High Resolution Melt Assays to Detect and Identify Vibrio parahaemolyticus, Bacillus cereus, Escherichia coli, and Clostridioides difficile Bacteria


Abstract
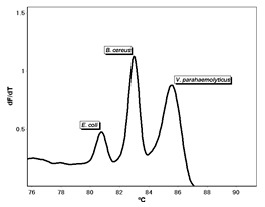
:
1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Bacterial Samples
2.2. DNA Extraction
2.3. Primer Design
2.4. Multiplex Assay Development
2.5. PCR Reaction Conditions
2.6. Sensitivity and Specificity Testing
2.7. Evaluation of Multiplex Assay for Bacterial Identification
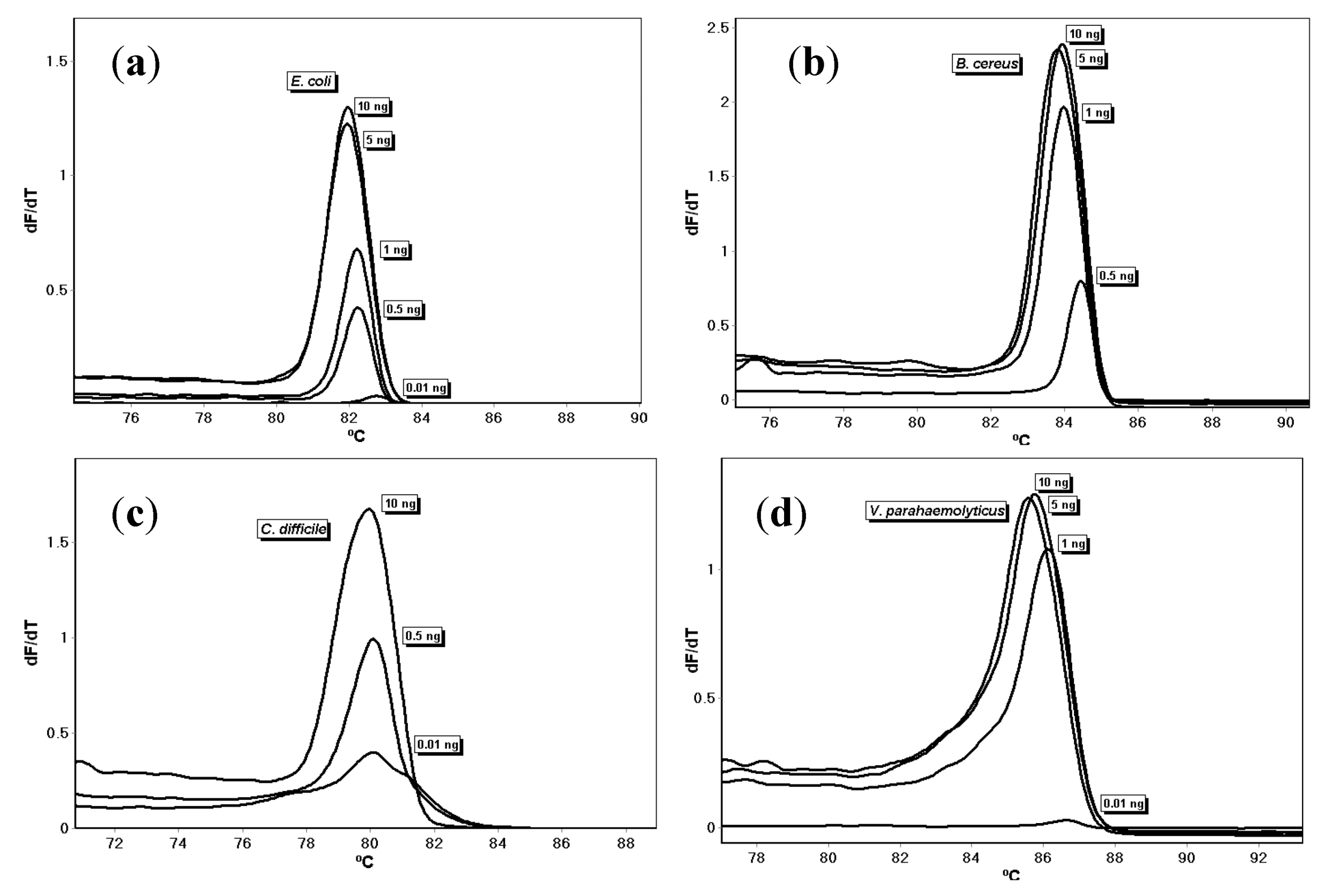
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Switai, T.L.; Winter, K.J.; Christensen, S.R. Diagnosis and Management of Foodborne Illness. Am. Fam. Physician 2015, 92, 358–365. [Google Scholar]
- Estimates of Foodborne Illness in the United States. Centers for Disease Control and Prevention. Available online: https://www.cdc.gov/foodborneburden/estimates-overview.html (accessed on 15 December 2019).
