Molecular Characterization, Intra-Species Diversity and Abundance of Freshwater Plesiomonas shigelloides Isolates


1
SAMRC Microbial Water Quality Monitoring Centre, University of Fort Hare, Alice 5700, Eastern Cape, South Africa
2
Applied and Environmental Microbiology Research Group, Department of Biochemistry and Microbiology, University of Fort Hare, Alice 5700, Eastern Cape, South Africa
3
Department of Biological Sciences, University of Medical Sciences, Ondo City PMB 536, Ondo State, Nigeria
*
Author to whom correspondence should be addressed.
Microorganisms 2020, 8(7), 1081; https://doi.org/10.3390/microorganisms8071081
Submission received: 2 February 2020
/
Revised: 28 February 2020
/
Accepted: 3 March 2020
/
Published: 20 July 2020
(This article belongs to the Section Environmental Microbiology)
Abstract
:Molecular signatures of Plesiomonas shigelloides strain specific to pathogenic and nonpathogenic variants are not well established till present. There is a need for intra-species barcoding of P. shigelloides to aid infection control. This study aims at characterizing and assessing intra-species diversity and abundance of P. shigelloides isolated from three freshwaters in the Eastern Cape Province. The study used a Plesiomonas-specific PCR to characterize the isolates. Intra-species (dis)similarities were assessed using ERIC-PCR and (GTG)5-PCR techniques. The DNA fingerprints produced were electrophoresed, digitized, and documented via computer-assisted pattern analysis. The fingerprints were analyzed using neighbor-joining clustering (NJC) based on Euclidean similarity index. Results revealed 80%, 83.64%, and 80% of the water samples from Tyhume, Kat, and Kubusie rivers, respectively, positive for P. shigelloides isolation. The prevalence of P. shigelloides from sites ranged from 13.5% to 88.9%. NJC delineated 48 isolates to 8 clades (ERIC-fingerprints) and 34 isolates into 7 clades ((GTG)5-fingerprints). The relative abundance of unique strains ranged from 6.3% to 22.9% via the two methods. Both fingerprinting approaches have strain-differentiating potential for P. shigelloides, however ERIC-PCR possessed higher resolution (D = 37.46) advantage over (GTG)5-PCR (D = 29.64). In conclusion, the study achieved intra-species diversity and abundance of P. shigelloides from aquatic milieu and provide further opportunity for intra-species-specific barcoding.
1. Introduction
Molecular signatures of Plesiomonas strains specific to virulent and nonvirulent strains still defy understanding and remain a diagnostic challenge. With a clinical obligation to differentiate pathogenic strains from nonpathogenic variants, there is a need to understand intra-species/strain diagnostic features that would aid discrimination of a pathogenic strain of P. shigelloides from nonpathogenic counterparts. Generally, pathogenic strains of a bacterium species have unique genetic signature(s)/trait(s) that are not present in its nonpathogenic strains, thus, these signature(s)/trait(s) form the basis for differentiating virulent strains from avirulent strains. Presently, strain-typing of P. shigelloides irrespective of pathogenic potential relies on somatic (O) and flagella (H) antigens serotyping [1]. Improved development of P. shigelloides O/H antigens serotyping was advanced by Aldova [2] and Aldova and Schubert [3]. Unfortunately, a good number of P. shigelloides strains are not serotypeable [4]. Aside from the inability to serotype some strains, P. shigelloides also cross-react with Shigella species, resulting in a false-positive reaction [5]. Hence, serology is not a reliable way to type P. shigelloides strains. Also, P. shigelloides antisera is not commercially available [6]. The need for rapid turnaround diagnostics in both strain-typing and pathogenic strain delineation of P. shigelloides is eminent for infection control. A more recent method is still in the pipeline for detecting 12 serovars of P. shigelloides involving O-antigen-specific suspension array-based molecular typing [7] and still has the inability to surmount pathogenic and nonpathogenic discernment.
According to González-Rey et al. [8], genomic diversity of P. shigelloides isolates that shared similar serotype data from different sources has been reported using fingerprinting techniques. González-Rey et al. [8] disclosed that the pulsed-field gel electrophoresis (PFGE), random amplified polymorphic DNA-PCR (RAPD), repetitive extragenic palindromic-PCR (rep-PCR), and enterobacterial repetitive intergenic consensus-PCR (ERIC-PCR) delineated 24 strains involved in the study into 22, 21, 19, and 17 genotypes respectively, based on manual examination, thus suggesting the inadequacy in the use of serotyping in P. shigelloides strain discrimination. Shigematsu et al. [9] examined strain heterogeneity of P. shigelloides isolates using PFGE and DNA macrorestriction analysis in an epidemiological study of P. shigelloides travelers’ diarrhea in Kansai Airport, Japan. The study initially aimed at discriminating and detecting strains associated with disease severity and/or most prevalent in the specific locations. Although Shigematsu et al. [9] showed strain heterogeneity of the 39 P. shigelloides isolates and 3 reference strains via their PFGE fingerprints (each isolate had a unique profile and was highly variable), strain-specific signatures connected to disease severity were not reported or discoverable by the DNA profiles. Molecular fingerprinting only provides clues of strains’ diversities in the P. shigelloides isolates. In a similar manner, Gu and Levin [10] studied strain diversity among 6 fish, 10 freshwater’s samples, and 10 human clinical isolates of P. shigelloides using RAPD analysis. Genetic variability was notable among the majority of the isolates, unlike composite RAPD profiles. Some of the most outstanding results from the Gu and Levin [10] studies include notable similar RAPD profiles of isolates from the same source, similar profiles of the fish and the human clinical isolates, certain strains of the human clinical isolates with specific features, and genotypic similarity of freshwater and fish isolates, thus highlighting the possibility of cross-transmission among the various matrices. Another known method for assessing strain diversity such as 16S-23S ITS (Internal Transcribed Spacer) region in many microorganisms was further reported to be unable to detect heterogeneity amongst P. shigelloides strains [9]. Also, matrix-assisted laser-desorption/ionization time-of-flight mass spectrometry (MALDITOF MS) instrumental methods have been used for proteomics, structural and molecular characterization of P. shigelloides’ lipopolysaccharide (LPS), and serotype (strain) delineation based on the O-PS side chain of LPS-variable chemical units as well as genetic variability in the wb cluster genes [11]. However, it has been shown that MALDITOF MS characterization of P. shigelloides lacks similarity between the same sero-strains [12].
Some authors believed the superiority of RAPD and PFGE as rapid, simple, and veritable sub-species typing techniques in strain-typing compared with restriction enzyme analysis, multilocus enzyme electrophoresis, and ribotyping [13], while many others have questioned their usefulness and reproducibility in the study of pathogens [14]. Notwithstanding that RAPD, rep-PCR, ERIC-PCR, RAPD, and PFGE have been used on P. shigelloides in different studies [8,9,13], many cogent questions about its strain diagnostic features remain unanswered. There also exist some unverified hypotheses about its pathogenicity potential. For instance, the proposition that the pathogenicity potential and/or the ability to cause gastroenteritis is ubiquitous to P. shigelloides isolates on the basis of the high variability of Plesiomonas chromosomal DNA patterns requires critical evaluation [9].
