington, C.; Dobson, A.D.W.; O’Gara, F. Emerging strategies and integrated systems microbiology technologies for biodiscovery of marine bioactive compounds. Mar. Drugs 2014, 12, 3516–3559. [Google Scholar] [CrossRef]
- Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association-a review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef] [Green Version]
- Proksch, P.; Edrada, R.A.; Ebel, R. Drugs from the seas—current status and microbiological implications. Appl. Microbiol. Biotechnol. 2002, 59, 125–134. [Google Scholar] [CrossRef]
- Oclarit, J.M.; Okada, H.; Ohta, S.; Kaminura, K.; Yamaoka, Y.; Iizuka, T.; Miyashiro, S.; Ikegami, S. Anti-bacillus substance in the marine sponge, Hyatella species, produced by an associated Vibrio species bacterium. Microbios 1994, 78, 7–16. [Google Scholar]
- Kobayashi, M.; Aoki, S.; Gato, K.; Matsunami, K.; Kurosu, M.; Kitagawa, I. Marine natural products. XXXIV. Trisindoline, a new antibiotic indole trimer, produced by a bacterium of Vibrio sp. separated from the marine sponge Hyrtios altum. Chem. Pharm. Bull. 1994, 42, 2449–2451. [Google Scholar] [CrossRef] [Green Version]
- König, G.M.; Kehraus, S.; Seibert, S.F.; Abdel-Lateff, A.; Müller, D. Natural products from marine organisms and their associated microbes. ChemBioChem 2006, 7, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Rinehart, K.L.; Gloer, J.B.; Hughes, R.G.; Renis, H.E.; McGovren, J.P.; Swynenberg, E.B.; Stringfellow, D.A.; Kuentzel, S.L.; Li, L.H. Didemnins: Antiviral and antitumor depsipeptides from a Caribbean tunicate. Science 1981, 212, 933–935. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Kersten, R.D.; Nam, S.-J.; Lu, L.; Al-Suwailem, A.M.; Zheng, H.; Fenical, W.H.; Dorrestein, P.C.; Moore, B.S.; Qian, P.-Y. Bacterial biosynthesis and maturation of the didemnin anti-cancer agents. J. Am. Chem. Soc. 2012, 134, 8625–8632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukimoto, M.; Nagaoka, M.; Shishido, Y.; Fujimoto, J.; Nishisaka, F.; Matsumoto, S.; Harunari, E.; Imada, C.; Matsuzaki, T. Bacterial production of the tunicate-derived antitumor cyclic depsipeptide didemnin B. J. Nat. Prod. 2011, 74, 2329–2331. [Google Scholar] [CrossRef]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine microorganisms as a promising and sustainable source of bioactive molecules. Mar. Environ. Res. 2017, 128, 58–69. [Google Scholar] [CrossRef]
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Structure and function of the global ocean microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef] [Green Version]
- Venter, J.C.; Remington, K.; Heidelberg, J.F.; Halpern, A.L.; Rusch, D.; Eisen, J.A.; Wu, D.; Paulsen, I.T.; Nelson, K.E.; Nelson, W.; et al. Environmental genome shotgun sequencing of the Sargasso Sea. Science 2004, 304, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Rusch, D.B.; Halpern, A.L.; Sutton, G.; Heidelberg, K.B.; Williamson, S.; Yooseph, S.; Wu, D.; Eisen, J.A.; Hoffman, J.M.; Remington, K.; et al. The Sorcerer II Global Ocean Sampling Expedition: Northwest Atlantic through Eastern Tropical Pacific. PLoS Biol. 2007, 5, e77. [Google Scholar] [CrossRef]
- Ballestriero, F.; Daim, M.; Penesyan, A.; Nappi, J.; Schleheck, D.; Bazzicalupo, P.; Di Schiavi, E.; Egan, S. Antinematode activity of violacein and the role of the insulin/IGF-1 pathway in controlling violacein sensitivity in Caenorhabditis elegans. PLoS ONE 2014, 9, e109201. [Google Scholar] [CrossRef] [Green Version]
- Ballestriero, F.; Thomas, T.; Burke, C.; Egan, S.; Kjelleberg, S. Identification of compounds with bioactivity against the nematode Caenorhabditis elegans by a screen based on the functional genomics of the marine bacterium Pseudoalteromonas tunicata D2. Appl. Environ. Microbiol. 2010, 76, 5710–5717. [Google Scholar] [CrossRef] [Green Version]