- Omer, M.K.; Álvarez-Ordoñez, A.; Prieto, M.; Skjerve, E.; Asehun, T.; Alvseike, O.A. A Systematic Review of Bacterial Foodborne Outbreaks Related to Red Meat and Meat Products. Foodborne Pathog. Dis. 2018, 15, 598–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candel-Pérez, C.; Ros-Berruezo, G.; Martínez-Graciá, C. A review of Clostridioides [Clostridium] difficile occurrence through the food chain. Food Microbiol. 2019, 77, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Rohlke, F.; Stollman, N. Fecal microbiota transplantation in relapsing Clostridium difficile infection. Therap Adv. Gastroenterol. 2012, 5, 403–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diarrhoeal Disease. World Health Organization. Available online: http://who.int/mediacentre/factsheets/fs330/en/index.html (accessed on 19 December 2019).
- Guerrant, R.L.; Van Gilder, T.; Steiner, T.S.; Thielman, N.M.; Slutsker, L.; Tauxe, R.V.; Hennessy, T.; Griffin, P.M.; DuPont, H.; Sack, R.B.; et al. Practice guidelines for the management of infectious diarrhea. Clin Infect. Dis. 2001, 32, 331–351. [Google Scholar] [CrossRef]
- Humphries, R.M.; Linscott, A.J. Laboratory diagnosis of bacterial gastroenteritis. Clin. Microbiol. Rev. 2015, 28, 3–31. [Google Scholar] [CrossRef] [Green Version]
- Ward, L.N.; Bej, A.K. Detection of Vibrio parahaemolyticus in Shellfish by Use of Multiplexed Real-Time PCR with TaqMan Fluorescent Probes. Appl. Environ. Microbiol. 2006, 72, 2031–2042. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, H.P.; Jaykus, L. Detection of pathogens in foods: The current state-of-the-art and future directions. Crit. Rev. Microbiol. 2011, 37, 40–63. [Google Scholar] [CrossRef]
- Alahi, M.E.E.; Mukhopadhyay, S.C. Detection Methodologies for Pathogen and Toxins: A Review. Sensors 2017, 17, 1885. [Google Scholar] [CrossRef] [Green Version]
- Murasova, P.; Kovarova, A.; Kasparova, J.; Brozkova, I.; Hamiot, A.; Pekarkova, J.; Dupuy, B.; Drbohlavova, J.; Bilkova, Z.; Korecka, L. Direct culture-free electrochemical detection of Salmonella cells in milk based on quantum dots-modified nanostructured dendrons. J. Electroanal. Chem. 2020, 863, 114051. [Google Scholar] [CrossRef]
- Sánchez-Chica, J.; Correa, M.M.; Aceves-Diez, A.E.; Castañeda-Sandoval, L.M. A novel method for direct detection of Bacillus cereus toxin genes in powdered dairy products. Int. Dairy J. 2020, 103, 104625. [Google Scholar] [CrossRef]
- Jayan, H.; Pu, H.; Sun, D.-W. Recent development in rapid detection techniques for microorganism activities in food matrices using bio-recognition: A review. Trends Food Sci. Technol. 2020, 95, 233–246. [Google Scholar] [CrossRef]
- Sharifi, S.; Vahed, S.Z.; Ahmadian, E.; Dizaj, S.M.; Eftekhari, A.; Khalilov, R.; Ahmadi, M.; Hamidi-Asl, E.; Labib, M. Detection of pathogenic bacteria via nanomaterials-modified aptasensors. Biosens. Bioelectron. 2020, 150, 111933. [Google Scholar] [CrossRef] [PubMed]
- Elkins, K.M. Introduction to Forensic Chemistry; Taylor and Francis/CRC Press: Boca Raton, FL, USA, 2018; pp. 251–286. [Google Scholar]