Generally, DNA-based techniques have been proposed by some authors to be characterized by high discriminatory power, high throughput, low cost, and considerable reliability of strain classification as well as typing of Gram-negative and Gram-positive bacteria [15]. The rep-PCR fingerprinting is a method in bacterial taxonomy that has been effectively applied in classifying lactobacilli [16], mycobacteria [17], staphylococci [18], vancomycin-resistant Enterococcus faecium strains [17,19], streptomycetes [20], and E. faecium clinical strains [21,22].
P. shigelloides is a single-species genus in the family Enterobacteriaceae [23]. It is well known to cause infections such as travelers’ diarrhea, gastroenteritis, to severe extraintestinal infections [9,24,25,26,27]. Also, some foodborne and waterborne outbreaks have been solely credited to P. shigelloides with sound microbiological and epidemiological validation [28,29]). Particularly, the incidence of Plesiomonas travelers’ diarrhea increased from 23.2% to 77.8% between 1987 and 1999 at Kansai Airport, Japan [9]. These and many more attest to the need to identify virulence signatures in P. shigelloides and the need for virulent strains’ diagnostic features. Strain-specific or virulent strain-specific diagnostic features would give room for discriminating its virulent strains from avirulent variants. Presently, no suitable phenotypic or molecular methods have been described for P. shigelloides virulent strain diagnosis.
From the foregoing, there is a need to understand and identify diagnostic traits of virulent and avirulent strains of P. shigelloides. However, the present study aims at characterizing and assessing intra-species heterogeneity and abundance of P. shigelloides isolated from aquatic environments in the Eastern Cape Province using molecular methods. This was aimed at enabling grouping of the strains to facilitate comparative studies of different groups to uncover the groups’ diagnostic traits. This approach will hopefully assist in extensive studies relevant for determining strain-specific molecular signatures of P. shigelloides that may provide insights for future endeavors in specific tagging of virulent (pathogenic) strains from avirulent (nonpathogenic) variants. For, we hypnotized that virulent strains of P. shigelloides have diagnostic trait(s) different from avirulent ones as it is common in other pathogens.
2. Materials and Methods
2.1. Cultural Isolation of P. shigelloides from River Water
P. shigelloides strains were isolated from 165 river water samples collected from freshwater resources at various points where human activities were prominent (see detailed sampling point descriptions in Table 1) across three popular rivers, viz. Tyhume, Kat, and Kubusie, in the Eastern Cape, South Africa. Sampling was done monthly and sequentially from the same sampling locations throughout February to December 2017. Water samples were collected aseptically in 1 L sterile glass bottle. Standard serial dilution of the water samples was carried out according to the standard protocol of APHA (American Public Health Association) [30]. Then, 100 mL aliquots of the respective dilutions were filtered through a 0.45µ millipore filter (Ø 47 mm) [30]. Each membrane filter, according to the dilution, was plated aseptically on a pre-labeled dried plate of inositol brilliant green bile agar (IBGBA) (HiMedia Laboratories, Mumbai-400086, India) using sterile forceps. After a period of 24 h incubation at 39 °C, pink colonies on the plates were counted and recorded as presumptive P. shigelloides isolates. Some randomly selected pink colonies were further streaked on a fresh IBGBA to purify them, subsequently grown on nutrient agar, and assayed for oxidase enzyme production using oxidase strips [31]. Oxidase-positive isolates were stored on glycerol stocks (−80 °C) for further studies.
2.2. DNA Extraction and Molecular Characterization of P. shigelloides
The purified oxidase-positive isolates were re-streaked onto fresh nutrient agar plates and cultured overnight at 37 °C. Total DNA of the 24 h culture was extracted by direct boiling procedure [32]. Two to three single colonies of the 24 h culture were picked and reconstituted in 200 µL sterile distilled water by vortex using a vortex mixer (DiGiSystem Laboratory instruments INC, Taiwan, Republic of China (ROC). The reconstituted cells were washed repeatedly with three changes of sterile DH2O at 15,000 rpm/2 min. The final cell pellet was re-suspended in nuclease-free water and boiled (100 °C/10 min) using Dri-Block®DB-3D (Bibby Scientific LTD, Staffordshire, UK). The boiled cells suspension was then centrifuged at 15,000 rpm/10 min in a micro-centrifuge (HEMLE Labrtechnik GmbH, Germany) to separate cell debris. The supernatant (DNA) was collected in a sterile Eppendorf tube and stored in a freezer (−20 °C) until further use.
The presumptive isolates were characterized using Plesiomonas-specific 23S rRNA polymerase chain reaction [4]. The primer used was PS23FW3 5′-CTCCGAATACCGTAGAGTGCTATCC-3′ and PS23RV3 5′-CTCCCCTAGCCCAATAAC ACCTAAA-3′, with the expected amplicon product of 284 bp (Gu and Levin, 2006) [10]. The PCR reaction consisted in 10× reaction buffer (2.5 µL), dNTPs (1 µL of 2.5 mM), MgCl2 (1.25 µL of 50 mM), primers (0.6 µL each, 10 µM), Taq polymerase (0.1 µL of 5 u/µL), 2.5 µL deoxyribonucleic acid of each isolate, and 16.45 µL sterilized nuclease-free water in a 25 µL reaction volume. The negative control was devoid of any DNA template. The thermal scheme for the reaction included initiation (1 cycle, 95 °C, 5 min), 35 cycles of denaturation (94 °C, 1 min), annealing (68 °C, 1 min), and extension (72 °C, 1 min), and final extension (72 °C, 10 min). Five microliters (5 µL) of the amplicons from each tube was electrophoresed in a 1% w/v agarose gel (Laboratorois Conda, Madrid, Spain). The gel had 2 µL ethidium bromide (0.2 µg/mL) incorporated. All electrophoresis was performed in a TBE (Tris-borate-EDTA) buffer (pH 8.0) (0.089 M Tris, 0.089 M boric acid, and 0.002 M EDTA (ethylenediaminetetraacetic acid) at 100 V/45 min (2.22 V/min). A 100 bp DNA standard served as gene ruler for the gel. At the completion of electrophoresis, bands were visualized using UV trans-illumination and photographed.
2.3. Fingerprinting, Fingerprint Treatment, and Computer-Assisted Analysis
Molecular diversities of selected, confirmed P. shigelloides isolates were determined by rep-PCR [33]. Two sets of primers including ERIC1 5′-ATGTAAGCTCCTGGGGATTCAC-3′, ERIC2 5′-AAGTAAGTGACTGGGGTGAGCG-3′, and (GTG)5 5′-GTGGTGGTGGTGGTG-3′ were used. The reactions for the two sets of primers were carried out in 20 µL total reaction volumes, made up of 1 µL of ERIC primer or 2 µL of (GTG)5 primer, 1.5 µL of MgCl2, 10 µL of the master mix, 2 µL of isolate’s DNA template, and 3.5 µL of sterilized nuclease-free water. While the thermal programs for ERIC-PCR consisted of initiation (94 °C/6 min), 35 cycles of denaturation (94 °C/0.5 min), annealing (48 °C/1 min), extension (72 °C/5 min), and final extension (72 °C/7 min), the thermal program for (GTG)5-PCR likewise comprised initiation step (94 °C/4 min), 40 cycles of denaturation (94 °C/1 min), annealing (40 °C/2 min), and extension (72 °C/2 min); and final extension (72 °C/10 min). Five/ten microliters (5/10 µL) of the ERIC-PCR and (GTG)5-PCR products was electrophoresed using TakaRa Mupid-ONE (Takara Bio Inc, Shiga, Japan). A 1.5% agarose gel (Laboratorois Conda, Madrid, Spain) with 2 µL ethidium bromide (1 µg/mL; Sigma-Aldrich, USA) incorporated in a TBE buffer (pH 8.0) comprising 0.089 M boric acid, 0.089 M Tris, and 0.002 M EDTA) was used. Equivalent volume of the PCBIO ladder (PCR Biosystems Ltd., London, UK) made up of a mixture of marker sizes ranging from 100 bp to 10 kb was filled into terminal of the gel. The gel was visualized for DNA fingerprints and documented using a UV transilluminator.
