
Biotransformation of Flavonoids by Newly Isolated and Characterized Lactobacillus pentosus NGI01 Strain from Kimchi


1
School of Biological Sciences and Technology, Chonnam National University, 77 Yongbongro, Gwangju 61186, Korea
2
Department of Research and Development, Namhae Garlic Research Institute, 2465-8 Namhaedaero, Namhae 52430, Korea
*
Author to whom correspondence should be addressed.
Microorganisms 2021, 9(5), 1075; https://doi.org/10.3390/microorganisms9051075
Submission received: 26 April 2021
/
Revised: 13 May 2021
/
Accepted: 14 May 2021
/
Published: 17 May 2021
(This article belongs to the Section Microbial Biotechnology)
Abstract
:Lactic acid bacteria (LAB) are generally recognized as safe (GRAS) microorganisms. This study aimed to identify novel LAB strains that can transform flavonoids into aglycones to improve bioavailability. We isolated 34 LAB strains from kimchi. The biotransformation activity of these 34 LAB strains was investigated based on α-L-rhamnosidase and β-D-glucosidase activities. Among them, 10 LAB strains with high activities were identified by 16S rRNA sequencing analysis. All tested LAB strains converted hesperidin to hesperetin (12.5–30.3%). Of these, only the Lactobacillus pentosus NGI01 strain produced quercetin from rutin (3.9%). The optimal biotransformation conditions for the L. pentosus NGI01 producing hesperetin and quercetin were investigated. The highest final product concentrations of hesperetin and quercetin were 207 and 78 μM, respectively. Thus, the L. pentosus NGI01 strain can be a biocatalyst for producing flavonoid aglycones in the chemical and food industries.
1. Introduction
Biotransformation is a chemical modification of substances by organisms and enzymes. Whole-cell systems and isolated enzymes are widely used as biocatalysts in the chemical and food industries. Biotransformation is thought to be an eco-friendly and cost-effective alternative to conventional chemical synthesis [1,2].
Flavonoids are polyphenolic secondary metabolites with diverse chemical structures found in plants. They have been reported to have beneficial effects on human health, including antioxidant, anti-inflammatory, and anti-cancer activities [3,4,5]. Ingested flavonoids are metabolized and absorbed in the small intestine, colon, and liver; they are absorbed after deglycosylation in the small intestine. Flavonoids not absorbed in the small intestine are catabolized to aglycone and phenolic acids by the microbiota and are then absorbed in the colon [6,7,8]. Generally, the aglycone forms of flavonoids are more efficiently absorbed than flavonoid glycosides because of their ability to cross through the cell membrane [9,10,11]. However, the bioavailability of most natural flavonoids is low because they are conjugated with rutinose (6-O-α-L-rhamnosyl-d-glucose) or neohesperidoside (2-O-α-L-rhamnosyl-d-glucose), and human cells lack α-rhamnosidase activity [12,13].
α-l-rhamnosidase (EC 3.2.1.40) and β-d-glucosidase (EC 3.2.1.21) are glycosyl hydrolases that cleave terminal α-L-rhamnose and β-d-glucose, respectively, from several natural flavonoids [14,15,16]. For example, α-l-rhamnosidase converts hesperidin and rutin to hesperetin-7-O-glucoside and isoquercitrin, respectively. β-d-glucosidase then hydrolyzes these glycosides to hesperetin and quercetin, respectively (Figure 1). In the food industry, these enzymes have been applied for de-bittering fruit juices and enhancing the aroma of wine [17,18]. In addition, they are used in the production of flavonoid aglycones to increase bioavailability [10,19].
Lactic acid bacteria (LAB) are Gram-positive bacteria that produce lactic acid as the major end-product of carbohydrate fermentation. They also catabolize proteins and lipids during fermentation. LAB play important roles in the fermentation processes of foods such as cheese, yogurt, kimchi, and chongkukjang. Kimchi is a Korean tradition food, and LAB are used for kimchi fermentation [20,21,22]. LAB are generally recognized as safe (GRAS) microorganisms and can be used as safe biocatalysts. LAB strains have α-l-rhamnosidase and β-d-glucosidase activities involved in the deglycosylation of secondary metabolites during fermentation. Biotransformation using LAB has been applied for the hydrolysis of flavonoid glycosides based on these enzyme activities [13,23,24,25].
In the current study, LAB strains were isolated from kimchi, a traditional Korean fermented food, to develop novel LAB strains that could improve the bioavailability of flavonoids and be used as biocatalysts. The potential of LAB strains to transform flavonoids was evaluated by screening for the glycosyl hydrolase activity. In addition, LAB strains with a high glycosyl hydrolase activity were used as whole-cell biocatalysts for the biotransformation of hesperidin and rutin.