- Penesyan, A.; Ballestriero, F.; Daim, M.; Kjelleberg, S.; Thomas, T.; Egan, S. Assessing the effectiveness of functional genetic screens for the identification of bioactive metabolites. Mar. Drugs 2013, 11, 40–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, F.P.; Ingham, C.; Hurtado-Ortiz, R.; Bizet, C.; Tasdemir, D.; De Pascale, D. Isolation by miniaturized culture chip of an Antarctic bacterium Aequorivita sp. with antimicrobial and anthelmintic activity. Biotechnol. Rep. 2018, 20, e00281. [Google Scholar] [CrossRef] [PubMed]
- Nappi, J. Discovery of Novel Bioactive Metabolites from Marine Epiphytic Bacteria and Assessment of Their Ecological Role. Ph.D. Thesis, The University of New South Wales, Sydney, Australia, February 2019. [Google Scholar]
- Yu, J.; Du, G.; Li, R.; Li, L.; Li, Z.; Zhou, C.; Chen, C.; Guo, D. Nematicidal activities of bacterial volatiles and components from two marine bacteria, Pseudoalteromonas marina strain H-42 and Vibrio atlanticus strain S-16, against the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2015, 17, 1011–1025. [Google Scholar] [CrossRef]
- Huang, D.; Yu, C.; Shao, Z.; Cai, M.-M.; Li, G.-Y.; Zheng, L.; Yu, Z.-N.; Zhang, J. Identification and characterization of nematicidal volatile organic compounds from deep-sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neu, A.-K.; Månsson, M.; Gram, L.; Prol-García, M.J. Toxicity of bioactive and probiotic marine bacteria and their secondary metabolites in Artemia sp. and Caenorhabditis elegans as eukaryotic model organisms. Appl. Environ. Microbiol. 2014, 80, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Gram, L.; Melchiorsen, J.; Bruhn, J.B. Antibacterial activity of marine culturable bacteria collected from a global sampling of ocean surface waters and surface swabs of marine organisms. Mar. Biotechnol. 2010, 12, 439–451. [Google Scholar] [CrossRef]
- Wietz, M.; Månsson, M.; Bowman, J.S.; Blom, N.; Ng, Y.; Gram, L. Wide Distribution of closely related, antibiotic-producing Arthrobacter strains throughout the Arctic Ocean. Appl. Environ. Microbiol. 2012, 78, 2039–2042. [Google Scholar] [CrossRef] [Green Version]
- Mathew, M.D.; Mathew, N.D.; Miller, A.; Simpson, M.; Au, V.; Garland, S.; Gestin, M.; Edgley, M.L.; Flibotte, S.; Balgi, A.; et al. Using C. elegans forward and reverse genetics to identify new compounds with anthelmintic activity. PLoS Negl. Trop. Dis. 2016, 10, e0005058. [Google Scholar] [CrossRef]
- Blasco-Costa, I.; Poulin, R. Parasite life-cycle studies: A plea to resurrect an old parasitological tradition. J. Helminthol. 2017, 91, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Burns, A.R.; Luciani, G.M.; Musso, G.; Bagg, R.; Yeo, M.; Zhang, Y.; Rajendran, L.; Glavin, J.; Hunter, R.; Redman, E.; et al. Caenorhabditis elegans is a useful model for anthelmintic discovery. Nat. Commun. 2015, 6, 7485. [Google Scholar] [CrossRef]
- Leung, M.C.K.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Queirós, L.; Pereira, J.; Gonçalves, F.J.M.; Pacheco, M.; Aschner, M.; Pereira, P. Caenorhabditis elegans as a tool for environmental risk assessment: Emerging and promising applications for a “nobelized worm”. Crit. Rev. Toxicol. 2019, 49, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Park, H.; Liu, J.; Sternberg, P.W. An efficient genome editing strategy to generate putative null mutants in Caenorhabditis elegans using CRISPR/Cas9. G3 2018, 8, 3607–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddle, D.L.; Blumenthal, T.; Meyer, B.J.; Priess, J.R. C elegans II, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1997; Section I, The Biological Model. Available online: https://www.ncbi.nlm.nih.gov/books/NBK20086/ (accessed on 5 July 2020).