All fingerprint images were digitized for a computer-assisted pattern analysis using GelJ version 2.0 software [34]. The DNA fingerprints occurrence matrices and molecular weight of ERIC2-PCR and (GTG)5-PCR bands were generated from their gel images using the unweighted pair group arithmetic mean algorithm at 1.0% tolerance level for quality control.
2.4. Assessment of Intra-Species/Strain Diversity of P. shigelloides Isolates
The band occurrence matrices of (GTG)5-PCR and ERIC-PCR fingerprints generated by the GelJ 2.0 software were imported into PAleontological Statistics Version 3.23 (PAST3.23) [35] for diversity studies. The absence/presence of a fingerprint across the isolates formed the basis for strain homogeneity or heterogeneity (associations) assessment. Firstly, dendrograms of the two matrices were created by neighbor-joining (NJ) [36] using a Euclidean similarity index (Equation (1)).
Here, NJ defines the distance between any strain pair i and j as dij that minimizes Q criterion (7.2).
where r is the current number of bands (representing strains) and the sums run on the band (strain) set. Secondly, the abundance of strains that form a clade was computed. Then, the study compared the resolution power of each fingerprinting technique using the Shannon’s index (H) and Simpson’s index based on the number of clades from the neighbor-joining clustering (NJC).
3. Results
Fifty-five samples were processed for each of the three sampled rivers during the eleven months (February–December 2017). From Tyhume, Kat, and Kubusie rivers, 80% (44/55), 83.64% (46/55), and 80% (44/55) of samples were positive for P. shigelloides, respectively. Overall, 134 (81.21%) out of 165 samples examined from the rivers yielded P. shigelloides (Figure 1).
Seven hundred and forty-eight randomly selected presumptive colonies of P. shigelloides were oxidase-positive (Table 2). Out of these, two hundred and eleven isolates (n = 211, 28.21%) were identified as P. shigelloides by P. shigelloides-specific 23S rRNA polymerase chain reaction. These 211 P. shigelloides isolates yielded a band of 284 bp that confirmed their identity as P. shigelloides [4]. The sites 2TY and 5KT contributed 1.42% (lowest) and 18.01% (highest) of the total P. shigelloides confirmed from the rivers, respectively. A representative P. shigelloides-specific 23S rRNA gel image is shown in Figure 2.
A representative ERIC-PCR fingerprint image is presented in Figure 3. The ERIC-PCR banding patterns of isolates ranged from 0 to 10 bands. Molecular weight of bands also varied from 380 to 5665 bp (Figure 3). Certain isolates did not produce any band and appeared not typeable by the ERIC-PCR. The ERIC-PCR fingerprints dendrogram constructed by NJ using a Euclidean similarity index is shown in Figure 4. All the isolates clustered together. However, eight clades of strains were observed. The clades were as follows, numbering from origin (0) along the horizontal axis: clade 1: 614 2KT, 617 4KT, 618 1KT, 609 1KT, 631 3KT, 633 3KT, 620 4KT, 626 3KT, 627 3KT, and 670 1Kb; clade 2: 646 1KT, 671 2Kb, 692 3TY, and 628 3KT; clade 3: 655 1KT, 612 2KT, and 632 3KT; clade 4: 690 4KB, 691 6TY, 629 3KT, 654 5KT, 696 3TY, 663 4TY, 613 2KT, 662 1TY, 611 2KT, and 640 2KT; clade 5: 679 1KB, 630 3KT, 645 1KT, 664 4TY, 666 1TY, and 667 1Kb; clade 6: 681 3TY, 685 3TY, 615 2KT, 639 2KT, and 642 2KT; clade 7: 635 2KT, 638 2KT, 651 5KT, 693 5TY, and 643 2KT; and clade 8: 675 5Kb, 6765Kb, 687 4KB, and 636 2Kb. The relative abundance (%) of the isolates that made up each clade was 22.9% (11/48), 8.3% (4/48), 6.3% (3/48), 22.8% (10/48), 12.5% (6/48), 10.4% (5/48), 10.4% (5/48), and 8.3% (4/48) for clade1, clade 2, clade 3, clade 4, clade 5, clade 6, clade 7, and clade 8, respectively. ERIC-PCR strain typing resolution power was estimated as 17.39 (H) and 37.46 (D).
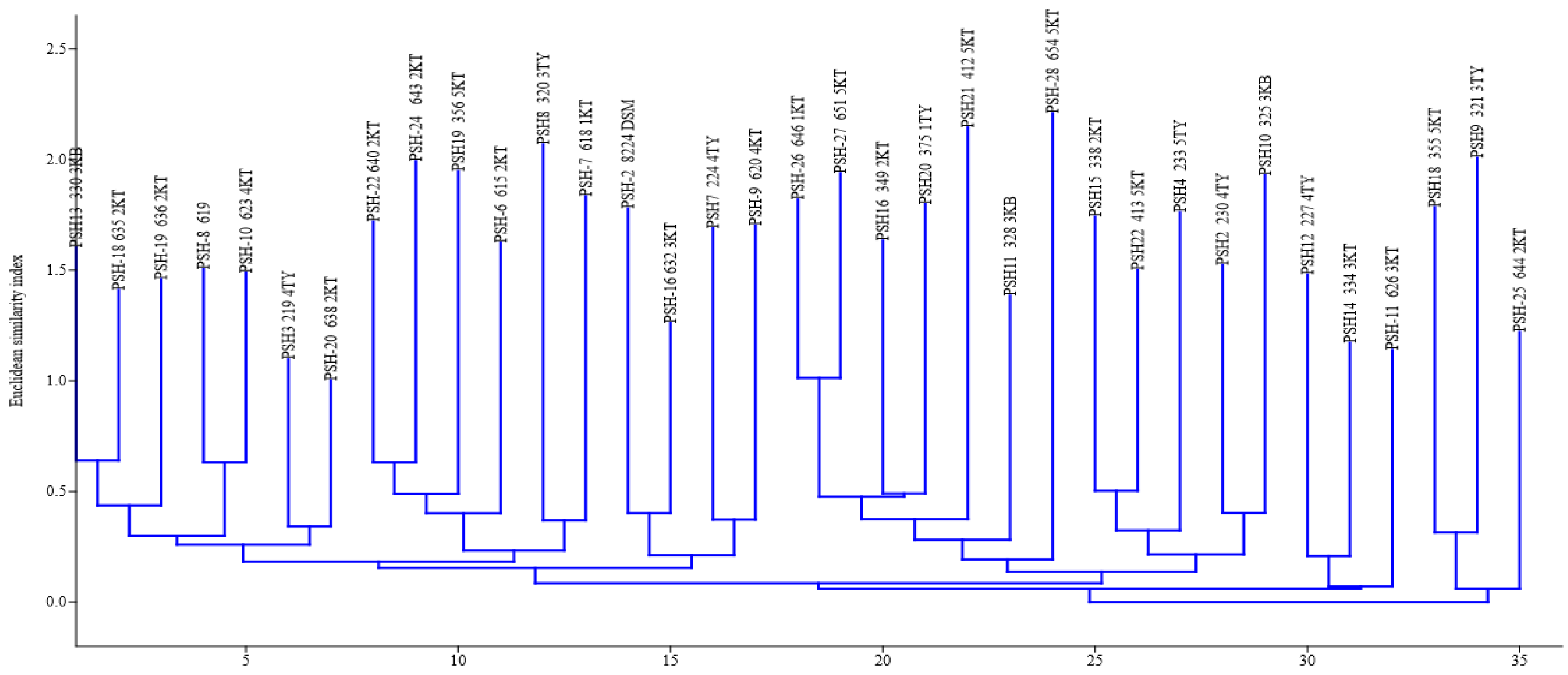
A typical digitized (GTG)5-PCR fingerprint image is presented in Figure 5. (GTG)5 fingerprints produced band patterns that ranged from 4 to 14. The (GTG)5 fingerprint bands ranged in size from 147.76 to 5304.98 bp. All the isolates produced (GTG)5 fingerprint bands and clustered together. The NJ clustering of (GTG)5 fingerprint resulted in seven clades. The composition of each clade, numbering from origin (0) along the horizontal axis to 35 scale, was as listed (Figure 6): clade 1: 330 2KB, 635 2KT, 636 2KT, PSH-8 619, 623 4KT, 219 4TY, and 638 2KT; clade 2: 640 2KT, 643 2KT, 356 5KT, 615 2KT, 320 3TY, and 618 1KT; clade 3: PSH-2 8224 DSM, 632 3KT, 224 4TY, and 620 4KT; clade 4: 646 1KT, 651 5KT, 349 2KT, 375 1TY, 412 5KT, 328 3KB, and 654 5KT; clade 5: 338 2KT, 413 5KT, 233 5KT, 233 5TY, 230 4TY, and 325 3KB; clade 6: 227 4TY, 334 3KT, and 626 3KT, and clade 7: 355 5KT, 321 3TY and 355 5KT, 321 3TY, and 644 2KT. The relative abundance (%) of strains making up each clade was 20.6% (7/34), 17.7% (6/34), 8.8% (3/34), 20.6% (7/34), 414.7% (7/34), 8.8% (3/34), and 8.8% (3/34) for clade 1, clade 2, clade 3, clade 4, clade 5, clade 6, and clade 7, respectively. (GTG)5-PCR fingerprint resolution power was calculated as 13.76 (H) and 29.64 (D).