- Priyanka, B.; Patil, R.K.; Dwarakanath, S. A review on detection methods used for foodborne pathogens. Indian J. Med Res. 2016, 144, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Law, J.W.-F.; Mutalib, N.-S.A.; Chan, K.-G.; Lee, L.-H. Rapid methods for detection of foodborne bacterial pathogens: Principles, applications, advantages and limitations. Front. Microbiol. 2015, 5, 770. [Google Scholar] [CrossRef] [Green Version]
- Elkins, K.M.; Perez, A.C.U.; Sweetin, K.C. Rapid and inexpensive species differentiation using a multiplex real-time polymerase chain reaction high-resolution melt assay. Anal. Biochem. 2016, 500, 15–17. [Google Scholar] [CrossRef]
- Rajapaksha, P.; Elbourne, A.J.; Gangadoo, S.; Brown, R.; Cozzolino, D.; Chapman, J. A review of methods for the detection of pathogenic microorganisms. Analyst 2019, 144, 396–411. [Google Scholar] [CrossRef]
- Toma, C.; Lu, Y.; Higa, N.; Nakasone, N.; Chinen, I.; Baschkier, A.; Rivas, M.; Iwanaga, M. Multiplex PCR Asaay for Identification of Human Diarrhaegenic Escherichia coli. J. Clin. Microbiol. 2003, 41, 2669–2671. [Google Scholar] [CrossRef] [Green Version]
- Kagkli, D.-M.; Folloni, S.; Barbau-Piednoir, E.; Van den Eede, G.; Van den Bulcke, M. Towards a Pathogenic Escherichia coli Detection Platform Using Multiplex SYBR Green Real-Time PCR Methods and High Resolution Melting Analysis. PLoS ONE 2012, 7, e39287. [Google Scholar] [CrossRef] [Green Version]
- Harrison, L.B.; Hanson, N.D. High-Resolution Melting Analysis for Rapid Detection of Sequence Type 131 Escherichia coli. Antimicrob. Agents Chemother. 2017, 61, e00265-17. [Google Scholar] [CrossRef] [Green Version]
- Bélanger, S.D.; Boissinot, M.; Clairoux, N.; Picard, F.J.; Bergeron, M.G. Rapid Detection of Clostridium difficile in Feces by Real-Time PCR. J. Clin. Microbiol. 2003, 41, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natarajan, M.; Walk, S.T.; Young, V.B.; Aronoff, D.M. A Clinical and Epidemiological Review of Non-toxigenic Clostridium difficile. Anaerobe 2013, 22, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, B.M.; Hendriksen, N.B. Detection of Enterotoxic Bacillus cereus and Bacillus thuringiensis Strains by PCR Analysis. Appl. Environ. Microbiol. 2001, 67, 185–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blattner, F.R.; Plunkett, G., 3rd; Bloch, C.A.; Perna, N.T.; Burland, V.; Riley, M.; Collado-Vides, J.; Glasner, J.D.; Rode, C.K.; Mayhew, G.F.; et al. The complete genome sequence of Escherichia coli K-12. Science 1997, 277, 1453–1462. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez West, C.K.; Klein, S.L.; Lovell, C.R. High Frequency of Virulence Factor Genes tdh, trh, and tlh in Vibrio parahaemolyticus Strains Isolated from a Pristine Estuary. Appl. Environ. Microbiol. 2013, 79, 2247–2252. [Google Scholar] [CrossRef] [Green Version]
- Chumbler, N.M.; Farrow, M.A.; Lapierre, L.A.; Franklin, J.L.; Lacy, D.B. Clostridium difficile Toxins TcdA and TcdB Cause Colonic Tissue Damage by Distinct Mechanisms. Infect. Immun. 2016, 84, 2871–2877. [Google Scholar] [CrossRef] [Green Version]
- Helgasonm, E.; Okstad, O.A.; Caugant, D.A.; Johansen, H.A.; Fouet, A.; Mock, M.; Hegna, I.; Kolstø, A.B. Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis—One Species on the Basis of Genetic Evidence. Appl. Environ. Microbiol. 2000, 66, 2627–2630. [Google Scholar] [CrossRef] [Green Version]
- EcoCyc. Available online: https://biocyc.org/gene?orgid=ECOLI&id=G7043 (accessed on 19 December 2019).