2. Materials and Methods
2.1. Materials
Buffered peptone water and De Man, Rogosa, and Sharpe (MRS) medium were purchased from Difco (Sparks, MD, USA). MRS medium without glucose was obtained from KisanBio (Seoul, Korea). Bromocresol purple, p-nitrophenyl β-d-glucopyranoside (pNPG), p-nitrophenyl α-l-rhamnopyranoside (pNPR), dimethyl sulfoxide (DMSO), hesperidin, hesperetin, rutin, and quercetin were purchased from Sigma-Aldrich (St. Louis, MO, USA). All chemicals were of analytical grade.
2.2. Isolation of LAB Strains from Kimchi
The cabbage kimchi and cucumber kimchi samples (homemade; 1 g) were cut with sterilized scissors and homogenized in 10 mL of buffered peptone water using BagMixer (Interscience, Saint-Nom la Bretèche Arpents, France). The homogenized sample was serially diluted to 10−5 in peptone water, and 100 μL of the dilution was spread on an MRS agar plate containing 0.0015% (w/v) bromocresol purple. After incubating at 30 °C for 24 h, yellow-colored colonies were selected. The colonies were inoculated into 10 mL of MRS medium and cultured at 30 °C until reaching an absorbance of 1 at 600 nm, which corresponded to 5 × 108 colony forming unit per mL (CFU/mL). The LAB strain stocks were prepared from these cultures and stored at −80 °C.
2.3. Glycosyl Hydrolase Activity Assay
The α-L-rhamnosidase and β-d-glucosidase activities of LAB strains isolated from kimchi were measured by following the protocol described in a previous study [26], with modifications. The cell stock (100 μL) was inoculated into 10 mL of MRS medium without glucose, supplemented with 1 mM pNPR or pNPG. After incubating at 30 °C for 3 days, the cultures were centrifuged at 3500× g for 20 min, and supernatants were used for the enzyme activity assay. The supernatant (2 mL) was mixed with 5 mL of 67 mM potassium phosphate buffer (pH 6.8) and 8 mL of 100 mM sodium carbonate. The absorbance of these mixtures was measured at 405 nm using an Epoch micro plate spectrophotometer (Biotek Instruments Inc., Winooski, VT, USA). The molar extinction coefficient for p-nitrophenol (pNP) at 405 nm was calculated to be ε = 18.3 mM−1 cm−1 [27]. The whole cells’ glycosyl hydrolase activities were presented as the final concentrations of the product (pNP, μM).
2.4. Identification of LAB Strains Isolated from Kimchi
The LAB strains isolated from kimchi were identified by 16S rRNA sequencing analysis. The cell stock was inoculated into MRS broth and incubated at 30 °C for 2 days. The cultures were sent to Macrogen (Seoul, Korea) for sequencing analysis. Genomic DNA of the LAB strains was amplified using two PCR primers (27F: 5′-AGAGTTTGATCMTGGCTCAG-3′; 1492R: 5′-TACGGYTACCTTGTTACGACTT-3′). The PCR product was purified using a Montage PCR Clean up kit (Millipore, Burlington, MA, USA). The purified PCR product was sequenced using two sequencing primers (85F: 5′-GGATTAGATACCCTGGTA-3′; 907R: 5′-CCGTCAATTCMTTTRAGTTT-3′). Sequencing was performed on the Big Dye Terminator Cycle Sequencing Kit v.3.1 (Applied BioSystems, Foster City, CA, USA). Sequencing products were resolved on an Applied Biosystems model 3730XL automated DNA sequencing system (Applied BioSystems, Foster City, CA, USA) at Macrogen. The 16S rRNA sequences of LAB strains were compared with data in GenBank using the Basic Local Alignment Search Tool (BLAST).