4. Discussion
High detection (positive) rates of P. shigelloides were observed in the sampled waters in this present study. The P. shigelloides cultural prevalence rates of 80% (44/55), 83.64% (46/55), and 80% (44/55) observed in river water from Tyhume, Kat, and Kubusie, respectively, were higher than those in earlier reports from river water in literature.
The observed high recovery rate compared with previous studies in part could be attributed to a combined use of filtration technique and Inositol Brilliant Green Bile Agar for the isolation of P. shigelloides in this study. Most previous studies used some other media and culture techniques [37,38] that allowed P. shigelloides to compete with a host of other bacteria, which, in most cases, have higher growth advantages in the media compared with P. shigelloides. Some rates of positive detection of P. shigelloides from river, well, or pond water in other works include 40.7% [37], 16.67% (4/24) in Rio de Janeiro City in Brazil [38], 0.6% (well) and 7.4% (pond) in Zaria, Nigeria [39], 12.8% in Japan [40], and 13.3% in Dhaka, Bangladesh [41].
The overall P. shigelloides yield of 81.21% (134/165) from river water observed in this study could have arisen from probably organic and inorganic contaminants in the rivers that support the growth of P. shigelloides. For instance, herbicides, fertilizers, and pesticide input into the rivers occurs frequently at 3TY, 5TY, 2KT, 4KT, 3KB, and 5KB. More so, anthropogenic resuspension of the riverbeds, as frequently observed in the area during animal watering in the sites, might increase the levels of P. shigelloides in the overlying water because sediment resuspensions release P. shigelloides trapped in riverbeds, biofilms, and other matrices into the water column. P. shigelloides has been isolated from pond sediment, hydrophytes, and phytoplankton [41]. Some authors have reported isolation rates of P. shigelloides from freshwater sediment, hydrophytes, and phytoplankton as 29.2%, 20.8%, and 18.3%, respectively [41]. Islam et al. [41] noted that matrices associated with pond such as soil, sediment, phytoplankton and hydrophytes had 62.5%, 41.7%, and 33.3% P. shigelloides positive isolation rate in their study.
Generally, the choice of IBGA and membrane filtration in isolation encouraged the recovery performance of P. shigelloides in this study. The use of Plesiomonas isolation agar and membrane filtration for cultivation of P. shigelloides from the Nilufer Stream in Bursa, Turkey found an 83% (30/36) positive rate [42]. Most media for isolation of coliform and members of the Enterobacteriaceae allow the growth of P. shigelloides with varying recovery efficiencies and specificities. These media include deoxycholate-hydrogensulphide lactose agar [38], Salmonella-Shigella agar [34,43,44], MacConkey [43,45,46], Hektoen enteric agar [45], modified Salmonella-Shigella agar [40,44], deoxycholate citrate agar [46], taurocholate tellurite gelatin agar [41], xylose lysine deoxycholate agar [39,45], Endo agar [47], Plesiomonas differential agar [37,41], and peptone inositol bile and broth tetrathionate broth without iodine [37].
Another possible explanation for the high recovery rate of P. shigelloides from the sampled rivers without enrichment in this study could be due to favorable water temperature. The average temperature of the rivers ranged from 4.7–25.8 °C during the sampling period. The relative high water temperatures, known to be favorable for P. shigelloides multiplication, may have resulted in the higher rate of detection. However, isolation of P. shigelloides from freshwater water samples in the temperate and colder regions of the world, such as Czech Republic [48], Hungary [37]; the Netherlands [49], Slovakia [50], subpolar region of Sweden [47,50], and Serbia [37], has been reported.
Also, insanitary activities such as in-stream flow of domestic wastewater (3KB), poultry wastewaters (2KB, 5KB), slaughterhouse wastewaters, piggery wastewater (2KB, 5KB), livestock manure and litters (throughout the sites except 1TY), wastewater treatment plant effluents (1KT and 5KB), leachates from manhole and community dumpsite (5KT, 5TY), and fertilizers applications (2KT, 5KB, 5TY) offer conditions that could have led to direct input and proliferation of microorganisms, including P. shigelloides, along the sampled waters. All sampled sites are livestock watering sites coupled with several other uses with the exception of 1TY (a swimming/recreational hotspot) [51].