- Standard Nucleotide BLAST. NCBI. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE_TYPE=BlastSearch (accessed on 19 December 2019).
- Chakraborty, S.; Alam, M.; Scobie, H.M.; Sack, D.A. Adaptation of a simple dipstick test for detection of Vibrio cholerae O1 and O139 in environmental water. Front. Microbiol. 2013, 29, 320. [Google Scholar]
- Proctor, L.M. The National Institutes of Health Human Microbiome Project. Semin. Fetal Neonatal Med. 2016, 21, 368–372. [Google Scholar] [CrossRef]
- Proctor, L. Priorities for the next 10 years of human microbiome research. Nature 2019, 569, 623–625. [Google Scholar] [CrossRef] [Green Version]
- The Integrative HMP (iHMP) Research Network Consortium. The Integrative Human Microbiome Project: Dynamic analysis of microbiome-host omics profiles during periods of human health and disease. Nature 2019, 569, 641–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
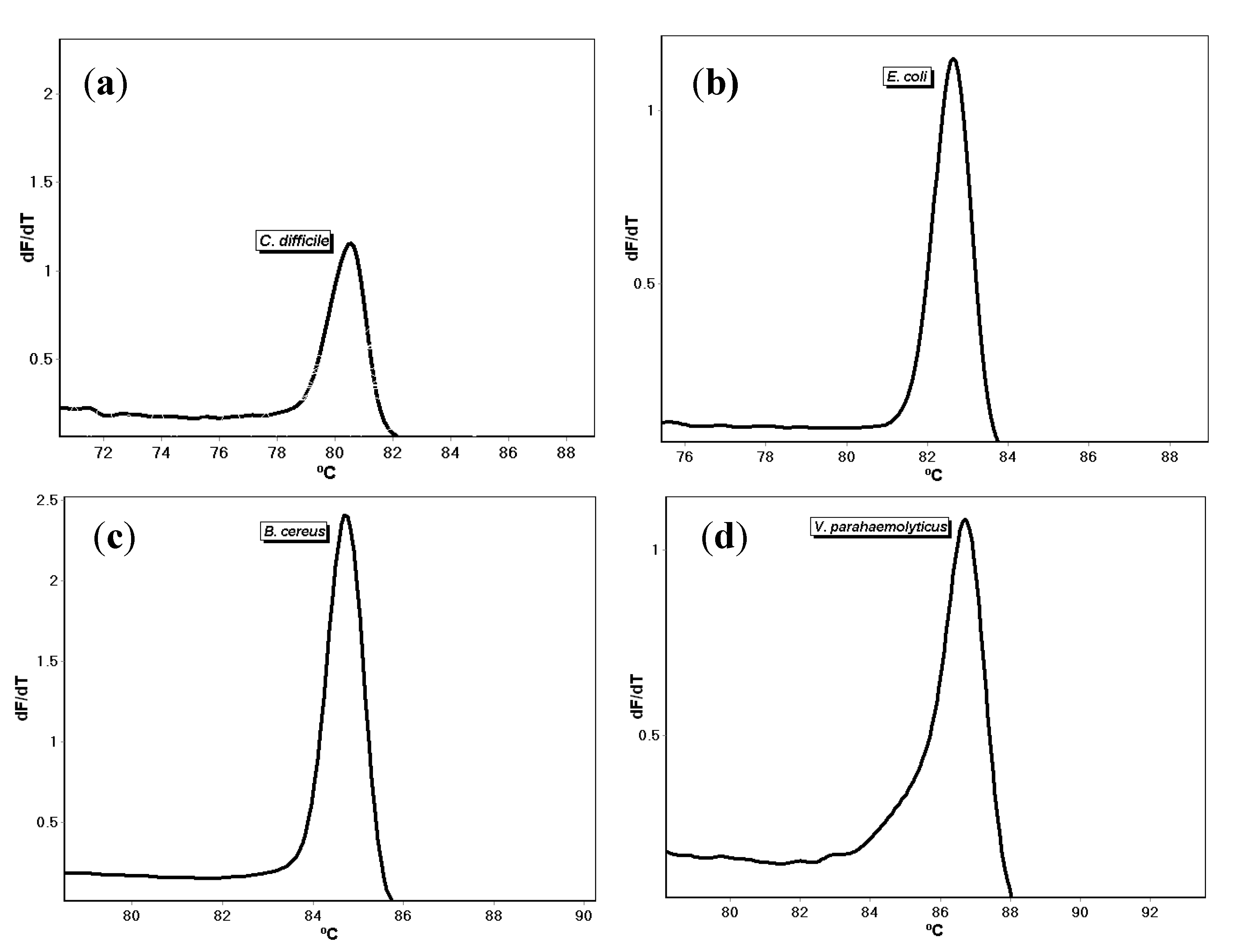
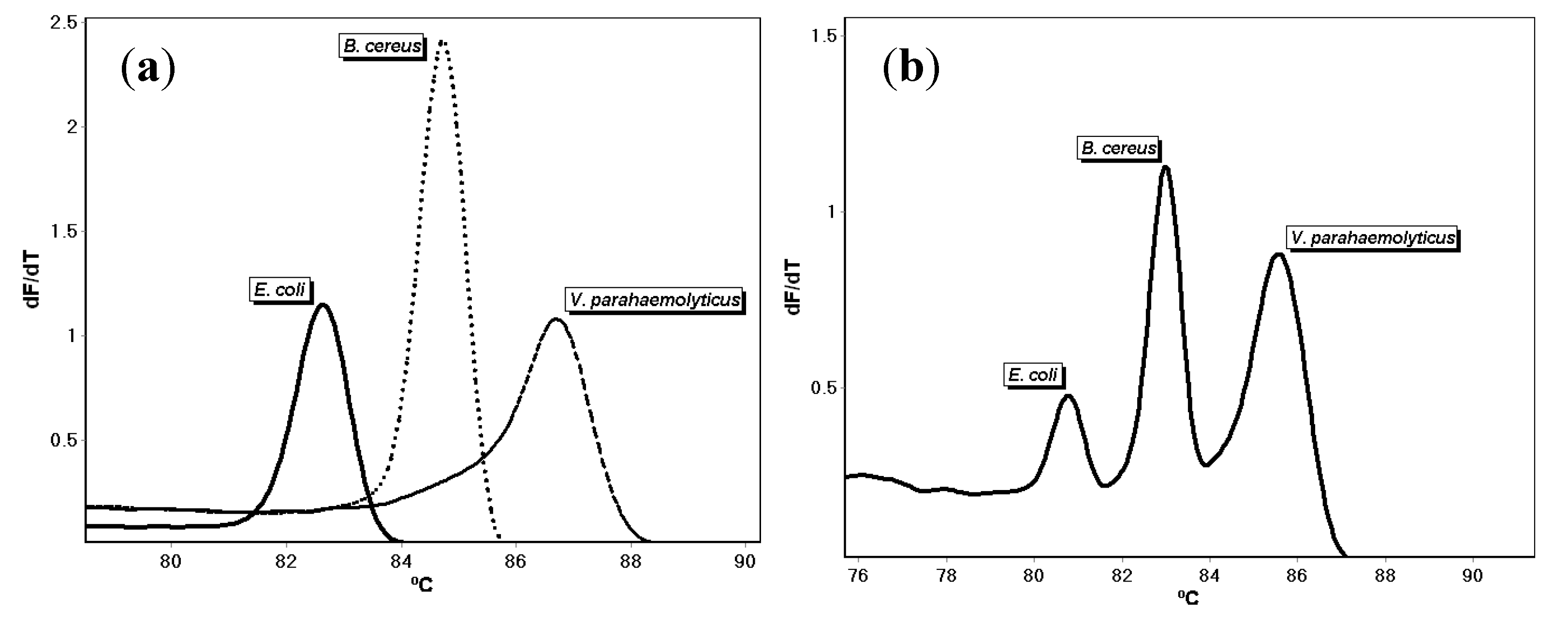
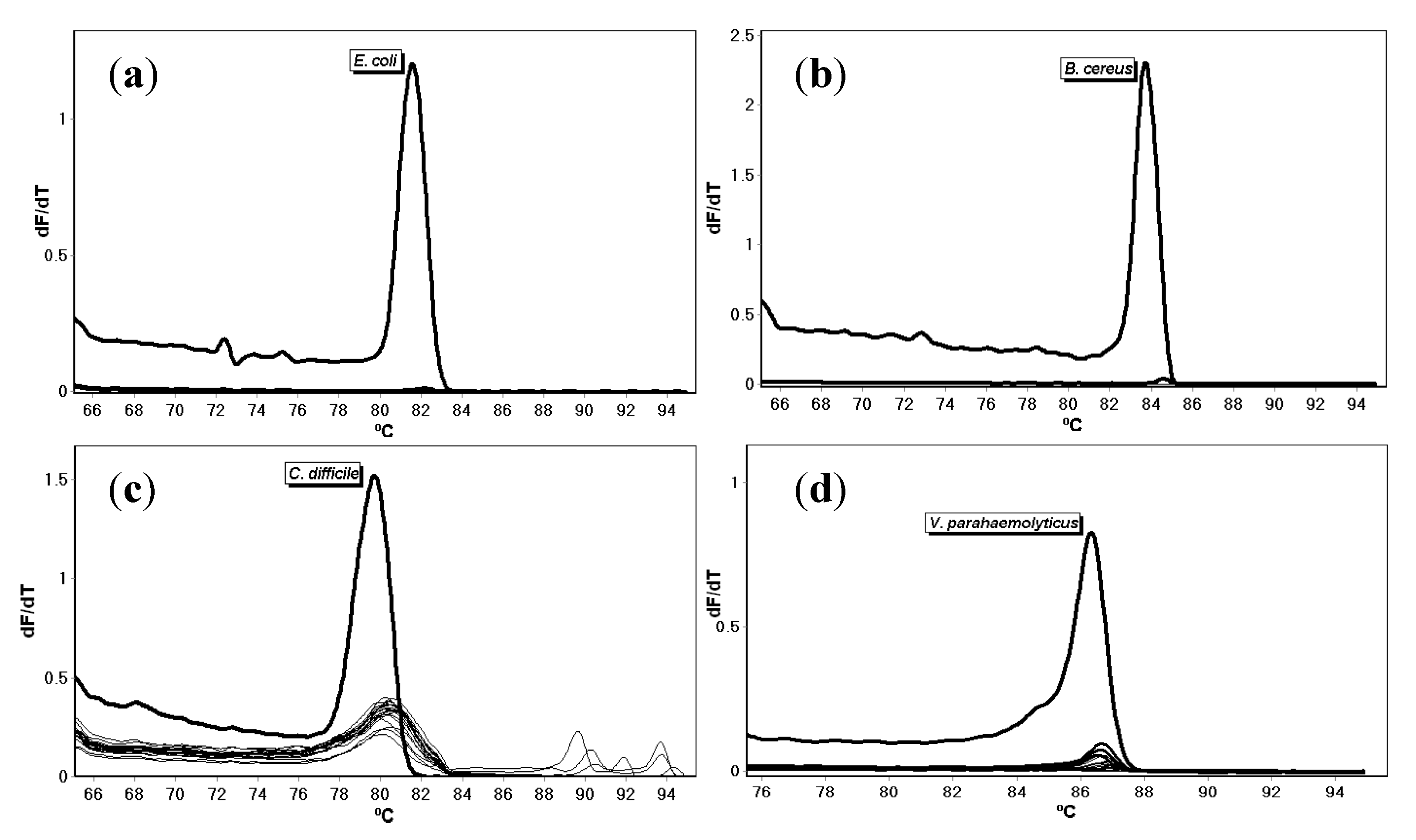

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterium | Source |
|---|---|
| Bacillus cereus str. Frankland and Frankland | ATCC (10876) |
| Bacillus megaterium | TU Biology Department |
| Bacillus subtilis str. Ehrenberg Cohn Str. 168 | ATCC (23857) |
| Bacillus thurigiensis serovar Israelensis | ATCC (35646) |
| Citrobacter freudii | Carolina Biological |
| Clostridioides difficile 90556-M6S | ATCC (9689D-5) |
| Enterobacter aerogenes | TU Biology Department |
| Escherichia coli K-12 MG1655 | ATCC (700926) |
| Escherichia coli (Migula) Castellani and Chalmers | ATCC (25922) |
| Klebsiella oxytoca MsA1 | ATCC |
| Listeria monocytogenes str. EGD | ATCC (BAA-679D-5) |
| Micrococcus luteus | Midwest Culture Service |
| Salmonella enterica serovar Typhimurium str. LT2 | ATCC (700720) |