2.5. Biotransformation of Hesperidin and Rutin by LAB Strains
To investigate whether the LAB strains could convert hesperidin and rutin to their aglycone forms, the cell stocks (100 μL) were inoculated into MRS medium without glucose, supplemented with 1 mM hesperidin or rutin. After incubating at 30 °C for 7 days, the cultures were freeze-dried using a freeze dryer (Operon, Gimpo, Korea). The dried samples were dissolved in 10 mL of DMSO and then filtered using a 0.2 μm GHP syringe filter (Pall Corporation, Port Washington, NY, USA). The samples were analyzed using a 1260 DAD VL detector on an Agilent 1260 Infinity HPLC system (Agilent Technologies, Santa Clara, CA, USA). The samples of 20 µL were injected into an SB-C18 column (150 × 4.6 mm, 5 μm; Agilent Technologies, Santa Clara, CA, USA). The mobile phases were 0.1% formic acid in water (A) and 0.1% formic acid in acetonitrile (B). The samples were separated via a binary program with 5% B (0–1 min), 25% B (1–30 min), 30% B (30–34 min), 30% B (34–36 min), 53% B (36–50 min), 100% B (50–51 min), and 100% B (51–55 min) at a flow rate of 1 mL/min. Hesperidin and hesperetin were detected at 280 nm. Rutin and quercetin were detected at 340 nm. The retention times of hesperidin, hesperetin, rutin, and quercetin were 26.7, 42.6, 21.9, and 34.8 min, respectively (Figure S1). The molar amounts of substrates and products were calculated from the peak area of the standards. The product yield (%) was calculated using Equation (1) [28]:
2.6. Production of Hesperetin and Quercetin by L. pentosus NGI01 Strain
Biotransformation of hesperidin and rutin by NGI01 was performed at different concentrations. The cell stock (100 uL) was inoculated into MRS medium without glucose, supplemented with 100–400 μM hesperidin or rutin. After biotransformation at 30 °C for 0–21 days, the cultures were freeze-dried. The products were analyzed using HPLC as described above.
2.7. Statistical Analysis
All experiments were performed three times. The values are presented as the mean and standard error of the mean (SEM) from values of the replicates for each sample. Statistical analysis of the experimental results was carried out using Prism 5 (GraphPad Software, San Diego, CA, USA).
3. Results
3.1. Glycosyl Hydrolase Activity of LAB Strains from Kimchi
To identify strains that can biotransform flavonoids, we isolated 34 LAB strains from cabbage kimchi and cucumber kimchi. The α-l-rhamnosidase and β-d-glucosidase activities of isolated LAB strains were investigated using pNPR and pNPG, respectively, as substrates. The LAB strains were inoculated in MRS medium without glucose, supplemented with 1 mM pNPR or pNPG. After 3 days, the pNP produced by strains in the culture was measured. All isolated LAB strains produced over 150 μM pNP from pNPG. Among them, the NGI24 strain exhibited the highest activity, 268 μM. When pNPR was the substrate, 12 LAB strains (NGI01, NGI02, NGI06, NGI11, NGI12, NGI14, NGI17, NGI19, NGI21, NGI22, NGI23, and NGI25) produced pNP, and the NGI19 strain exhibited the highest activity, 78 μM (Figure 2).
3.2. Identification of LAB Strains from Kimchi
Among 34 LAB strains isolated from kimchi, 10 strains (NGI01, NGI02, NGI06, NGI14, NGI17, NGI19, NGI21, NGI22, NGI23, and NGI25) produced over 30 μM pNP from pNPR by α-l-rhamnosidase activity and more than 150 μM pNP from pNPG by β-d-glucosidase activity. Thus, these were selected for identification. Based on 16S rRNA sequencing analysis, NGI01, NGI06, NGI17, NGI19, NGI22, and NGI23 belonged to L. pentosus. NGI02, NGI14, NGI21, and NGI25 belonged to L. senmaizukei, L. plantarum, L. paraplantarum, and L. curvatus, respectively. The GenBank database accession numbers of the LAB strains are shown in Table 1.
3.3. Biotransformation of Hesperidin and Rutin by the L. pentosus NGI01 Strain
To investigate whether the 10 LAB strains could convert hesperidin and rutin to their aglycone forms, strains were cultured in MRS medium without glucose, supplemented with 1 mM hesperidin or rutin for 7 days. After whole-cell biotransformation with LAB, the yield of hesperetin and quercetin was analyzed by HPLC. All tested LAB strains converted hesperidin to hesperetin. Of these, the NGI01 strain produced the most hesperetin, with a yield of 30.3%. The hesperetin yield of the NGI06 strain was the second highest, 28.3%. The NGI23 strain yielded the least hesperetin, with a yield of 12.5%. When rutin was present in the culture, only NGI01 produced quercetin from rutin, with a product yield of 3.9% (Table 2).
3.4. Production of Hesperetin and Quercetin by L. pentosus NGI01 Strain
The optimal conditions to produce hesperetin and quercetin using the NGI01 strain were determined by varying the substrate concentration and biotransformation time. Hesperetin production increased with increasing hesperidin concentration and biotransformation time, reaching saturation after 10 days. The NGI01 strain produced the highest concentration of hesperetin, 207 μM, from 300 μM hesperidin after 10 days. The product yield of hesperetin under these conditions was 69.1% (Figure 3A). In addition, the NGI01 strain converted rutin to quercetin. Quercetin production increased with rutin concentration and biotransformation time and was saturated within 17 days. The most quercetin produced was 78 μM, obtained when rutin concentration was 400 μM, and the production yield of quercetin was 19.4% (Figure 3B). Thus, the optimal biotransformation conditions for producing hesperetin by the NGI01 strain were: MRS medium without glucose, supplemented with 300 μM hesperidin, at 30 °C, and for 10 days. The optimal conditions for quercetin production by NGI01 were: MRS medium without glucose, supplemented with 400 μM rutin, at 30 °C, and for 17 days.
