Microbial Resources as a Tool for Enhancing Sustainability in Winemaking


CREA—Council for Agricultural Research and Economics, Research Centre for Viticulture and Enology, Viale XXVIII Aprile 26, 31015 Conegliano, Italy
Microorganisms 2020, 8(4), 507; https://doi.org/10.3390/microorganisms8040507
Submission received: 28 February 2020
/
Revised: 30 March 2020
/
Accepted: 31 March 2020
/
Published: 2 April 2020
(This article belongs to the Special Issue Microbial Dynamics in Wine Production)
Abstract
:In agriculture, the wine sector is one of the industries most affected by the sustainability issue. It is responsible for about 0.3% of annual global greenhouse gas emissions from anthropogenic activities. Sustainability in vitiviniculture was firstly linked to vineyard management, where the use of fertilizers, pesticides and heavy metals is a major concern. More recently, the contribution of winemaking, from grape harvest to bottling, has also been considered. Several cellar processes could be improved for reducing the environmental impact of the whole chain, including microbe-driven transformations. This paper reviews the potential of microorganisms and interactions thereof as a natural, environmentally friendly tool to improve the sustainability aspects of winemaking, all along the production chain. The main phases identified as potentially interesting for exploiting microbial activities to lower inputs are: (i) pre-fermentative stages, (ii) alcoholic fermentation, (iii) stage between alcoholic and malolactic fermentation, (iv) malolactic fermentation, (v) stabilization and spoilage risk management, and (vi) by-products and wastewater treatment. The presence of proper yeast or bacterial strains, the management and timing of inoculation of starter cultures, and some appropriate technological modifications that favor selected microbial activities can lead to several positive effects, including (among other) energy savings, reduction of chemical additives such as sulfites, and reuse of certain residues.
{kind=link}
{kind=link}
1. Introduction
In agriculture, the wine sector is one of the industries most affected by the environmental sustainability issue. On a global scale, it is responsible for around 0.3% of annual global greenhouse gas (GHG) emissions from anthropogenic activities; this corresponds to about 2% of agriculture contribution, which in turn is estimated to contribute to 14% of the total anthropogenic-derived GHG emissions [1,2].
The mounting interest regarding the environmental impact of wine production and its potential to modify regional climate patterns has prompted many wine producers to move toward sustainable grape growing and wine production practices. Moreover, recent analyses of consumer perceptions, preferences, and willingness to pay for wine showed that producing and marketing wine with sustainability features is a promising strategy for quality differentiation, providing an additional stimulus for the wine industry to proceed toward a larger adoption of sustainable practices [3]. Therefore, improvements in terms of energy and water consumption, use of pesticides and additives, and the polluting effects that these inputs may have on the biosphere, together with their potential concern for consumer acceptance, have prompted recent research activities.
The exploitation of microbial resources to improve sustainability of the winemaking process, nonetheless, is a very recent approach and only a few research studies have addressed it. Sustainability in vitiviniculture was first linked to vineyard management, where crucial issues are related to environmental emissions arising from the use of fertilizers, pesticides, and heavy metals [4,5]. Afterward, some research surveys performed under EU programs and via industry audits [1,4,5] assessed the environmental impact of each input used all along the wine production chain, including winemaking phases. Consequently, several programs for wine life cycle assessment (including initiatives following the European Regulation EMAS [6]) accounted, among other factors, equivalent emissions for electricity consumption and for sulfur dioxide used in the vinification phase [7], which are in turn influenced by microbial transformations and their management.
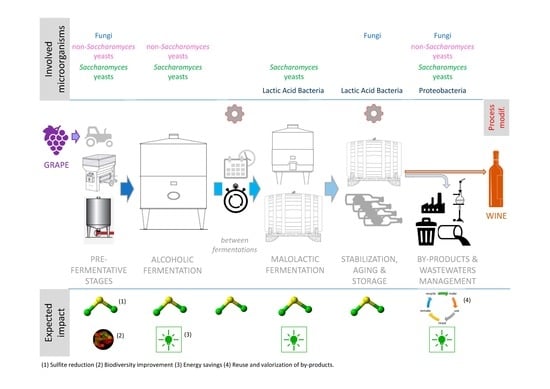
In this regard, one of the aspects of a two-way relationship between microorganisms and climate change in winemaking has been recently addressed and reviewed: the microbial potential as an adaptation strategy for the effects of climate change [8,9,10]. The abovementioned papers described the potential of pro-technological microbes as agents capable of mitigating the negative features of the evolving climatic influence (e.g., microbial solutions to reduce ethanol content, improve organic acids content, and reduce pH). However, the microbial potential for reducing the environmental impact of winemaking (and therefore for lowering the contribution of wine production to climate change) has not been comprehensively described to date. The aim of this paper is to review the potential of microorganisms and interactions thereof as a natural, environmentally friendly tool to improve the sustainability aspects of winemaking, all along the production chain, in different phases. Microorganisms play a role in several steps of the winemaking process, from grape harvest to bottling and waste treatment, and most of these steps can be improved for reducing the environmental impact of the whole chain, including fermentation stages. In this context, the exploitation of microbial resources and the best management of their interactions will be considered for their potential effects on the environmental impact of winemaking, including (i) limiting the use of chemical additives such as sulfites used as preservatives against microbial spoilage, taking advantage of beneficial microorganisms’ activities; (ii) allowing a better energy management and achieving energy savings during fermentations; (iii) avoiding additional inputs such as filtrations, cellulose-based adjuvants, additional warming and cooling steps, and other technological corrections that are eventually used in case of stuck or sluggish fermentations; (iv) improving, in certain cases, microbial biodiversity; and (v) valorizing and/or reducing the environmental impact of wine industry by-products and wastewater. Special attention will be paid to microbial resources and processes which are already, or about to become, available for the winemaking sector at the industrial scale.
2. Improving Sustainability in Pre-Fermentative Stages
Grape transport after harvest and grape crushing are typically the first steps where juice is released and therefore, sulfite is usually added for its antioxidant, antioxidasic, and antimicrobial properties [11]. The antimicrobial effects reduce the activity of “apiculated” yeasts and some bacteria that are present on the grapes and can develop in the cellar, causing off-flavors and defects either before or after fermentations [11,12,13,14]. In this context, the early inoculation of grapes or must with different yeasts has developed considerably in recent years in order to bioprotect the must by directly colonizing the environment and preventing the development of spoilage microorganisms. The industrial objective is to reduce the dose of sulfites and to substitute their effect as much as possible [15,16,17,18], as displayed in Figure 1. Early yeast inoculation, which is also recommended by the International Organisation of Vine and Wine (OIV) as an useful practice to achieve this goal [19], was first intended for moving up the timing of Saccharomyces addition for launching alcoholic fermentation (AF) [20]. More recently, the early inoculation of yeasts belonging to the heterogeneous group of non-Saccharomyces genera is also becoming a common and innovative practice. Indeed, not only the role of non-Saccharomyces yeasts in winemaking has been re-evaluated for their benefits on the quality and sensory properties of wine (reviewed in [21,22,23,24,25]), but also for the additional advantage they provide for must bioprotection by directly colonizing the environment. Even though these yeasts do not necessarily play a role in sugar fermentation, they contribute to preventing the development of undesired microorganisms [15,16,18]. It is worth to note that limiting sulfite addition at this stage can, at times, generate a double advantage: it can indirectly have a positive effect on the production of SO2 by Saccharomyces yeasts that will later on ferment the sugars. It is known that certain enological strains can overproduce this molecule in the presence of high starting concentrations in the must [26].
This approach can, in certain cases, also increase microbial biodiversity. The use of a wide range of yeasts (non-Saccharomyces and Saccharomyces) and lactic acid bacteria (LAB) as starters can tackle the problem of uniformity caused by the use of a few active dry yeast (ADY) strains when this practice was first introduced (in the 1960s) [13]. Some winemakers have tried to increase the influence of the native yeasts by delaying or reducing the use of starter cultures. However, this can lead to uncontrolled fermentations [14]. The noncontrolled fermentations might lead to economic losses due to the risk of spoilage and are hardly compatible with sustainable sulfite management for wine stabilization. The alternative proposed by a recent approach is to incorporate the features of “native” microorganisms to starter cultures: the use of a wide array of different cultures aims at reproducing the vineyard natural microbiota, exploiting different strains as well as different species [14,15]. As the range of available microorganisms increases over time, this gives winemakers the possibility to increase biodiversity also in guided fermentations [16], as reported in Figure 1. The selection of local or autochthonous strains to be included in the starter cultures would further enhance this possibility [17], meanwhile aiming at bioprotecting the must [27].
Focusing on non-Saccharomyces yeasts, the goal of grape and must microbial bioprotection can be achieved through the exploitation of different properties thereof, including (i) specific bioprotective features conferred to certain species and strains by their ability to produce molecules with antimicrobic activities, such as killer toxins or pigments with antifungal action and (ii) more generally, an effective competition for nutrients with natural microbiota that prevents the development of other microorganisms, including possible spoilers. Moreover, a side effect that also contributes to sulfite substitution is potential protection against oxidation; this could be exerted by selected non-Saccharomyces yeasts due to their rapid consumption of oxygen, which prevents oxygen utilization by oxidative yeasts [16].
2.1. Non-Saccharomyces Yeasts Producing Antimicrobial Compounds
One of the most interesting, studied and discussed yeasts to be inoculated in early stages of winemaking (on grapes or on must in pre-fermentative stages or at the beginning of alcoholic fermentation, a few hours before a Saccharomyces strain) is the genus Metschnikowia, with its species Metschnikowia pulcherrima and Metschnikowia fructicola.