The computer-assisted analysis of the ERIC-PCR and (GTG)5-PCR fingerprints of the P. shigelloides clustered the isolates into eight and seven clades, respectively, thus suggesting that the P. shigelloides from the freshwater resources possessed intra-species or strain diversity. The clustering of strains from different sampling sites together is suggestive of an evolutionary relationship. This is in agreement with [8], who reported that clustered strains originated from different matrices (human and animal sources) or geographical location depicts clonal association. Uniqueness of P. shigelloides strains from diarrheic travelers among the Japanese was also demonstrated by Shigematsu et al. [9] using PFGE and DNA macrorestriction digests. Identical DNA-based profile of two pairs of P. shigelloides strains from human and animal origin has also been reported [8]. While González-Rey et al. [8] used DNA-based techniques for comparative delineation of a population of single/the same serovar strains, our study did not consider serotypes of the isolates since it has been previously reported that some strains are not serotypeable. Also, subclade (strain clade) diversity was observed in some clades. This explains potential within-clade strain dissimilarity. Notable in this study is the inability of ERIC-PCR to type some strains. Other authors have reported inability of ERIC-PCR to type certain E. coli strains [52,53]. In the study of Prabhu et al. [52], 13 out of 40 E. coli isolates were not ERIC-PCR typeable, and Ramazanzadeh et al. [54] observed 25 of 230 E. coli isolates not typeable by ERIC-PCR. The González-Rey et al. [8] study involved strains that belonged to the same serovar and this might be accounted for the differences observed in our study that involved strains from undifferentiated serovars. (GTG)5-PCR yielded bands for all the strains and thus allowed the differentiation of all P. shigelloides strains more efficiently compared with ERIC-PCR.
The relative abundance of the isolates that made up each clade of strain varied significantly. This connotes differences in the occurrence of P. shigelloides strains in nature. Heterogeneity in P. shigelloides has been reported by many studies, even at the same serovar level [8,9,37]. However, establishing unique identities that discriminate pathogenic strains from nonpathogenic strains has been a challenge. A combination and integrated fingerprinting techniques will be needed to fully identify the intra-species signatures of P. shigelloides strains. Besides, strain-specific signatures are now being sought for P. shigelloides delineation [54]. Although the ability to cause diarrheal illness has been hypothesized to be universal among P. shigelloides’ strains from DNA-based fingerprints [9], the resolution of such procedures is not sufficient to support the assumption.
5. Conclusions
This study reports intra-species genetic diversity of P. shigelloides isolated from freshwaters in the Eastern Cape province, South Africa. The high detection rate of P. shigelloides was similar in the three sampled waters (Tyhume, Kat, and Kubusie). The overall P. shigelloides’ positive isolation rate observed in the study could be attributed to the high level of pollution along the sampled rivers’ courses. Also, abundance and occurrences of P. shigelloides strains vary in the aquatic milieu. Both fingerprinting approaches have intra-species/strain characterizing potential for P. shigelloides’ preliminary grouping purposes. However, ERIC-PCR possessed potentially superior resolution merits over (GTG)5-PCR in P. shigelloides intra-species typing.
Author Contributions
Conceptualization, T.E. and A.O.; methodology, T.E.; validation, T.E., and A.O.; formal analysis, T.E.; investigation, T.E. resources, A.O.; data curation, T.E.; writing—original draft preparation, T.E..; writing—review and editing, T.E. and A.O.; visualization, T.E.; supervision, A.O.; project administration, A.O.; funding acquisition, A.O. and T.E. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the South African Medical Research Council (SAMRC), the National Research Foundation, and The World Academy of Science (NRF-TWAS), grant numbers 99796 and 116382 and “The APC was funded by Govan Mbeki Research and Development Centre, University of Fort Hare.
Acknowledgments
The authors thank the South African Medical Research Council (SAMRC) and the National Research Foundation, The World Academy of Science (NRF-TWAS) for financial support (Grant Numbers: 99796 and 116382). Conclusions arrived at and opinions expressed in this article are those of the authors and are not necessarily to be attributed to SAMRC or NRF-TWAS.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Shimada, T.; Sakazaki, R. On the serology of Plesiomonas shigelloides. Jpn. J. Med. Sci. Biol. 1978, 31, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldova, E.V.A. Serovars of Plesiomonas shigelloides. Zentralbl. Bakteriol. 1994, 281, 38–44. [Google Scholar] [CrossRef]
- Aldova, E.; Schubert, R.H.W. Serotyping of Plesiomonas shigelloides—A tool for understanding ecological relationships. Med. Microbiol. Lett. 1996, 5, 33–39. [Google Scholar]
- González-Rey, C.; Svenson, S.B.; Bravo, L.; Rosinsky, J.; Ciznar, I.; Krovacek, K. Specific detection of Plesiomonas shigelloides isolated from aquatic environments, animals and human diarrhoeal cases by PCR based on 23S rRNA gene. FEMS Immunol. Med. Microbiol. 2000, 29, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, J.; Gosselin, F.; Ismaïl, J.; Lorange, M.; Lior, H.; Woodward, D. Evaluation of commercial antisera for Shigella serogrouping. J. Clin. Microbiol. 1995, 33, 1997–2001. [Google Scholar] [CrossRef] [Green Version]
- Ekundayo, T.C.; Okoh, A.I. Plesiomonas shigelloides seventy years of systematics and taxonomy in perspective of the present-day diagnostic demands. Res. J. Med. Sci. 2017, 11, 103–113. [Google Scholar] [CrossRef]