- Kong, C.; Eng, S.-A.; Lim, M.-P.; Nathan, S. Beyond traditional antimicrobials: A Caenorhabditis elegans model for discovery of novel anti-infectives. Front. Microbiol. 2016, 7, 1956. [Google Scholar] [CrossRef]
- Frézal, L.; Félix, M.-A. The natural history of model organisms: C. elegans outside the Petri dish. eLife 2015, 4, e05849. [Google Scholar] [CrossRef]
- Artal-Sanz, M.; de Jong, L.; Tavernarakis, N. Caenorhabditis elegans: A versatile platform for drug discovery. Biotechnol. J. 2006, 1, 1405–1418. [Google Scholar] [CrossRef]
- Lok, J.B.; Unnasch, T.R. Transgenesis in Animal Parasitic Nematodes: Strongyloides spp. and Brugia spp. In WormBook: The Online Review of C. elegans Biology; The C. elegans Research Community, Ed.; WormBook, 2018. Available online: https://www.ncbi.nlm.nih.gov/books/NBK174830/ (accessed on 23 June 2020).
- Stiernagle, T. Maintenance of C. elegans. In WormBook: The Online Review of C. elegans Biology; The C. elegans Research Community, Ed.; WormBook, 2006; Available online: http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html (accessed on 30 May 2020).
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar]
- Fraietta, I.; Gasparri, F. The development of high-content screening (HCS) technology and its importance to drug discovery. Expert Opin. Drug Discov. 2016, 11, 501–514. [Google Scholar] [CrossRef]
- Midkiff, D.; San-Miguel, A. Microfluidic technologies for high throughput screening through sorting and on-chip culture of C. elegans. Molecules 2019, 24, 4292. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, M.; Dean, E.; Reilly, E.; Bergholz, E.; Chalfie, M. Genetic and molecular analysis of a Caenorhabditis elegans beta-tubulin that conveys benzimidazole sensitivity. J. Cell Biol. 1989, 109, 2993–3003. [Google Scholar] [CrossRef] [Green Version]
- Haber, C.L.; Heckaman, C.L.; Li, G.P.; Thompson, D.P.; Whaley, H.A.; Wiley, V.H. Development of a mechanism of action-based screen for anthelmintic microbial metabolites with avermectin like activity and isolation of milbemycin-producing Streptomyces strains. Antimicrob. Agents Chemother. 1991, 35, 1811–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marroquin, L.D.; Elyassnia, D.; Griffitts, J.S.; Feitelson, J.S.; Aroian, R.V. Bacillus thuringiensis (Bt) toxin susceptibility and isolation of resistance mutants in the nematode Caenorhabditis elegans. Genetics 2000, 155, 1693–1699. [Google Scholar]
- Mitreva, M.; McCarter, J.P.; Martin, J.; Dante, M.; Wylie, T.; Chiapelli, B.; Pape, D.; Clifton, S.W.; Nutman, T.B.; Waterston, R.H. Comparative genomics of gene expression in the parasitic and free-living nematodes Strongyloides stercoralis and Caenorhabditis elegans. Genome Res. 2004, 14, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumarasingha, R.; Young, N.D.; Manurung, R.; Lim, D.S.L.; Tu, C.-L.; Palombo, E.A.; Shaw, J.M.; Gasser, R.B.; Boag, P.R. Transcriptional alterations in Caenorhabditis elegans following exposure to an anthelmintic fraction of the plant Picria fel-terrae Lour. Parasites Vectors 2019, 12, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, D.A.; Balamurugan, K. Global proteomic response of Caenorhabditis elegans against PemKSa toxin. Front. Cell. Infect. Microbiol. 