4. Discussion
Flavonoids have a potential as biomaterials in the development of functional foods and medicines because they have beneficial effects on human health [4,5]. Most natural flavonoids are glycoside forms such as rutinose and neohesperidoside. To improve the bioavailability of flavonoids, ingestion of flavonoids in aglycone forms or activities of gut microbiota are necessary because α-rhamnosidase activity is absent in humans [8,13]. Since LAB are GRAS microorganisms and have both α-l-rhamnosidase and β-d-glucosidase activities [25], they can be biocatalysts for biotransformation of flavonoids to aglycones.
Here, LAB strains with high glycosidase activity were isolated from kimchi, a Korean traditional food, and identified by 16S rRNA sequence analysis (Table 1). All LAB strains exhibited β-d-glucosidase activity and 12 LAB strains showed α-l-rhamnosidase activity. The β-d-glucosidase activity was higher than α-l-rhamnosidase activity in tested LAB strains (Figure 2). These results suggest the LAB strains isolated from kimchi have a potential as whole-cell biocatalysts for glycosyl hydrolysis of flavonoid glucosides.
To investigate whether the LAB strains could biotransform flavonoids, hesperidin and rutin were used as substrates in whole-cell biotransformation. After whole-cell reactions, 10 LAB strains produced hesperetin from hesperidin and only the NGI01 strain produced quercetin from rutin (Table 2). We also tested whether naringin was converted to naringenin by the 10 LAB strains, but no naringenin was produced by the strains (data not shown). These results suggested that LAB strains isolated from kimchi showed different substrate specificity [16,29,30]. In addition, hesperetin-7-O-glucoside and isoquercitrin, products of α-L-rhamnosidase (Figure 1), were not detected in the whole-cell reactions using NGI01 (Figure S1). These results are related to Figure 2 which shows that the β-d-glucosidase activity of LAB strains was higher than α-l-rhamnosidase activity.
A previous study described the biotransformation of hesperidin and rutin using probiotics such as L. plantarum and L. acidophilus LA-5. When the concentration of hesperidin and rutin was 100 μM, the conversion rates of the two by L. acidophilus were 84% and 3%, respectively, after 10 days. L. plantarum yielded 70% hesperetin and 5% quercetin after 10 days [13]. In this study, the product yields of hesperetin and quercetin by the L. pentosus NGI01 strain after 10 days of biotransformation were 71.8% and 18.8% from 100 μM hesperidin and rutin, respectively (Figure 3). To investigate the stability of hesperidin and rutin, the flavonoids were incubated without whole cells for 21 days. The concentration of hesperidin was not apparently changed. Although the rutin concentration decreased over the time after 7 days of incubation, the product of biotransformation, quercetin, was not detected (Figures S2 and S3). In addition, cell growth was monitored during the NGI01-induced biotransformation. The growth of the NGI01 strain was stationary for 21 days in media containing hesperidin. In case of media containing rutin, the growth of the NGI01 strain decreased after 7 days, but quercetin, a product of biotransformation, was continuously produced after 7 days. (Figure S4 and Figure 3). These current results suggest that the L. pentosus NGI01 strain can produce hesperetin and quercetin with a high product yield. The L. pentosus NGI01 strain identified in this study was deposited at the Korean Culture Center of Microorganisms as KCCM12592P.
In conclusion, the L. pentosus NGI01 strain isolated from kimchi was a LAB strain and was a GRAS microorganism. This NGI01 strain had α-l-rhamnosidase and β-d-glucosidase activities. In addition, it produced high yields of hesperetin and quercetin from hesperidin and rutin, respectively. Therefore, the NGI01 strain can be applied as a safe whole-cell biocatalyst for chemical synthesis with applications in the food industry.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/microorganisms9051075/s1, Figure S1: Chromatogram of flavonoids after the biotransformation by the L. pentosus NGI01 strain, Figure S2: Stability of hesperidin and rutin, Figure S3: Chromatograms of flavonoids after incubation without whole-cell biocatalysts, Figure S4: Growth of the L. pentosus NGI01 strain during biotransformation.