Metschnikowia pulcherrima can be used as a biological control agent due to its ability to produce natural antimicrobial compounds, namely pulcherrimin, an insoluble red pigment with antifungal activity. This peculiar antimicrobial activity is produced by the depletion of iron in the medium through the precipitation of iron(III) ions caused by interaction with pulcherriminic acid, a precursor of pulcherrimin secreted by M. pulcherrima [28]. In this way, the environment becomes inhospitable to other microorganisms that require iron for their development. Pulcherrimin has shown effective inhibitory activity against several yeasts: Candida tropicalis, Candida glabrata, and Candida albicans, as well as yeasts belonging to Dekkera/Brettanomyces, Hanseniaspora, and Pichia genera [28,29,30]; and fungi: Botrytis cinerea, as well as Penicillium, Alternaria, Fusarium, Rhizopus, Verticillium, and Monilia spp. [30]. Metschnikowia pulcherrima has therefore been described as a biofungicide capable of effectively reducing the incidence of Botrytis development in post-harvest fruits, such as apples [31,32,33], citrus [34], and cherry [35]. In most cases, its antagonistic mechanism is completed by its competition for nutrients [28]. Some research papers have proposed the application of Metschnikowia pulcherrima on grape berries [36,37,38], since S. cerevisiae seems not to be affected by M. pulcherrima antimicrobial activity [29], so the use of this yeast as a selected starter at any stage prior to Saccharomyces inoculation is compatible with alcoholic fermentation [28]. To our knowledge, the most industrially relevant enological applications of this species are proposed by some companies for protecting must and controlling the indigenous microbiota (pre-fermentative control and bioprotection), either alone or in association with Torulaspora delbrueckii [39], with early-stage inoculation (grape bunches before crushing and grape must after crushing). Moreover, its application in pre-fermentative stages (namely, pre-fermentative cold maceration, PCM) has been tested in sulfite-free must at the winery level and also compared with strains of Metschnikowia fructicola [40,41]. Although these studies were set up for elucidating the positive effects of Metschnikowia on chromatic and aromatic wine characteristics, the results also indirectly confirmed the potential of this species for bioprotection purposes in early winemaking stages, by showing good implantation and population dynamics in PCM.
Besides Metschnikowia, other yeast species have a broad killer spectrum against spoilage yeasts, including, among others, Wickerhamomyces anomalus (formerly Pichia anomala), Kluyeromyces wickerhamii, and Torulaspora delbrueckii [42,43,44]. After its initial discovery in S. cerevisiae, the killer phenotype was described in non-Saccharomyces yeasts [45] and consequently, later on, killer toxins have been proposed as a biocontrol strategy alternative to the use of chemical preservatives or physical methodologies during the winemaking process [43,46]. Killer toxins are generally defined as antimicrobial proteinaceous compounds that inhibit susceptible yeast species or strains, although they remain immune to their own toxins [46]. Although killer proteins have been reported as antimicrobial agents against diverse undesired microorganisms present in different foods [43], they are mainly tested against the prevailing wine spoilage microorganisms Dekkera/Brettanomyces in the enological environment [17,46]. Despite this wide diversity, the killing action of all of the characterized killer toxins is generally mediated by a two-step mechanism, where cell wall is generally the primary site of action. Therefore, cell wall components such as β-1,3-D-glucans and β-1,6-D-glucans are common receptors for the majority of killer toxins characterized to date, although mannoproteins and chitin also serve as first receptors for a number of killer toxins [43]. Recent studies have focused on Kwkt and Pikt, zymocins produced by Kluyeromyces wickerhamii and Wickerhamomyces anomalus, respectively, with antimicrobial properties [47], and on TdKT produced by T. delbrueckii [48]. Recently, the killer toxin KTCf20 secreted by the strain W. anomalus Cf20 was also suggested to bind to β-1,3 and β-1,6 glucans of the cell wall of sensitive strains [42]. Interestingly, these killer toxins are not affected by the pH, temperature, and ethanol concentrations that are typical of winemaking conditions. Furthermore, they do not inhibit the fermenting S. cerevisiae strains or the lactic acid bacteria and are therefore hypothesized not to have a negative impact on alcoholic and malolactic fermentation [43]. Thus, it was hypothesized that the use of W. anomalus starter cultures can partially replace SO2 during grape must fermentation, in order to reduce the wine sulfite content; antimicrobial activity was also reported toward other minor yeast species present during the early stages of grape fermentation, such as Pichia guilliermondii or Pichia membranifaciens [17,42]. Moreover the potential use of the purified toxin Pikt from W. anomalus D2 as an alternative to sulfur dioxide (SO2) has been proposed because Pikt, unlike SO2, produced irreversible damage on sensitive yeasts, ensuring the complete control of spoilage Brettanomyces yeasts [42,47,49].
Nevertheless, it is worth to note that none of these microbe-based innovations (neither killer yeast starter cultures nor purified toxins thereof) are currently used in winemaking at the industrial scale. Indeed, to our knowledge, there are no suppliers in the enology sector proposing these solutions anymore, although for a while, a company used to sell W. anomalus and K. wickerhamii starter cultures in fresh cream [39,50]. Several reasons may explain this situation, including cost-effectiveness issues. Beyond their undoubtful antimicrobial activity, protective cultures/toxins should be produced at moderate costs in order to make them available (in adequate concentrations for industrial winemaking) at affordable prices for wineries.
2.2. Non-Saccharomyces Yeasts Exerting Indirect Bioprotective Effect
Besides specific antimicrobial activities, biocontrol strategies are based on the activity of living microorganisms that counteract the evolution and harmful effects of spoilage microorganisms, without interfering with the life cycle of useful microorganisms or creating risks for human health. Concerning grape bunches and berries, some biocontrol agents, including Aureobasidium pullulans, Metschnikowia pulcherrima, and Pichia guilliermondii, have been proposed against Botrytis cinerea to protect fresh fruit and table or wine grapes [32,37,38,51]. Moreover, some authors examined the microbiota associated with dried grapes in traditional wine production as a source of biocontrol agents against B. cinerea and found interesting activities in several other genera, including Hanseniaspora, Cryptococcus, and Issatchenkia [52], together with epiphytic bacteria mainly belonging to the Bacillus taxon [53]. In another recent work, fermenting must obtained from overripe grape berries and therefore more susceptible to fungal infection was considered for the selection of yeasts with antifungal activity. Promising antifungal activity against B. cinerea was demonstrated in Starmerella bacillaris species and the production of volatile organic compounds (VOCs), tested in vitro, was found to be mainly responsible for the observed effects [54].
To our knowledge, the most industrially relevant enological non-Saccharomyces yeast used in this context is a strain of Metschnikowia fructicola, a species also known for its post-harvest biocontrol potential on other fruits [55,56]. In particular, the abovementioned strain is currently being considered for withering of grapes [57], in order to biocontrol Botrytis infection during the natural drying process of grape bunches for “passito” wine production. Since S. cerevisiae seems not to be affected by M. fructicola presence, the use of the latter as a selected starter in all the pre-fermentative stages, including cold macerations, has gained great interest in modern enology, both for the many benefits on wine quality associated with this yeast and for the possibility of working with low sulfite doses [40,41].
During the crushing stage, the strategy to bioprotect must as soon as possible at the biochemical level, avoiding undesired microorganisms’ metabolism, by inoculating selected starter cultures is increasingly being employed [18]. Indeed, it has been shown that some non-Saccharomyces species support or inhibit the growth of other non-Saccharomyces and Saccharomyces species in multispecies consortiums, and that the relative performance of each yeast species is dependent on its fermentation capacity, initial cell density, and ecological interactions as well as tolerance to environmental factors [58]. A large number of research work has addressed this topic in recent years, most of which focused on the limitation of spoilage occurrence, mainly due to Brettanomyces bruxellensis growth (for reviews, see [17,39,59]). Narrowing the field to those studies that tested strains and strategies at the winemaking scale (pilot or industrial), interesting results about yeast inoculation were observed from a trial aimed at understanding the impact of a Torulaspora delbrueckii strain used as a bioprotective agent instead of sulfite addition. The authors demonstrated the effects of the T. delbrueckii strain, inoculated at the beginning of the white winemaking process, in two Burgundian wineries and proposed it as an alternative to the use of sulfites [16]. A further improvement of this strategy is the use of both non-Saccharomyces yeasts and lactic acid bacteria for obtaining must and minimizing the sulfur dose [60,61], as recently shown in a study that reported the sequential inoculation of Lactobacillus plantarum and Lachancea thermotolerans as a promising winemaking alternative in contrast to traditional vinification and also showed the advantage of producing wines with higher titratable acidity and lower pH [18].
The indirect antagonistic activity of selected non-Saccharomyces yeasts on undesired wine yeast species is considered to be a key property for many starter cultures that display possible biocontrol applications, but are also interesting for the winemaker as tools, for improving the sensory properties of the wine, that are susceptible to modulate the sensory profile/volatile aroma composition and/or exploitable to modify acidity or color in wine [8,39,41,60]. Therefore, the potential application of starter cultures of species which harbor bioprotective potential (either direct or indirect) coupled with other enological features seems to be wider than the application of species that do not bring any technological/sensory property other than bioprotection through specific antimicrobial mechanisms (e.g., killer toxins).
3. Managing Alcoholic Fermentation for Sustainability
The use of selected starter cultures to “guide” grape must fermentations was first introduced by inoculating S. cerevisiae yeasts with the aim of securing a rapid and reliable fermentation process, avoiding organoleptic defects and obtaining wines with desired quality and sensory characteristics [20]. The different potentialities of this step for improving sustainability are described below and summarized in Figure 1.
3.1. Control of Microbial Spoilage during Alcoholic Fermentation
Concerning the impact of alcoholic fermentation (AF) management on sustainability, that is, on the reduction of sulfites and other interventions linked to the risk of microbial spoilage, important guidelines have been provided by the OIV, in 2014, within the resolution “Code of good vitivinicultural practices in order to avoid or limit contamination by Brettanomyces” [62]. The document explains that Brettanomyces can grow as AF slows down or stops and therefore, enological practices commonly recommended for the management of alcoholic fermentation must be implemented, including inoculation of must with selected yeasts that help to achieve a more reliable AF. The fact that the environment becomes more favorable to the multiplication of Brettanomyces if alcoholic fermentation slows down or stops is pointed out, and in the case of the latter, using a process to restart alcoholic fermentation as soon as possible is recommended (as endorsed also in reference scientific studies) [63]. Finally, the OIV document recalls that residual sugars (mainly glucose and fructose) are substrates for Brettanomyces growth [62] and therefore, their leftover should be avoided by carefully monitoring the completion of AF. All the above mentioned goals can be achieved through an effective management of AF, which is usually pursued by taking advantage of a fruitful inoculation of Saccharomyces starters [61,64] and with careful management of nutrient supplementation, including both nitrogen and lipid-based molecules [65,66].