- Xi, D.; Wang, X.; Ning, K.; Liu, Q.; Jing, F.; Guo, X.; Cao, B. O-antigen gene clusters of Plesiomonas shigelloides serogroups and its application in development of a molecular serotyping scheme. Front. Microbiol. 2019, 10, 741. [Google Scholar] [CrossRef]
- González-Rey, C.; Siitonen, A.; Pavlova, A.; Ciznar, I.; Svenson, S.B.; Krovacek, K. Molecular evidence of Plesiomonas shigelloides as a possible zoonotic agent. Folia Microbiol. 2011, 56, 178. [Google Scholar] [CrossRef]
- Shigematsu, M.; Kaufmann, M.E.; Charlett, A.; Niho, Y.; Pitt, T.L. An epidemiological study of Plesiomonas shigelloides diarrhoea among Japanese travellers. Epidemiol. Infect. 2000, 125, 523–530. [Google Scholar] [CrossRef]
- Gu, W.; Levin, R.E. Factors affecting quantitative PCR assay of Plesiomonas shigelloides. Food Biotechnol. 2006, 20, 219–230. [Google Scholar] [CrossRef]
- Kaszowska, M.; Jachymek, W.; Lukasiewicz, J.; Niedziela, T.; Kenne, L.; Lugowski, C. The unique structure of complete lipopolysaccharide isolated from semi-rough Plesiomonas shigelloides O37 (strain CNCTC 39/89) containing (2S)-O-(4-oxopentanoic acid)-α-D-Glcp (α-D-Lenose). Carbohydr. Res. 2013, 378, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Kolínská, R.; Dřevínek, M.; Aldová, E.; Žemličková, H. Identification of Plesiomonas spp.: Serological and MALDI-TOF MS methods. Folia Microbiol. 2010, 55, 669–672. [Google Scholar] [CrossRef] [PubMed]
- González-Rey, C. ThesisStudies on Plesiomonas shigelloides Isolated from Different Environments. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2003; pp. 10–27. [Google Scholar]
- Giacometti, F.; Piva, S.; Vranckx, K.; De Bruyne, K.; Drigo, I.; Lucchi, A.; Manfreda, G.; Serraino, A. Application of MALDI-TOF MS for the subtyping of Arcobacter butzleri strains and comparison with their MLST and PFGE types. Int. J. Food Microbiol. 2018, 277, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Olive, D.M.; Bean, P. Principles and applications of methods for DNA-based typing of microbial organisms. J. Clin. Microbiol. 1999, 37, 1661–1669. [Google Scholar] [CrossRef] [Green Version]
- Gevers, D.; Huys, G.; Swings, J. Applicability of rep-PCR fingerprinting for identification of Lactobacillus species. FEMS Microbiol. Lett. 2001, 205, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cangelosi, G.A.; Freeman, R.J.; Lewis, K.N.; Livingston-Rosanoff, D.; Shah, K.S.; Milan, S.J.; Goldberg, S.V. Evaluation of a high-throughput repetitive-sequence-based PCR system for DNA fingerprinting of Mycobacterium tuberculosis and Mycobacterium avium complex strains. J. Clin. Microbiol. 2004, 42, 2685–2693. [Google Scholar] [CrossRef] [Green Version]
- Wieser, M.; Busse, H.J. Rapid identification of Staphylococcus epidermidis. Int. J. Syst. Evol. Microbiol. 2000, 50, 1087–1093. [Google Scholar] [CrossRef] [Green Version]
- Dunne, W.M.; Wang, W. Clonal dissemination and colony morphotype variation of vancomycin-resistant Enterococcus faecium isolates in metropolitan Detroit, Michigan. J. Clin. Microbiol. 1997, 35, 388–392. [Google Scholar] [CrossRef] [Green Version]
- Lanoot, B.; Vancanneyt, M.; Dawyndt, P.; Cnockaert, M.; Zhang, J.; Huang, Y.; Liu, Z.; Swings, J. BOX-PCR fingerprinting as a powerful tool to reveal synonymous names in the genus Streptomyces. Emended descriptions are proposed for the species Streptomyces cinereorectus, S. fradiae, S. tricolor, S. colombiensis, S. filamentosus, S. vinaceus and S. phaeopurpureus. Syst. Appl. Microbiol. 2004, 27, 84–92. [Google Scholar]
- Willems, R.J.; Top, J.; Braak van den, N.; van Belkum, A.; Endtz, H.; Mevius, D.; Stobberingh, E.; Van den Bogaard, A.; van Embden, J.D. Host specificity of vancomycin-resistant Enterococcus faecium. J. Infect. Dis. 2000, 182, 816–823. [Google Scholar] [CrossRef] [Green Version]
- Bedendo, J.; Pignatari, A.C.C. Typing of Enterococcus faecium by polymerase chain reaction and pulsed field gel electrophoresis. Braz. J. Med. Biol. Res. 2000, 33, 1269–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrity, G.M.; Winters, M.; Searles, D.B. Taxonomic outline of the prokaryotic genera. In Bergey’s Manual of Systematic Bacteriology; Garrity, G.M., Ed.; Springer-Verlag: New York, NY, USA, 2001; pp. 1–39. [Google Scholar]
- Xia, F.Q.; Liu, P.N.; Zhou, Y.H. Meningoencephalitis caused by Plesiomonas shigelloides in a Chinese neonate: Case report and literature review. Ital. J. Pediatr. 2015, 41, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, J.K.; Zhang, X.C.; Hack, J.B. Plesiomonas shigelloides meningitis in an adult in the ED. Am. J. Emerg. Med. 2016, 34, 1329.e1. [Google Scholar] [CrossRef] [PubMed]
- Novoa-Farías, O.; Frati-Munari, A.C.; Peredo, M.A.; Flores-Juárez, S.; Novoa-García, O.; Galicia-Tapia, J.; Romero-Carpio, C.E. Susceptibilidad de las bacterias aisladas de infecciones gastrointestinales agudas a la rifaximina y otros agentes antimicrobianos en México. Rev. Gastroenterol. México. 2016, 81, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hustedt, J.W.; Ahmed, S. Plesiomonas shigelloides Periprosthetic Knee Infection After Consumption of Raw Oysters. Am. J. Orthop. (Belle Mead, NJ) 2017, 46, E32–E34. [Google Scholar] [PubMed]
- Wouafo, M.; Pouillot, R.; Kwetche, P.F.; Tejiokem, M.C.; Kamgno, J.; Fonkoua, M.C. An acute foodborne outbreak due to Plesiomonas shigelloides in Yaounde, Cameroon. Foodborne Pathog. Dis. 2006, 3, 209–211. [Google Scholar] [CrossRef]
- Wang, S.; Duan, H.; Zhang, W.; Li, J.W. Analysis of bacterial foodborne disease outbreaks in China between 1994 and 2005. FEMS Immunol. Med. Microbiol. 2007, 51, 8–13. [Google Scholar] [CrossRef] [Green Version]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater, 21st ed.; American Public Health Association, American Water Works Association, Water Environment Federation: Washington, DC, USA, 2005. [Google Scholar]
- Public Health England. Identification of Vibrio and Aeromonas Species. UK Standards for Microbiology Investigations. ID 19 Issue 3. 2015. Available online: https://www.gov.uk/ukstandards-for-microbiology-investigations-smi-quality-andconsistency-in-clinical-laboratories (accessed on 6 January 2020).