Author Contributions
Conceptualization, C.-M.P. and G.-S.C.; methodology, C.-M.P. and G.-S.C.; software, C.-M.P. and G.-S.C.; validation, C.-M.P. and G.-S.C.; formal analysis, C.-M.P., G.-M.K. and G.-S.C.; investigation, C.-M.P., G.-M.K. and G.-S.C.; resources, G.-S.C.; data curation, G.-S.C.; writing—original draft preparation, C.-M.P.; writing—review and editing, G.-S.C.; visualization, C.-M.P.; supervision, G.-S.C.; project administration, G.-S.C.; funding acquisition, G.-S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (No. 2019R1C1C1011142).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The accession numbers in Table 1 are available in GenBank database (https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 5 April 2021)). The L. pentosus NGI01 strain is available in Korean Culture Center of Microorganisms (http://www.kccm.or.kr/ (accessed on 5 April 2021)).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pollard, D.J.; Woodley, J.M. Biocatalysis for pharmaceutical intermediates: The future is now. Trends Biotechnol. 2007, 25, 66–73. [Google Scholar] [CrossRef]
- Porter, J.L.; Rusli, R.A.; Ollis, D.L. Directed Evolution of Enzymes for Industrial Biocatalysis. ChemBioChem 2016, 17, 197–203. [Google Scholar] [CrossRef] [PubMed]
- El Gharras, H. Polyphenols: Food sources, properties, and applications—A review. Int. J. Food Sci. Technol. 2009, 44, 2512–2518. [Google Scholar] [CrossRef]
- Gupta, C.; Prakash, D. Phytonutrients as therapeutic agents. J. Complement. Integr. Med. 2014, 11, 151–169. [Google Scholar] [CrossRef]
- Tripoli, E.; Guardia, M.L.; Giammanco, S.; Majo, D.D.; Giammanco, M. Citrus flavonoids: Molecular structure, biological activity and nutritional properties: A review. Food Chem. 2007, 104, 466–479. [Google Scholar] [CrossRef]
- Day, A.J.; DuPont, M.S.; Ridley, S.; Rhodes, M.; Rhodes, M.J.C.; Morgan, M.R.A.; Williamson, G. Deglycosylation of flavonoid and iso-flavonoid glycosides by human small intestine and liver β-glucosidase activity. FEBS Lett. 1998, 436, 71–75. [Google Scholar] [CrossRef] [Green Version]
- Hollman, P.C.H. Absorption, Bioavailability, and Metabolism of Flavonoids. Pharm. Biol. 2004, 42, 74–83. [Google Scholar] [CrossRef]
- Thilakarathna, S.H.; Rupasinghe, H.P.V. Flavonoid Bioavailability and Attempts for Bioavailability Enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, G.B.; Smagghe, G.; Grootaert, C.; Zotti, M.; Raes, K.; Van Camp, J. Flavonoid interactions during digestion, absorption, distribution, and metabolism: A sequential structure-activity/property relationship-based approach in the study of bioa-vailability and bioactivity. Drug Metab. Rev. 2015, 47, 175–190. [Google Scholar] [CrossRef]
- Hollman, P.C.H.; Bijsman, M.N.C.P.; van Gameren, Y.; Cnossen, E.P.J.; de Vries, J.H.M.; Katan, M.B. The sugar moiety is a major deter-minant of the absorption of dietary flavonoid glycosides in man. Free Radic. Res. 1999, 31, 569–573. [Google Scholar] [CrossRef]
- Nielsen, I.L.; Chee, W.S.; Poulsen, L.; Offord-Cavin, E.; Rasmussen, S.E.; Frederiksen, H.; Enslen, M.; Barron, D.; Horcajada, M.N.; Wil-liamson, G. Bioavailability is improved by enzymatic modification of the citrus flavonoid hesperidin in humans: A ran-domized, double-blind, crossover trial. J. Nutr. 2006, 136, 404–408. [Google Scholar] [CrossRef] [Green Version]
- Amaretti, A.; Raimondi, S.; Leonardi, A.; Quartieri, A.; Rossi, M. Hydrolysis of the rutinose-conjugates flavonoids rutin and hes-peridin by the gut microbiota and bifidobacteria. Nutrients 2015, 7, 2788–2800. [Google Scholar] [CrossRef] [Green Version]