Besides the inoculation of a robust strain able to consume sugars, performed early enough to avoid the development of undesired indigenous microorganisms, the choice of a Saccharomyces starter strain can bring along some other features useful for reducing the final amount of sulfites in wine. The proper choice of wine yeasts and bacteria is a key factor in determining the final levels of both SO2 and acetaldehyde produced [26,67,68]. Acetaldehyde is a key component of wine, formed by yeasts during alcoholic fermentation, that can bind with SO2, and since wines with high levels of acetaldehyde require more exogenous SO2, this can be a concern [69]. At the same time, Saccharomyces yeasts, including commercial starters, widely differ for SO2 production: the production of sulfites by wine yeasts is highly strain-dependent, and despite strong selective processes, some commercial yeast still produce high amounts of these sulfur compounds in some circumstances [67]. Therefore, a correct and careful choice of the Saccharomyces strain for alcoholic fermentation is a key issue for managing acetaldehyde and SO2 production during the winemaking process [70,71,72]. Wine yeasts selected or breaded for having low SO2 and acetaldehyde production are, in this context, a valuable tool for achieving the abovementioned goal [73]. Besides Saccharomyces wine yeasts, selected strains of non-Saccharomyces species can represent a further improvement: in a recent work, Starmerella bacillaris, used in sequential inoculation with S. cerevisiae, produced less SO2 and acetaldehyde compared with Lachancea thermotolerans and Metschnikowia spp. or with S. cerevisiae alone [74].
Moreover, the production of sulfites depends on environmental factors, including the concentration of nutrients in the media, in particular, nitrogen-containing compounds (ammonium, amino acids and especially sulfur-containing amino acids) [75], and on starting sulfite levels [26,75]; therefore, the management of alcoholic fermentation (including the choice of yeast nutrients) might be a tool for modulating SO2 production by yeasts.
Additionally, the careful choice of yeast strain and nutrition protocol will also bring benefits for winemaking stages, mainly malolactic fermentation. Low-SO2-producing strains usually do not inhibit malolactic fermentation (MLF) and therefore, favorize further stages (see par. 5).
3.2. Energy Savings Associated with Alcoholic Fermentation
Literature has extensively described the effect of temperature on yeast metabolism during wine fermentation [76]. As shown in the last decade [76,77,78], the effect of low temperature on fermentation efficiency and aroma production varies markedly for different S. cerevisiae strains.
Certainly, temperature control during fermentation significantly impacts the energy demand of wineries. The majority of the electricity used by wineries (about 90%) is consumed by refrigeration systems for process cooling, that is, fermentation control, cold stabilization, and cold storage [79,80]. The fermentation process takes place at a controlled temperature for quality purposes, to which the wine needs to be cooled at the beginning of fermentation and throughout the process; and the fermentation reaction also generates heat that needs to be removed [79]. As expressed before, the increased interest of consumers in the environmental aspects of winemaking, combined with economic pressure, compels winemakers to address concerns over energy consumption during wine production and to identify potential energy savings. Moreover, the gained awareness among retailers and distribution chains will drive wine suppliers to provide quantitative information on their energy saving solutions and their impact on the environment thereof [81,82,83]. As a consequence, recent research studies addressed the quantification of required heat dissipation during AF, showing interesting results [84,85].
In an initial work, a newly selected Saccharomyces wine strain was tested in the production of sparkling base wine, fermented at a temperature higher than the winery standard. The quantification of electric energy consumption and estimation of energy conservation showed that increasing the temperature from 15 °C to 19 °C during the fermentation process yielded an energy saving of ~65% [84]. No significant differences were found in the main chemical wine parameters and sensory characteristics (through a triangular panel test) between the two temperatures. This was consistent with volatile compound quantitation, as only 25% of the tested aromatic molecules showed a change in concentration with the fermentation temperature (most of them were higher at 19 °C than at 15 °C), and many were below the sensory threshold. Moreover, no measurable SO2 was produced by the yeast (low-producer strain) in any of the fermentations, confirming that temperature did not affect sulfite production for this specific strain at the tested conditions. This study was the first to quantify energy conservation from sustainable temperature management during base wine fermentation, showing the benefits of such an approach [84].
In a more recent work, required heat dissipation was measured in Riesling fermentation and the results confirmed and further illustrated the relevance of the temperature program employed with regard to energy demand for cooling [85]. Approximately 70% less heat had to be dissipated for fermentation at 19 °C, compared with that for fermentation at 14 °C. Approximately 30% less heat had to be dissipated under a 16–11–17 °C temperature program, compared with that for fermentation at 14 °C. Thus, high savings in electrical energy can be expected, although depending on the technical configuration of the cooling system. The formation of most esters was more pronounced in the second half of fermentation at higher temperature. No difference was found in the final concentration of acetate esters or acetic acid. Acetaldehyde concentration was 35% lower for fermentations at 19 °C, compared with those at 14°C. A descriptive analysis, at 5 and 11 months after bottling, revealed no significant difference in wine sensory profiles.
Overall, these studies, carried out with different selected yeast strains, show that energy savings can be achieved by reducing the required dissipated heat through temperature management of fermentations, without compromising wine composition. Yeast characteristics and expected aromatic profile should be carefully considered as the strain choice criteria, when deciding on temperature management and related saving potential. Moreover, in the reported studies, the use of innovative thermal protocols allowed the wineries to adopt more sustainable winemaking processes with low SO2 [84] and acetaldehyde [85] production together with low energy consumption, and consequently, to propose ecolabeling strategies and price premium policies that presently have marketing benefits [3].
4. Improving Sustainability between Alcoholic and Malolactic Fermentation
In modern winemaking, the timing of occurrence of malolactic fermentation (MLF, traditionally happens during storage) is advancing, taking place right after AF or even during AF when co-inoculated is used. This is due to climate change, which determines modifications in grape/must composition such as pH increase, as well as an enlarged need to better manage this step and avoid risky situations in the time frame between AF and MLF [8,61]. Indeed, wines waiting for MLF cannot be stabilized and, in some situations, need to be warmed to favorize bacterial development, but these situations also favor the growth of microbial spoilers. As such, early MLF management has been recommended by the OIV as a good winemaking practice to avoid wine spoilage that causes major economic losses [19] (outlined in Figure 1).
Indeed, MLF management strongly affects the development of spoilers, mainly Brettanomyces, during subsequent wine aging. Some studies showed that wines that underwent rapid MLF inhibited the growth of Brettanomyces, resulting in a product containing little or no volatile phenols. Conversely, wines that did not undergo MLF or underwent late spontaneous MLF that proceeded slowly allowed the proliferation of Brettanomyces, resulting in a product containing more volatile phenols [86,87]. The abovementioned OIV resolution [19] attests that if MLF is delayed, the risk of production of volatile phenols increases because Brettanomyces can take advantage of the time between alcoholic and malolactic fermentations to multiply, benefiting from the absence of SO2. Thus, the use of malolactic starters is proposed as a good way to limit Brettanomyces development and volatile phenol production. Co-inoculation or early sequential inoculation is presented as the best tool to prevent Brettanomyces contamination by reducing the lag phase between AF and MLF, as also shown in scientific studies [86,88].
5. Managing Malolactic Fermentation for Sustainability
MLF is the microbial transformation that, more than others, affects post-fermentation stages such as aging, color stabilization, and microbial stabilization. As expressed before and also outlined in Figure 1, an effective and reliable MLF with no lag phases and no nutrient leftovers is an essential step for avoiding further microbial spoilage and consequent overutilization of sulfites or other corrective tools [89,90]. Moreover, stuck or sluggish MLFs not only put the final wine quality at risk, but also require enological interventions, such as additional racking-off operations, cellulose-based adjuvants, and cooling or warming (depending on the situation), that burden the environmental impact of the winemaking process. In this context, the current knowledge on yeast–bacteria characteristics and interactions can be implemented by winemakers in protocols for avoiding fermentation slowdowns in risky situations.
5.1. Control of Microbial Spoilage during Malolactic Fermentation
Over the last 20 years, various studies have reported many factors that influence the development of LAB in wine, providing to winemakers an interpretation key to understand MLF problems and some tools to manage the MLF process [89,91,92,93,94]. Indeed, if some of the parameters affecting MLF feasibility are not easy to change (grape variety, alcohol or potential alcohol, pH, and malic acid content), many others can be managed by the winemakers in order to minimize risks of stuck or sluggish fermentations (SO2 concentration, temperature, nutrients, and yeast strain), as reviewed in [61]. Moreover, difficulties arising from the impact of two or more of the abovementioned conditions together may cause a problem of much greater difficulty than what would have been predicted by a single parameter acting alone. Therefore, each step of the winemaking process needs to be approached with as complete an understanding as possible to favor bacterial development and fermentative activity. Among other factors, the yeast strain chosen for AF deserves a special attention in this paper, as yeast–bacteria compatibility is a key parameter for exploiting microbial resources and their interactions for a fruitful MLF [95,96]. The ability of LAB to undergo MLF is affected by many factors directly or indirectly influenced by the yeast strain carrying out AF, including inhibitor content (e.g., SO2 and medium chain fatty acids), nutrient consumption/limitation, other potential as-yet-unknown factors, and interactions with the indigenous microflora of the fermentation [97]. Therefore, the use of yeast strains known to inhibit MLF should be avoided, despite their potential enological interest (e.g., for aromatic or fermentative features) at least in difficult must conditions [98]. Various studies have addressed the interactions between bacteria using different yeast/bacteria pairs, as summarized in earlier reviews [61,91,95,96], giving the winemaker useful information for strain choice. Particular attention to yeast strain individuation should be paid when using non-Saccharomyces and Saccharomyces, in order to keep the environment favorable to MLF later on [60,99].
For successful induction of malolactic fermentations when starter cultures containing malolactic bacteria are used, it is critical that the most appropriate bacterial strain is selected for the prevailing wine conditions. Since the four main limiting factors (alcohol, pH, temperature, and SO2) have a cumulative stress effect on cultures, all should be considered for the best choice [61,100]. This may lead to the choice of inoculating either an Oenococcus oeni or Lactobacillus plantarum strain, depending on the must condition. Another point on MLF management that can be important to prevent spoilage is biogenic amine (BA) formation. Previous work showed that most of the commercial malolactic bacteria did not produce BA, and that the application of commercial malolactic starters in wines is useful to reduce the BA amounts, since BA concentrations in inoculated wines were significantly lower compared with those in non-inoculated wines [101]. These results suggest that the use of selected malolactic starters can minimize BA production [102]. When BA-producing strains are present in indigenous microbiota, a winemaker is particularly encouraged to inoculate selected malolactic starters to replace the indigenous microorganisms. Nevertheless, when the dominance of starter cultures on the indigenous BA-producing microbiota is not sufficient, this does not represent the definitive solution. Thus, a recent study reports the selection of autochthonous strains of L. plantarum able to degrade BA and their suitability as malolactic starters in wine production [103]. This represents one more scenario in which BA could be controlled, in fermented foods, by modulating microbial resources as MLF.