2019, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Angstadt, J.D.; Donmoyer, J.E.; Stretton, A.O. Retrovesicular ganglion of the nematode Ascaris. J. Comp. Neurol. 1989, 284, 374–388. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.D.; Stretton, A.O. Localization of choline acetyltransferase within identified motoneurons of the nematode Ascaris. J. Neurosci. 1985, 5, 1984–1992. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.D.; Stretton, A.O. GABA-immunoreactivity in inhibitory motor neurons of the nematode Ascaris. J. Neurosci. 1987, 7, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Davis, R. Neurophysiology of glutamatergic signalling and anthelmintic action in Ascaris suum: Pharmacological evidence for a kainate receptor. Parasitology 1998, 116, 471–486. [Google Scholar] [CrossRef]
- Weeks, J.C.; Robinson, K.J.; Lockery, S.R.; Roberts, W.M. Anthelmintic drug actions in resistant and susceptible C. elegans revealed by electrophysiological recordings in a multichannel microfluidic device. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 607–628. [Google Scholar] [CrossRef]
- Lewis, J.; Wu, C.-H.; Levine, J.; Berg, H. Levamisole-resitant mutants of the nematode Caenorhabditis elegans appear to lack pharmacological acetylcholine receptors. Neuroscience 1980, 5, 967–989. [Google Scholar] [CrossRef]
- Kaminsky, R.; Ducray, P.; Jung, M.; Clover, R.; Rufener, L.; Bouvier, J.; Weber, S.S.; Wenger, A.; Wieland-Berghausen, S.; Goebel, T.; et al. A new class of anthelmintics effective against drug-resistant nematodes. Nature 2008, 452, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, M.; Okuyama, E.; Kobayashi, M.; Inoue, H. The structure of paraherquamide, a toxic metabolite from Penicillium paraherquei. Tetrahedron Lett. 1981, 22, 135–136. [Google Scholar] [CrossRef]
- Hernando, G.; Turani, O.; Bouzat, C. Caenorhabditis elegans muscle Cys-loop receptors as novel targets of terpenoids with potential anthelmintic activity. PLoS Negl. Trop. Dis. 2019, 13, e0007895. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, J.M.; Blizzard, T.A.; Ondeyka, J.; Goegelman, R.; Sinclair, P.J.; Mrozik, H. [3H]Paraherquamide binding to Caenorhabditis elegans: Studies on a potent new anthelmintic agent. Biochem. Pharmacol. 1992, 43, 679–684. [Google Scholar] [CrossRef]
- Ruiz-Lancheros, E.; Viau, C.; Walter, T.N.; Francis, A.; Geary, T. Activity of novel nicotinic anthelmintics in cut preparations of Caenorhabditis elegans. Int. J. Parasitol. 2011, 41, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z. The modification of natural products for medical use. Acta Pharm. Sin. B 2017, 7, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Taman, A.; Azab, M. Present-day anthelmintics and perspectives on future new targets. Parasitol. Res. 2014, 113, 2425–2433. [Google Scholar] [CrossRef]
Figure 1.
Surface-associated marine bacteria live on the nutrient-rich marine surfaces (for example macroalgae or cnidaria) in the form of biofilms. These marine biofilms are exposed to biotic (intra- and/or inter-species interaction with other microorganisms or predators i.e., protozoa and nematodes) and abiotic physical–chemical stressors. The predator–prey interactions lead to the production of nematicidal metabolites by the surface-associated marine bacteria, while the harsh environmental conditions enhance the chemical, molecular and functional properties of the produced microbial compounds. Image created with BioRender.com.