- Mueller, M.; Zartl, B.; Schleritzko, A.; Stenzl, M.; Viernstein, H.; Unger, F.M. Rhamnosidase activity of selected probiotics and their ability to hydrolyse flavonoid rhamnoglucosides. Bioprocess Biosyst. Eng. 2017, 41, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Beekwilder, J.; Marcozzi, D.; Vecchi, S.; De Vos, R.; Janssen, P.; Francke, C.; Vlieg, J.V.H.; Hall, R.D. Characterization of Rhamnosidases from Lactobacillus plantarum and Lactobacillus acidophilus. Appl. Environ. Microbiol. 2009, 75, 3447–3454. [Google Scholar] [CrossRef] [Green Version]
- Ketudat Cairns, J.R.; Esen, A. β-Glucosidases. Cell. Mol. Life Sci. 2010, 67, 3389–3405. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Yadav, P.K.; Yadav, S.; Yadav, K.D.S. α-L-Rhamnosidase: A review. Process Biochem. 2010, 45, 1226–1235. [Google Scholar] [CrossRef]
- Busto, M.; Meza, V.; Ortega, N.; Perez-Mateos, M. Immobilization of naringinase from Aspergillus niger CECT 2088 in poly(vinyl alcohol) cryogels for the debittering of juices. Food Chem. 2007, 104, 1177–1182. [Google Scholar] [CrossRef]
- Spagna, G.; Barbagallo, R.N.; Martino, A.; Pifferi, P.G. A simple method for purifying glycosidases: α-l-rhamnopyranosidase from Aspergillus niger to increase the aroma of Moscato wine. Enzym. Microb. Technol. 2000, 27, 522–530. [Google Scholar] [CrossRef]
- Németh, K.; Plumb, G.W.; Berrin, J.G.; Juge, N.; Jacob, R.; Naim, H.Y.; Williamson, G.; Swallow, D.M.; Kroon, P.A. Deglycosylation by small intestinal epithelial cell beta-glucosidases is a critical step in the absorption and metabolism of dietary flavonoid glycosides in humans. Eur. J. Nutr. 2003, 42, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. Open Access. 2018, 6, 89–94. [Google Scholar]
- Patra, J.K.K.; Das, G.; Paramithiotis, S.; Shin, H.-S. Kimchi and Other Widely Consumed Traditional Fermented Foods of Korea: A Review. Front. Microbiol. 2016, 7, 1493. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.J.; Lee, J.-E.; Lee, C.-H. Importance of lactic acid bacteria in Asian fermented foods. Microb. Cell Factories 2011, 10, S5. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.-B.; Kim, K.-S.; Rhee, J.-S. Hydrolysis of soybean isoflavone glucosides by lactic acid bacteria. Biotechnol. Lett. 2002, 24, 2113–2116. [Google Scholar] [CrossRef]
- Lee, N.-K.; Paik, H.-D. Bioconversion Using Lactic Acid Bacteria: Ginsenosides, GABA, and Phenolic Compounds. J. Microbiol. Biotechnol. 2017, 27, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Michlmayr, H.; Kneifel, W. β-Glucosidase activities of lactic acid bacteria: Mechanisms, impact on fermented food and human health. FEMS Microbiol. Lett. 2014, 352, 1–10. [Google Scholar] [CrossRef] [Green Version]
- McMahon, H.; Zoecklein, B.W.; Fugelsang, K.; Jasinski, Y. Quantification of glycosidase activities in selected yeasts and lactic acid bacteria. J. Ind. Microbiol. Biotechnol. 1999, 23, 198–203. [Google Scholar] [CrossRef]
- Bowers, G.N.; McComb, R.B.; Christensen, R.C.; Schaffer, R. High-Purity 4-Nitrophenol: Purification, Characterization, and Speci-fications for Use as a Spectrophotometric Reference Material. Clin. Chem. 1980, 26, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Liu, X.Q.; Du, X.P.; Wu, L.; Jiang, Z.D.; Ni, H.; Li, Q.B.; Chen, F. Preparation of isoquercitrin by biotransformation of rutin using α-L-rhamnosidase from Aspergillus niger JMU-TS528 and HSCCC purification. Prep. Biochem. Biotechnol. 2019, 50, 1–9. [Google Scholar] [CrossRef]
- Langston, J.; Sheehy, N.; Xu, F. Substrate specificity of Aspergillus oryzae family 3 β-glucosidase. Biochim. Biophys. Acta Proteins Proteom. 2006, 1764, 972–978. [Google Scholar] [CrossRef]
- Volkov, P.V.; Rozhkova, A.M.; Zorov, I.N.; Sinitsyn, A.P. Cloning, purification and study of recombinant GH3 family β-glucosidase from Penicillium verruculosum. Biochimie 2020, 168, 231–240. [Google Scholar] [CrossRef]
Figure 1.
Biotransformation pathway of hesperidin (A) and rutin (B) by α-l-rhamnosidase and β-d-glucosidase.
Figure 1.
Biotransformation pathway of hesperidin (A) and rutin (B) by α-l-rhamnosidase and β-d-glucosidase.