5.2. Energy Savings Associated with Malolactic Fermentation
As inoculated fermentations of both yeast and bacteria are nowadays practiced in most wine regions of the world, there has been considerable research aimed at optimizing the time point for the inoculation of different yeasts for AF and bacteria for MLF, the latter resulting in a growing interest in the use of co-inoculation (inoculation of LAB starters at 24 to 48 h after yeast inoculation) in the production of many red and some white wines [104]. As expressed before (Section 4), co-inoculation is often proposed as a worthwhile alternative for winemaking compared with traditional post-alcoholic fermentation LAB inoculation or spontaneous MLF [105] (as more and more findings illustrate that co-inoculated MLF is an effective and novel way of modulating the volatile and aroma compound profile of wine [104,106,107]). Indeed, relative to sequential inoculation, co-inoculation reduces overall vinification time. This has at least two important consequences for the wine industry; firstly, speeding up the vinification rate leads to more rapid wine stabilization and reduces the risk of spoilage [90,104], as also recommended by the OIV [19]; secondly, this can significantly reduce the necessity to heat tanks or the whole cellar, a step that is necessary to start the MLF when a sequential inoculation or spontaneous MLF is desired. The heat that is naturally produced by yeasts during alcoholic fermentation favors malolactic fermentation (MLF), thus the early management and accomplishment of malolactic fermentation allows to avoid tank warming, necessary to achieve malic acid transformation in winter/spring season [61]. In this context, more research will be needed to accurately quantify energy savings related to co-inoculation, as less data are currently available. Some rough estimations have been published in Italy in 2009 [108], in a study where total money savings from co-inoculation (including mainly the cost for energy saved for warming tanks, but also extra costs kept back, such as wine analyses) were estimated as €0.08/bottle. In the frame of an ongoing European research project, a study was carried out in Spain comparing controlled malolactic fermentations using co-inoculation with spontaneous MLF [109]. The wines were kept at 20 °C until MLF was completed. Preliminary results showed that the co-inoculated MLF was very fast (completed 5 days after the end of AF), whereas the spontaneous tank started MLF very late (completed 45 days after the end of AF). In this long period of time, the energy consumption to heat the tank was measured and resulted in a significant value, in the order of 150 kWh/hL. This energy expenditure for the spontaneous MLF trial had a calculated cost in the order of €10/hL, although cost may vary according to the price of kWh depending on the country, the power of the heating equipment, the outside temperature, the volume of wine, and the duration of MLF [109,110].
6. Sustainable Procedures in Post-Fermentation, Stabilization, Aging, and Storage
As stated before, sulfur dioxide (SO2) is the key additive for the preservation of wines; therefore, its role is not limited to must treatment, but is also crucial for stabilizing wines in post-fermentative stages [11]. In this context, the correct management of fermentation steps (alcoholic and malolactic) can be critical to obtain the best ratio between free and bound SO2. Beyond the contribution of AF (production and consumption of both SO2 and its binding-molecule acetaldehyde by yeasts, described in Section 3.1), late stages of malolactic fermentation have been known to have an impact on bound SO2, potentially reducing its levels. Some studies suggest that microbiological wine stabilization at one week after malic acid depletion is an effective strategy for maximum removal of SO2 binders. This time frame is optimal for exploiting acetaldehyde consumption by O. oeni, still controlling the risk of possible post-MLF spoilage by bacteria leading to the production acetic acid and biogenic amines [111,112].
In case of microbial detrimental contamination, the addition of chitosan is another microbe-based tool for controlling the growth of undesirable microorganisms, particularly Brettanomyces [113], but also acetic acid bacteria [114]. Chitosan is a linear polysaccharide composed of two repeating units (D-glucosamine units (GlcN) and N-acetyl-D-glucosamine (GLcNAc) units) randomly distributed along the polymer chain and linked by β(1-4)-bonds [115]. The chitosan preparation specifically allowed in winemaking is microbial (“of fungoid origin” [116]) and is currently produced in Aspergillus niger [113]. Recent studies have shown the impact of chitosan application on wines contaminated with Brettanomyces bruxellensis, leading to a drop in B. bruxellensis cells, even at population levels as high as 105–106 CFU/mL [117]. In some studies, the chitosan preparation was added to the wine under storage and the wine was racked off (usually after 10 days), and the efficiency of the treatment was evaluated in a short delay after the wines were racked off [118,119]. In other cases, chitosan was successfully employed to control wine microbiological stability during the period of aging in barrels, in order to prevent wine from B. bruxellensis contamination along the aging period (up to 9 months) [87].
Furthermore, some investigations may propose in the future new strategies to select Oenococcus oeni strains holding competitive advantages for surviving in wine after fermentation, preventing microbial spoilage, and improving the wine organoleptic profile, thanks to their biofilm formation. Indeed, prior observations showed that O. oeni was able to survive for several months in harsh wine conditions in oak barrels. Since biofilm is a prevailing microbial lifestyle in natural environments, the capacity of O. oeni to form biofilms was recently investigated on winemaking materials, such as stainless steel and oak chips [120]. Promising results showed that biofilm could be considered as a novel approach for performing MLF and as an alternative way of adapting MLF starters to wine stress.
7. Sustainable Management of By-Products and Wastewater
Bioeconomy and circular economy have gained political traction during the second decade of this century. The movement of bioeconomy toward the use of wastes, co-products, and residue sources resonates well with circular economy principles of making the most efficient uses of natural resources. Microbial transformations, such as fermentations for metabolite production, composting, and controlled oxidizations may contribute to this effort in the case of winemaking for transforming co-products and by-products, including grape marc and pomace, vine shoots, and winery wastewater (as outlined in Figure 1). Indeed, up to 210 million tons of grapes (Vitis vinifera L.) are produced annually, with 15% of the produced grapes addressed to the wine-making industry. This socio-economic activity generates a large amount of wastes (up to 30%, w/w of the material used) [121].
7.1. Microbial Valorization of Solid Co-Products
Grape marc is the most important by-product of the winemaking industry. It consists of the solid residue left after juice extraction from grapes and contains skins, seeds, and, in some cases, stalks. It can represent a co-product to be further valorized or a waste to be treated, depending on the situation. Spirits obtained from grape pomace distillation are produced in almost all the Mediterranean countries, allowing economical valorization of marc, which can therefore represent a co-product in this area. The fermented material to be distilled is usually produced by extended storage of the marc, which allows alcoholic fermentation, therefore involving a further microbial transformation, especially in the case of white pomace [122]. Grape marc from red grapes has already undergone alcoholic fermentation with the must and can be distilled immediately, whereas marc from white grapes does not contain ethanol, but contains sugars that are fermented by spontaneous anaerobic fermentation during a storage period. Marc is stored for a period lasting from a few days to several weeks, when fermentation of residual sugars occurs mainly by yeast activity, but bacterial populations can also develop and are often associated with off-flavor production [123,124,125]. Therefore, a careful management of the fermentation process during storage is increasingly being applied by distilleries, often employing marc acidification [122], temperature control [125], and yeast inoculation [126]. Research studies showed that the lowering of the pH [122] caused significant changes in the yeast–bacteria populations ratio and in yeast species turnover, determining an improvement of the aromatic profile of the distillate, due to the reduction of the main volatile products associated with potential off-flavors [122]. A significant impact on yeast ecology variability under marc storage and on sensory quality of the distillate was also shown for temperature [125]. Moreover, results demonstrated that effective inoculation of yeast strains (although not easy to achieve due to the solid state of grape marc) has a great impact on the fermentation of grape marc during storage, by leading not only to the increased development of aroma molecules, but also to the control of spoilage microorganisms that could greatly affect product quality [126].
Grape and wine co-products are also good sources of carbon and have been used to generate various high-value products like citric acid, lactic acid, gluconic acid, and ethanol through submerged and solid-state fermentation [127,128]. Trichoderma harzianum, Aspergillus niger, Penicillium chrysogenum, and Penicillium citrinum have been used in order to degrade winery biomass, leading to the production of commercially important metabolites such as, among others, stigmasterol, glycerol, maleic acid, xylitol, and citric acid [129]. Moreover, protein-rich products can be used as feedstock for animals. The protein content of grape marc increased from 7% to 27% in five days using the solid-state fermentation process and certain fungal strains and managing specific conditions like temperature and moisture content [130,131]. Submerged fermentation of grape wastes using Monascus purpureus produced a red pigment to be used in food industry and the usage of Lactococcus lactis and Lactobacillus pentosus led to the production of lactic acid and Trametes pubescens to produce laccase [131].
Finally, there is interest in wine co-products as a substrate for Aureobasidium pullulans growth and the production of pullulan, an extracellular and unbranched homopolysaccharide useful for biofilms and for applications in medical sciences, particularly drug delivery [51]. Grape skin pulp is considered as one of the best substrates for pullulan production, especially hot water extracts of the pulp. The product is of higher molecular weight and rather pure. Moreover, poly(β-L-malic acid) (PMA) is a natural biopolyester produced by many microorganisms including A. pullulans. The interest in this molecule could be attributed to its properties of being biodegradable, water-soluble, and biocompatible, and its uses in the pharmaceutical industry. No applications in the wine industry have been reported, but possible relationships can be explored via wine wastes as substrates for PMA production and PMA as a coating for post-harvest protection of grapes, similar to that previously proposed for pullulan production [51].
As a last remark, grape pomace (pulp and skins) was also investigated as a new biosorbent for removing mycotoxins from liquid media. In vitro adsorption experiments showed that the pomace obtained from Primitivo grapes was able to rapidly and simultaneously sequester different mycotoxins. Aflatoxin B1 was the most adsorbed mycotoxin, followed by zearalenone and ochratoxin A [132]. An innovative winemaking procedure involving the use of grape pomace has been suggested as a corrective measure to reduce ochratoxin A (OTA) levels in must and wines.