| Serratia macrescens | Carolina Biological |
| Shigella flexneri str. 2547T | ATCC (29903) |
| Staphylococcus capitis subsp. capitis Kloos and Schleifer | ATCC (35661) |
| Staphylococcus saprophyticus | Wards Natural Science |
| Vibrio parahaemolyticus str. EB 101 | ATCC (17802D-5) |
| Target Specie | Primer Sequence (5’–3’) | Gene Target | Amplicon Length (bp) | Reference |
|---|---|---|---|---|
| C. difficile | F-GTTAGCATCCGTATTAGCAGGTGC R-ACAGCTATGGGTGCGAATGG | tcdA | 135 | This study |
| E. coli | F-TCCTGGATTGAGGTGCTTTATC R-CTACGGAGACCTGGGTAATTCC | yedN | 142 | Elkins et al. [19] |
| V. parahaemolyticus | F-ACTGGATTTCGCTTTGCCCTCAATGA R-GTTCTGAGTTCGATAACCTCTTGTGTGGATTAAG | tlh | 146 | This study |
| B. cereus | F-GAAAAGTACAGTGGCAAAAGTTGTTGCG R-CGCTAACTCTTGCTGACGACGT | cmk | 313 | This study |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bender, A.C.; Faulkner, J.A.; Tulimieri, K.; Boise, T.H.; Elkins, K.M. High Resolution Melt Assays to Detect and Identify Vibrio parahaemolyticus, Bacillus cereus, Escherichia coli, and Clostridioides difficile Bacteria. Microorganisms 2020, 8, 561. https://doi.org/10.3390/microorganisms8040561
Bender AC, Faulkner JA, Tulimieri K, Boise TH, Elkins KM. High Resolution Melt Assays to Detect and Identify Vibrio parahaemolyticus, Bacillus cereus, Escherichia coli, and Clostridioides difficile Bacteria. Microorganisms. 2020; 8(4):561. https://doi.org/10.3390/microorganisms8040561
Chicago/Turabian StyleBender, Allison C., Jessica A. Faulkner, Katherine Tulimieri, Thomas H. Boise, and Kelly M. Elkins. 2020. "High Resolution Melt Assays to Detect and Identify Vibrio parahaemolyticus, Bacillus cereus, Escherichia coli, and Clostridioides difficile Bacteria" Microorganisms 8, no. 4: 561. https://doi.org/10.3390/microorganisms8040561