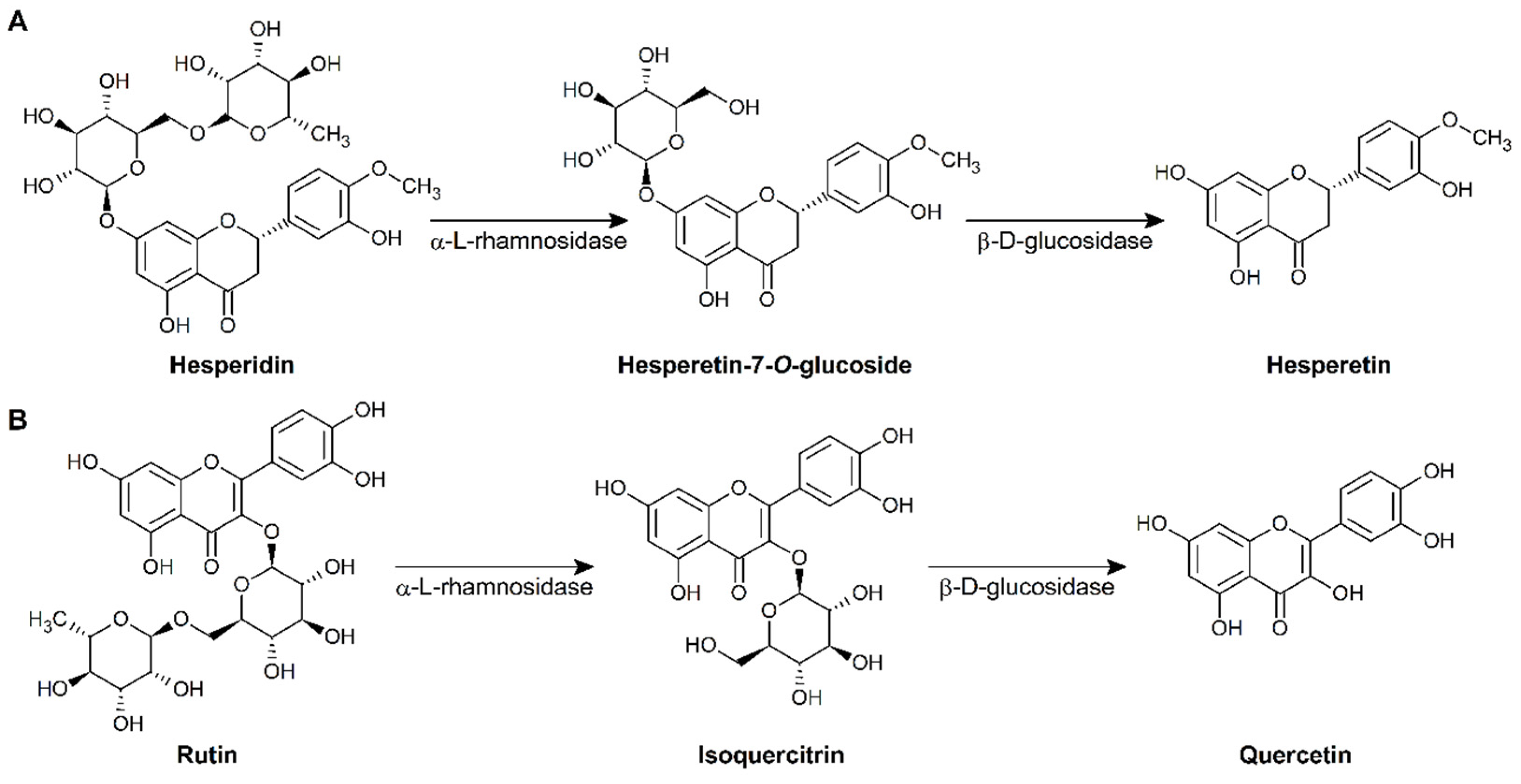
Figure 2.
α-l-rhamnosidase and β-d-glucosidase activities of 34 LAB strains isolated from kimchi.