7.2. Microbial Treatment of Wastewater and Solid Residues
Wastewater sources are major causes for environmental pollution in surface and ground water bodies. Current wastewater treatment technologies are not sustainable because they are energy- and cost-intensive, leaving latitude for the development of technologies that are energy-conservative or energy-yielding. For the present and future context, microbial fuel cell technology may present a sustainable and environmentally friendly route to meet the water sanitation needs [133]. Microbial fuel cells (MFCs) are electrochemical devices that use the metabolic activity of microorganisms to oxidize fuels, generating electric current by direct or mediated electron transfer to electrodes. In the anodic compartment, organic matter is oxidized by microbial metabolism, which transfers the electrons to the anode. In the cathodic compartment, oxygen or oxidized compounds are reduced either via an abiotic process or by microbially mediated reduction [134]. The bacterial communities that develop in these systems show great diversity, ranging from primarily δ-Proteobacteria that predominate in sediment MFCs to communities composed of α-, β-, γ-, or δ-Proteobacteria, Firmicutes, and uncharacterized clones in other types of MFCs [135]. Microbial fuel cells can treat agro-industrial wastewater, and a few studies have reported the treatment of winery wastes. Dual chamber MFCs were used to treat real effluents from wine-processing factories. Results demonstrated that electricity can be produced efficiently and that the unbalanced nutrients/COD ratio was a major challenge in the treatment of winery wastewater, in spite of the very high organic load contained in this type of wastewater [136,137]. In another study, a single-chamber MFC was used to treat white wine lees and red wine lees [138]. Different degradability, due to different substrate composition, gave different results: white wine lees produced much more electricity and degradation, i.e., total-COD removal, than red wine lees (the high presence of polyphenols in the latter, played a role in reducing MFC performance). Different substrates led, also, to different microbial consortia. Electricity and degradation obtained with white wine lees indicated their suitability to be treated by MFC. At present, the technology is proposed for wineries at the industrial level for feasibility studies [139].
Besides, wine production determines the creation of large amounts of solid residues, such as vine branches from winter pruning and grape marc from winemaking (when not transferred to distilleries). Composting is a process transforming the organic matter by an aerobic biological process, allowing organic matter degradation and stabilization. Composting is becoming an ecological and economical microbially based alternative for reusing plant biomass residues [140] and therefore, residues from vitiviniculture. Indeed, by-products such as pruning residues and grape marcs can be exploited for the production of compost. In particular, grape marc [141,142] and branches have been composted, separately or together [143], showing beneficial results. Quite recently, wine by-products utilization by co-composting with olive mill wastewater was also proposed with promising results [144]. This represents an affordable and useful tool for both grape growers and winemakers, as the reutilization of residues produced by winemaking-related activities can lead to obtain a compost that can be reintroduced in the vineyard from where the plant biomass came from [143,145]. Additionally, it is worth to note that suppression of soil-borne diseases of horticultural crops by compost has been proved and attributed to the activities of antagonistic microorganisms, as a great diversity of biological control agents naturally colonize compost [140]. Focusing on winemaking by-products, the suppressive capacity of grape marc compost against Pythium aphanidermatum and Phytophthora parasitica was determined with promising results on cucumber [146]. In a further study, the large number of microbes which appeared in the microbiological analyses of grape marc compost was characterized and most microorganisms were bacteria. Antagonist in vitro assays were performed showing effective antagonistic activities against all the fungal pathogens tested [141]. This opens up further positive implications in terms of sustainability improvement. Most soil-borne pathogens are difficult to control by conventional strategies such as the use of synthetic fungicides. The lack of reliable chemical controls, the occurrence of fungicide resistance in pathogens, and the breakdown or circumvention of host resistance by pathogen populations are among the key factors underlying efforts to develop more sustainable control measures [141]. Therefore, the use of winery compost would represent an environmentally friendly tool, even in terms of circular economy if applied on vineyards [145].
8. Conclusions
In agriculture, the wine sector is one of the industries most affected by the environmental sustainability issue. Recently, the contribution of winemaking, from grape harvest to bottling, has been considered together with vineyard management in assessing the environmental impact of vitiviniculture. Several cellar processes could be improved for reducing the environmental impact of the whole chain, including microbe-driven transformations. The aim of this paper was to review the potential of microorganisms and interactions thereof as a natural, environmentally friendly tool to improve the sustainability aspects of winemaking, all along the production chain, including waste treatment. Microorganisms play a role in several steps of the winemaking process, most of which can be improved for reducing the environmental impact. These steps include pre-fermentative stages, alcoholic and malolactic fermentations and their management and timing, post-fermentation and stabilization processes, and valorization or treatment of by-products. Microbial resources exploitable for sustainability improvements include a wide array of genera and species comprising yeasts (both non-Saccharomyces and Saccharomyces), fungi, and bacteria. Moreover, microbial interactions and their exploitation also play a crucial role. Special attention was paid to microbial resources and processes which are already, or about to become, available for the winemaking sector at the industrial scale. In conclusion, the paper illustrates how the presence of proper yeast or bacterial strains, the management and timing of starter cultures inoculation, and some appropriate technological modifications that favor selected microbial activities can lead to several positive effects, including (among others) energy savings, reduction of chemical additives such as sulfites, and reuse of certain residues.
List of abbreviations
| ADY | Active dry yeast |
| AF | Alcoholic fermentation |
| BA | Biogenic amine |
| COD | Chemical oxygen demand |
| GHG | Greenhouse gas |
| LAB | Lactic acid bacteria |
| MLF | Malolactic fermentation |
| MFC | Microbial fuel cells |
| OIV | International Organisation of Vine and Wine |
| PCM | Pre-fermentative cold maceration |
| VOC | Volatile organic compound |
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Trioli, G.; Sacchi, A.; Corbo, C.; Trevisan, M. Environmental impact of vinegrowing and winemaking inputs: An european survey. Internet J. Viticult. Enol. 2015, 7, 2. [Google Scholar]
- De Matos, C.T.; Garcia, J.C.; Aurambout, J.-P.; Manfredi, S. Environmental sustainability assessment of bioeconomy products and processes—Progress report 1. Eur. Comm. Rep. EUR 27356 EN 2015. [Google Scholar] [CrossRef]
- Pomarici, E.; Vecchio, R. Will sustainability shape the future wine market? Wine Econ. Policy 2019, 8, 1–4. [Google Scholar] [CrossRef]
- Santini, C.; Cavicchi, A.; Casini, L. Sustainability in the wine industry: Key questions and research trends a. Agric. Food Econ. 2013, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Bonamente, E.; Scrucca, F.; Rinaldi, S.; Merico, M.C.; Asdrubali, F.; Lamastra, L. Environmental impact of an Italian wine bottle: Carbon and water footprint assessment. Sci. Total Environ. 2016, 560–561, 274–283. [Google Scholar] [CrossRef] [PubMed]
- REGULATION (EC) No 1221/2009 on the Voluntary Participation by Organisations in a Community Eco-Management and Audit Scheme (EMAS). Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/HTML/?uri=OJ:L:2009:342:FULL&from=EN (accessed on 4 December 2019).
- Merli, R.; Preziosi, M.; Acampora, A. Sustainability experiences in the wine sector: Toward the development of an international indicators system. J. Clean. Prod. 2018, 172, 3791–3805. [Google Scholar] [CrossRef]
- Berbegal, C.; Fragasso, M.; Russo, P.; Bimbo, F.; Grieco, F.; Spano, G.; Capozzi, V. Climate Changes and Food Quality: The Potential of Microbial Activities as Mitigating Strategies in the Wine Sector. Fermentation 2019, 5, 85. [Google Scholar] [CrossRef] [Green Version]
- Castrillo, D.; Rabuñal, E.; Neira, N.; Blanco, P. Oenological potential of non-Saccharomyces yeasts to mitigate effects of climate change in winemaking: Impact on aroma and sensory profiles of Treixadura wines. FEMS Yeast Res. 2019, 19, foz065. [Google Scholar] [CrossRef]
- Dequin, S.; Escudier, J.-L.; Bely, M.; Noble, J.; Albertin, W.; Masneuf-Pomarède, I.; Marullo, P.; Salmon, J.-M.; Sablayrolles, J.M. How to adapt winemaking practices to modified grape composition under climate change conditions. OENO One 2017, 51, 205–214. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology, the Microbiology of Wine and Vinifications; John Wiley & Sons: Hoboken, NJ, USA, 2006; Volume 1. [Google Scholar]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Pretorius, I.S. Microbial formation and modification of flavor and off-flavor compounds in wine. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer: Berlin/Heidelberg, Germany, 2009; pp. 209–231. [Google Scholar]
- Malfeito-Ferreira, M. Yeasts and wine off-flavours: A technological perspective. Annals Microbiol. 2011, 61, 95–102. [Google Scholar] [CrossRef]
- Mas, A. Microbial challenges in sustainable winemaking. OENOVITI Int. Netw. 2018, 25, 38. [Google Scholar]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a bio-protection agent in winemaking. Food Res. Int. 2018, 107, 451–461. [Google Scholar] [CrossRef]
- Berbegal, C.; Spano, G.; Fragasso, M.; Grieco, F.; Russo, P.; Capozzi, V. Starter cultures as biocontrol strategy to prevent Brettanomyces bruxellensis proliferation in wine. Appl. Microbiol. Biotechnol. 2018, 102, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Rubio-Bretón, P.; Gonzalo-Diago, A.; Iribarren, M.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. Bioprotection as a tool to free additives winemaking: Effect on sensorial, anthocyanic and aromatic profile of young red wines. LWT 2018, 98, 458–464. [Google Scholar] [CrossRef]
- OIV Oenology Resolutions—OIV/OENO 462/2014. Available online: http://www.oiv.int/en/technical-standards-and-documents/resolutions-of-the-oiv/oenology-resolutions (accessed on 29 October 2016).