Figure 1.
Surface-associated marine bacteria live on the nutrient-rich marine surfaces (for example macroalgae or cnidaria) in the form of biofilms. These marine biofilms are exposed to biotic (intra- and/or inter-species interaction with other microorganisms or predators i.e., protozoa and nematodes) and abiotic physical–chemical stressors. The predator–prey interactions lead to the production of nematicidal metabolites by the surface-associated marine bacteria, while the harsh environmental conditions enhance the chemical, molecular and functional properties of the produced microbial compounds. Image created with BioRender.com.

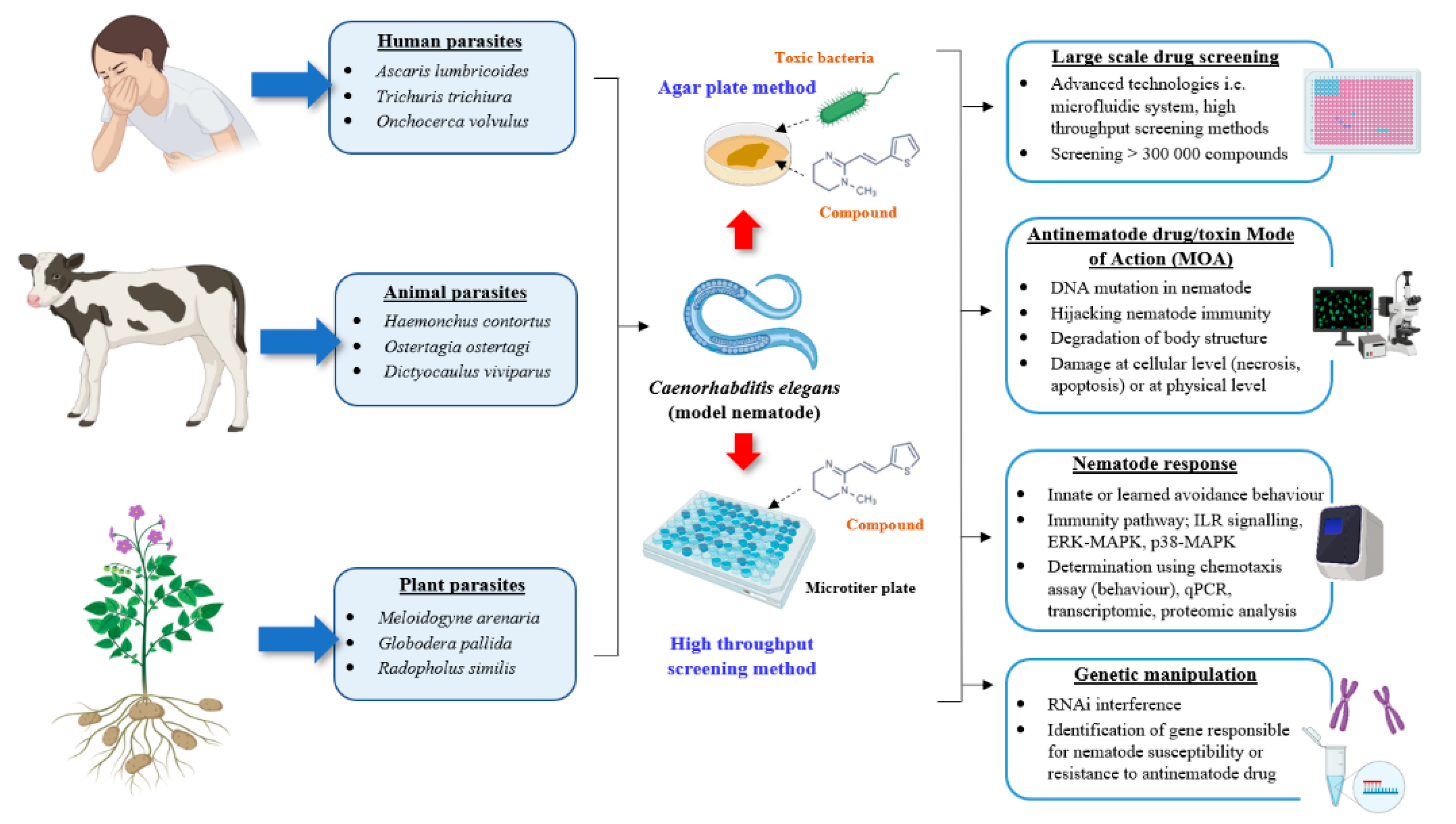
Figure 2.
Schematic diagram representing the functional role of C. elegans as a model nematode for the development of antinematode drugs. The propagation and handling methods of human, animal or plant-parasitic nematodes in the laboratory are challenging. C. elegans can be used as a surrogate nematode for the initial screening against the potential compounds or microorganisms with nematotoxic properties either via the conservative agar plate method or through high throughput screening technologies. C. elegans-based research offers several advantages including nematode genetic manipulation, determination of drug MOA, evaluation of the resulting nematode responses and a large-scale initial drug screening due to easy nematode maintenance and propagation in the laboratory. Image created with BioRender.com.
Figure 2.
Schematic diagram representing the functional role of C. elegans as a model nematode for the development of antinematode drugs. The propagation and handling methods of human, animal or plant-parasitic nematodes in the laboratory are challenging. C. elegans can be used as a surrogate nematode for the initial screening against the potential compounds or microorganisms with nematotoxic properties either via the conservative agar plate method or through high throughput screening technologies. C. elegans-based research offers several advantages including nematode genetic manipulation, determination of drug MOA, evaluation of the resulting nematode responses and a large-scale initial drug screening due to easy nematode maintenance and propagation in the laboratory. Image created with BioRender.com.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Salikin, N.H.; Nappi, J.; Majzoub, M.E.; Egan, S. Combating Parasitic Nematode Infections, Newly Discovered Antinematode Compounds from Marine Epiphytic Bacteria. Microorganisms 2020, 8, 1963. https://doi.org/10.3390/microorganisms8121963
AMA Style
Salikin NH, Nappi J, Majzoub ME, Egan S. Combating Parasitic Nematode Infections, Newly Discovered Antinematode Compounds from Marine Epiphytic Bacteria. Microorganisms. 2020; 8(12):1963. https://doi.org/10.3390/microorganisms8121963
Chicago/Turabian StyleSalikin, Nor Hawani, Jadranka Nappi, Marwan E. Majzoub, and Suhelen Egan. 2020. "Combating Parasitic Nematode Infections, Newly Discovered Antinematode Compounds from Marine Epiphytic Bacteria" Microorganisms 8, no. 12: 1963. https://doi.org/10.3390/microorganisms8121963
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.