- Salerno, A.; Delétoile, A.; Lefevre, M.; Ciznar, I.; Krovacek, K.; Grimont, P.; Brisse, S. Recombining population structure of Plesiomonas shigelloides (Enterobacteriaceae) revealed by multilocus sequence typing. J. Bacteriol. 2007, 189, 7808–7818. [Google Scholar] [CrossRef] [Green Version]
- Versalovic, J.; Schneider, M.; De Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell. Biol. 1994, 5, 25–40. [Google Scholar]
- Heras, J.; Domínguez, C.; Mata, E.; Pascual, V.; Lozano, C.; Torres, C.; Zarazaga, M. GelJ–a tool for analyzing DNA fingerprint gel images. BMC Bioinform. 2015, 16, 270. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST-palaeontological statistics, ver. 1.89. Palaeontol. Electron 2001, 4, 1–9. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Petrušić, M.; Vidaković, D.O.; Lazić, S.; Radnović, D.; Knežević, P. Prevalence and genetic variability of Plesiomonas shigelloides in temperate climate surface waters of the Pannonian Plain. Arch. Biol. Sci. 2018, 70, 099–108. [Google Scholar] [CrossRef]
- Mondino, S.S.B.D.; Nunes, M.P.; Ricciardi, I.D. Occurrence of Plesiomonas shigelloides in water environments of Rio de Janeiro city. Memórias Inst. Oswaldo Cruz 1995, 90, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Kwaga, J.K.P.; Adesiyun, A.A.; Abdullahi, S.U.; Bello, C.S.S. Prevalence of salmonellae, shigellae and Plesiomonas shigelloides in dogs in Zaria, Nigeria. Br. Vet. J. 1989, 145, 174–177. [Google Scholar] [CrossRef]
- Arai, T.; Ikejima, N.; Itoh, T.; Sakai, S.; Shimada, T.; Sakazaki, R. A survey of Plesiomonas shigelloides from aquatic environments, domestic animals, pets and humans. Epidemiol. Infect. 1980, 84, 203–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.S.; Alam, M.J.; Khan, S.I. Distribution of Plesiomonas shigelloides in various components of pond ecosystems in Dhaka, Bangladesh. Microbiol. Immunol. 1991, 35, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Bülger, B. Occurrence of Plesiomonas shigelloides and relationship with faecal pollution in Nilufer Stream, Bursa, Turkey. Turk. Electron J. Biotechnol. 2004, 2, 22–29. [Google Scholar]
- Holmberg, S.D.; Farmer, J.J., III. Aeromonas hydrophila and Plesiomonas shigelloides as causes of intestinal infections. Rev. Infect. Dis. 1984, 6, 633–639. [Google Scholar] [CrossRef]
- Tsukamoto, T.; Kinoshita, Y.; Shimada, T.; Sakazaki, R. Two epidemics of diarrhoeal disease possibly caused by Plesiomonas shigelloides. Epidemiol. Infect. 1978, 80, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Penn, R.G.; Giger, D.K.; Knoop, F.C.; Preheim, L.C. Plesiomonas shigelloides overgrowth in the small intestine. J. Clin. Microbiol. 1982, 15, 869–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huq, M.I.; Islam, M.R. Microbiological and clinical studies in diarrhoea due to Plesiomonas shigelloides. Indian J. Med Res. 1983, 77, 793–797. [Google Scholar]
- Krovacek, K.; Eriksson, L.M.; González-Rey, C.; Rosinsky, J.; Ciznar, I. Isolation, biochemical and Serological characterisation of Plesiomonas shigelloides from freshwater in Northern Europe. Comp. Immunol. Microbiol. Infect. Dis. 2000, 23, 45–51. [Google Scholar] [CrossRef]
- Aldova, E.; Melter, O.; Chýle, P.; Slosarek, M.; Kodym, P. Plesiomonas shigelloides in water and fish. Cent. Eur. J. Public Health 1999, 7, 172–175. [Google Scholar] [PubMed]
- Medema, G.; Schets, C. Occurrence of Plesiomonas shigelloides in surface water: Relationship with faecal pollution and trophic state. Zent. Hyg. Umweltmed. Int. J. Hyg. Environ. Med. 1993, 194, 3. [Google Scholar]
- Gonzalez-Rey, C.; Svenson, S.B.; Eriksson, L.M.; Ciznar, I.; Krovacek, K. Unexpected finding of the” tropical” bacterial pathogen Plesiomonas shigelloides from lake water north of the Polar Circle. Polar Biol. 2003, 26, 495–499. [Google Scholar] [CrossRef]
- Prabhu, V.; Isloor, S.; Balu, M.; Suryanarayana, V.V.S.; Rathnamma, D. Genotyping by ERIC-PCR of Escherichia coli isolated from bovine mastitis cases. Indian J. Biotechnol. 2010, 9, 298–301. [Google Scholar]
- Ramazanzadeh, R.; Zamani, S.; Zamani, S. Genetic diversity in clinical isolates of Escherichia coli by enterobacterial repetitive intergenic consensus (ERIC)–PCR technique in Sanandaj hospitals. Iran. J. Microbiol. 2013, 5, 126. [Google Scholar]
- Ekundayo, T.C.; Okoh, A.I. A global bibliometric analysis of Plesiomonas-related research (1990–2017). PloS ONE 2018, 13, e0207655. [Google Scholar] [CrossRef] [Green Version]
- Ekundayo, T.C.; Okoh, A.I. Modelling the effects of physicochemical variables and anthropogenic activities as ecological drivers of Plesiomonas shigelloides distribution and freshwaters quality. Sci. Total Environ. 2019, 682, 765–778. [Google Scholar] [CrossRef]
Figure 1.
Heatmap of cultural detection of P. shigelloides across months in water samples collected from selected rivers in the Eastern Province. Red color represents positive isolation of P. shigelloides, while blue color represents no isolation. 1KB, Stutt. Game Reserve*; 1KT, Seymour; 1TY, Hogsberg; 2KB, StuttTW1*; 2KT, Katberg; 2TY, Hillfoot*; 3KB, StuttVBridge1*; 3KT, Balfour; 3TY, Kayalethu; 4KB, StuttEbBridge*; 4KT, Blinkwater; 4TY, Binfield; 5KB, StuttFbridge*; 5KT, Fort Beaufort; 5TY, Melani. * Arbitrary name.
Figure 1.
Heatmap of cultural detection of P. shigelloides across months in water samples collected from selected rivers in the Eastern Province. Red color represents positive isolation of P. shigelloides, while blue color represents no isolation. 1KB, Stutt. Game Reserve*; 1KT, Seymour; 1TY, Hogsberg; 2KB, StuttTW1*; 2KT, Katberg; 2TY, Hillfoot*; 3KB, StuttVBridge1*; 3KT, Balfour; 3TY, Kayalethu; 4KB, StuttEbBridge*; 4KT, Blinkwater; 4TY, Binfield; 5KB, StuttFbridge*; 5KT, Fort Beaufort; 5TY, Melani. * Arbitrary name.

Figure 2.
A representative P. shigelloides-specific 23S rDNA gel image. Line 1 = Positive control (P. shigelloides DSM 8224); lines 2 to 12 = positive isolates.
Figure 2.
A representative P. shigelloides-specific 23S rDNA gel image. Line 1 = Positive control (P. shigelloides DSM 8224); lines 2 to 12 = positive isolates.

Figure 3.
A representative ERIC-PCR fingerprint gel image. A = 1kb molecular ladder, lines 1 to 38 = P. shigelloides isolates.
Figure 3.
A representative ERIC-PCR fingerprint gel image. A = 1kb molecular ladder, lines 1 to 38 = P. shigelloides isolates.
Figure 4.
Neighbor-joining dendrogram clusters of ERIC-fingerprints of P. shigelloides strains. Isolates were encoded, for example, 32 696 3TY representing strain number (32), assigned number for PCR confirmed (696), and origin of the isolate (3TY).
Figure 4.
Neighbor-joining dendrogram clusters of ERIC-fingerprints of P. shigelloides strains. Isolates were encoded, for example, 32 696 3TY representing strain number (32), assigned number for PCR confirmed (696), and origin of the isolate (3TY).