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aroma-a review. S. Afr. J. Enol. Viticult. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Aranda, A. Enological Repercussions of Non-Saccharomyces Species. Fermentation 2019, 5, 68. [Google Scholar] [CrossRef] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial Resources and Enological Significance: Opportunities and Benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef] [Green Version]
- Nardi, T.; Nadai, C.; Bovo, B.; Treu, L.; Campanaro, S.; Giacomini, A.; Corich, V. Yeast selection criteria for improvement of sulphite management in winemaking. In Proceedings of the Oeno2011-Actes de Colloques du 9 e Symposium International D’oenologie de Bordeaux, Bordeaux, France, 15–17 June 2012; pp. 445–450. [Google Scholar]
- Berbegal, C.; Garofalo, C.; Russo, P.; Pati, S.; Capozzi, V.; Spano, G. Use of autochthonous yeasts and bacteria in order to control Brettanomyces bruxellensis in wine. Fermentation 2017, 3, 65. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in Wine Biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Pawlikowska, E.; James, S.A.; Breierova, E.; Antolak, H.; Kregiel, D. Biocontrol capability of local Metschnikowia sp. isolates. Antonie Leeuwenhoek 2019, 112, 1425–1445. [Google Scholar] [CrossRef] [Green Version]
- Piano, S.; Neyrotti, V.; Migheli, Q.; Gullino, M.L. Biocontrol capability of Metschnikowia pulcherrima against Botrytis postharvest rot of apple. Postharvest Biol. Technol. 1997, 11, 131–140. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Ciavorella, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Metschnikowia pulcherrima strain MACH1 outcompetes Botrytis cinerea, Alternaria alternata and Penicillium expansum in apples through iron depletion. Postharvest Biol. Technol. 2008, 49, 121–128. [Google Scholar] [CrossRef]
- Spadaro, D.; Vola, R.; Piano, S.; Gullino, M.L. Mechanisms of action and efficacy of four isolates of the yeast Metschnikowia pulcherrima active against postharvest pathogens on apples. Postharvest Biol. Technol. 2002, 24, 123–134. [Google Scholar] [CrossRef]
- Kinay, P.; Yildiz, M. The shelf life and effectiveness of granular formulations of Metschnikowia pulcherrima and Pichia guilliermondii yeast isolates that control postharvest decay of citrus fruit. Biol. Control 2008, 45, 433–440. [Google Scholar] [CrossRef]
- Sharma, R.R.; Singh, D.; Singh, R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control 2009, 50, 205–221. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia Strains Isolated from Botrytized Grapes Antagonize Fungal and Bacterial Growth by Iron Depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmer, P.A.G.; Reglinski, T. Biosuppression of Botrytis cinerea in grapes. Plant Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Raspor, P.; Miklič-Milek, D.; Avbelj, M.; Čadež, N. Biocontrol of Grey Mould Disease on Grape Caused by Botrytis cinerea with Autochthonous Wine Yeasts. Food Technol. Biotechnol. 2010, 48, 336–343. [Google Scholar]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. Recent Patents food Nutr. Agric. 2019, 10, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Benucci, I.; Luziatelli, F.; Cerreti, M.; Liburdi, K.; Nardi, T.; Vagnoli, P.; Ruzzi, M.; Esti, M. Pre-fermentative cold maceration in the presence of non-Saccharomyces strains: Effect on fermentation behaviour and volatile composition of a red wine. Aust. J. Grape Wine Res. 2017, 24, 135–281. [Google Scholar] [CrossRef]
- Benucci, I.; Cerreti, M.; Liburdi, K.; Nardi, T.; Vagnoli, P.; Ortiz-Julien, A.; Esti, M. Pre-fermentative cold maceration in presence of non-Saccharomyces strains: Evolution of chromatic characteristics of Sangiovese red wine elaborated by sequential inoculation. Food Res. Int. 2018, 107, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Padilla, B.; Gil, J.V.; Manzanares, P. Challenges of the Non-Conventional Yeast Wickerhamomyces anomalus in Winemaking. Fermentation 2018, 4, 68. [Google Scholar] [CrossRef] [Green Version]
- Mannazzu, I.; Domizio, P.; Carboni, G.; Zara, S.; Zara, G.; Comitini, F.; Budroni, M.; Ciani, M. Yeast killer toxins: From ecological significance to application. Crit. Rev. Biotechnol. 2019, 39, 603–617. [Google Scholar] [CrossRef]
- Velázquez, R.; Zamora, E.; Álvarez, M.L.; Hernández, L.M.; Ramírez, M. Effects of new Torulaspora delbrueckii killer yeasts on the must fermentation kinetics and aroma compounds of white table wine. Front. Microbiol. 2015, 6, 1222. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.-L.; Chi, Z.; Wang, G.-Y.; Wang, Z.-P.; Li, Y.; Chi, Z.-M. Yeast killer toxins, molecular mechanisms of their action and their applications. Crit. Rev. Biotechnol. 2015, 35, 222–234. [Google Scholar] [CrossRef]
- Mehlomakulu, N.N.; Setati, M.E.; Divol, B. Non-Saccharomyces Killer Toxins: Possible Biocontrol Agents Against Brettanomyces in Wine? S. Afr. J. Enol. Viticult. 2015, 36, 94–104. [Google Scholar] [CrossRef] [Green Version]
- Oro, L.; Ciani, M.; Bizzaro, D.; Comitini, F. Evaluation of damage induced by Kwkt and Pikt zymocins against Brettanomyces/Dekkera spoilage yeast, as compared to sulphur dioxide. J. Appl. Microbiol. 2016, 121, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Villalba, M.L.; Sáez, J.S.; del Monaco, S.; Lopes, C.A.; Sangorrín, M.P. TdKT, a new killer toxin produced by Torulaspora delbrueckii effective against wine spoilage yeasts. Int. J. Food Microbiol. 2016, 217, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Kuchen, B.; Maturano, Y.P.; Mestre, M.V.; Combina, M.; Toro, M.E.; Vazquez, F. Selection of Native Non-Saccharomyces Yeasts with Biocontrol Activity against Spoilage Yeasts in Order to Produce Healthy Regional Wines. Fermentation 2019, 5, 60. [Google Scholar] [CrossRef] [Green Version]
- Infowine Non Saccharomyces contro Brett: Nuove Prospettive nell’Impiego di Lieviti ad Attività Antimicrobica—Video—Piacenza. Available online: https://www.infowine.com/it/video/non_saccharomyces_contro_brett_nuove_prospettive_nellimpiego_di_lieviti_ad_attivita_antimicrobica_sc_13312.htm (accessed on 17 February 2020).
- Bozoudi, D.; Tsaltas, D. The Multiple and Versatile Roles of Aureobasidium pullulans in the Vitivinicultural Sector. Fermentation 2018, 4, 85. [Google Scholar] [CrossRef] [Green Version]
- Guzzon, R.; Franciosi, E.; Larcher, R. A new resource from traditional wines: Characterisation of the microbiota of “Vino Santo” grapes as a biocontrol agent against Botrytis cinerea. Eur. Food Res. Technol. 2014, 239, 117–126. [Google Scholar] [CrossRef]
- Lorenzini, M.; Zapparoli, G. Epiphytic bacteria from withered grapes and their antagonistic effects on grape-rotting fungi. Int. J. Food Microbiol. 2020, 319, 108505. [Google Scholar] [CrossRef]
- Nadai, C.; Junior, W.J.F.L.; Favaron, F.; Giacomini, A.; Corich, V. Biocontrol activity of Starmerella bacillaris yeast against blue mold disease on apple fruit and its effect on cider fermentation. PLoS ONE 2018, 13, e0204350. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Droby, S. Metschnikowia fructicola, a new ascosporic yeast with potential for biocontrol of postharvest fruit rots. Syst. Appl. Microbiol. 2001, 24, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Karabulut, O.A.; Smilanick, J.L.; Gabler, F.M.; Mansour, M.; Droby, S. Near-harvest applications of Metschnikowia fructicola, ethanol, and sodium bicarbonate to control postharvest diseases of grape in central California. Plant Disease 2003, 87, 1384–1389. [Google Scholar] [CrossRef] [Green Version]
- Torriani, S.; Tornielli, G. Metschnikowia fructicola, un lievito contro la botrite. Vite&Vino 2019, 2, 38. [Google Scholar]
- Bagheri, B.; Zambelli, P.; Vigentini, I.; Bauer, F.F.; Setati, M.E. Investigating the Effect of Selected Non-Saccharomyces Species on Wine Ecosystem Function and Major Volatiles. Front. Bioeng. Biotechnol. 2018, 6, 139. [Google Scholar] [CrossRef] [PubMed]
- García, M.; Esteve-Zarzoso, B.; Arroyo, T. Non-Saccharomyces yeasts: Biotechnological role for wine production. In Grape and Wine Biotechnology; Morata, A., Ed.; IntechOpen: London, UK, 2016; pp. 249–271. [Google Scholar]
- Nardi, T.; Panero, L.; Petrozziello, M.; Guaita, M.; Tsolakis, C.; Cassino, C.; Vagnoli, P.; Bosso, A. Managing wine quality using Torulaspora delbrueckii and Oenococcus oeni starters in mixed fermentations of a red Barbera wine. Eur. Food Res. Technol. 2019, 245, 293–307. [Google Scholar] [CrossRef]
- Nardi, T.; Bordiga, M. Fermentation Process. In Post-Fermentation and-Distillation Technology: Stabilization, Aging, and Spoilage; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- OIV Code of Good Vitivinicultural Practices in Order to Avoid or Limit Contamination by Brettanomyces. Available online: http://www.oiv.int/en/technical-standards-and-documents/good-practices-guidelines/code-of-good-vitivinicultural-practices-in-order-to-avoid-or-limit-contamination-by-brettanomyces (accessed on 18 February 2020).
- Alexandre, H.; Charpentier, C. Biochemical aspects of stuck and sluggish fermentation in grape must. J. Ind. Microbiol. Biotechnol. 1998, 20, 20–27. [Google Scholar] [CrossRef]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Duc, C.; Pradal, M.; Sanchez, I.; Noble, J.; Tesniere, C.; Blondin, B. A set of nutrient limitations trigger yeast cell death in a nitrogen-dependent manner during wine alcoholic fermentation. PLoS ONE 2017, 12, e0184838. [Google Scholar] [CrossRef]
- Childs, B.C.; Bohlscheid, J.C.; Edwards, C.G. Impact of available nitrogen and sugar concentration in musts on alcoholic fermentation and subsequent wine spoilage by Brettanomyces bruxellensis. Food Microbiol. 2015, 46, 604–609. [Google Scholar] [CrossRef]
- Noble, J.; Sanchez, I.; Blondin, B. Identification of new Saccharomyces cerevisiae variants of the MET2 and SKP2 genes controlling the sulfur assimilation pathway and the production of undesirable sulfur compounds during alcoholic fermentation. Microb. Cell Fact. 2015, 14, 68. [Google Scholar] [CrossRef] [Green Version]
- Morgan, S.C.; Haggerty, J.J.; Johnston, B.; Jiranek, V.; Durall, D.M. Response to Sulfur Dioxide Addition by Two Commercial Saccharomyces cerevisiae Strains. Fermentation 2019, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Mira de Orduña, R.; Lamon, J. Acetaldehyde Management during Winemaking. Available online: https://www.winemak-in.com/en/publications/acetaldehyde-management-during-winemaking (accessed on 18 February 2020).