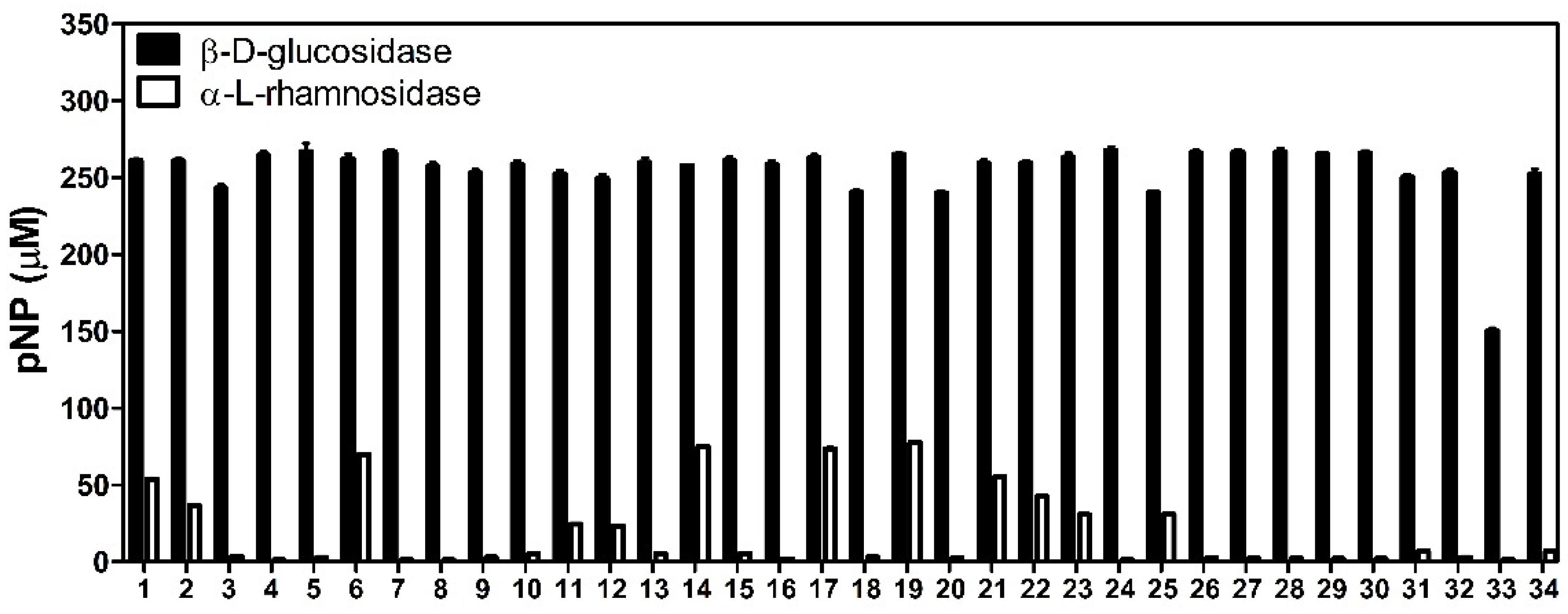
Figure 3.
Production of hesperetin (A) and quercetin (B) by L. pentosus NGI01.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Identification of LAB strains with high enzyme activities.
| Strain | Isolation Source | Species | Accession Number |
|---|---|---|---|
| NGI01 | Cucumber kimchi | L. pentosus | MT898558 |
| NGI02 | Cucumber kimchi | L. senmaizukei | MT898562 |
| NGI06 | Cabbage kimchi | L. pentosus | MT898563 |
| NGI14 | Cabbage kimchi | L. plantarum | MT898568 |
| NGI17 | Cabbage kimchi | L. pentosus | MT919355 |
| NGI19 | Cabbage kimchi | L. pentosus | MT898576 |
| NGI21 | Cabbage kimchi | L. paraplantarum | MT898577 |
| NGI22 | Cabbage kimchi | L. pentosus | MT898642 |
| NGI23 | Cabbage kimchi | L. pentosus | MT898646 |
| NGI25 | Cabbage kimchi | L. curvatus | MT898647 |
Table 2.
Biotransformation of hesperidin and rutin by 10 LAB strains isolated form kimchi.
| Strain | Product Yield (%) | |
|---|---|---|
| Hesperidin → Hesperetin | Rutin → Quercetin | |
| NGI01 | 30.3 ± 0.1 | 3.9 ± 0.2 |
| NGI02 | 26.3 ± 2.3 | 0 |
| NGI06 | 28.3 ± 0.8 | 0 |
| NGI14 | 20.4 ± 0.1 | 0 |
| NGI17 | 26.7 ± 0.1 | 0 |
| NGI19 | 24.9 ± 0.2 | 0 |
| NGI21 | 22.7 ± 0.1 | 0 |
| NGI22 | 23.1 ± 0.1 | 0 |
| NGI23 | 12.5 ± 0.1 | 0 |
| NGI25 | 15.5 ± 0.1 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, C.-M.; Kim, G.-M.; Cha, G.-S. Biotransformation of Flavonoids by Newly Isolated and Characterized Lactobacillus pentosus NGI01 Strain from Kimchi. Microorganisms 2021, 9, 1075. https://doi.org/10.3390/microorganisms9051075
AMA Style
Park C-M, Kim G-M, Cha G-S. Biotransformation of Flavonoids by Newly Isolated and Characterized Lactobacillus pentosus NGI01 Strain from Kimchi. Microorganisms. 2021; 9(5):1075. https://doi.org/10.3390/microorganisms9051075
Chicago/Turabian StylePark, Chan-Mi, Gyoung-Min Kim, and Gun-Su Cha. 2021. "Biotransformation of Flavonoids by Newly Isolated and Characterized Lactobacillus pentosus NGI01 Strain from Kimchi" Microorganisms 9, no. 5: 1075. https://doi.org/10.3390/microorganisms9051075
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.