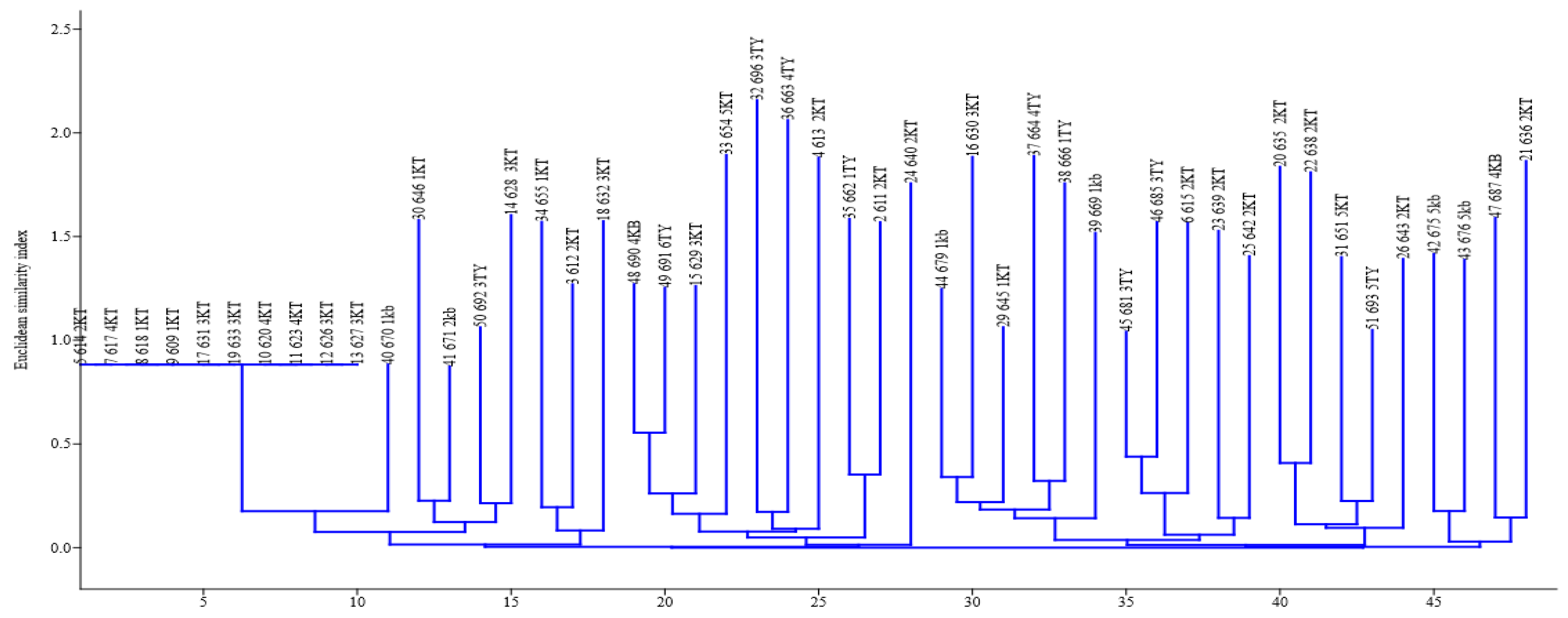
Figure 5.
A (GTG)5-PCR fingerprint digitized image. Isolates were encoded, for example, PSH-26 646 1KT, representing strain number (PSH-26), assigned number for PCR confirmed (646), and origin of the isolate (1KT).
Figure 5.
A (GTG)5-PCR fingerprint digitized image. Isolates were encoded, for example, PSH-26 646 1KT, representing strain number (PSH-26), assigned number for PCR confirmed (646), and origin of the isolate (1KT).

Figure 6.
A neighbor-joining dendrogram cluster of (GTG)5-PCR fingerprints of P. shigelloides strains. Isolates were encoded, for example, PSH-15 338 2KT, representing strain number (PSH-15), assigned number for PCR confirmed (338), and origin of the isolate (2KT).
Figure 6.
A neighbor-joining dendrogram cluster of (GTG)5-PCR fingerprints of P. shigelloides strains. Isolates were encoded, for example, PSH-15 338 2KT, representing strain number (PSH-15), assigned number for PCR confirmed (338), and origin of the isolate (2KT).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Coordinate and name descriptions of the sampling sites.
| Sites | Coordinates | Name |
|---|---|---|
| 1KB | S32°35.429′ E027°21.277′ | Stutt. Game Reserve * |
| 1KT | S32°32.891′ E026°45.835′ | Seymour |
| 1TY | S32°36.683′; E022°57.612′ | Hogsberg |
| 2KB | S32°35.249′ E027°23.916′ | StuttTW1 * |
| 2KT | S32°33.870′ E026°43.252′ | Katberg |
| 2TY | S32°36.652′ E026°54.564′ | Hillfoot * |
| 3KB | S32°35.278′ E027°24.35′ | StuttVBridge1 * |
| 3KT | S32°32.450′ E026°40.568′ | Balfour |
| 3TY | S32°38.374′ E026°56.163′ | Kayalethu |
| 4KB | S32°35.190′ E027°25.415′ | StuttEbBridge * |
| 4KT | S32°42.748′ E026°35.682′ | Blinkwater |
| 4TY | S32°40.980′ E026°54.080′ | Binfield |
| 5KB | S32°35.853′ E027°27.114′ | StuttFbridge * |
| 5KT | S32°46.753′ E026°37.250′ | Fort Beaufort |
| 5TY | S32°43.223′ E026°51.646′ | Melani |
* Arbitrary name.
Table 2.
Molecular confirmation of P. shigelloides among randomly selected isolates from IBGA plates.
Table 2.
Molecular confirmation of P. shigelloides among randomly selected isolates from IBGA plates.
| Sites | Coordinates | Name | Isolates | PCR+(%) | PCR+/211 × 100 |
|---|---|---|---|---|---|
| 1KB | S32°35.429′ E027°21.277′ | Stutt.Game Reserve * | 23 | 07(30.44) | 3.32 |
| 1KT | S32°32.891′ E026°45.835′ | Seymour | 89 | 12(13.5) | 5.69 |
| 1TY | S32°36.683′; E022°57.612′ | Hogsberg | 09 | 08(88.9) | 3.79 |
| 2KB | S32°35.249′ E027°23.916′ | StuttTW1 * | 30 | 06(20.0) | 2.84 |
| 2KT | S32°33.870′ E026°43.252′ | Katberg | 65 | 26(40.0) | 12.32 |
| 2TY | S32°36.652′ E026°54.564′ | Hillfoot * | 10 | 03 (30.0) | 1.42 |
| 3KB | S32°35.278′ E027°24.35′ | StuttVBridge1 * | 97 | 15(15.5) | 7.11 |
| 3KT | S32°32.450′ E026°40.568′ | Balfour | 30 | 20(66.7) | 9.48 |
| 3TY | S32°38.374′ E026°56.163′ | Kayalethu | 19 | 09(47.4) | 4.27 |
| 4KB | S32°35.190′ E027°25.415′ | StuttEbBridge * | 42 | 14(33.33) | 6.64 |
| 4KT | S32°42.748′ E026°35.682′ | Blinkwater | 54 | 13(24.1) | 6.16 |
| 4TY | S32°40.980′ E026°54.080′ | Binfield | 59 | 18(30.5) | 8.53 |
| 5KB | S32°35.853′ E027°27.114′ | StuttFbridge * | 23 | 07(30.4) | 3.32 |
| 5KT | S32°46.753′ E026°37.250′ | Fort Beaufort | 119 | 38(31.9) | 18.01 |
| 5TY | S32°43.223′ E026°51.646′ | Melani | 79 | 19(24.1) | 9.01 |
| Total | 748 | 211(28.21) | - |
* Arbitrary name.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ekundayo, T.; Okoh, A. Molecular Characterization, Intra-Species Diversity and Abundance of Freshwater Plesiomonas shigelloides Isolates. Microorganisms 2020, 8, 1081. https://doi.org/10.3390/microorganisms8071081
AMA Style
Ekundayo T, Okoh A. Molecular Characterization, Intra-Species Diversity and Abundance of Freshwater Plesiomonas shigelloides Isolates. Microorganisms. 2020; 8(7):1081. https://doi.org/10.3390/microorganisms8071081
Chicago/Turabian StyleEkundayo, Temitope, and Anthony Okoh. 2020. "Molecular Characterization, Intra-Species Diversity and Abundance of Freshwater Plesiomonas shigelloides Isolates" Microorganisms 8, no. 7: 1081. https://doi.org/10.3390/microorganisms8071081
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.