- Li, E.; De Orduña, R.M. Acetaldehyde kinetics of enological yeast during alcoholic fermentation in grape must. J. Ind. Microbiol. Biotechnol. 2017, 44, 229–236. [Google Scholar] [CrossRef]
- Coetzee, C.; Buica, A.; Du Toit, W.J. The Use of SO2 to Bind Acetaldehyde in Wine: Sensory Implications. S. Afr. J. Enol. Viticult. 2018, 39, 1–6. [Google Scholar]
- Osborne, J.P.; de Orduña, R.M.; Pilone, G.J.; Liu, S.-Q. Acetaldehyde metabolism by wine lactic acid bacteria. FEMS Microbiol. Lett. 2000, 191, 51–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondin, B.; Noble, J.; Sanchez, I. Method for Controlling the Production of Sulphites, of Hydrogen Sulphide and of Acetaldehyde by Yeasts—Global Patent Index—EP 2807247 B1; European Patent Office: Munich, Germany, 2017. [Google Scholar]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Inte. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef] [PubMed]
- Andorrà, I.; Martín, L.; Nart, E.; Puxeu, M.; Hidalgo, C.; Ferrer-Gallego, R. Effect of grape juice composition and nutrient supplementation on the production of sulfur dioxide and carboxylic compounds by Saccharomyces cerevisiae. Aust. J. Grape Wine Res. 2018, 24, 260–266. [Google Scholar] [CrossRef]
- Masneuf-Pomarède, I.; Mansour, C.; Murat, M.-L.; Tominaga, T.; Dubourdieu, D. Influence of fermentation temperature on volatile thiols concentrations in Sauvignon blanc wines. Int. J. Food Microbiol. 2006, 108, 385–390. [Google Scholar]
- Torija, M.J.; Beltran, G.; Novo, M.; Poblet, M.; Guillamón, J.M.; Mas, A.; Rozes, N. Effects of fermentation temperature and Saccharomyces species on the cell fatty acid composition and presence of volatile compounds in wine. Int. J. Food Microbiol. 2003, 85, 127–136. [Google Scholar] [CrossRef]
- Molina, A.M.; Swiegers, J.H.; Varela, C.; Pretorius, I.S.; Agosin, E. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. Biotechnol. 2007, 77, 675–687. [Google Scholar] [CrossRef]
- Galitsky, C.; Worrell, E.; Radspieler, A.; Healy, P.; Zechiel, S. BEST Winery Guidebook: Benchmarking and Energy and Water Savings Tool for the Wine Industry; Alameda Lab: Alameda, CA, USA, 2005. [Google Scholar]
- Malvoni, M.; Congedo, P.M.; Laforgia, D. Analysis of energy consumption: A case study of an Italian winery. Energy Procedia 2017, 126, 227–233. [Google Scholar] [CrossRef]
- Flint, D.J.; Golicic, S.L. Searching for competitive advantage through sustainability. Int. J. Phys. Distrib. Logist. Manag. 2009, 39, 841–860. [Google Scholar] [CrossRef]
- Szolnoki, G. A cross-national comparison of sustainability in the wine industry. J. Clean. Prod. 2013, 53, 243–251. [Google Scholar] [CrossRef]
- Galletto, L.; Barisan, L. Carbon footprint as a lever for sustained competitive strategy in developing a smart oenology: Evidence from an exploratory study in Italy. Sustainability 2019, 11, 1483. [Google Scholar] [CrossRef] [Green Version]
- Giovenzana, V.; Beghi, R.; Vagnoli, P.; Iacono, F.; Guidetti, R.; Nardi, T. Evaluation of Energy Saving Using a New Yeast Combined with Temperature Management in Sparkling Base Wine Fermentation. Am. J. Enol. Viticult. 2016, 67, 308–314. [Google Scholar] [CrossRef]
- Schwinn, M.; Durner, D.; Wacker, M.; Delgado, A.; Fischer, U. Impact of fermentation temperature on required heat dissipation, growth and viability of yeast, on sensory characteristics and on the formation of volatiles in Riesling. Aust. J. Grape Wine Res. 2019, 25, 173–184. [Google Scholar] [CrossRef]
- Gerbaux, V.; Briffox, C.; Dumont, A.; Krieger, S. Influence of inoculation with malolactic bacteria on volatile phenols in wines. Am. J. Enol. Viticult. 2009, 60, 233–235. [Google Scholar]
- Nardi, T.; Vagnoli, P.; Minacci, A.; Gautier, S.; Sieczkowski, N. Evaluating the impact of a fungal-origin chitosan preparation on Brettanomyces bruxellensis in the context of wine aging. Wine Stud. 2014, 3. [Google Scholar] [CrossRef] [Green Version]
- Coulon, J.; Perello, M.C.; Lonvaud-Funel, A.; De Revel, G.; Renouf, V. Brettanomyces bruxellensis evolution and volatile phenols production in red wines during storage in bottles. J. Appl. Microbiol. 2010, 108, 1450–1458. [Google Scholar] [CrossRef]
- Bauer, R.; Dicks, L.M.T. Control of malolactic fermentation in wine. A review. S. Afr. J. Enol. Vitic. 2004, 25, 74–88. [Google Scholar] [CrossRef] [Green Version]
- Bartowsky, E.J. Bacterial spoilage of wine and approaches to minimize it. Lett. Appl. Microbiol. 2009, 48, 149–156. [Google Scholar] [CrossRef]
- Alexandre, H.; Costello, P.J.; Remize, F.; Guzzo, J.; Guilloux-Benatier, M. Saccharomyces cerevisiae–Oenococcus oeni interactions in wine: Current knowledge and perspectives. Int. J. Food Microbiol. 2004, 93, 141–154. [Google Scholar] [CrossRef]
- Lerm, E.; Engelbrecht, L.; Du Toit, M. Malolactic fermentation: The ABC’s of MLF. S. Afr. J. Enol. Vitic. 2010, 31, 186–212. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.-Q. Malolactic fermentation in wine–beyond deacidification. J. Appl. Microbiol. 2002, 92, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Terrade, N.; de Orduña, R.M. Determination of the essential nutrient requirements of wine-related bacteria from the genera Oenococcus and Lactobacillus. Int. J. Food Microbiol. 2009, 133, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rousseaux, S.; Tourdot-Maréchal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine microbiome, a dynamic world of microbial interactions. Crit. Rev. Food Sci. Nutr. 2017, 57, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Bartle, L.; Sumby, K.; Sundstrom, J.; Jiranek, V. The microbial challenge of winemaking: Yeast-bacteria compatibility. FEMS Yeast Res. 2019, 19, foz040. [Google Scholar] [CrossRef] [PubMed]
- Sumby, K.M.; Bartle, L.; Grbin, P.R.; Jiranek, V. Measures to improve wine malolactic fermentation. Appl. Microbiol. Biotechnol. 2019, 103, 2033–2051. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Forcisi, S.; Harir, M.; Deleris-Bou, M.; Krieger-Weber, S.; Lucio, M.; Longin, C.; Degueurce, C.; Gougeon, R.D.; Schmitt-Kopplin, P.; et al. New molecular evidence of wine yeast-bacteria interaction unraveled by non-targeted exometabolomic profiling. Metabolomics 2016, 12, 1–16. [Google Scholar] [CrossRef]
- Berbegal, C.; Borruso, L.; Fragasso, M.; Tufariello, M.; Russo, P.; Brusetti, L.; Spano, G.; Capozzi, V. A Metagenomic-Based Approach for the Characterization of Bacterial Diversity Associated with Spontaneous Malolactic Fermentations in Wine. Int. J. Mol. Sci. 2019, 20, 3980. [Google Scholar] [CrossRef] [Green Version]
- Krieger-Weber, S. Application of yeast and bacteria as starter cultures. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer: Berlin/Heidelberg, Germany, 2017; pp. 605–634. [Google Scholar]
- Lonvaud-Funel, A. Biogenic amines in wines: Role of lactic acid bacteria. FEMS Microbiol. Lett. 2001, 199, 9–13. [Google Scholar] [CrossRef]
- Marques, A.P.; Leitão, M.C.; San Romão, M.V. Biogenic amines in wines: Influence of oenological factors. Food Chem. 2008, 107, 853–860. [Google Scholar] [CrossRef]
- . Capozzi, V.; Russo, P.; Ladero, V.; Fernández, M.; Fiocco, D.; Alvarez, M.A.; Grieco, F.; Spano, G. Biogenic amines degradation by Lactobacillus plantarum: toward a potential application in wine. Front. Microbiol. 2012, 3, 122. [Google Scholar] [CrossRef] [Green Version]
- Bartowsky, E.J.; Costello, P.J.; Chambers, P.J. Emerging trends in the application of malolactic fermentation. Aust. J. Grape Wine Res. 2015, 21, 663–669. [Google Scholar] [CrossRef]
- Guzzon, R.; Villega, T.R.; Pedron, M.; Malacarne, M.; Nicolini, G.; Larcher, R. Simultaneous yeast–bacteria inoculum. A feasible solution for the management of oenological fermentation in red must with low nitrogen content. Ann. Microbiol. 2013, 63, 805–808. [Google Scholar] [CrossRef]
- Knoll, C.; Fritsch, S.; Schnell, S.; Grossmann, M.; Krieger-Weber, S.; Du Toit, M.; Rauhut, D. Impact of different malolactic fermentation inoculation scenarios on Riesling wine aroma. World J. Microbiol. Biotechnol. 2012, 28, 1143–1153. [Google Scholar] [CrossRef] [PubMed]
- Guzzon, R.; Davide, S.; Villegas, T.R.; Malacarne, M.; Larcher, R.; Nardi, T.; Vagnoli, P.; Krieger-Weber, S. Exploitation of Simultaneous Alcoholic and Malolactic Fermentation of Incrocio Manzoni, a Traditional Italian White Wine. S. Afr. J. Enol. Viticult. 2016, 37, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Mo, S.; Mo, F. Differenti protocolli di gestione della fermentazione malolattica in vini Barbera d’Asti. In L’ENOLOGO; Assoenologi: Milano, Italy, 2009; pp. 77–81. [Google Scholar]
- CDTI VINYSOST—EU Project. Available online: http://vinysost.com/ (accessed on 26 February 2020).
- Vagnoli, P. “I Giorni della Sostenibilità 2020” Seminari Formazione Avanzata—Personal Communication; CIRVE, Università di Padova: Padova, Italy, 2020. [Google Scholar]
- Jackowetz, J.N.; Mira de Orduña, R. Metabolism of SO2 binding compounds by Oenococcus oeni during and after malolactic fermentation in white wine. Int. J. Food Microbiol. 2012, 155, 153–157. [Google Scholar] [CrossRef]
- Osborne, J.P.; Dubé Morneau, A.; Mira de Orduna, R. Degradation of free and sulfur-dioxide-bound acetaldehyde by malolactic lactic acid bacteria in white wine. J. Appl. Microbiol. 2006, 101, 474–479. [Google Scholar] [CrossRef]
- Taillandier, P.; Joannis-Cassan, C.; Jentzer, J.-B.; Gautier, S.; Sieczkowski, N.; Granes, D.; Brandam, C. Effect of a fungal chitosan preparation on Brettanomyces bruxellensis, a wine contaminant. J. Appl. Microbiol. 2015, 118, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Valera, M.J.; Sainz, F.; Mas, A.; Torija, M.J. Effect of chitosan and SO2 on viability of Acetobacter strains in wine. Int. J. Food Microbiol. 2017, 246, 1–4. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef]
- EU Commission Regulation (EU) No 53/2011 of 21 January 2011 Amending Regulation (EC) No 606/2009 Laying down Certain Detailed Rules for Implementing Council Regulation (EC) No 479/2008 as Regards the Categories of Grapevine Products, Oenological Practices and the Applicable Restrictions; Official Journal L019/2011. Available online: http://data.europa.eu/eli/reg/2011/53/oj/eng (accessed on 25 February 2020).
- Petrova, B.; Cartwright, Z.M.; Edwards, C.G. Effectiveness of chitosan preparations against Brettanomyces bruxellensis grown in culture media and red wines. Oeno One 2016, 50, 49–56. [Google Scholar] [CrossRef]
- Zuehlke, J.M.; Petrova, B.; Edwards, C.G. Advances in the control of wine spoilage by Zygosaccharomyces and Dekkera/Brettanomyces. Annu. Rev. Food Sci. Technol. 2013, 4, 57–78. [Google Scholar] [CrossRef] [PubMed]
- Malfeito-Ferreira, M. Two Decades of “Horse Sweat” Taint and Brettanomyces Yeasts in Wine: Where do We Stand Now? Beverages 2018, 4, 32. [Google Scholar] [CrossRef] [Green Version]
- Bastard, A.; Coelho, C.; Briandet, R.; Canette, A.; Gougeon, R.D.; Alexandre, H.; Guzzo, J.; Weidmann, S. Effect of biofilm formation by Oenococcus oeni on malolactic fermentation and the release of aromatic compounds in wine. Front. Microbiol. 2016, 7, 613. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.; Baenas, N.; Dominguez-Perles, R.; Barros, A.; Rosa, E.; Moreno, D.A.; Garcia-Viguera, C. Natural Bioactive Compounds from Winery By-Products as Health Promoters: A Review. Int. J. Mol. Sci. 2014, 15, 15638–15678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovo, B.; Nardi, T.; Fontana, F.; Carlot, M.; Giacomini, A.; Corich, V. Acidification of grape marc for alcoholic beverage production: Effects on indigenous microflora and aroma profile after distillation. Int. J. Food Microbiol. 2012, 152, 100–106. [Google Scholar] [CrossRef]
- Campanaro, S.; Treu, L.; Vendramin, V.; Bovo, B.; Giacomini, A.; Corich, V. Metagenomic analysis of the microbial community in fermented grape marc reveals that Lactobacillus fabifermentans is one of the dominant species: Insights into its genome structure. Appl. Microbiol. Biotechnol. 2014, 98, 6015–6037. [Google Scholar] [CrossRef] [Green Version]
- Maragkoudakis, P.A.; Nardi, T.; Bovo, B.; D’Andrea, M.; Howell, K.S.; Giacomini, A.; Corich, V. Biodiversity, dynamics and ecology of bacterial community during grape marc storage for the production of grappa. Int. J. Food Microbiol. 2013, 162, 143–151. [Google Scholar] [CrossRef]
- Iacumin, L.; Manzano, M.; Cecchini, F.; Orlic, S.; Zironi, R.; Comi, G. Influence of specific fermentation conditions on natural microflora of pomace in “Grappa” production. World J. Microbiol. Biotechnol. 2012, 28, 1747–1759. [Google Scholar] [CrossRef]
- Bovo, B.; Fontana, F.; Giacomini, A.; Corich, V. Effects of yeast inoculation on volatile compound production by grape marcs. Ann. Microbiol. 2011, 61, 117–124. [Google Scholar] [CrossRef]
- Bordiga, M. Valorization of Wine Making by-Products; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Bordiga, M.; Travaglia, F.; Locatelli, M. Valorisation of grape pomace: An approach that is increasingly reaching its maturity—A review. Int. J. Food Sci. Technol. 2019, 54, 933–942. [Google Scholar] [CrossRef]
- Karpe, A.V.; Beale, D.J.; Harding, I.H.; Palombo, E.A. Optimization of degradation of winery-derived biomass waste by Ascomycetes. J. Chem. Technol. Biotechnol. 2015, 90, 1793–1801. [Google Scholar] [CrossRef]
- Zepf, F.; Jin, B. Bioconversion of grape marc into protein rich animal feed by microbial fungi. Chem. Eng. Process Tech. 2013, 1, 1011–1018. [Google Scholar]
- Sotiropoulou, E.I.; Liouni, M.; Calokerinos, A.C.; Nerantzis, E. Utilization of grape pomace for the production of microbial protein-A review. In Proceedings of the 5th International Conference on Sustainable Solid Waste Management, Athens, Greece, 21–24 June 2017; pp. 21–24. [Google Scholar]
- Avantaggiato, G.; Greco, D.; Damascelli, A.; Solfrizzo, M.; Visconti, A. Assessment of multi-mycotoxin adsorption efficacy of grape pomace. J. Agricult. Food Chem. 2014, 62, 497–507. [Google Scholar] [CrossRef] [PubMed]
- Gude, V.G. Wastewater treatment in microbial fuel cells–an overview. J. Clean. Prod. 2016, 122, 287–307. [Google Scholar] [CrossRef]
- Cercado-Quezada, B.; Delia, M.-L.; Bergel, A. Testing various food-industry wastes for electricity production in microbial fuel cell. Bioresour. Technol. 2010, 101, 2748–2754. [Google Scholar] [CrossRef] [Green Version]
- Logan, B.E.; Regan, J.M. Electricity-producing bacterial communities in microbial fuel cells. Trends Microbiol. 2006, 14, 512–518. [Google Scholar] [CrossRef]
- Penteado, E.D.; Fernandez-Marchante, C.M.; Zaiat, M.; Cañizares, P.; Gonzalez, E.R.; Rodrigo, M.A. Energy recovery from winery wastewater using a dual chamber microbial fuel cell. J. Chem. Technol. Biotechnol. 2016, 91, 1802–1808. [Google Scholar] [CrossRef]
- Penteado, E.D.; Fernandez-Marchante, C.M.; Zaiat, M.; Gonzalez, E.R.; Rodrigo, M.A. Influence of carbon electrode material on energy recovery from winery wastewater using a dual-chamber microbial fuel cell. Environ. Technol. 2017, 38, 1333–1341. [Google Scholar] [CrossRef]
- Sciarria, T.P.; Merlino, G.; Scaglia, B.; D’Epifanio, A.; Mecheri, B.; Borin, S.; Licoccia, S.; Adani, F. Electricity generation using white and red wine lees in air cathode microbial fuel cells. J. Power Sources 2015, 274, 393–399. [Google Scholar] [CrossRef]
- Biovale. Available online: https://www.progettoager.it/index.php/settori/trasferimento-tecnologico-i-progetti/trasferimento-tecnologico-i-progetti-biovale (accessed on 28 February 2020).
- Insam, H.; Riddech, N.; Klammer, S. Microbiology of Composting; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Santos, M.; Diánez, F.; del Valle, M.G.; Tello, J.C. Grape marc compost: Microbial studies and suppression of soil-borne mycosis in vegetable seedlings. World J. Microbiol. Biotechnol. 2008, 24, 1493–1505. [Google Scholar] [CrossRef]
- Patti, A.F.; Issa, G.; Smernik, R.; Wilkinson, K. Chemical composition of composted grape marc. Water Sci. Technol. 2009, 60, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Viel, A.; Stellin, F.; Carlot, M.; Nadai, C.; Concheri, G.; Stevanato, P.; Squartini, A.; Corich, V.; Giacomini, A. Characteristics of Compost Obtained from Winemaking Byproducts. Waste Biomass Valoriz. 2018, 9, 2021–2029. [Google Scholar] [CrossRef] [Green Version]
- Majbar, Z.; Lahlou, K.; Ben Abbou, M.; Ammar, E.; Triki, A.; Abid, W.; Nawdali, M.; Bouka, H.; Taleb, M.; El Haji, M. Co-composting of Olive Mill Waste and Wine-Processing Waste: An Application of Compost as Soil Amendment. J. Chem. 2018, 2018, 7918583. [Google Scholar] [CrossRef]
- Excessive, K. Using composted grape marc in the vineyard. Water Sci. Technol. 2018, 60, 1265–1271. [Google Scholar]
- Diánez, F.; Santos, D.M.; Tello, J.C. Suppressive effects of grape marc compost on phytopathogenic oomycetes. Arch. Phytopathol. Plant Prot. 2007, 40, 1–18. [Google Scholar] [CrossRef]
Figure 1.
Scheme showing the different winemaking phases and displaying the critical points considered in the paper for enhancing sustainability. Symbols legend: (1) Sulfite reduction, (2) biodiversity improvement, (3) energy savings, (4) reuse and valorization of by-products.
Figure 1.
Scheme showing the different winemaking phases and displaying the critical points considered in the paper for enhancing sustainability. Symbols legend: (1) Sulfite reduction, (2) biodiversity improvement, (3) energy savings, (4) reuse and valorization of by-products.

© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nardi, T. Microbial Resources as a Tool for Enhancing Sustainability in Winemaking. Microorganisms 2020, 8, 507. https://doi.org/10.3390/microorganisms8040507
AMA Style
Nardi T. Microbial Resources as a Tool for Enhancing Sustainability in Winemaking. Microorganisms. 2020; 8(4):507. https://doi.org/10.3390/microorganisms8040507
Chicago/Turabian StyleNardi, Tiziana. 2020. "Microbial Resources as a Tool for Enhancing Sustainability in Winemaking" Microorganisms 8, no. 4: 507. https://doi.org/10.3390/microorganisms8040507
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.